Embed Size (px)
Citation preview

Marine sponges (phylum Porifera) are functionally important members of marine benthic communities1. These sessile, filter-feeding animals can occupy up to 80% of available surfaces in polar regions2 and are often a dominant faunal component of tropical and temper-ate reefs3–5. However, the importance of sponges extends beyond the sea floor and into the water column itself, where their impressive pumping capacity (thousands of litres per kilogram of sponge per day) enables sponges to sequester food particles and other nutrients from the pelagic habitat1.
Having arisen some 600 million years ago, marine sponges are among the most ancient of all extant animal lineages6,7. However, bacteria were already well established in the oceans at this time, meaning that sponges evolved in a literal sea of potential pathogens and parasites. Their simple body plan has served sponges well throughout their evolution and has remained essentially unchanged (FIG. 1), but their lack of morphological complexity belies a remark-able and multifaceted arsenal of defences against invad-ing microorganisms. What is arguably most interesting about the humble sea sponges is not their ability to ward off harmful microorganisms, but rather their ability to house enormously dense and diverse communities of symbiotic bacteria, archaea and unicellular eukaryotes in their tissues8. These microorganisms can constitute up to 35% of the total sponge biomass and are found at den-sities exceeding 109 microbial cells per cubic centimetre
of sponge tissue (3–4 orders of magnitude greater than the density in the surrounding sea water)8. Most sponge-associated microorganisms inhabit the mesohyl tissue, an extracellular matrix that is populated by sponge cells and constitutes most of the sponge body (FIG. 1a,b). However, some symbionts may also exist intracellularly, as evidenced by early electron microscopy studies9. The mesohyl pres-ents a conundrum: it is both the site of digestion of micro-organisms (which can be food particles) and, in many sponges, the home for extensive communities of symbiotic microorganisms (FIG. 1b). How do these symbionts avoid being digested? Feeding studies have revealed that putative symbionts which are ingested by their hosts pass through the host unscathed, whereas ingested non- symbiont bacteria are generally consumed10,11. This implies that specific symbionts are either recognized and ignored by the host sponge or not recognized at all, per-haps because the microorganisms shield themselves from detection, for example by producing slime capsules10. Sponges also have a surprisingly well-developed innate immune system12–15 and produce a wide range of anti-microbial compounds16, a factor that adds to their com-plexity as a habitat for microorganisms. Conversely, microbial symbionts of sponges enjoy some benefits, including a stable nutrient supply as a result of the filter-feeding activities of the animal, as well as a supply of the scarce element nitrogen, which is excreted by the sponge in the form of ammonia as a metabolic end product.
1Julius-von-Sachs-Institute for Biological Sciences, University of Würzburg, Julius-von-Sachs Platz 3, 97082 Würzburg, Germany.2Kekulé Institute of Organic Chemistry and Biochemistry, University of Bonn, Gerhard-Domagk Straße 1, 53121 Bonn, Germany.3School of Biological Sciences, University of Queensland, Brisbane, Queensland 4072, Australia.4Centre for Microbial Innovation, School of Biological Sciences, University of Auckland, Private Bag 92019, Auckland, New Zealand.Correspondence to U.H. e-mail: [email protected]:10.1038/nrmicro2839Published online 30 July 2012
BenthicPertaining to organisms: living in or on the sea floor.
Genomic insights into the marine sponge microbiomeUte Hentschel1, Jörn Piel2, Sandie M. Degnan3 and Michael W. Taylor4
Abstract | Marine sponges (phylum Porifera) often contain dense and diverse microbial communities, which can constitute up to 35% of the sponge biomass. The genome of one sponge, Amphimedon queenslandica, was recently sequenced, and this has provided new insights into the origins of animal evolution. Complementary efforts to sequence the genomes of uncultivated sponge symbionts have yielded the first glimpse of how these intimate partnerships are formed. The remarkable microbial and chemical diversity of the sponge–microorganism association, coupled with its postulated antiquity, makes sponges important model systems for the study of metazoan host–microorganism interactions, and their evolution, as well as for enabling access to biotechnologically important symbiont-derived natural products. In this Review, we discuss our current understanding of the interactions between marine sponges and their microbial symbiotic consortia, and highlight recent insights into these relationships from genomic studies.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 1
Nature Reviews Microbiology | AOP, published online 30 July 2012; doi:10.1038/nrmicro2839
© 2012 Macmillan Publishers Limited. All rights reserved
Nature Reviews | Microbiology
Sea water
Sea floor
Choanoderm
Choanocyte
PinacocyteArchaeocyte
PinacodermMesohyl
MicroorganismSiliceous spicule
Osculum
MesohylChoanocyte chamber
Ostium
Cyanobacteriaa b
c
f
i
g
j
h
k
d e
Slime capsulesExtracellular masking layers that encapsulate bacterial cells, putatively shielding the cells from detection by a host (in the context of this Review, the sponge).
Biologically activeExhibiting activity in a biological assay (for example, a screen for antitumour agents).
Collectively, marine sponges and their micro-biomes boast an impressive metabolic and chemical repertoire that not only contributes to their nutri-tional ecology but also has elicited the interest of the pharmaceutical industry owing to their production of biologically active compounds16,17. This Review focuses on the genomic interplay between sponges and their microbial symbionts and considers the respective parts played by all partners in maintaining these complex associations. We begin by describing how the diverse
microbial communities associated with sponges are established and maintained. We then examine genom-ics-based findings about symbiont function and host innate immunity, before we finally outline the syn-thesis of biologically active natural products by the sponge–microorganism consortium. Other important research, such as the cultivation of sponge-associated bacteria18, is beyond the scope of this Review, and we refer interested readers to a previously published article covering these topics8.
R E V I E W S
2 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

AposymbioticPertaining to an organism: lacking its symbionts.
16S rRNA gene libraryA vector library created by the amplification and cloning of the 16S rRNA genes in a population. The library can be used for subsequent sequencing and phylogenetic analysis, as a method of describing the composition of a microbial community.
The sponge microbiome: a community affairEstablishment and maintenance of sponge–microorgan-ism associations. In many stable, long-term symbioses, microbial partners are passed between generations by maternal or vertical transmission, with no aposymbiotic phase19. The pea aphid, Acyrthosiphon pisum, and its obligate bacterial symbiont, Buchnera aphidicola20,21, exemplify this strategy, and there is strong evidence that co-evolution between these ancient partners has led to the markedly reduced bacterial genome in the symbiont. By contrast, juveniles of the bobtail squid Euprymna scolopes do not contain symbionts at hatch-ing but efficiently recruit the bioluminescent bacterium Vibrio fischeri from the surrounding sea water22. Our current understanding for marine sponges is that a com-bination of these two processes may apply8,23,24, analo-gous to the ‘leaky’ vertical transmission proposed for deep-sea mussels25. In the case of mussels, it seems that
bacterial symbionts are sometimes lost from the host but reacquired by horizontal transmission from the sea water. Transmission of a diverse microbial assemblage in sponge eggs and larvae has been shown for numerous sponge species using both molecular and microscopy techniques26–29. The fact that the adults and reproduc-tive stages of sponges have largely overlapping microbial community compositions suggests that many or most of the associated microorganisms are vertically trans-mitted27,30, although little is known about whether these communities actually persist in settled juveniles. Vertical transmission therefore provides a mechanism by which sponge–microorganism associations can be maintained over evolutionarily long periods of time (see BOX 1 and REFS 8,31,32), although an apparent lack of co-speciation between host and symbiont8 — coupled with tentative observations of “sponge bacteria” in sea water30 — suggest that sponges also recruit potential symbionts from the sea water.
Community composition and diversity. Although the antiquity of certain associations between extant sponges and microorganisms remains the subject of debate8,33,34, it is clear that sponges and microorganisms in general have been interacting since ancient times. The marine sponges of the present contain a striking phylogenetic diversity of microorganisms (FIG. 2). To date, 28 bacterial phyla (of which 18 are formally described and 10 are candidate phyla known only from their 16S rRNA sequences) have been recorded to be associated with sponges, on the basis of cultivation and/or conventional molecular approaches such as 16S rRNA gene library construction32. Several more phyla have been detected using deep pyrosequencing of 16S rRNA genes35,36, suggesting that these taxa are confined to the rare biosphere. The dominant sponge-associated bacterial phyla, regardless of the detection technique used, include Proteobacteria (especially the classes Alpha-, Gamma- and Deltaproteobacteria), Chloroflexi, Actinobacteria, Acidobacteria, Nitrospirae
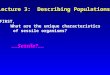
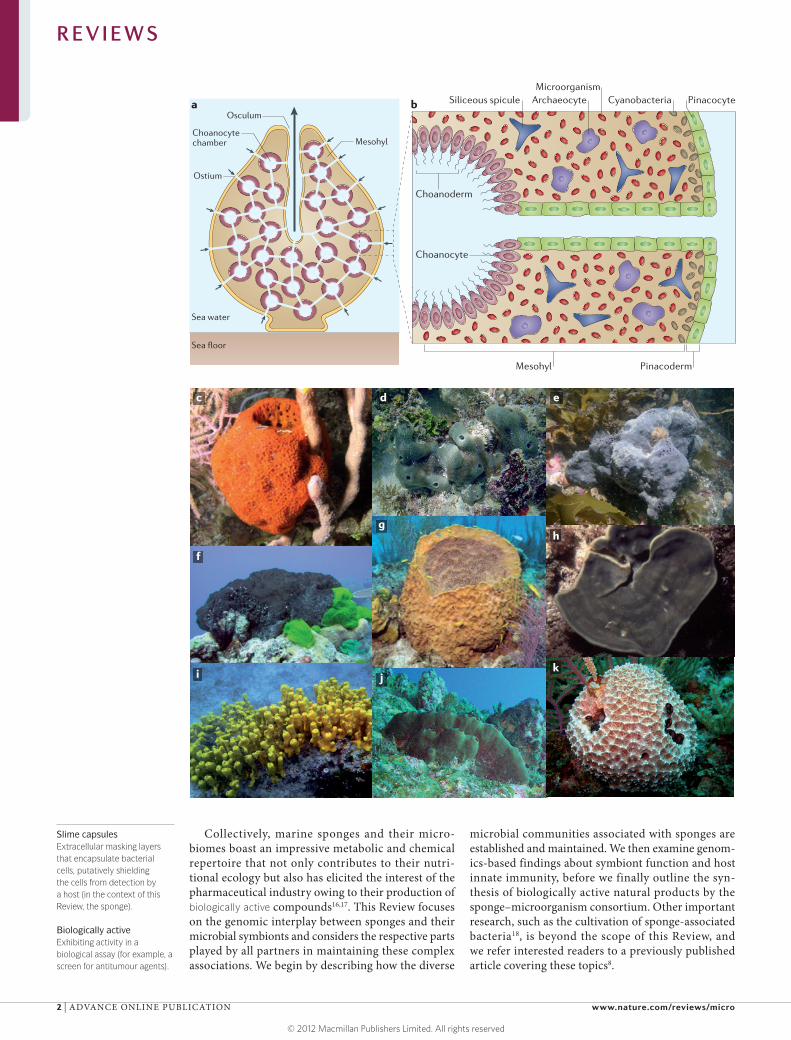
Figure 1 | Body plan and underwater images of marine sponges. a | A schematic overview of a typical demosponge. b | An enlargement of the internal structure of a typical demosponge. Sea water is conveyed into the sponge through small inhalant openings (ostia) in the outer pinacoderm tissue owing to a surge created by the beating of flagellated choanocyte cells lining the sponge’s aquiferous system. Near-sterile water (up to 96% of microbial cells are removed11,149) is then discharged via the exhalant opening (osculum). The choanocytes, which are specialized cells that are organized in chambers, serve to capture food particles in the sea water, including bacteria, unicellular algae and even viruses149–152. From the choanocyte chambers, microorganisms are transferred to the inner mesohyl layer, where they are engulfed by archaeocyte cells. Dense communities of symbiotic microorganisms exist in the mesohyl of many demosponges, with cyanobacteria being largely confined to the light-exposed outer regions. Siliceous spicules provide structural support to most demosponges. c–k | Underwater photography of important model sponge species: Mycale laxissima (c), Amphimedon queenslandica (d), Ancorina alata (e), Rhopaloeides odorabile (f), Xestospongia muta (g), Cymbastela concentrica (h), Aplysina aerophoba (i), Theonella swinhoei (j) and Ircinia felix (k). Images courtesy of J. Pawlik, University of North Carolina, Wilmington, USA (c,g); M. Taylor, University of Auckland, New Zealand (h); J. Tanaka, University of the Ryukus, Japan (j); and P. Erwin, University of Barcelona, Spain (k). Part d is reproduced, with permission, from REF. 153 © (2010) Bentham Science. Parts e,f,i are reproduced, with permission, from REF. 24 © (2011) John Wiley & Sons.
◀
Box 1 | Sponge-specific clusters and the evolutionary and ecological mechanisms that shape them
The term sponge-specific cluster refers to an important concept in sponge microbiology, but the evolutionary and ecological mechanisms that shape these clusters are still not fully understood. A sponge-specific, monophyletic 16S rRNA gene cluster is defined as a group of at least three 16S rRNA gene sequences that have been recovered from different sponge species and/or from different geographical locations and are more closely related to each other than to any other sequence from non-sponge sources31. Sponges that come from entirely different oceans and are distantly related to each other have microbiotas that share particular clades of microorganisms which seem to be absent (or at least rare) in other environments. These microorganisms are those that are most likely to represent true symbionts of sponges, rather than food bacteria that were present at the time of sampling. A recent study32 showed that nearly one-third of 7,500 sponge-derived 16S rRNA gene sequences fell into such sponge-specific clusters, although their presence varied greatly among phyla (FIG. 2). The identification of shared clusters between sponges and corals (for example, clusters in the phyla Nitrospirae and Gemmatimonadetes) is noteworthy, but their ecological significance remains to be investigated32. The mechanism (or mechanisms) that lead to the formation of sponge-specific clusters is also a topic of debate8. Microbial adaptation to life in the sponge mesohyl (specifically, resistance to phagocytosis by sponge archaeocytes) is likely to be an important selection factor. Moreover, a combination of vertical and horizontal microbial transmission is likely to occur, leading to highly specific associations on the one hand and ensuring global distributions on the other. The concept of sponge-specific clusters is clearly different from many other intimate and evolutionarily long-standing host–symbiont interactions that are characterized by co-evolution and congruent phylogenies. Rather than ‘one host, one symbiont’ associations, sponges represent ‘many hosts, many symbionts’ associations, probably as a result of the ample exchange with sea water that occurs during filter feeding, and also the lack of microbial compartmentation to specific cells or organs.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 3
© 2012 Macmillan Publishers Limited. All rights reserved
Nature Reviews | Microbiology
proteobacteria
OP3Verrucomicrobia
LentisphaeraeChlamydiae
PlanctomycetesCandidate phylum Poribacteria
SAULBRC1
Actinobacteria
GemmatimonadetesNitrospirae
BacteroidetesCaldithrix–Deferribacteres
Gamma-
Beta-Alpha-
Delta-Epsilon-TM6
AcidobacteriaCyanobacteria
FirmicutesFusobacteria
SpirochaetesChloroflexi
TM7Armatimonadetes
OP11OD1
OP1Thaumarchaeota
Euryarchaeota
Deinococcus–ThermusBD1-5
FungiOther eukaryotes
0.10
Bacteria
Archaea
Eukarya
% of sequences in sponge-specific clusters
01–2526–5051–7576–100
PyrosequencingA next-generation sequencing method based on the detection of pyrophosphate release on nucleotide incorporation. The technique allows thousands or millions of sequences to be obtained from a given sample.
Rare biosphereThe low-abundance microorganisms that are found in a microbial community.
Operational taxonomic units(OTUs). An arbitrary definition for a specific taxonomic grouping; for example, 97% 16S rRNA sequence similarity is frequently used to approximate a bacterial species.
PhylotypeA sequence type; analogous to an operational taxonomic unit.
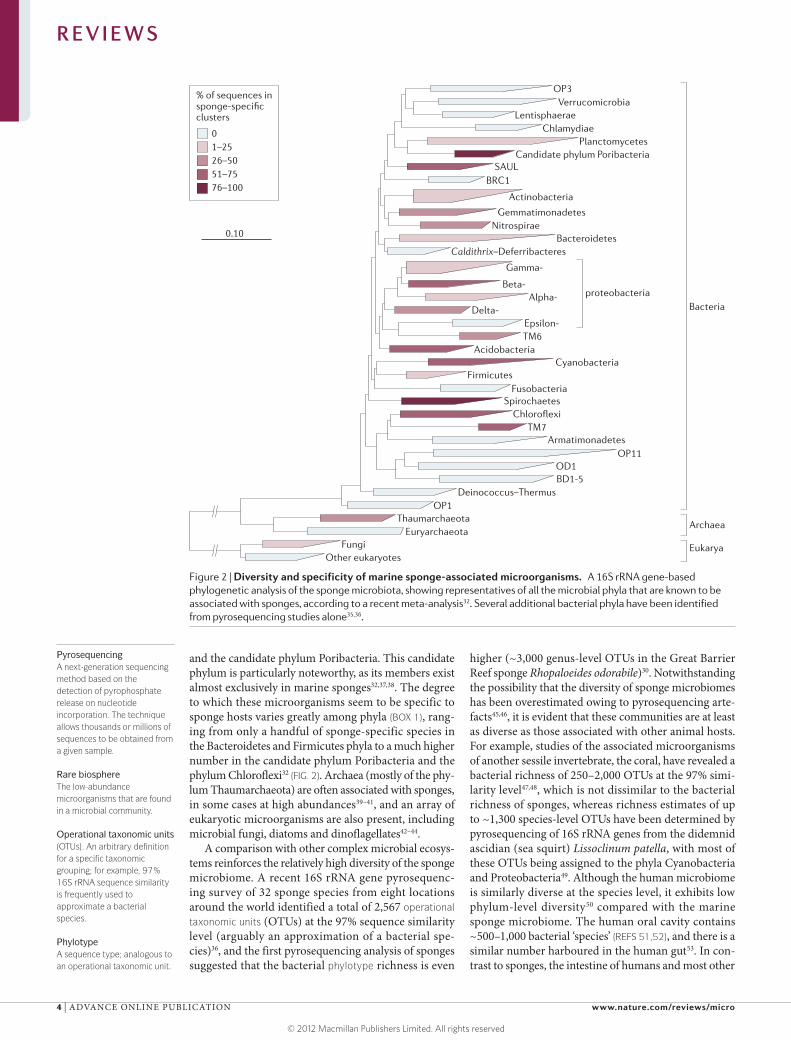
and the candidate phylum Poribacteria. This candidate phylum is particularly noteworthy, as its members exist almost exclusively in marine sponges32,37,38. The degree to which these microorganisms seem to be specific to sponge hosts varies greatly among phyla (BOX 1), rang-ing from only a handful of sponge-specific species in the Bacteroidetes and Firmicutes phyla to a much higher number in the candidate phylum Poribacteria and the phylum Chloroflexi32 (FIG. 2). Archaea (mostly of the phy-lum Thaumarchaeota) are often associated with sponges, in some cases at high abundances39–41, and an array of eukaryotic microorganisms are also present, including microbial fungi, diatoms and dinoflagellates42–44.
A comparison with other complex microbial ecosys-tems reinforces the relatively high diversity of the sponge microbiome. A recent 16S rRNA gene pyrosequenc-ing survey of 32 sponge species from eight locations around the world identified a total of 2,567 operational taxonomic units (OTUs) at the 97% sequence similarity level (arguably an approximation of a bacterial spe-cies)36, and the first pyrosequencing analysis of sponges suggested that the bacterial phylotype richness is even
higher (~3,000 genus-level OTUs in the Great Barrier Reef sponge Rhopaloeides odorabile)30. Notwithstanding the possibility that the diversity of sponge microbiomes has been overestimated owing to pyrosequencing arte-facts45,46, it is evident that these communities are at least as diverse as those associated with other animal hosts. For example, studies of the associated microorganisms of another sessile invertebrate, the coral, have revealed a bacterial richness of 250–2,000 OTUs at the 97% simi-larity level47,48, which is not dissimilar to the bacterial richness of sponges, whereas richness estimates of up to ~1,300 species-level OTUs have been determined by pyrosequencing of 16S rRNA genes from the didemnid ascidian (sea squirt) Lissoclinum patella, with most of these OTUs being assigned to the phyla Cyanobacteria and Proteobacteria49. Although the human microbiome is similarly diverse at the species level, it exhibits low phylum-level diversity50 compared with the marine sponge microbiome. The human oral cavity contains ~500–1,000 bacterial ‘species’ (REFS 51,52), and there is a similar number harboured in the human gut53. In con-trast to sponges, the intestine of humans and most other
Figure 2 | Diversity and specificity of marine sponge-associated microorganisms. A 16S rRNA gene-based phylogenetic analysis of the sponge microbiota, showing representatives of all the microbial phyla that are known to be associated with sponges, according to a recent meta-analysis32. Several additional bacterial phyla have been identified from pyrosequencing studies alone35,36.
R E V I E W S
4 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

HolobiontA host animal and all of its associated microorganisms.
CRISPRs(Clustered regularly interspaced short palindromic repeats). Short repeats that are found in the genomes of many bacteria and archaea, and provide resistance to genetic elements such as viruses and plasmids.
vertebrates is dominated by only two phyla: Firmicutes and Bacteroidetes50,54. On human skin, members of the phyla Proteobacteria and Actinobacteria may also be abundant54,55, but overall the bacterial communities associated with humans seem to exhibit lower evenness (that is, the relative numbers of the different bacteria are highly dissimilar) at phylum level than those associated with sponges.
Emergence and maintenance of diversity. That sponge-associated microbial communities are diverse is beyond question, but what mechanisms are responsible for the emergence and maintenance of this diversity? It is likely that ancient filter-feeding sponges retained seawater-derived ‘food’ bacteria as symbionts, but whether this was essentially a non-selective process that led to the immediate acquisition of the ≥30 bacterial phyla that are known to exist in sponges today, or whether it was a more gradual colonization over hundreds of millions of years, is mere speculation. Exactly why sponges maintain such high microbial diversity is also open to conjecture. For example, why does Xestospongia testudinaria har-bour >100 species of Chloroflexi bacteria36? Does each of these species have a different functional role, or is there extensive functional redundancy in the sponge micro-biome? The lifestyle of the filter-feeding sponge may also come into play, as the continuous exposure of sponges to microorganisms in the surrounding sea water ensures a constant influx of fresh DNA (with potentially novel functions) to the sponge holobiont. It is also important to note that certain environmental stressors, such as increased temperature, can result in the loss of microbial symbionts from sponges (BOX 2).
One potentially important determinant of microbial diversity in sponges is the degree of host specificity — that is, whether microorganisms are associated with only one or a few (specialist) or many (generalist) host species56. Molecular evidence suggests that, despite the occurrence of particular microbial lineages among mul-tiple sponge hosts (that is, sponge-specific clusters; see BOX 1), most bacterial species and species-level OTUs are in fact limited to a single species of host sponge36,56. The recent pyrosequencing analysis of 32 sponge spe-cies from eight locations around the world identified few
bacterial species that are common to more than a hand-ful of sponge species36. Among the 2,567 species-level OTUs that were recorded, only three were detected in at least 70% of tested sponges: an alphaproteo bacterium and two Chloroflexi-affiliated sequence types. The authors suggest that the members of this very small, globally distributed ‘core’ community are probably dis-persed by horizontal transmission through the water column36. Although these findings are somewhat unex-pected, given earlier observations of overlapping bacte-rial communities in different sponge hosts8,31, they do imply an element of selection for specific bacterial types by different sponge species. Further studies are required to establish whether such selection has a prominent role in determining the composition of the sponge microbiome.
Horizontal gene transfer. Horizontal gene transfer (HGT) is a mechanism by which microorganisms can generate diversity in their genomes57. Mobile genetic ele-ments such as transposons and integrons are widespread in nature58,59, and it is likely that HGT is prevalent in the dense microbial communities of many sponges, espe-cially given the probable liberation of bacterial DNA as a result of phagocytotic feeding by sponge cells. To date, there have been no unequivocal cases of HGT among sponge-associated microorganisms or between a micro-organism and the host sponge. A number of potential HGT cases have been identified in the genome of the sponge Amphimedon queenslandica, although an inves-tigation is underway into whether the genome sequence contains these ‘bacterial contaminants’ as a result of sequence data misassembly (S.M.D., unpublished observations).
In addition, although little is known about viruses in sponges24, viral lysis of microbial cells may also contrib-ute to interspecies transfer of genetic material60. Viral abundance in the ocean exceeds bacterial abundance by an order of magnitude61, and sponges are likely to contain extensive communities of bacteriophages and other viruses62. The recently sequenced metagenome of the sponge Cymbastela concentrica revealed a prevalence of CRISPRs (clustered regularly interspaced short palindromic repeats) that may serve to protect
Box 2 | Sponges and microorganisms in a changing global environment
Global climate change is expected to have a direct and substantial impact on marine ecosystems, with global surface temperatures predicted to rise by 1.8–4 °C by 2100 (REF. 141). It is thus plausible that elevated temperatures, and possibly other environmental stressors (for example, eutrophication and sedimentation) will affect the stability of the sponge–microorganism association. Indeed, several studies have reported on the loss of microbial symbionts and a system breakdown on exposure to thermal stress142–144. The Great Barrier Reef sponge Rhopaloeides odorabile does not tolerate temperatures above 33 °C, which result in loss of microbial symbionts and sponge death143. The effects of increased temperatures are also evident in the animal itself, which responds with rapid molecular shutdown of many indicator genes and increased expression of heat shock proteins144,145. Although some cases of sponge diseases may be unrelated to microbial pathogens146,147, reports of sponge disease have nevertheless increased markedly over the past two decades (reviewed in REF. 98). In most cases, the aetiological agent (or agents) of sponge diseases have not been elucidated. One notable exception is an alphaproteobacterium that was isolated from an infected sample of R. odorabile148. This bacterium degrades collagenous spongin fibres to result in tissue necrosis of almost the entire sponge surface. Importantly, this work has altered our perception of the influence of environmental stressors, suggesting that sponge–microorganism symbioses are sensitive indicators of global environmental change.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 5
© 2012 Macmillan Publishers Limited. All rights reserved
HeterotrophicPertaining to an organism: requiring dissolved organic matter for its carbon and energy sources.
AutotrophicPertaining to an organism: growing on carbon dioxide as the sole carbon source.
sponge-associated bacteria from viral attack63. Notably, the genome of A. queenslandica encodes core compo-nents of the RNAi pathway, which is known to target invading viral RNA64.
Genomic insights into symbiont functionAlthough much work has focused on the functional genomic repertoire of sponge symbionts, the surface has barely been scratched. The inability to cultivate most symbionts is a substantial constraint to the field, as is the lack of an acknowledged experimental model sponge65, the absence of an accessible laboratory host assay and the failure to obtain any substantial sponge growth in seawater aquaria. Although laboratory and field experiments have provided many useful insights into the uptake and excretion rates of carbon and nitro-gen (reviewed in REFS 8,24), our understanding of these processes and of many others has been greatly enhanced through information revealed by genomic (including single-cell methods) and proteomic approaches. In the following section, we focus on those metabolic pathways that have been identified by genomics.
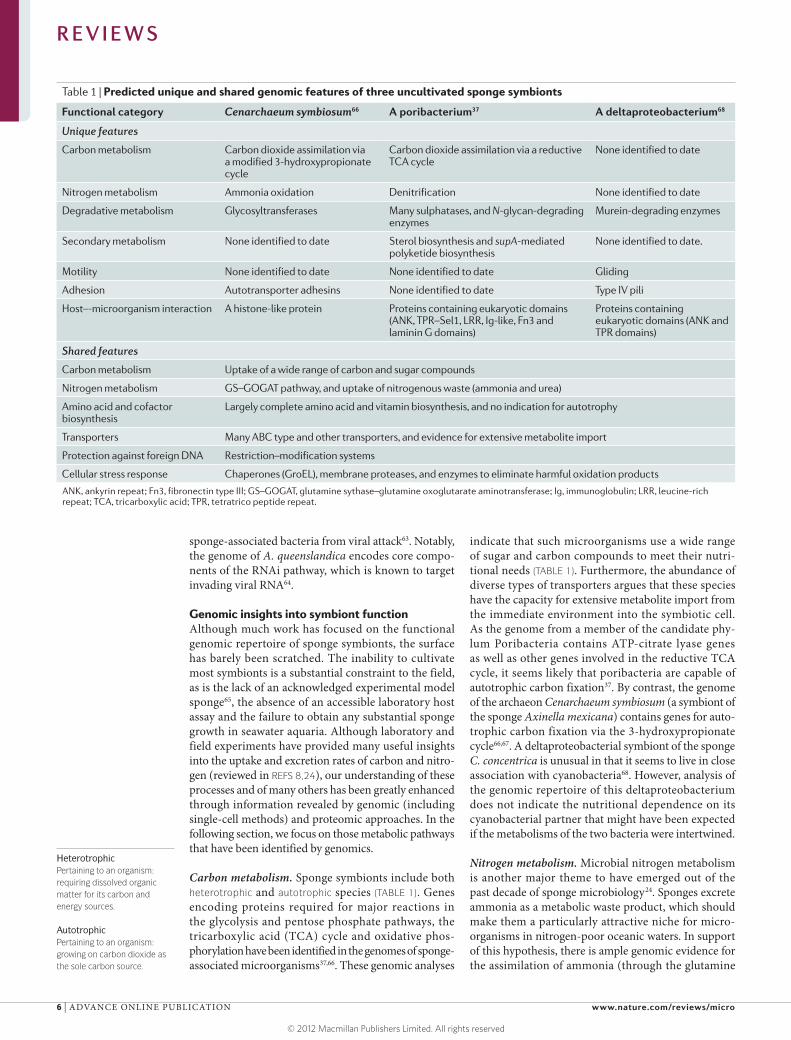
Carbon metabolism. Sponge symbionts include both heterotrophic and autotrophic species (TABLE 1). Genes encoding proteins required for major reactions in the glycolysis and pentose phosphate pathways, the tricarboxylic acid (TCA) cycle and oxidative phos-phorylation have been identified in the genomes of sponge- associated microorganisms37,66. These genomic analyses
indicate that such microorganisms use a wide range of sugar and carbon compounds to meet their nutri-tional needs (TABLE 1). Furthermore, the abundance of diverse types of transporters argues that these species have the capacity for extensive metabolite import from the immediate environment into the symbiotic cell. As the genome from a member of the candidate phy-lum Poribacteria contains ATP-citrate lyase genes as well as other genes involved in the reductive TCA cycle, it seems likely that poribacteria are capable of autotrophic carbon fixation37. By contrast, the genome of the archaeon Cenarchaeum symbiosum (a symbiont of the sponge Axinella mexicana) contains genes for auto-trophic carbon fixation via the 3-hydroxypropionate cycle66,67. A deltaproteobacterial symbiont of the sponge C. concentrica is unusual in that it seems to live in close association with cyanobacteria68. However, analysis of the genomic repertoire of this deltaproteobacterium does not indicate the nutritional dependence on its cyanobacterial partner that might have been expected if the metabolisms of the two bacteria were intertwined.
Nitrogen metabolism. Microbial nitrogen metabolism is another major theme to have emerged out of the past decade of sponge microbiology24. Sponges excrete ammonia as a metabolic waste product, which should make them a particularly attractive niche for micro-organisms in nitrogen-poor oceanic waters. In support of this hypothesis, there is ample genomic evidence for the assimilation of ammonia (through the glutamine
Table 1 | Predicted unique and shared genomic features of three uncultivated sponge symbionts
Functional category Cenarchaeum symbiosum66 A poribacterium37 A deltaproteobacterium68
Unique features
Carbon metabolism Carbon dioxide assimilation via a modified 3-hydroxypropionate cycle
Carbon dioxide assimilation via a reductive TCA cycle
None identified to date
Nitrogen metabolism Ammonia oxidation Denitrification None identified to date
Degradative metabolism Glycosyltransferases Many sulphatases, and N-glycan-degrading enzymes
Murein-degrading enzymes
Secondary metabolism None identified to date Sterol biosynthesis and supA-mediated polyketide biosynthesis
None identified to date.
Motility None identified to date None identified to date Gliding
Adhesion Autotransporter adhesins None identified to date Type IV pili
Host–-microorganism interaction A histone-like protein Proteins containing eukaryotic domains (ANK, TPR–Sel1, LRR, Ig-like, Fn3 and laminin G domains)
Proteins containing eukaryotic domains (ANK and TPR domains)
Shared features
Carbon metabolism Uptake of a wide range of carbon and sugar compounds
Nitrogen metabolism GS–GOGAT pathway, and uptake of nitrogenous waste (ammonia and urea)
Amino acid and cofactor biosynthesis
Largely complete amino acid and vitamin biosynthesis, and no indication for autotrophy
Transporters Many ABC type and other transporters, and evidence for extensive metabolite import
Protection against foreign DNA Restriction–modification systems
Cellular stress response Chaperones (GroEL), membrane proteases, and enzymes to eliminate harmful oxidation products
ANK, ankyrin repeat; Fn3, fibronectin type III; GS–GOGAT, glutamine sythase–glutamine oxoglutarate aminotransferase; Ig, immunoglobulin; LRR, leucine-rich repeat; TCA, tricarboxylic acid; TPR, tetratrico peptide repeat.
R E V I E W S
6 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

Ankyrin repeats(ANKs). Common eukaryotic structural protein motifs that occur in functionally diverse proteins and mediate protein–protein interactions.
Tetratrico peptide repeatsCommon structural protein motifs that mediate protein–protein interactions and are frequently involved in the assembly of multiprotein complexes.
Leucine-rich repeatsStructural protein motifs that are unusually rich in the hydrophobic amino acid leucine and are involved in the formation of protein–protein interactions.
Quorum sensingA chemical language by which bacteria communicate in and across populations through the use of small diffusible molecules. Bacterial production of quorum sensing molecules, as well as the bacterial response to these molecules, is correlated with population density.
synthase–glutamine oxoglutarate aminotransferase (GS–GOGAT) pathway63) and nitrite37 by sponge-associated microorganisms (TABLE 1). Furthermore, urease-encoding gene clusters, urea transporters and accessory genes have been identified in two sponge symbiont genomes37,66. Urea, a by-product of nucleic acid and amino acid degradation, is probably present in the sponge environment at low concentrations and might thus serve as an alternative nitrogen source to ammonia, nitrate and nitrite. Both aerobic (nitrifica-tion) and anaerobic (denitrification) branches of the nitrogen cycle occur in sponges24. Ammonia oxidation, the first step of nitrification, is catalysed by bacterial or archaeal ammonia monooxygenases, encoded by amo genes. The study of ammonia oxidation in sponges pro-vides an elegant example of the effective combination of molecular genomics-, transcriptomics- and proteomics-derived data with physiological experiments and field studies41,69–74. As a result of these investigations, we now know that ammonia-oxidizing archaea (AOAs) and ammonia-oxidizing bacteria (AOBs) are widespread in sponges from temperate73, tropical71 and cold-water regions41; that AOAs are vertically transmitted through sponge larvae72; that AOAs are metabolically active and undergo seasonal cycling73; that the archaeal amoA genes are highly expressed in sponges (as inferred by metatranscriptomics)69; and that AOAs, rather than AOBs, are the main drivers of sponge nitrification, at least in cold-water sponges41. In addition to nitrifica-tion, several genes with products that are important for denitrification have been identified by a single-cell genomics analysis of a member of the candidate phylum Poribacteria37, and proteins related to both nitrification and denitrification have been identified through a com-bined metagenomic and metaproteomic analysis of the microbial consortium in C. concentrica75. The efficient recycling of sponge-excreted nitrogenous wastes is one obvious benefit for symbionts in return for an existence in sponge tissues.
Vitamin biosynthesis. Vitamins have also come under scrutiny, and the three genomic studies of sponge symbionts have reported on the presence of genes encoding enzymes involved in the synthesis of cobala-min (vitamin B12)
37,63,66, riboflavin (vitamin B2)66,68, biotin
(vitamin B7)66, thiamine (vitamin B1)
66 and pyridoxine (a form of vitamin B6)
66. Interestingly, the metagenome of the human distal-gut microbiome is substantially enriched for genes encoding enzymes with roles in vita-min biosynthesis76, and a functional requirement for cobalamin biosynthesis influences microbial coloniza-tion77. It is therefore conceivable not only that microbial symbionts supplement the diet of their sponge hosts with vitamins, but also that vitamin biosynthesis confers a selective advantage on vitamin-producing symbionts.
Symbiosis factors. The possession of eukaryotic-like proteins is a characteristic feature of bacterial pathogens that have a long history of co-evolution with their hosts78. These eukaryotic-domain-containing proteins mediate protein–protein interactions in many biological processes,
including those pertaining to an intracellular lifestyle. Genes encoding such proteins, which contain motifs such as ankyrin repeats (ANKs), tetratrico peptide repeats (TPRs) and leucine-rich repeats (LRRs), are also represented in the genomes of two sponge symbionts37,63,68,75 (TABLE 1).
Originally considered to be eukaryote specific, ANK-containing proteins have recently been found in high abundance in the genomes of many intracellular pathogens79,80, including endosymbionts of amoeba81. The function of these proteins is best illustrated by the ANK-containing proteins of Legionella pneumophila, the causative agent of Legionnaire’s disease82. In this disease, ANK-containing proteins are secreted through a type IV secretion system (T4SS) into eukaryotic cells, where they interfere with host cell functions such as polyubiquitylation83 and vesicular transport79. Mutations in ankB result in a defect in L. pneumophila replication within both macrophages and proto-zoan hosts83,84. Interestingly, in Coxiella burnetii (the causative agent of Q fever), AnkG prevents pathogen-induced apoptosis, and this activity is interpreted as a strategy to secure the pathogen’s survival in mammalian cells85. ANKs may, by analogy, allow the sponge symbi-onts to interfere with host phagocytosis prior to, during or after symbiont uptake by the sponge cell. The TPR-containing proteins, in particular the Sel1 subclass, might also mediate bacterium–eukaryote interactions — for example, allowing entry of L. pneumo phila into epithelial cells and macrophages, or regulating exopoly-saccharide synthesis in Pseudomonas aeruginosa86,87. The genes encoding ANK- or TPR-containing pro-teins are located in close proximity in the genome of the C. concentrica-associated deltaproteobacterium68, suggesting that this particular genomic region encodes a bona fide symbiosis factor.
Several genes identified in the poribacterial genome encode products containing eukaryotic-like domains such as fibronectin type III (Fn3) domains, laminin G domains and immunoglobulin-like domains37 (TABLE 1). There is little precedent for the presence of these pro-tein domains in other bacteria. Fn3 domains have been found in the extracellular glycohydrolases that aid in cellulose degradation by Clostridium thermocellum88. Finding such motifs in a sponge symbiont genome is therefore noteworthy, as the sponge extracellular matrix is rich in proteoglycans, adhesive glycoproteins (laminin-like subunits and fibronectin) and other structural proteins (fibrillar collagen and spongin)89–91. It is thus tempting to speculate that these poribacterial genes function in modifying or utilizing structures in the sponge extracellular matrix. Microorganisms that are capable of quorum sensing92 have also been isolated from marine sponges93–96; for example, the genome sequence of Ruegeria sp. str. KLH11 revealed the presence of two quorum sensing circuits94. The types of signalling mole-cules involved in these bacterial communication circuits may also facilitate communication between microorgan-isms and their hosts97. Indeed, gene families that have been implicated in inter-kingdom signalling, including those encoding receptor tyrosine kinases, epidermal growth factor receptors and guanine nucleotide-binding
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 7
© 2012 Macmillan Publishers Limited. All rights reserved
Extracellular(secreted)
Site of PRR localization
PRR gene family
Sponge genestructures
Potentialligand
Human gene structures
MAMPsPAMPsLPS?
MAMPsPAMPsLPS?
MAMPsPAMPsDAMPsPGN?
SRCRs(SR-As)
TIRsTLRs
NLRsIntracellular
Protein domains
Lysyl oxidase
SRCR
LRR
Ig-like
TIR
NACHT
NAD
DEATH
CARD
PYD
Membrane-bound
Nature Reviews | Microbiology
protein-coupled receptors, are highly expanded in the A. queenslandica genome14. Investigating the role of quorum sensing in microorganism–microorganism communication, and possibly also in microorganism–host communication, is a tantalizing avenue for future research.
Genomic insights into sponge immunityAs a result of their sessile marine filter-feeding lifestyle, sponges are continuously challenged to differentiate between and respond appropriately to beneficial micro-organisms — both food sources destined for ingestion and symbionts destined for retention — and harmful pathogens10,11,98. This challenge is met primarily by the innate immune system, which was discovered in sponges more than a century ago99. The near-complete genome of A. queenslandica14 has provided the first view of a full complement of sponge innate immunity genes that regulate host–microorganism interactions.
Pattern recognition receptors. The first points of contact between a sponge host and incoming microorganisms are generally genome-encoded pattern recognition recep-tors (PRRs)100, which have the crucial task of recog nizing microbial ligands known as microbial- or pathogen- associated molecular patterns (MAMPs or PAMPs, respectively)101. Common examples of ligands that are recognized by PRRs are lipopolysaccharide (LPS) of Gram-negative bacteria, peptidoglycan of both Gram-negative and Gram-positive bacteria, and the mannose sugar component of microbial glycolipids and glyco proteins100. Binding of a microbial ligand to a PRR typically trig-gers a signal transduction cascade that leads to tran-scription of immune response effector genes encoding products such as antibacterial proteins101. Some animal immunity genes originated early in eukaryote evolution, reflecting the ancient need for the discrimination of self from non-self, whereas others are restricted to the eumetazoans, a group that excludes sponges12,14. However, sponges possess a surprising number of the innate immunity genes that are found in eumetazoans.
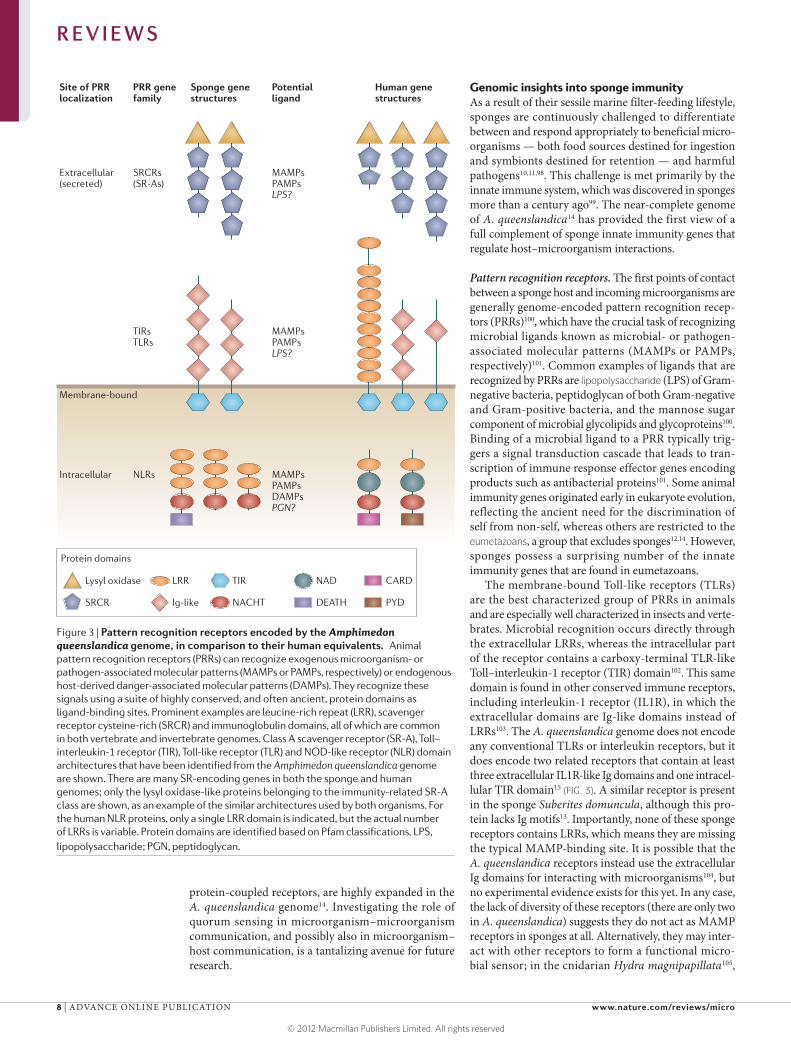
The membrane-bound Toll-like receptors (TLRs) are the best characterized group of PRRs in animals and are especially well characterized in insects and verte-brates. Microbial recognition occurs directly through the extracellular LRRs, whereas the intracellular part of the receptor contains a carboxy-terminal TLR-like Toll–interleukin-1 receptor (TIR) domain102. This same domain is found in other conserved immune receptors, including interleukin-1 receptor (IL1R), in which the extracellular domains are Ig-like domains instead of LRRs103. The A. queenslandica genome does not encode any conventional TLRs or interleukin receptors, but it does encode two related receptors that contain at least three extracellular IL1R-like Ig domains and one intracel-lular TIR domain15 (FIG. 3). A similar receptor is present in the sponge Suberites domuncula, although this pro-tein lacks Ig motifs13. Importantly, none of these sponge receptors contains LRRs, which means they are missing the typical MAMP-binding site. It is possible that the A. queenslandica receptors instead use the extracellular Ig domains for interacting with microorganisms104, but no experimental evidence exists for this yet. In any case, the lack of diversity of these receptors (there are only two in A. queenslandica) suggests they do not act as MAMP receptors in sponges at all. Alternatively, they may inter-act with other receptors to form a functional micro-bial sensor; in the cnidarian Hydra magnipapillata105,
Figure 3 | Pattern recognition receptors encoded by the Amphimedon queenslandica genome, in comparison to their human equivalents. Animal pattern recognition receptors (PRRs) can recognize exogenous microorganism- or pathogen-associated molecular patterns (MAMPs or PAMPs, respectively) or endogenous host-derived danger-associated molecular patterns (DAMPs). They recognize these signals using a suite of highly conserved, and often ancient, protein domains as ligand-binding sites. Prominent examples are leucine-rich repeat (LRR), scavenger receptor cysteine-rich (SRCR) and immunoglobulin domains, all of which are common in both vertebrate and invertebrate genomes. Class A scavenger receptor (SR-A), Toll–interleukin-1 receptor (TIR), Toll-like receptor (TLR) and NOD-like receptor (NLR) domain architectures that have been identified from the Amphimedon queenslandica genome are shown. There are many SR-encoding genes in both the sponge and human genomes; only the lysyl oxidase-like proteins belonging to the immunity-related SR-A class are shown, as an example of the similar architectures used by both organisms. For the human NLR proteins, only a single LRR domain is indicated, but the actual number of LRRs is variable. Protein domains are identified based on Pfam classifications. LPS, lipopolysaccharide; PGN, peptidoglycan.
R E V I E W S
8 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

LipopolysaccharideA large endotoxic molecule consisting of a lipid and a polysaccharide; the major component of the outer membrane of Gram-negative bacteria. LPS induces a strong immune response in animals.
EumetazoansThe ‘true’ animals, as defined by having a nervous system and true tissues; this group includes cnidarians, ctenophores and bilaterians.
BilaterianAn animal that has three germ layers (endoderm, mesoderm and ectoderm) and bilateral symmetry; this group includes all animals except for sponges, ctenophores and cnidarians.
Danger-associated molecular patterns(DAMPS). Molecules that are released by stressed or damaged cells and act as endogenous signals to initiate a non-infectious repair (or inflammatory) response. Also called damage-associated molecular patterns.
Complement systemThe part of the vertebrate innate immune system that complements the activity of antibodies by opsonizing bacteria and inducing inflammatory responses which help to fight infection. Several complement-like components have also been identified in many invertebrates.
Secondary metabolitesMetabolites that are not essential for the survival of the organism in which they are found (unlike primary metabolites, such as fatty acids, amino acids, and so on, which are essential).
EpibiontsOrganisms that live on the surface of another living organism.
PolyketidesSecondary metabolites that are biosynthesized from short acyl-CoA units and often exhibit potent bioactivities. Examples are erythromycin and tetracycline.
for example, a TIR domain-containing protein that lacks LRRs interacts with an LRR-containing protein to mediate the induction of antimicrobial peptides in response to PAMPs105. Of interest in this context are three genes encoding LPS-binding-protein-like (LBL) proteins in the A. queenslandica genome; the products of these genes seem to be structurally capable of detect-ing and/or removing the LPS of Gram-negative bacte-ria, either directly or via interactions with TLR-related receptors14,15.
Although the NOD-like receptor (NLR)-encoding genes are not as well characterized as the TLR- encoding genes, they are the most diverse family of eukary otic immune system genes106. NLRs have an important role in sensing a wide range of microorgan-isms106–108. Most NLRs are located in the cytosol, where they probably respond to bacteria that manage to invade the cell and to bacterial products remaining after phago-cytosis106. However, they can also be activated by non-invasive bacteria, and in these cases it is unclear how the bacterial ligands come into contact with the intracellular receptors. Early work in this area — although none has yet been carried out in sponges — has raised the intrigu-ing possibility of cell- or species-level variation in these entry mechanisms; there is evidence to suggest that NLR activators, such as peptidoglycan fragments, are trans-ported into host cells via bacterial type IV secretion sys-tems and pore-forming toxins, and via host cell pannexin hemichannels and peptide transporter channels. The A. queenslandica genome encodes seven putative NLRs, all containing a NACHT domain in combination with variable numbers and classes of LRRs (J. Bayes and S.M.D., unpublished observations) (FIG. 3). No NLRs have yet been reported from other sponges, and nothing is known about specific functions in A. queenslandica or about which tissues or cells specifically express these proteins. In bilaterian animals, NLRs primarily detect pep tides derived from the degradation of bacterial peptido glycan106 and are therefore differentially sensitive to Gram-positive and Gram-negative species, owing to the differing pep-tidoglycan contents of these two groups. Peptidoglycan is not recognized by TLRs108, so NLRs could provide an alternative line of defence in sponges, in which TLR sig-nalling seems to be reduced. Importantly, NLRs function not only to detect exogenous MAMPs, but also to sense endogenous danger-associated molecular patterns (DAMPs) that are produced by the host in response to tissue damage and cellular stress106. DAMPs thus indicate the presence of pathogenic infection or the occurrence of an environ-mental insult, and their release promotes the recruitment of immune cells to the damaged area to initiate repair. The capacity to detect both MAMPs and DAMPs helps the host to discriminate between pathogenic and symbi-otic bacteria109, suggesting that NLRs have a pivotal role in sponge–microorganism interactions.
A third suite of PRRs of likely importance to sponge–microorganism crosstalk is the scavenger receptor cysteine-rich (SRCR) proteins, characterized by one or several repeats of the ancient SRCR domain110. Of particular interest are the macrophage class A scav-enger receptors (SR-A proteins), which have already
been shown to mediate endocytic clearance of bacte-ria, environmental particles and DNA in vertebrates110. In humans, SR-A proteins bind to the LPS on multiple bacterial species, resulting in phagocytosis111. At least one SR-A protein has been identified from the sponge Geodia cydonium112,113, and a gene encoding an SRCR-containing protein in the sponge Petrosia ficiformis is upregulated in the presence of cyanobacterial symbi-onts114. The A. queenslandica genome sequence encodes hundreds of SRCR domains, a few of which have simi-larity to secreted SR-A proteins that are implicated in vertebrate innate immunity (S.M.D., unpublished obser-vations, and REF. 111) (FIG. 3). Other A. queenslandica SRCR domains, from the SR-B class, are associated with complement control domains (S.M.D., unpublished observations) similar to those used by the verte-brate complement system, although no recognizable comple ment pathway genes have been detected in the A. queenslandica genome14. Because the SRCR super-family seems to be capable of generating an enormous diversity of multi-ligand and multifunctional receptors, as would be required by an organism for meeting the challenges of hosting an immensely diverse microbial community, these receptors are ripe for future study in sponge–microorganism systems.
A cooperative chemical factoryOne of the most remarkable properties of marine sponges is their capacity to produce a diverse range of natural products. Sponges are among the richest known sources of biologically active secondary metabolites and have yielded more compounds than any other group of marine organisms16. The fact that many of these substances act as protective agents against predators and epibionts5 may have contributed to the evolutionary and ecological success of sponges. Numerous sponge-derived natural products exhibit promising activities against various diseases, most notably cancer. For instance, eribulin, a synthetic analogue of halichondrin B from Halichondria and Lissodendoryx spp. sponges, was recently approved as a drug for the treatment of metastatic breast cancer115. Its long history of development involved a veritable tour de force: establishing a 90-step protocol for total syn-thesis of the compound116, collecting 1 metric tonne of sponges to secure 300 milligrams of the natural product for preclinical trials117, and synthesizing and testing hun-dreds of structurally complex variants. Notwithstanding this single success story, the challenge of providing long-term supplies for structurally complex metabolites can represent a crucial bottleneck in sponge-based drug development.
In light of these obstacles, the possibility that at least some sponge compounds are of bacterial origin offers an attractive opportunity to create sustainable biotech-nological production systems. This could be realized either by cultivating the producer itself or by transfer-ring the biosynthetic genes into a culturable surrogate host. The observation that many substances isolated from sponges structurally resemble typical products of microbial pathways — in particular, complex polyketides or non-ribosomal peptides — provided initial support for
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 9
© 2012 Macmillan Publishers Limited. All rights reserved
the symbiont hypothesis118. Substantial efforts have therefore been invested in pinpointing the metabolic sources of the natural products that are produced during sponge–microorganism associations. These efforts have included strategies based on cultivation17, mechanical cell separation119–123 and microscopy121,124.
In addition, a number of recent studies have begun to investigate the synthetic pathways at the genetic and genomic levels. This strategy not only provides an unam-biguous link between the natural-product chemistry and producer cell type (sponge or symbiont), but also provides the genetic information that is necessary for heterologous
Table 2 | Natural products that are suspected or known to be produced by bacterial symbionts of sponges
Chemical structure Natural products Suspected bacterial source Host sponges Ref.
Brominated biphenylethers
Oscillatoria spongeliae Lamellodysidea spp. 121
Chlorinated peptides O. spongeliae Lamellodysidea spp. 135
Theopalauamide ‘Candidatus Entotheonella palauensis’
Theonella swinhoei 123
Swinholide A Unicellular bacterium T. swinhoei 123
Onnamide A Unidentified bacterium T. swinhoei 127
Psymberin Unidentified bacterium Psammocinia bulbosa
126
Br
Br
BrBrBrBrBr
O OBr
Br
OH OHOMe
N
N
Me
O
O CCl3
Cl3C
Me
Cl3C N CCl3
O
Me
O NH
N
S
BrHN
O
HN
O
NHHN
NH
HN NH
NH
HNNH
HN
H2NOC
H2NOC
OHO
O
O
HOO
Ph
HO
O PhO
OH
O
OHO
O
N
N
NHO
HO
+
-
OHO
HO
OHHO
OOC
O
O
O
O
O
O
OMe
OH OH OH OMe
OH
MeO
OHOHOHMeO
O
O
HO
OHN
OMeO
OMeO OH
OH
O O
O
NH
COOHHN
NH2
HN
OH
OH
OH
OH
O
O
O
O
OMe
OMe
MeHN
R E V I E W S
10 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

Non-ribosomal peptidesPeptides that are assembled by large multifunctional enzymes (termed non-ribosomal- peptide synthetases) and often contain non-proteinogenic amino acids. Many of these compounds (for example, cyclosporine and daptomycin) are of biomedical relevance.
CARD–FISH(Catalysed reporter deposition–fluorescence in situ hybridization). A technique that uses horseradish peroxi-dase-labelled oligonucleotide probes to obtain signals in samples with low-abundance targets. The signal is generated after incubation with fluorescently labelled tyramine, which is deposited at the labelling site by enzymatic polymerization.
biotechnological production of the compounds. However, the complexity and diversity of homologous genes asso-ciated with sponges of biomedical interest still represent major challenges for identification of biosynthetic genes. The success of gene-based strategies therefore depends on the ability to reliably predict biosynthetic-gene sequences from natural-product structures and to isolate these genes from vast metagenomic clone libraries. This process is most straightforward with complex polyketides and non-ribosomal peptides, which are synthesized in an assembly- line-like manner by multidomain enzymes; in these cases, a tight correlation usually exists between enzymatic archi-tecture and the chemical structure of the product, mak-ing it possible to identify probable biosynthetic genes on the basis of a knowledge of the compound structure, and vice versa125. This principle was recently used to search the total metagenomes of the sponges Theonella swinhoei and Psammocinia bulbosa and identify the genes that are important for the production of the highly potent anti-tumor polyketides onnamide A and psymberin, respec-tively126,127 (TABLE 2). The genes belong to a bacterial polyketide synthase (PKS) family, are clustered in oper-ons and are located in genomic regions that have typical bacterial features, suggesting a symbiont origin for these metabolites. Interestingly, both PKSs fall into an enzy-matic subgroup, termed trans-acyltransferase PKSs128, that an increasing number of studies shows to be prevalent among symbiotic bacteria. Other examples of enzymes that have been identified in this way include the PKSs for pederin from a Pseudomonas sp. symbiont of beetles129, bryostatin from the bryozoan-associated symbiont ‘Candidatus Endobugula sertula’ (REF. 130), rhizoxin from fungus-associated Burkholderia spp. symbionts131,132 and patellazole from an unidentified tunicate symbiont133. Although the functional relevance of this correlation between enzymatic architecture and product chemi-cal structure remains unexplained, it is a noteworthy correlation, as it allows us to develop more reliable bio-synthetic predictions, to search metagenomes in a more targeted and streamlined manner and to develop hosts that are specifically adapted to express symbiont-derived PKS genes126,134.
One of the disadvantages of metagenomic gene tar-geting is that it rarely provides information about the exact taxonomic identity of the producer, because only the biosynthetic loci and adjacent regions are obtained. Several studies have started to address this issue by applying techniques that localize genes at the single-cell level. The earliest example of such a study in sponge research was carried out for chlorinated peptides from the sponge Lamellodysidea herbacea135 (TABLE 2). A halo-genase gene that is thought to be involved in the attach-ment of chlorine residues in these peptides was linked to the cyanobacterium Oscillatoria spongeliae using CARD–FISH (catalysed reporter deposition–fluorescence in situ hybridization). Recently, single-cell genome amplification and sequencing methods have provided exciting new perspectives for studying the chemistry of uncultivated consortia at unprecedented depth and resolution136,137. Using these techniques, PKS-like genes that are thought to be involved in methyl-branched fatty
acid biosynthesis have been linked to members of the unculti vated candidate phylum Poribacteria37,138, and non-ribosomal peptide synthetase genes have been linked to a member of the phylum Chloroflexi138. In a similar way, several natural products have been unam-biguously associated with cyanobacteria in an intimately mixed free-living bacterial assemblage139,140.
From all of these studies, the general picture that emerges is one in which bacterial symbionts are of cen-tral importance in the chemistry of sponges. This could lead to a potential solution to the problem of supply-ing many pharmacologically important ‘sponge’ natural products, as the bacteria that produce these products could be grown on a large scale. Do all secondary metabolites found in sponges originate from bacteria? Circumstantial evidence suggests that this scenario is unlikely. For example, the structure of halichondrin B resembles the structure of typical polyether ladder polyketides isolated from dinoflagellates, indicating a related source for these compounds. In addition, the common presence of terpenoid secondary metabolites in sponges stands in contrast to the relative scarcity of such compounds in free-living bacteria and suggests a more prominent eukaryotic contribution in sponge– microorganism consortia. New high-throughput sequencing methods that can achieve sufficient cov-erage of sponge metagenomes at low cost will have an important future role in unravelling the chemistry and biotechnological potential of sponge-associated bacteria.
ConclusionsThe ecological, evolutionary and biotechnological rele-vance of marine sponges and their associated microbiota has spurred much recent research in this area. Although some pressing issues remain unresolved (such as the resistance of sponge symbionts to cultivation, the absence of a generally accepted sponge model species and of a tractable laboratory assay, and the inherent difficulties of studying sponges in seawater aquaria), sponges are now considered to be a valuable system for the study of high-diversity marine host–microorganism associations, much like the human gut for terrestrial systems. With the aid of next-generation sequencing technologies and the greater sequencing depth that they afford, a clearer picture of microbial diversity in these hosts is now being achieved, and the factors that influence this diversity are being iden-tified. Importantly, the metabolic capabilities of sponge-associated microorganisms are also being clarified, largely owing to metagenomic, metaproteogenomic and single-cell genomics studies of uncultivated sponge symbionts. The potential for novel drugs remains a major research focus, and metagenomics promises to revolutionize the field of marine-compound-based drug development by enabling biotechnological access to pharmacologically important host- and symbiont-derived compounds. Finally, the recently sequenced A. queenslandica genome is providing new insights into sponge–microorganism interactions and metazoan evolution in general. In con-cert with genomic studies of the symbionts themselves, the sponge genome offers a powerful tool for elucidating how these intimate partnerships are formed.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 11
© 2012 Macmillan Publishers Limited. All rights reserved

1. Bell, J. J. The functional roles of marine sponges. Estuar. Coast. Shelf Sci. 79, 341–353 (2008).
2. Dayton, P. K. Interdecadal variation in an antarctic sponge and its predators from oceanographic climate shifts. Science 245, 1484–1486 (1989).
3. Bell, J. J. & Smith, D. Ecology of sponge assemblages (Porifera) in the Wakatobi region, south-east Sulawesi, Indonesia: richness and abundance. J. Mar. Biol. Assoc. UK 84, 581–591 (2004).
4. Brusca, R. C. & Brusca, G. J. Invertebrates 2nd edn (Sinauer Associates, 2003).
5. Pawlik, J. R. The chemical ecology of sponges on Caribbean reefs: natural products shape natural systems. Bioscience 61, 888–898 (2011).
6. Li, C. W., Chen, J. Y. & Hua, T. E. Precambrian sponges with cellular structures. Science 279, 879–882 (1998).
7. Love, G. D. et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 457, 718–721 (2009).
8. Taylor, M. W., Radax, R., Steger, D. & Wagner, M. Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 71, 295–347 (2007).An excellent, comprehensive review of sponge microbiology.
9. Vacelet, J. & Donadey, C. Electron microscope study of the association between some sponges and bacteria. J. Exp. Mar. Biol. Ecol. 30, 301–314 (1977).
10. Wilkinson, C. R., Garrone, R. & Vacelet, J. Marine sponges discriminate between food bacteria and bacterial symbionts: electron microscope radioautography and in situ evidence. Proc. R. Soc. Lond. B 220, 519–528 (1984).An early paper in sponge microbiology that is noteworthy for its thought-provoking hypotheses, which are stimulating contemporary discussions.
11. Wehrl, M., Steinert, M. & Hentschel, U. Bacterial uptake by the marine sponge Aplysina aerophoba. Microb. Ecol. 53, 355–365 (2007).
12. Müller, W. E. G. & Müller, I. M. Origin of the metazoan immune system: identification of the molecules and their functions in sponges. Integr. Comp. Biol. 43, 281–292 (2003).
13. Wiens, M. et al. Toll-like receptors are part of the innate immune defense system of sponges (Demospongiae: Porifera). Mol. Biol. Evol. 24, 792–804 (2007).
14. Srivastava, M. et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 466, 720–726 (2010).The first, and still only, complete genome from a sponge reveals that most gene families which are characteristic of true animals, including humans, were already present in the last common ancestor of all animals.
15. Gauthier, E. A., Du Pasquier, L. & Degnan, B. M. The genome of the sponge Amphimedon queenslandica provides new perspectives into the origin of Toll-like and interleukin 1 receptor pathways. Evol. Dev. 12, 519–533 (2010).
16. Blunt, J. W., Copp, B. R., Munro, M. H. G., Northcote, P. T. & Prinsep, M. R. Marine natural products. Nat. Prod. Rep. 28, 196–268 (2011).
17. Piel, J. Bacterial symbionts: prospects for the sustainable production of invertebrate-derived pharmaceuticals. Curr. Med. Chem. 13, 39–50 (2006).
18. Sipkema, D. et al. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (gellius) sp. Appl. Environ. Microbiol. 77, 2130–2140 (2011).
19. Bright, M. & Bulgheresi, S. A complex journey: transmission of microbial symbionts. Nature Rev. Microbiol. 8, 218–230 (2010).
20. Wilson, A. C. et al. Genomic insight into the amino acid relations of the pea aphid, Acyrthosiphon pisum, with its symbiotic bacterium Buchnera aphidicola. Insect Mol. Biol. 19, 249–258 (2010).
21. Dale, C. & Moran, N. A. Molecular interactions between bacterial symbionts and their hosts. Cell 126, 453–465 (2006).
22. Nyholm, S. V. & McFall-Ngai, M. J. The winnowing: establishing the squid–Vibrio symbiosis. Nature Rev. Microbiol. 2, 632–642 (2004).A brilliant example of the intricate interactions that can evolve between animal host and bacterial symbiont.
23. Schmitt, S., Angermeier, H., Schiller, R., Lindquist, N. & Hentschel, U. Molecular microbial diversity survey of sponge reproductive stages and mechanistic insights into vertical transmission of microbial symbionts. Appl. Environ. Microbiol. 74, 7694–7708 (2008).
24. Webster, N. S. & Taylor, M. W. Marine sponges and their microbial symbionts: love and other relationships. Environ. Microbiol. 14, 335–346 (2012).
25. Won, Y.-J., Jones, W. J. & Vrijenhoek, R. C. Absence of cospeciation between deep-sea mytilids and their thiotrophic endosymbionts. J. Shellfish Res. 27, 129–138 (2008).
26. Usher, K. M. & Ereskovsky, A. V. Larval development, ultrastructure and metamorphosis in Chondrilla australiensis Carter, 1873 (Demospongiae, Chondrosida, Chondrillidae). Invertebr. Reprod. Dev. 47, 51–62 (2005).
27. Schmitt, S., Weisz, J. B., Lindquist, N. & Hentschel, U. Vertical transmission of a phylogenetically complex microbial consortium in the viviparous sponge Ircinia felix. Appl. Environ. Microbiol. 73, 2067–2078 (2007).
28. Sharp, K. H., Eam, B., Faulkner, D. J. & Haygood, M. G. Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl. Environ. Microbiol. 73, 622–629 (2007).
29. Lee, O. O., Chui, P. Y., Wong, Y. H., Pawlik, J. R. & Qian, P. Y. Evidence for vertical transmission of bacterial symbionts from adult to embryo in the Caribbean sponge Svenzea zeai. Appl. Environ. Microbiol. 75, 6147–6156 (2009).
30. Webster, N. S. et al. Deep sequencing reveals exceptional diversity and modes of transmission for bacterial sponge symbionts. Environ. Microbiol. 12, 2070–2082 (2010).The first study to apply 16S rRNA gene pyrosequencing to sponge bacteria, revealing the high degree of overlap between bacterial communities in adult and larval sponges.
31. Hentschel, U. et al. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 68, 4431–4440 (2002).The first molecular (16S rRNA gene) study to identify monophyletic sponge-specific clusters of bacteria.
32. Simister, R. L., Deines, P., Botte, E. S., Webster, N. S. & Taylor, M. W. Sponge-specific clusters revisited: a comprehensive phylogeny of sponge-associated microorganisms. Environ. Microbiol. 14, 517–524 (2012).
33. Wilkinson, C. R. Immunological evidence for the Precambrian origin of bacterial symbioses in marine sponges. Proc. R. Soc. Lond. B 220, 509–517 (1984).
34. Taylor, M. W., Thacker, R. W. & Hentschel, U. Genetics: evolutionary insights from sponges. Science 316, 1854–1855 (2007).
35. Lee, O. O. et al. Pyrosequencing reveals highly diverse and species-specific microbial communities in sponges from the Red Sea. ISME J. 5, 650–664 (2011).
36. Schmitt, S. et al. Assessing the complex sponge microbiota: core, variable and species-specific bacterial communities in marine sponges. ISME J. 6, 564–576 (2012).An investigation that uses 16S rRNA gene pyrosequencing to describe the biogeography and host specificity of bacteria in 32 sponge species from eight worldwide locations.
37. Siegl, A. et al. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 5, 61–70 (2011).The first study to apply single-cell techniques to investigate the genomes of uncultivated bacterial symbionts of sponges.
38. Fieseler, L., Horn, M., Wagner, M. & Hentschel, U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Appl. Environ. Microbiol. 70, 3724–3732 (2004).
39. Preston, C. M., Wu, K. Y., Molinski, T. F. & DeLong, E. F. A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov. Proc. Natl Acad. Sci. USA 93, 6241–6246 (1996).
40. Pape, T. et al. Dense populations of Archaea associated with the demosponge Tentorium semisuberites Schmidt, 1870 from Arctic deep-waters. Polar Biol. 29, 662–667 (2006).
41. Radax, R., Hoffmann, F., Rapp, H. T., Leininger, S. & Schleper, C. Ammonia-oxidizing archaea as main drivers of nitrification in cold-water sponges. Environ. Microbiol. 14, 909–923 (2012).
42. Wilkinson, C. R. in Algae and Symbioses: Plants, Animals, Fungi, Viruses, Interactions Explored (ed. W. Reisser) 111–151 (Biopress Limited, 1992).
43. Höller, U. et al. Fungi from marine sponges: diversity, biological activity and secondary metabolites. Mycol. Res. 104, 1354–1365 (2000).
44. Baker, P. W., Kennedy, J., Dobson, A. D. W. & Marchesi, J. R. Phylogenetic diversity and antimicrobial activities of fungi associated with Haliclona simulans isolated from Irish coastal waters. Mar. Biotechnol. 11, 540–547 (2009).
45. Quince, C. et al. Accurate determination of microbial diversity from 454 pyrosequencing data. Nature Methods 6, 639–641 (2009).
46. Kunin, V., Engelbrektson, A., Ochman, H. & Hugenholtz, P. Wrinkles in the rare biosphere: pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 12, 118–123 (2010).
47. Sunagawa, S., Woodley, C. M. & Medina, M. Threatened corals provide underexplored microbial habitats. PLoS ONE 5, e9554 (2010).
48. Cardenas, A., Rodriguez, L. M., Pizarro, V., Cadavid, L. F. & Arevalo-Ferro, C. Shifts in bacterial communities of two caribbean reef-building coral species affected by white plague disease. ISME J. 123, 502–512 (2012).
49. Behrendt, L. et al. Microbial diversity of biofilm communities in microniches associated with the didemnid ascidian Lissoclinum patella. ISME J. 6, 1222–1237 (2012).
50. Ley, R. E., Lozupone, C. A., Hamady, M., Knight, R. & Gordon, J. I. Worlds within worlds: evolution of the vertebrate gut microbiota. Nature Rev. Microbiol. 6, 776–888 (2008).
51. Dewhirst, F. E. et al. The human oral microbiome. J. Bacteriol. 192, 5002–5017 (2010).
52. Griffen, A. L. et al. Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J. 6, 1176–1185 (2012).
53. Zoetendal, E. G., Rajilic-Stojanovic, M. & de Vos, W. M. High-throughput diversity and functionality analysis of the gastrointestinal tract microbiota. Gut 57, 1605–1615 (2008).
54. Ley, R. E. et al. Evolution of mammals and their gut microbes. Science 320, 1647–1651 (2008).
55. Grice, E. A. & Segre, J. A. The skin microbiome. Nature Rev. Microbiol. 9, 244–253 (2011).
56. Taylor, M. W., Schupp, P. J., Dahllof, I., Kjelleberg, S. & Steinberg, P. D. Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity. Environ. Microbiol. 6, 121–130 (2004).
57. Wiedenbeck, J. & Cohan, F. M. Origins of bacterial diversity through horizontal genetic transfer and adaptation to new ecological niches. FEMS Microbiol. Rev. 35, 957–976 (2011).
58. Dobrindt, U., Hochhut, B., Hentschel, U. & Hacker, J. Genomic islands in pathogenic and environmental microorganisms. Nature Rev. Microbiol. 2, 414–424 (2004).
59. Sobecky, P. A. & Hazen, T. H. Horizontal gene transfer and mobile genetic elements in marine systems. Methods Mol. Biol. 532, 435–453 (2009).
60. Villareal, L. P. Viruses and the Evolution of Life (American Society for Microbiology Press, 2005).
61. Suttle, C. A. Marine viruses--major players in the global ecosystem. Nature Rev. Microbiol. 5, 801–812 (2007).
62. Claverie, J.-M. et al. Mimivirus and mimiviridae: giant viruses with an increasing number of potential hosts, including corals and sponges. J. Invertebr. Pathol. 101, 172–180 (2009).
63. Thomas, T. et al. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 4, 1557–1567 (2010).The first metagenomic analysis of sponge microbiota, revealing insights into mechanisms of bacterium–sponge interactions and highlighting ANK-containing proteins as putative symbiosis factors.
64. Grimson, A. et al. Early origins and evolution of microRNAs and Piwi-interacting RNAs in animals. Nature 455, 1193–1197 (2008).
65. Taylor, M. W., Hill, R. T. & Hentschel, U. Meeting report: first international symposium on sponge microbiology. Mar. Biotechnol. 13, 1057–1061 (2011).
66. Hallam, S. J. et al. Genomic analysis of the uncultivated marine crenarchaeote Cenarchaeum symbiosum. Proc. Natl Acad. Sci. USA 103, 18296–18301 (2006).The sequenced genome of C. symbiosum provides many new insights into the metabolic potential of this sponge symbiont.
R E V I E W S
12 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

67. Hallam, S. J. et al. Pathways of carbon assimilation and ammonia oxidation suggested by environmental genomic analyses of marine Crenarchaeota. PLoS Biol. 4, e95 (2006).
68. Liu, M. Y., Kjelleberg, S. & Thomas, T. Functional genomic analysis of an uncultured δ-proteobacterium in the sponge Cymbastela concentrica. ISME J. 5, 427–435 (2011).
69. Radax, R. et al. Metatranscriptomics of the marine sponge Geodia barretti: tackling phylogeny and function of its microbial community. Environ. Microbiol. 14, 1308–1324 (2012).
70. Lopez-Legentil, S., Erwin, P. M., Pawlik, J. R. & Song, B. Effects of sponge bleaching on ammonia-oxidizing Archaea: distribution and relative expression of ammonia monooxygenase genes associated with the barrel sponge Xestospongia muta. Microb. Ecol. 60, 561–571 (2010).
71. Turque, A. S. et al. Environmental shaping of sponge associated archaeal communities. PLoS ONE 5, e15774 (2010).
72. Steger, D. et al. Diversity and mode of transmission of ammonia-oxidizing archaea in marine sponges. Environ. Microbiol. 10, 1087–1094 (2008).
73. Bayer, K., Schmitt, S. & Hentschel, U. Physiology, phylogeny and in situ evidence for bacterial and archaeal nitrifiers in the marine sponge Aplysina aerophoba. Environ. Microbiol. 10, 2942–2955 (2008).
74. Hoffmann, F. et al. Complex nitrogen cycling in the sponge Geodia barretti. Environ. Microbiol. 11, 2228–2243 (2009).
75. Liu, M. Y., Fan, L., Zhong, L., Kjelleberg, S. & Thomas, T. Metaproteogenomic analysis of a community of sponge symbionts. ISME J. 6, 1515–1525 (2012).The first proteomic analysis of a sponge-associated microbiota, providing novel information on the activities, physiology and interactions of microorganisms with sponge hosts.
76. Gill, S. R. et al. Metagenomic analysis of the human distal gut microbiome. Science 312, 1355–1359 (2006).
77. Goodman, A. L. et al. Identifying genetic determinants needed to establish a human gut symbiont in its habitat. Cell Host Microbe 6, 279–289 (2009).
78. Cazalet, C. et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nature Genet. 36, 1165–1173 (2004).
79. Pan, X., Lührmann, A., Satoh, A., Laskowski-Arce, M. A. & Roy, C. R. Ankyrin repeat proteins comprise a diverse family of bacterial type IV effectors. Science 320, 1651–1654 (2008).
80. Voth, D. E. ThANKs for the repeat: intracellular pathogens exploit a common eukaryotic domain. Cell. Logist. 1, 128–132 (2011).
81. Schmitz-Esser, S. et al. The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” reveals common mechanisms for host cell interaction among amoeba-associated bacteria. J. Bacteriol. 192, 1045–1057 (2010).
82. Al-Khodor, S., Price, C. T., Kalia, A. & Abu Kwaik, Y. Functional diversity of ankyrin repeats in microbial proteins. Trends Microbiol. 18, 132–139 (2010).
83. Al-Khodor, S., Price, C. T., Habyarimana, F., Kalia, A. & Abu Kwaik, Y. A. Dot/Icm-translocated ankyrin protein of Legionella pneumophila is required for intracellular proliferation within human macrophages and protozoa. Mol. Microbiol. 70, 908–923 (2008).
84. Price, C. T., Al-Khodor, S., Al-Quadan, T. & Abu Kwaik, Y. Indispensable role for the eukaryotic-like ankyrin domains of the ankyrin B effector of Legionella pneumophila within macrophages and amoebae. Infect. Immun. 78, 2079–2088 (2010).
85. Lührmann, A., Nogueira, C. V., Carey, K. L. & Roy, C. R. Inhibition of pathogen-induced apoptosis by a Coxiella burnetii type IV effector protein. Proc. Natl Acad. Sci. USA 107, 18997–19001 (2010).
86. D’Andrea, L. D. & Regan, L. TPR proteins: the versatile helix. Trends Biochem. Sci. 28, 655–662 (2003).
87. Mittl, P. R. & Schneider-Brachert, W. Sel1-like repeat proteins in signal transduction. Cell. Signal. 19, 20–31 (2007).
88. Kataeva, I. A. et al. The fibronectin type 3-like repeat from the Clostridium thermocellum cellobiohydrolase CbhA promotes hydrolysis of cellulose by modifying its surface. Appl. Environ. Microbiol. 68, 4292–4300 (2002).
89. Exposito, J. Y. et al. Demosponge and sea anemone fibrillar collagen diversity reveals the early emergence
of A/C clades and the maintenance of the modular structure of type V/XI collagens from sponge to human. J. Biol. Chem. 283, 28226–28235 (2008).
90. Har-el, R. & Tanzer, M. L. Extracellular matrix. 3: evolution of the extracellular matrix in invertebrates. FASEB J. 7, 1115–1123 (1993).
91. Ozbek, S., Balasubramanian, P. G., Chiquet-Ehrismann, R., Tucker, R. P. & Adams, J. C. The evolution of extracellular matrix. Mol. Biol. Cell 21, 4300–4305 (2010).
92. Fuqua, C. & Greenberg, E. P. Listening in on bacteria: acyl-homoserine lactone signalling. Nature Rev. Mol. Cell Biol. 3, 685–695 (2002).
93. Mohamed, N. M. et al. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 10, 75–86 (2008).
94. Zan, J., Fricke, W. F., Fuqua, C., Ravel, J. & Hill, R. T. Genome sequence of Ruegeria sp. strain KLH11, an N-acylhomoserine lactone-producing bacterium isolated from the marine sponge Mycale laxissima. J. Bacteriol. 193, 5011–5012 (2011).
95. Zan, J., Fuqua, C. & Hill, R. T. Diversity and functional analysis of luxS genes in vibrios from marine sponges Mycale laxissima and Ircinia strobilina. ISME J. 5, 1505–1516 (2011).A report on quorum sensing, the presence of luxS genes and autoinducer 2 activity in sponge-associated bacterial communities.
96. Taylor, M. W. et al. Evidence for acyl homoserine lactone signal production in bacteria associated with marine sponges. Appl. Environ. Microbiol. 70, 4387–4389 (2004).
97. Hughes, D. T. & Sperandio, V. Inter-kingdom signalling: communication between bacteria and their hosts. Nature Rev. Microbiol. 6, 111–120 (2008).A summary of recently acquired evidence that bacterial quorum sensing signalling can also be used in crosstalk between microorganisms and their hosts.
98. Webster, N. S. Sponge disease: a global threat? Environ. Microbiol. 9, 1363–1375 (2007).
99. Wilson, H. V. On some phenomena of coalescence and regeneration in sponges. J. Exp. Zool. 5, 245–258 (1907).
100. Gordon, S. Pattern recognition receptors: doubling up for the innate immune response. Cell 111, 927–930 (2002).
101. Janeway, C. A. & Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 20, 197–216 (2002).
102. Liu, L. et al. Structural basis of toll-like receptor 3 signaling with double-stranded RNA. Science 320, 379–381 (2008).
103. Dunne, A. & O’Neill, L. A. The Interleukin-1 receptor/Toll-like receptor superfamily: signal transduction during inflammation and host defense. Sci. STKE 2003, re3 (2003).
104. Påisson-McDermott, E. M. & O’Neill, L. A. J. Building an immune system from nine domains. Biochem. Soc. Trans. 35, 1437–1444 (2007).
105. Bosch, T. C. G. et al. Uncovering the evolutionary history of innate immunity: the simple metazoan Hydra uses epithelial cells for host defence. Dev. Comp. Immunol. 33, 559–569 (2009).
106. Franchi, L., Eigenbrod, T., Munoz-Planillo, R. & Nunez, G. The inflammasome: a caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nature Immunol. 10, 241–247 (2009).
107. Elinav, E., Strowig, T., Henao-Mejia, J. & Flavell, R. A. Regulation of the antimicrobial response by NLR proteins. Immunity 34, 665–679 (2011).
108. Monie, T. P., C. E., B. & Gay, N. J. Activating immunity: lessons from the TLRs and NLRs. Trends Biochem. Sci. 34, 553–561 (2009).
109. Seong, S. & Matzinger, P. Hydrophobicity: an ancient damage-associated molecular pattern that initiates innate immune responses. Nature Rev. Immunol. 4, 469–478 (2004).
110. Sarrias, M. R. et al. The Scavenger Receptor Cysteine-Rich (SRCR) domain: an ancient and highly conserved protein module of the innate immune system. Crit. Rev. Immunol. 24, 1–37 (2004).
111. Plüddemann, A. et al. SR-A, MARCO and TLRs differentially recognise selected surface proteins from Neisseria meningitidis: an example of fine specificity in microbial ligand recognition by innate immune receptors. Innate Immun. 1, 153–163 (2009).
112. Pahler, S., Blumbach, B., Muller, I. & Muller, W. E. Putative multiadhesive protein from the marine
sponge Geodia cydonium: cloning of the cDNA encoding a fibronectin-, an SRCR-, and a complement control protein module. J. Exp. Zool. 282, 332–343 (1998).
113. Pancer, Z., Münker, J., Müller, I. M. & Müller, W. E. G. A novel member of an ancient superfamily: sponge (Geodia cydonium, Porifera) putative protein that features scavenger receptor cysteine-rich repeats. Gene 193, 211–218 (1997).
114. Steindler, L. et al. Differential gene expression in a marine sponge in relation to its symbiotic state. Mar. Biotechnol. 9, 543–549 (2007).
115. Huyck, T. K., Gradishar, W., Manuguid, F. & Kirkpatrick, P. Eribulin mesylate. Nature Rev. Drug Discov. 10, 173–174 (2011).
116. Aicher, T. D. et al. Total synthesis of halichondrin B and norhalichondrin B. J. Am. Chem. Soc. 114, 3162–3164 (1992).
117. Munro, M. G. H. et al. The discovery and development of marine compounds with pharmaceutical potential. J. Biotechnol. 70, 15–25 (1999).
118. Bewley, C. A. & Faulkner, D. J. Lithistid sponges: star performers or hosts to the stars. Angew. Chem. Int. Ed. Engl. 37, 2163–2178 (1998).
119. Faulkner, D. J., Unson, M. D. & Bewley, C. A. The chemistry of some sponges and their symbionts. Pure Appl. Chem. 66, 1983–1990 (1994).
120. Unson, M. D. & Faulkner, D. J. Cyanobacterial symbiont biosynthesis of chlorinated metabolites from Dysidea herbacea (Porifera). Experientia 49, 349–353 (1993).The first convincing experimental evidence that sponge-derived natural products can be of bacterial origin.
121. Unson, M. D., Holland, N. D. & Faulkner, D. J. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of the crystalline metabolite in the sponge tissue. Mar. Biol. 119, 1–11 (1994).
122. Schmidt, E. W., Obraztsova, A. Y., Davidson, S. K., Faulkner, D. J. & Haygood, M. G. Identification of the antifungal peptide-containing symbiont of the marine sponge Theonella swinhoei as a novel δ-proteobacterium, “Candidatus Entotheonella palauensis”. Mar. Biol. 136, 969–977 (2000).
123. Bewley, C. A., Holland, N. D. & Faulkner, D. J. Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia 52, 716–722 (1996).
124. Gillor, O., Carmeli, S., Rahamim, Y., Fishelson, Z. & Ilan, M. Immunolocalization of the toxin latrunculin B within the Red Sea sponge Negombata magnifica (Demospongiae, Latrunculiidae). Mar. Biotechnol. 2, 213–223 (2000).
125. Fischbach, M. A. & Walsh, C. T. Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: logic, machinery, and mechanisms. Chem. Rev. 106, 3468–3496 (2006).
126. Fisch, K. M. et al. Polyketide assembly lines of uncultivated sponge symbionts from structure-based gene targeting. Nature Chem. Biol. 5, 494–501 (2009).A report of a strategy that allows the identification of biosynthetic-gene clusters from microbial metagenomes on the basis of chemical moieties in the compounds produced by the microbial community. Using this strategy, the complete pathway for psymberin is recovered from the sponge P. bulbosa.
127. Piel, J. et al. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl Acad. Sci. USA 101, 16222–16227 (2004).An investigation that provides unambiguous evidence for the bacterial production of a potent antitumor compound from the sponge T. swinhoei.
128. Piel, J. Biosynthesis of polyketides by trans-AT polyketide synthases. Nat. Prod. Rep. 27, 996–1047 (2010).
129. Piel, J. A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proc. Natl Acad. Sci. USA 99, 14002–14007 (2002).
130. Sudek, S. et al. Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan Bugula neritina. J. Nat. Prod. 70, 67–74 (2007).
131. Partida-Martinez, L. P. & Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 437, 884–888 (2005).
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY ADVANCE ONLINE PUBLICATION | 13
© 2012 Macmillan Publishers Limited. All rights reserved

132. Partida-Martinez, L. P. & Hertweck, C. A gene cluster encoding rhizoxin biosynthesis in “Burkholderia rhizoxina”, the bacterial endosymbiont of the fungus Rhizopus microsporus. Chembiochem 8, 41–45 (2007).
133. Donia, M. S. et al. Complex microbiome underlying secondary and primary metabolism in the tunicate-Prochloron symbiosis. Proc. Natl Acad. Sci. USA 108, e1423–e1432 (2011).
134. Nguyen, T. et al. Exploiting the mosaic structure of trans-acyltransferase polyketide synthases for natural product discovery and pathway dissection. Nature Biotech. 26, 225–233 (2008).
135. Flatt, P. M. et al. Identification of the cellular site of polychlorinated peptide biosynthesis in the marine sponge Dysidea (Lamellodysidea) herbacea and symbiotic cyanobacterium Oscillatoria spongeliae by CARD-FISH analysis. Mar. Biol. 147, 761–774 (2005).
136. Lasken, R. S. Single-cell genomic sequencing using multiple displacement amplification. Curr. Opin. Microbiol. 10, 510–516 (2007).
137. Wang, D. J. & Bodovitz, S. Single cell analysis: the new frontier in ‘omics’. Trends Biotechnol. 28, 281–290 (2010).
138. Siegl, A. & Hentschel, U. PKS and NRPS gene clusters from microbial symbiont cells of marine sponges by whole genome amplification. Environ. Microbiol. Rep. 2, 507–513 (2010).
139. Grindberg, R. V. et al. Single cell genome amplification accelerates identification of the apratoxin biosynthetic pathway from a complex microbial assemblage. PLoS ONE 6, e18565 (2011).
140. Engene, N. et al. Underestimated biodiversity as a major explanation for the perceived rich secondary metabolite capacity of the cyanobacterial genus Lyngbya. Environ. Microbiol. 13, 1601–1610 (2011).
141. Intergovernmental Panel on Climate Change. Fourth Assessment Report of the Intergovernmental Panel on Climate Change (Geneva, 2007).
142. Lemoine, N., Buell, N., Hill, A. & Hill, M. in Porifera Research: Biodiversity, Innovation and Sustainability (eds M.R. Custódio, G. Lôbo-Hajdu, E. Hajdu, & G. Muricy) 419–425 (Série Livros, 2007).
143. Webster, N. S., Cobb, R. E. & Negri, A. P. Temperature thresholds for bacterial symbiosis with a sponge. ISME J. 2, 830–842 (2008).
144. Lopez-Legentil, S., Song, B., McMurray, S. E. & Pawlik, J. R. Bleaching and stress in coral reef ecosystems: hsp70 expression by the giant barrel sponge Xestospongia muta. Mol. Ecol. 17, 1840–1849 (2008).
145. Pantile, R. & Webster, N. Strict thermal threshold identified by quantitative PCR in the sponge Rhopaloeides odorabile. Mar. Ecol. Prog. Ser. 431, 97–105 (2011).
146. Luter, H. M., Whalan, S. & Webster, N. S. Exploring the role of microorganisms in the disease-like syndrome affecting the sponge Ianthella basta. Appl. Environ. Microbiol. 76, 5736–5744 (2010).
147. Angermeier, H. et al. The pathology of sponge orange band disease affecting the Caribbean barrel sponge Xestospongia muta. FEMS Microb. Ecol. 75, 218–230 (2011).
148. Webster, N. S., Negri, A. P., Webb, R. I. & Hill, R. T. A spongin-boring α-proteobacterium is the etiological agent of disease in the Great Barrier Reef sponge, Rhopaloeides odorabile. Mar. Ecol. Prog. Ser. 232, 305–309 (2002).
149. Reiswig, H. M. Particle feeding in natural populations of three marine demosponges. Biol. Bull. 141, 568–591 (1971).
150. Reiswig, H. M. Bacteria as food for temperate-water marine sponges. Can. J. Zool. 53, 582–589 (1975).
151. Pile, A. J., Patterson, M. R. & Witman, J. D. In situ grazing on plankton <10μm by the boreal sponge Mycale lingua. Mar. Ecol. Prog. Ser. 141, 95–102 (1996).
152. Hadas, E., Marie, D., Shpigel, M. & Ilan, M. Virus predation by sponges is a new nutrient-flow pathway in coral reef food webs. Limnol. Oceanogr. 51, 1548–1550 (2006).
153. Maritz, K., Calcino, A., Fahey, B., Degnan, B. & Degnan, S. M. Remarkable consistency of larval supply in the spermcast-mating demosponge Amphimedon queenslandica (Hooper and van Soest). Open Mar. Biol. J. 4, 57–64 (2010).
AcknowledgementsThe authors acknowledge the valuable contributions of all cur-rent and past members of their respective research groups. They are grateful to the participants of the First International Symposium on Sponge Microbiology (Würzburg, Germany, March 2011), for inspiring discussions and insights into many aspects of sponge microbiology. The pioneering efforts of early sponge microbiologists such as H. Reiswig, J. Vacelet and C. Wilkinson are also acknowledged. Funding for sponge and microbiology research in the authors’ laboratories was provided by the German Research Foundation (DFG) grants SFB567/TPC3 and SFB630/TPA5 (to U.H.); by the DFG, the German Federal Ministry of Education and Research (BMBF), the Japan Society for the Promotion of Sciences, the German Academic Exchange Service (DAAD), the Human Frontier Science Program, the Max Planck Society (MPG) and the German Alexander von Humboldt Foundation (to J.P.); by the Australian Research Council and the University of Queensland (to S.M.D.); and by the University of Auckland (to M.W.T.).
Competing interests statementThe authors declare no competing financial interests.
FURTHER INFORMATIONUte Hentschel’s homepage: http://www.uni-wuerzburg.de/?id=85392
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
R E V I E W S
14 | ADVANCE ONLINE PUBLICATION www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved