Embed Size (px)
Citation preview

Original Article
Folia Primatol 2011;82:25–32 DOI: 10.1159/000326056
Geophagy in Brown Spider Monkeys
(Ateles hybridus) in a Lowland Tropical
Rainforest in Colombia
Andres Link a, b Ana Gabriela de Luna a, d Ricardo Arango a, c Maria Clara Diaz a, c a Fundación Proyecto Primates, Colombia, b Departamento de Ciencias Biológicas, Universidad de Los Andes, Bogotá , y c Departamento de Biología, Universidad de Caldas, Manizales , Colombia; d Facultad de Ciencias Biológicas, Universidad Complutense de Madrid, Madrid , España
Key Words
Ateles hybridus � Camera traps � Mineral lick � Soil consumption � Spider monkeys
Abstract
Spider monkeys and howler monkeys are the only Neotropical primates that eat soil from mineral licks. Not all species within these genera visit mineral licks, and ge-ophagy has been restricted to populations of Ateles belzebuth belzebuth, Ateles belzebuth chamek and Alouatta seniculus in western Amazonian rainforests. With the aid of a cam-era trap we studied the visitation patterns of a group of brown spider monkeys (Ateles hybridus) to a mineral lick at Serrania de Las Quinchas, in Colombia. Spider monkeys vis-ited the lick frequently throughout the year, with a monthly average of 21.7 8 7.2 visits per 100 days of camera trapping (n = 14 months). Spider monkeys visited the mineral lick almost always on days with no rain, or very little ( ! 3 mm) rain, suggesting that prox-imate environmental variables might determine spider monkeys’ decisions to come to the ground at the licks. This study expands the geographical occurrence of mineral lick use by spider monkeys providing additional data for future assessments on the biogeo-graphical correlates of mineral lick use by platyrrhines.
Copyright © 2011 S. Karger AG, Basel
Introduction
Geophagy, the intentional consumption of soil by animals, has been observed in several primate species amongst catarrhines and platyrrhines [Krishnamani and Mahaney, 2000; Ferrari et al., 2008]. For many primate species, or populations, soil is an important and frequently consumed item in their diets (e.g. Ateles and Alouat-
Received: October 27, 2009 Accepted after revision: February 11, 2011 Published online: April 15, 2011
Andres Link Department of Anthropology, New York University 25 Waverly Place New York, NY 10003 (USA) Tel. +1 212 998 3813, E-Mail al898 @ nyu.edu
© 2011 S. Karger AG, Basel0015–5713/11/0821–0025$38.00/0
Accessible online at:www.karger.com/fpr
Fax +41 61 306 12 34E-Mail [email protected]

Folia Primatol 2011;82:25–32 26 Link /de Luna /Arango /Diaz
ta ) [Izawa, 1993; Blake et al., 2010; Link et al., 2011] most often obtained from termite mounds, leaf-cutting ant mounds, the forest floor or mineral licks [Ferrari et al., 2008]. Mineral licks are specific sites where not only primates come to eat soil or drink water, but also a large diversity of other mammals and birds use them for sim-ilar purposes [Izawa, 1993; Montenegro, 2004; Tobler et al., 2009]. Even though ge-ophagy in platyrrhines seems to occur in several genera, only two species of spider monkeys ( Ateles spp.) and one species of howler monkeys (Alouatta seniculus) are regular visitors at mineral licks [Ferrari et al., 2008].
Several non-exclusive hypotheses have been proposed to explain the functional-ity of geophagy in mammals and birds [Johns and Duquette, 1991; Krishnamani and Mahaney, 2000; Brightsmith and Muñoz-Najar, 2004; Brightsmith et al., 2008]. The most accepted hypotheses propose that soil (1) can help detoxify secondary plant metabolites and tannins [Gilardi et al., 1999; Johns and Duquette, 1991], (2) can pro-vide supplementary minerals that might be absent or in very low concentration in an animal’s diet [Oates, 1978; Brightsmith et al., 2008; Lee et al., 2009], (3) can act as an anti-diarrhoeal drug [Krishnamani and Mahaney, 2000] or (4) can be associated with the enhancement of the pharmacological properties of secondary plant com-pounds [Klein et al., 2008]. The higher concentration of minerals such as calcium, sodium or potassium, amongst others, from the mineral lick soils when compared to control sites [Emmons and Starck, 1979; Izawa, 1993; Montenegro, 2004; Mahaney et al., 2005], combined with the frequent use of mineral licks by many bird and mam-mal species, suggests that it has beneficial effects on their nutrition and health.
Only scant data exist on geophagy by atelids [Izawa, 1975; Ayres and Ayres, 1979; Izawa, 1993; Blake et al., 2010]. Recently, Ferrari et al. [2008] compiled the data on geophagy by New World monkeys and proposed two potential trends: (1) geoph-agy is more common amongst herbivorous species compared to those that include insects (at least partially) in their diets, and (2) geophagy is more common for Ama-zonian primates compared to other Neotropical primates. From these meta-analyses (see table 3 in Ferrari et al. [2008]) and the review on spider monkey diet done by Di Fiore et al. [2008], it is noticeable that all reports of geophagy at mineral licks by Neo-tropical primates are restricted to long-haired spider monkeys (Ateles belzebuth bel-zebuth), black spider monkeys (Ateles belzebuth chamek) and red howler monkeys (Alouatta seniculus), particularly in populations living in western Amazonia. Data on the temporal patterns of mineral lick visitation by primates or on the influence of environmental factors on primate geophagy are still scant.
Spider monkeys are one of the largest Neotropical primates. They are ripe-fruit specialists generally relying on a wide variety of fruits usually complement their diet with young leaves and other plant items [Di Fiore et al., 2008], insectivory occurs in only a few populations and accounts for a very small portion ( 1 1%) of their diet [Link, 2004]. They have fission-fusion grouping patterns in which subgroups fre-quently change their size and composition [Aureli and Schaffner, 2008].
This study aims to provide a descriptive analysis of the temporal patterns of min-eral lick use by a group of wild brown spider monkeys in Colombia. We specifically test if spider monkey visitation patterns at Las Quinchas respond to immediate weath-er conditions as described for parrots in Amazonia [Brightsmith, 2004], and discuss our results based on the hypothesis that primates perceive high predation risk when accessing mineral licks on the forest floor. This study also expands the geographical range of places where primate visits to mineral licks are frequent in the Neotropics.

Geophagy in Brown Spider Monkeys 27Folia Primatol 2011;82:25–32
Methods
This research took place at the Private Reserve ‘El Paujil’ (06°02 � 48 � � N, 74°16 � 00 � � W) from Fundación ProAves, in the Serrania de Las Quinchas, a small mountain chain in the west-ern slopes of the Eastern Andes in Colombia. Serrania de Las Quinchas is one of the largest Inter-Andean lowland forests on the eastern bank of the Magdalena River. Annual rainfall av-erages 3,544 8 336 mm (2006–2008) and temperature is relatively constant throughout the year with minimum and maximum daily average temperatures of 24.5 and 32.0 ° C, respectively. The study site contains a mixture of rainforest and recently formed pastures. It comprises a forest fragment of ca. 80 ha that connected to a larger forested area during the study through several ‘canopy bridges’. Our main study group of brown spider monkeys has been habituated and stud-ied since January 2006. This group visits a single mineral lick which consists of a cliff wall about 30 m high, located next to a dirt road that was built over 10 years ago. The primates access the lick by descending the trees at the top of the cliff.
During 13 months (December 14, 2007 through January 20, 2009), we monitored the min-eral lick using one camera trap (CuddeBack Excite � ) aimed at the base of the only tree where primates could access the mineral lick. The camera was set to take a picture every minute(the shortest available time interval between pictures) upon detecting movement. This allowed us to record mineral lick visits even if we were not following the group. We collected daily data on rainfall at 6: 00 and 18: 00 using a rain gauge and the daily temperature through a wall ther-mometer.
We assessed the frequency of spider monkey visits to the mineral lick by counting all in-dependent visits for those months in which the camera was working continuously, and esti-mated the frequency of visitation on those months when the camera was not working for 1 1 day. Expected visitation rates were calculated by multiplying the number of visits in each month by the quotient between the days the trap was working in a month and the total number of days in that particular month. Sampling effort is described in table 1 . We defined independent visits as visits that occurred in periods greater than 3 h, based on preliminary data on the time of vigi-lance displayed by spider monkey groups around mineral licks prior to coming down to the ground at several sites in Colombia and Ecuador [Link and Di Fiore, 2009].
Results
During 379 days of salt lick monitoring, our main study group of brown spider monkeys visited the mineral lick on 81 occasions. On 5 days, spider monkeys came twice to the mineral lick, thus visiting the lick on a total of 76 days. These data include only the times when primates came down to the ground and ate soil, as spider mon-keys sometimes came to the mineral lick area but left prior to eating soil [Link, un-publ. data]. Thus the rates presented in this article refer to the successful visitation of mineral licks where at least one spider monkey consumed soil from the ground.
Table 1. N umber of days per month with camera trap monitoring of the mineral lick used by brown spider monkeys
Dec07
Jan08
Feb08
Mar08
Apr08
May08
Jun08
Jul08
Aug08
Sep08
Oct08
Nov08
Dec08
Jan09
Days sampled 15 30 28 29 30 31 20 30 31 26 30 30 30 19Spider monkey visits 4 7 4 2 7 8 5 5 3 6 7 9 9 5
Folia Primatol 2011;82:25–32 28 Link /de Luna /Arango /Diaz
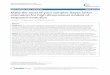
Based on approximately 800 h of behavioural follows collected continuously from January 2006 to January 2009 on our study group of brown spider monkeys, we observed that they used only one mineral lick to eat soil from the ground. Min-eral lick visitation by brown spider monkeys was frequent throughout the year, aver-aging 21.4 visits per 100 days that corresponds to on average slightly more than 1 visit every 5 days. The expected visitation rates were almost identical (as sampling was almost complete for all months) averaging 21.7 8 7.2 visits per 100 days (range: 6.9–30.0 visits per 100 days, n = 14 months) as shown in figure 1 .
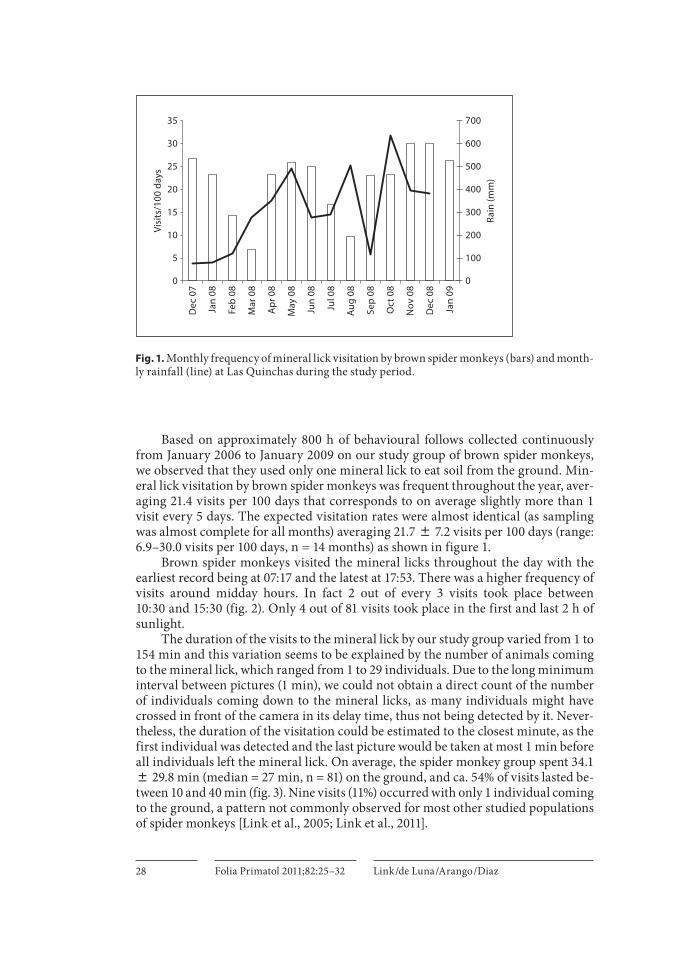
Brown spider monkeys visited the mineral licks throughout the day with the earliest record being at 07: 17 and the latest at 17: 53. There was a higher frequency of visits around midday hours. In fact 2 out of every 3 visits took place between10: 30 and 15: 30 ( fig. 2 ). Only 4 out of 81 visits took place in the first and last 2 h of sunlight.
The duration of the visits to the mineral lick by our study group varied from 1 to 154 min and this variation seems to be explained by the number of animals coming to the mineral lick, which ranged from 1 to 29 individuals. Due to the long minimum interval between pictures (1 min), we could not obtain a direct count of the number of individuals coming down to the mineral licks, as many individuals might have crossed in front of the camera in its delay time, thus not being detected by it. Never-theless, the duration of the visitation could be estimated to the closest minute, as the first individual was detected and the last picture would be taken at most 1 min before all individuals left the mineral lick. On average, the spider monkey group spent 34.1 8 29.8 min (median = 27 min, n = 81) on the ground, and ca. 54% of visits lasted be-tween 10 and 40 min ( fig. 3 ). Nine visits (11%) occurred with only 1 individual coming to the ground, a pattern not commonly observed for most other studied populations of spider monkeys [Link et al., 2005; Link et al., 2011].
Dec
07
Jan
08
Feb
08
Mar
08
Apr
08
May
08
Jun
08
Jul 0
8
Aug
08
Sep
08
Oct
08
Nov
08
Dec
08
Jan
09
0
5
10
15
20
25
30
35
0
100
200
300
400
500
600
700
Rain
(mm
)
Visi
ts/1
00 d
ays
Fig. 1. Monthly frequency of mineral lick visitation by brown spider monkeys (bars) and month-ly rainfall (line) at Las Quinchas during the study period.
Geophagy in Brown Spider Monkeys 29Folia Primatol 2011;82:25–32
Spider monkeys visited the lick during dry days ( ! 3 mm) more often than ex-pected by chance (Yates’ � 2 = 3.926, d.f. = 1, p = 0.047, n = 363 days). There was a tendency for spider monkeys to visit the mineral licks on days with no rain as 98% (72 out of 74) of visits took place on days with ! 3 mm of rain, and 80% of the days had no rain at all. Only 2 visits (2.7%) took place on days when there was substantial rain, even though we could not determine if this rain occurred after mineral lick visitations as rain gauge records cover 12-hour periods (6: 00 to 18: 00).
0
2
4
6
8
10
12
14
16
6:00 7:00 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Visi
ts
Fig. 2. Daily pattern of mineral lick visitation by brown spider monkeys.
0
5
10
15
20
25
Visi
ts
1
2–10
11–2
0
21–3
0
31–4
0
41–5
0
51–6
0
61–7
0
71–8
0
81–9
0
91–1
00
101–
110
>11
0
Duration (min)
Fig. 3. Duration of mineral lick visitation by brown spider monkeys.

Folia Primatol 2011;82:25–32 30 Link /de Luna /Arango /Diaz
Discussion
The results of this study suggest that mineral licks are important sites for some populations of brown spider monkeys (Ateles hybridus) as they visit them frequent-ly throughout the year. It is still uncertain if geophagy is providing brown spider monkeys with supplementary minerals, and/or aiding detoxification. Nevertheless, it is evident that soil is a regular resource in the brown spider monkeys’ diet, which at the study site is non-seasonal having only minor variation, with fruits comprising on average 93.4 8 6.7% (n = 27 months) of their monthly diets [Link et al., unpubl. data]. The importance of mineral licks for spider monkeys and howler monkeys has also been suggested in previous studies at Tinigua Park, Colombia [Izawa, 1993;Inaba, 2000; Shimooka, 2005], and ongoing studies at Yasuni Park, Ecuador [Blake et al., 2010; Link et al., 2011], where both species frequently visited mineral licks throughout the year [Izawa, 1993; Inaba, 2000]. Shimooka [2005] also suggested that mineral licks influenced the ranging behaviour of spider monkeys, and Spehar et al. [2010] and Link et al. [2011] found that the area around mineral licks was the most frequent and intensively used area for groups of spider monkeys in eastern Ecuador.
Consistent with the observations of environmental correlates of mineral lick use by parrots at Tambopata, Peru [Brightsmith, 2004], and by howler monkeys and spi-der monkeys at Tiputini Biological Station in Ecuador [Link et al., 2011], brown spi-der monkeys also visited the mineral licks on days without rain or adverse weather conditions. Brightsmith [2004] observed that birds would visit mineral licks on 94% of mornings that were sunny. When weather was ‘inclement’, there was a significant reduction in the percentage of mornings with birds visiting suggesting that rain re-duced bird visits to mineral licks.
For arboreal primates, mineral licks are areas with a relatively higher risk of predation from terrestrial predators. Izawa [1993] suggested that the selection of mineral licks by spider monkeys and howler monkeys could be influenced by their sense of predation risk. Although infrequent, indirect and direct evidence of preda-tion of howler monkeys and spider monkeys by pumas (Puma concolor) and jaguars (Panthera onca) exists [Ferrari, 2009]. In fact, Matsuda and Izawa [2008] observed predation of a spider monkey on the forest floor by a jaguar and another attempt of predation of a spider monkey by a puma at one of the mineral licks used by them. This evidence suggests that at least for large arboreal primates, mineral lick visitation is influenced by environmental and climatological conditions, as they might in turn influence the probability of detecting a predator on the forest ground [Link et al., 2011].
If predation is a major factor influencing the decisions of spider monkeys to come down to the ground to eat soil, it could be hypothesized that, in general, spider monkeys would come down to the salt lick in, on average, larger groups than the av-erage size of foraging or travelling subgroups. This seems to be the case for popula-tions of spider monkeys living in pristine ecosystems that have high densities of large felids [Link and Di Fiore, 2009]. From the results obtained in our study, it is impor-tant to take into account that the high percentage of visits when small subgroups of spider monkeys came to the ground could reflect the fact that the mineral lick used by our study group is located adjacent to a road and thus exposed to humans, a more influential predator of brown spider monkeys in this area in the recent past.

Geophagy in Brown Spider Monkeys 31Folia Primatol 2011;82:25–32
This study is the first report of mineral lick use by spider monkeys outside west-ern Amazonian populations. The reasons why only these populations of spider mon-keys and howler monkeys visit mineral licks remain unknown but seem consistent with the geographical patterns observed for other taxa (e.g. parrots in Lee et al. [2009]). This study thus broadens the geographical area where mineral licks are used by primates in the Neotropics allowing for stronger tests on the ecological and bio-geographical correlates of mineral lick availability and use by primates in the Neo-tropics. These data might contribute to an understanding of the specificity of min-eral lick use and help solve the mystery as to why licks are used by some populations of spider monkeys, while other populations never use them [Campbell et al., 2005].
Acknowledgments
We thank all our students and assistants who collaborated with us in the field. Erin Flem-ing, Lindsey Smith and two anonymous reviewers improved earlier versions of the manuscript. Fundación ProAves provided us with logistic support and collaboration to conduct our long-term study on brown spider monkeys and the primate community at their private reserve ‘El Paujil’. We specially thank Eliana Machado, and Euripides Montero for their kind support in the field. Fundación Biodiversa Colombia provided us with institutional support during this project, and we specially thank Alba Lucia Morales. This project was made possible thanks to the grants provided by Idea Wild, Primate Conservation Inc., Primate Action Fund (Conserva-tion International) and Rufford Small Grants Foundation for the conservation of Ateles hybri-dus in Colombia.
References
Aureli F, Schaffner CM (2008). Social interactions, social relationships and the social system of spider monkeys. In Spider Monkeys: Behavior, Ecology and Evolution of the Genus Ateles (Campbell CJ, ed.), pp 236–265. New York, Cambridge University Press.
Ayres JM, Ayres CM (1979). Aspectos da caça no alto Rio Aripuanã. Acta Amazonica 9: 287–298. Blake JG, Guerra J, Mosquera D, Torres R, Loiselle BA, Romo D (2010). Use of mineral licks by white-
bellied spider monkeys (Ateles belzebuth) and red howler monkeys (Alouatta seniculus) in eastern Ecuador. International Journal of Primatology 31: 471–483.
Brightsmith DJ (2004). Effects of weather on parrot geophagy at Tambopata, Peru. The Wilson Bulletin 116: 134–145.
Brightsmith DJ, Muñoz-Najar RA (2004). Avian geophagy and soil characteristics in southeastern Peru. Biotropica 36: 534–543.
Brightsmith DJ, Taylor J, Phillips OL (2008). The roles of soil characteristics and toxin adsorption in avian geophagy. Biotropica 40: 766–774.
Campbell CJ, Aureli F, Chapman CA, Ramos-Fernandez G, Matthews K, Russo SE, Suarez S, Vick L (2005). Terrestrial behavior of Ateles sp. International Journal of Primatology 26: 1039–1051.
Di Fiore A, Link A, Campbell CJ (2011). The atelines: Behavioral and socioecological diversity in a New World radiation. In Primates in Perspective (Campbell CJ, Fuentes A, MacKinnon KC, Panger M, Bearder SK, eds.), 2nd ed. New York, Oxford University Press.
Di Fiore A, Link A, Dew JL (2008). Diets of wild spider monkeys. In Spider Monkeys: Behavior, Ecology and Evolution of the Genus Ateles (Campbell CJ, ed.), pp 81–137. Cambridge, Cambridge Univer-sity Press.
Emmons LH, Starck NM (1979). Element composition of a natural mineral lick in Amazonia. Biotro-pica 11: 311–313.
Ferrari FF (2009). Predation risk and antipredator strategies. In South American Primates: Comparative Perspectives in the Study of Behavior, Ecology, and Conservation (Garber PA, Estrada A, Bicca-Marques JC, Heymann EW, Strier KB, eds.), pp 211–250. New York, Springer.
Ferrari FF, Veiga LM, Urbani B (2008). Geophagy in New World monkeys (Platyrrhini): ecological and geographic patterns. Folia Primato logica 79: 402–415.

Folia Primatol 2011;82:25–32 32 Link /de Luna /Arango /Diaz
Gilardi JD, Duffey SS, Jun CA, Tell LA (1999). Biochemical functions of geophagy in parrots: detoxifi-cation of dietary toxins and cytoprotective effects. Journal of Chemical Ecology 25: 897–922.
Inaba A (2000). Kumozaru no gun-kan kankei. (Intergroup relationships of wild spider monkeys in Macarena, Colombia.). In 16th Annual Meeting of Primate Society of Japan . Nagoya, Dogura and Co, Ltd, p 257.
Izawa K (1975). Food and feeding behavior of monkeys in the upper Amazon basin. Primates 16: 295–316.
Izawa K (1993). Soil-eating by Alouatta and Ateles . International Journal of Primatology 14: 229–242. Johns T, Duquette M (1991). Detoxification and mineral supplementation as functions of geophagy.
American Journal of Clinical Nutrition 53: 448–456. Klein N, Fröhlich F, Krief S (2008). Geophagy: soil consumption enhances the bioactivities of plants
eaten by chimpanzees. Naturwissenschaften 95: 325–331. Krishnamani R, Mahaney WC (2000). Geophagy among primates: adaptive significance and ecological
consequences. Animal Behavior 59: 899–915. Lee ATK, Kumar S, Brightsmith DJ, Marsden SJ (2009). Parrot claylick distribution in South America:
do patterns of ‘where’ help answer the question of ‘why’? Ecography 32: 1–11. Link A (2004). Insect eating by spider monkeys. Neotropical Primates 11: 104–107. Link A, Di Fiore A (2009). Effects of predation risk on the grouping patterns of spider monkeys. Folia
Primatologica 80: 125. Link A, Di Fiore A, Spehar S (2005). Predation risk affects subgroup size in spider monkeys (Ateles bel-
zebuth) at Yasuni National Park, Ecuador. Folia Primatologica 77: 318. Link A, Galvis N, Fleming E, Di Fiore A (2011). Pattern of mineral lick visitation by spider monkeys and
howler monkeys in Amazonia: are licks perceived as risky areas? American Journal of Primatol-ogy DOI: 10.1002/ajp.20910.
Mahaney WC, Milner MW, Aufreiter S, Hancock RGV, Wrangham R, Campbell S (2005). Soils con-sumed by chimpanzees of the Kenyawara community in the Kibale forest, Uganda. International Journal of Primatology 26: 1375–1398.
Matsuda I, Izawa K (2008). Predation of wild spider monkeys at La Macarena, Colombia. Primates 49: 65–68.
Montenegro OL (2004). Natural Licks as Keystone Resources for Wildlife and People in Amazonia . PhD thesis, University of Florida, Gainesville.
Oates JF (1978). Water-plant and soil consumption by guereza monkeys (Colobus guereza) : a relation-ship with minerals and toxins in the diet? Biotropica 10: 241–253.
Shimooka Y (2005). Sexual differences in the ranging behavior of Ateles belzebuth belzebuth at la Ma-carena, Colombia. International Journal of Primatology 26: 385–406.
Spehar S, Link A, Di Fiore A (2010). Male and female range use in a group of white-bellied spider mon-keys (Ateles belzebuth) in Yasuní National Park, Ecuador. American Journal of Primatology 72: 129–141.
Tobler MW, Carrillo-Percastegui SE, Powell G (2009). Habitat use, activity patterns and use of mineral licks by five species of ungulate in south-eastern Peru. Journal of Tropical Ecology 25: 261–270.
![Chemical Information Review Document for Butterbur ... · Chemical Information Review Document for Butterbur (Petasites hybridus, ext.) [CAS No. 90082-63-6] Supporting Nomination](https://img.pdfslide.us/doc/110x75/5e53e2025a104807131d0470/chemical-information-review-document-for-butterbur-chemical-information-review.jpg)