Embed Size (px)
Citation preview

iologicalsychiatry
Review BP
Genomic Approaches to PosttraumaticStress Disorder: The Psychiatric GenomicConsortium Initiative
Caroline M. Nievergelt, Allison E. Ashley-Koch, Shareefa Dalvie, Michael A. Hauser,Rajendra A. Morey, Alicia K. Smith, and Monica UddinISS
ABSTRACTPosttraumatic stress disorder (PTSD) after exposure to a traumatic event is a highly prevalent psychiatric disorder.Heritability estimates from twin studies as well as from recent molecular data (single nucleotide polymorphism–basedheritability) indicate moderate to high heritability, yet robust genetic variants for PTSD have not yet been identified andthe genetic architecture of this polygenic disorder remains largely unknown. To date, fewer than 10 large-scale genome-wide association studies of PTSD have been published, with findings that highlight the unique challenges for PTSDgenomics, including a complex diagnostic entity with contingency of PTSD diagnosis on trauma exposure and the largegenetic diversity of the study populations. The Psychiatric Genomics Consortium PTSD group has brought together morethan 200 scientists with the goal to increase sample size for genome-wide association studies and other genomic analysesto sufficient numbers where robust discoveries of molecular signatures can be achieved. The sample currently includesmore than 32,000 PTSD cases and 100,000 trauma-exposed control subjects, and collection is ongoing. The first resultsfound a significant shared genetic risk of PTSD with other psychiatric disorders and sex-biased heritability estimates withhigher heritability in female individuals compared with male individuals. This review describes the scope and current focusof the Psychiatric Genomics Consortium PTSD group and its expansion from the initial genome-wide association studygroup to nine working groups, including epigenetics, gene expression, imaging, and integrative systems biology. Wefurther briefly outline recent findings and future directions of “omics”-based studies of PTSD, with the ultimate goal ofelucidating the molecular architecture of this complex disorder to improve prevention and intervention strategies.
Keywords: Epigenetics, Gene expression, Genetics, Imaging, Psychiatric Genomics Consortium (PGC), Systems biology
https://doi.org/10.1016/j.biopsych.2018.01.020
GENETICS OF POSTTRAUMATIC STRESS DISORDERIN THE CONTEXT OF OTHER PSYCHIATRICDISORDERSPosttraumatic stress disorder (PTSD) is a debilitating psychi-atric disorder precipitated by traumatic experience, with sub-sequent pathological re-experiencing, avoidance, negativealterations in cognitions and mood, and hyperarousal symp-toms [DSM-5 (1)]. While most individuals exposed to traumaare resilient, PTSD prevalence is directly related to the severityand type of trauma, with rape and direct combat conferringvery high risk, and lifetime risk in women is twice that in men(2). PTSD prevalence varies by country, but lifetime prevalencein the United States is more than 7%—making it among themost common psychiatric disorders (3).
Genetic factors influence who develops PTSD; family andtwin studies have estimated heritability of PTSD from w40%to 70% following trauma (4–7). However, despite more than adecade of research on genetic candidate genes, robust ge-netic variants for PTSD have yet to be identified and thegenetic architecture of this polygenic disorder remainslargely unknown.
ª 2018 Society of BioN: 0006-3223 Bi
To address this gap in knowledge, the field of psychiatricgenomics has moved its focus over the last decade fromsmall studies on specific candidate genes (8) to agnostic,genome-wide association studies (GWASs) and ultimately towell-powered, large-scale meta-analyses made possiblethrough efforts such as the Psychiatric Genomics Con-sortium (PGC) (9). Recent success in the identification ofrobustly associated genetic variants in psychiatric disorderssuch as schizophrenia (10), bipolar disorder (11), and majordepressive disorder (MDD) (12) has confirmed that very largesample sizes are necessary to discover loci with the smallgenotypic relative risks typically seen in psychiatric disor-ders. Accordingly, the short-term goal of the PGC is toobtain GWAS data on 100,000 cases for each of its ninedisorders (9).
GENOME-WIDE ASSOCIATION APPROACHES TO PTSD
The PGC-PTSD (https://pgc-ptsd.com) was initiated in 2013by bringing together four groups with published GWASs ofPTSD (13–16), making it a relative latecomer to the PGC (17).
logical Psychiatry. Published by Elsevier Inc. All rights reserved. 831ological Psychiatry May 15, 2018; 83:831–839 www.sobp.org/journal
Genomic Approaches to PTSDBiologicalPsychiatry
To date, four additional association studies of PTSD withgenome-wide data have been published (18–21), and at leastone more study in Danish soldiers has been completed(Wang et al., Ph.D., personal communication, September 7,2017). Typically, these studies present findings that meetcriteria for genome-wide significance plus evidence ofreplication in at least one independent cohort (Table 1). Whilethis has been the gold standard for GWASs, none of theidentified genes in these GWASs has robustly replicated acrossmultiple studies. With the rapid availability of PGC-PTSD sum-mary data on a large number of studies, best practice guidelinesfor GWAS replication are currently being discussed (e.g., https://www.cohenveteransbioscience.org/2017/06/29/psychiatric-genomic-consortium-workshop-summary) and can nowinclude the prespecified selection of specific replication cohortsmatched for example on ancestry, gender, and trauma type.
The PGC-PTSD has adopted pipelines and protocols estab-lished by the PGC (https://data.broadinstitute.org/mpg/ricopili)that facilitate integration of data across disorders [e.g., (22)].However, the PGC-PTSD needs to consider some unique chal-lenges not faced by other groups, including the contingency ofPTSD diagnosis on trauma exposure, a complex and changingdiagnostic entity (23), and very diverse genetic ancestry within andacross study cohorts, resulting in considerable heterogeneity (17).
To address ancestral diversity, the PGC pipeline was extendedto include an ancestry inference module that allows for stableancestry determination across studies (https://github.com/nievergeltlab). It was designed to be portable to enable
Table 1. Genome-wide Significant Hits and Replication in Indiv
Study Hit CohortNo. PTSDCases
No.Contro
Logue, et al. (14) RORA NCPT 295 196
Replication NCPT 43 41
Replication DNHS 100 421
Guffanti, et al. (13) LINC01090 DNHS 94 319
Replication NHSII 578 1963
Xie, et al. (15) COBL GSD 300 1273
Replication GSD 207 1692
Xie, et al. (15) TLL1 GSD 300 1273
Replication GSD 207 1692
Solovieff, et al. (18) SLC18A2 NHSII 845 1693
Replication DNHS 748 748
Nievergelt, et al. (16) PRTFDC1 MRS 940 2554
Replication NCPT 313 178
Almli, et al. (84) rs717947 SBPBC 63 84
Replication GTP 2006 subjects
Replication GTP 862 subjects
Stein, et al. (20) ANKRD55 NSS 497 815
Replication MRS, PPDS, MIREC 947 4969
Stein, et al. (20) ZNF626 NSS 2140 2909
Replication PPDS 672 3335
AA, African American; DNHS, Detroit Neighborhood Health Study; EA, EuropGrady Trauma Project; GWAS, genome-wide association study; LD, linkageEducation and Clinical Center Study of Post-Deployment Mental Health; MRSStudy; NHSII, Nurses Health Study II; NS, nonsignificant; NSS, New Soldierposttraumatic stress disorder; SBPBC, Systems Biology PTSD Biomarkers Co
aTrauma: Cohorts have been separated by military and civilian; individua
832 Biological Psychiatry May 15, 2018; 83:831–839 www.sobp.org/jo
PGC-PTSD studies that cannot share individual-level genotypedata (e.g., some U.S. military and non-U.S. cohorts) to generatesummary-level results formeta-analysis. Themajority ofGWASs todate have been performed in subjects of European and AfricanAmerican ancestry groups while carefully addressing residualpopulation stratification (Figure 1). With the sustained PGC-PTSDefforts to increase sample size, other ancestries such as Latinosand East Asians are reaching considerable size. TransethnicGWASs have been generated using meta-analytical approaches(16,24), and alternative mega-analysis methods are currentlyemployed to leverage all available samples irrespective of ancestry(25).
The first PGC-PTSD publication in 2017 included 11 multi-ethnic studies of 5000 PTSD cases and 15,000 control cases(freeze 1) and is largest published genetic association study ofPTSD to date. Single nucleotide polymorphism (SNP)-basedheritability estimates for PTSD were w29% in females, butsubstantially lower in males (24), consistent with lower twin-based heritability estimates in males. In addition, the studyfound significant shared genetic risk of PTSD with schizophrenia,bipolar disorder, and MDD. Investigating the implications of sex-based heritability in PTSD and cross-disorder genetic risks is ahigh priority for future study design.
EXPANDING THE PGC-PTSD SCOPE
Although successful in demonstrating some aspects of the ge-netic architecture of PTSD, the PGC-PTSD freeze 1 dataset was
idual PTSD Studies
ls Ancestry Sex Traumaa p Value Comment
EA Pred. M Pred. military 2.50E-08
AA Pred. M Pred. military 0.05 Gene-base test
Pred. AA F Civilian NS Gene-base test
Pred. AA F Civilian 5.09E-08
EA F Civilian 0.07 Marginal
EA M1F Civilian 3.96E-08
EA M1F Civilian NS
EA M1F Civilian 2.99E-07 Marginal GWAS hit
EA M1F Civilian 6.30E-06
EA F Civilian 2.10E-05 Candidate-gene study
Pred. AA M1F Civilian 0.05 Haplotype
Mixed M Military 2.04E-09
EA Pred. M Military 0.14 Marginal forSNP in LD
Mixed Pred. M Military 1.28E-08
AA F Civilian 0.005 PTSD symptoms
AA M Civilian NS PTSD symptoms
AA Pred. M Military 2.34E-08
AA Pred. M Military NS Meta-analysis
EA Pred. M Military 4.59E-08
EA Pred. M Military NS
ean ancestry; F, female; GSD, Genetics of Substance Dependence; GTP,disequilibrium; M, male; MIREC, Mid-Atlantic Mental Illness Research,, Marine Resiliency Study; NCTP, VA Boston-National Center for PTSDStudy; PPDS, Pre/Post Deployment Study; Pred., predominantly; PTSD,nsortium; SNP, single nucleotide polymorphism.l type of trauma is not considered here.
urnal
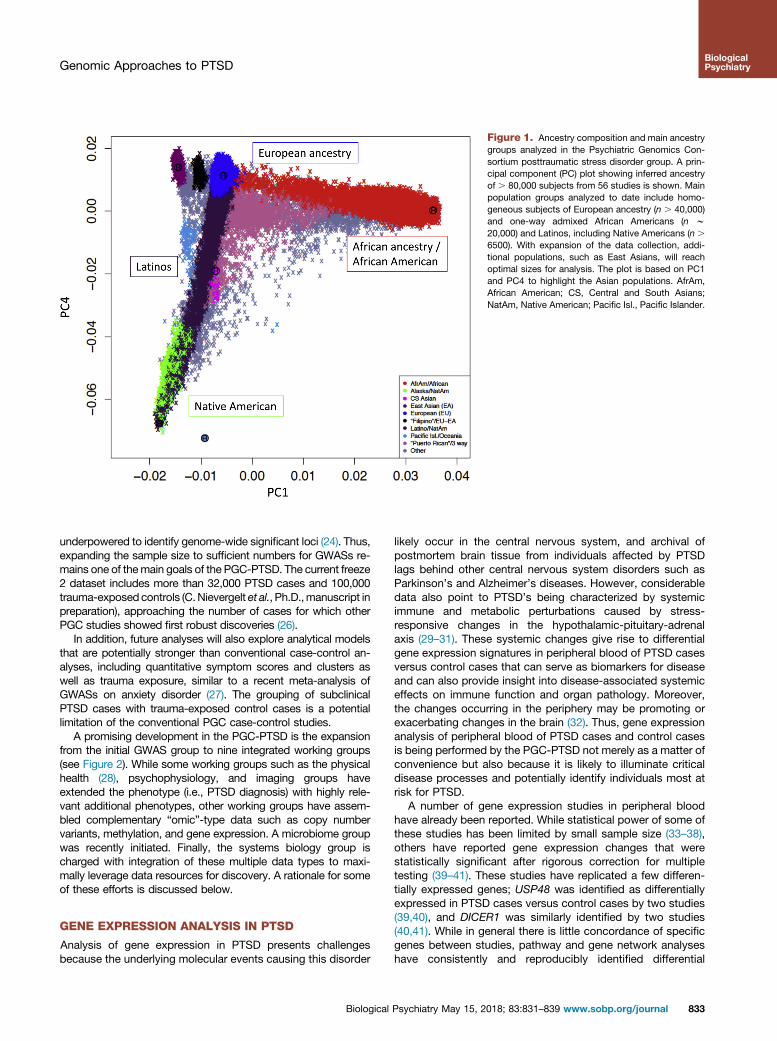
Figure 1. Ancestry composition and main ancestrygroups analyzed in the Psychiatric Genomics Con-sortium posttraumatic stress disorder group. A prin-cipal component (PC) plot showing inferred ancestryof . 80,000 subjects from 56 studies is shown. Mainpopulation groups analyzed to date include homo-geneous subjects of European ancestry (n . 40,000)and one-way admixed African Americans (n w20,000) and Latinos, including Native Americans (n .
6500). With expansion of the data collection, addi-tional populations, such as East Asians, will reachoptimal sizes for analysis. The plot is based on PC1and PC4 to highlight the Asian populations. AfrAm,African American; CS, Central and South Asians;NatAm, Native American; Pacific Isl., Pacific Islander.
Genomic Approaches to PTSDBiologicalPsychiatry
underpowered to identify genome-wide significant loci (24). Thus,expanding the sample size to sufficient numbers for GWASs re-mains one of themain goals of the PGC-PTSD. The current freeze2 dataset includes more than 32,000 PTSD cases and 100,000trauma-exposed controls (C.Nievergelt et al., Ph.D.,manuscript inpreparation), approaching the number of cases for which otherPGC studies showed first robust discoveries (26).
In addition, future analyses will also explore analytical modelsthat are potentially stronger than conventional case-control an-alyses, including quantitative symptom scores and clusters aswell as trauma exposure, similar to a recent meta-analysis ofGWASs on anxiety disorder (27). The grouping of subclinicalPTSD cases with trauma-exposed control cases is a potentiallimitation of the conventional PGC case-control studies.
A promising development in the PGC-PTSD is the expansionfrom the initial GWAS group to nine integrated working groups(see Figure 2). While some working groups such as the physicalhealth (28), psychophysiology, and imaging groups haveextended the phenotype (i.e., PTSD diagnosis) with highly rele-vant additional phenotypes, other working groups have assem-bled complementary “omic”-type data such as copy numbervariants, methylation, and gene expression. A microbiome groupwas recently initiated. Finally, the systems biology group ischarged with integration of these multiple data types to maxi-mally leverage data resources for discovery. A rationale for someof these efforts is discussed below.
GENE EXPRESSION ANALYSIS IN PTSD
Analysis of gene expression in PTSD presents challengesbecause the underlying molecular events causing this disorder
Biological
likely occur in the central nervous system, and archival ofpostmortem brain tissue from individuals affected by PTSDlags behind other central nervous system disorders such asParkinson’s and Alzheimer’s diseases. However, considerabledata also point to PTSD’s being characterized by systemicimmune and metabolic perturbations caused by stress-responsive changes in the hypothalamic-pituitary-adrenalaxis (29–31). These systemic changes give rise to differentialgene expression signatures in peripheral blood of PTSD casesversus control cases that can serve as biomarkers for diseaseand can also provide insight into disease-associated systemiceffects on immune function and organ pathology. Moreover,the changes occurring in the periphery may be promoting orexacerbating changes in the brain (32). Thus, gene expressionanalysis of peripheral blood of PTSD cases and control casesis being performed by the PGC-PTSD not merely as a matter ofconvenience but also because it is likely to illuminate criticaldisease processes and potentially identify individuals most atrisk for PTSD.
A number of gene expression studies in peripheral bloodhave already been reported. While statistical power of some ofthese studies has been limited by small sample size (33–38),others have reported gene expression changes that werestatistically significant after rigorous correction for multipletesting (39–41). These studies have replicated a few differen-tially expressed genes; USP48 was identified as differentiallyexpressed in PTSD cases versus control cases by two studies(39,40), and DICER1 was similarly identified by two studies(40,41). While in general there is little concordance of specificgenes between studies, pathway and gene network analyseshave consistently and reproducibly identified differential
Psychiatry May 15, 2018; 83:831–839 www.sobp.org/journal 833

Figure 2. Main interactions and dataflow among thenine Psychiatric Genomics Consortium posttraumaticstress disorder (PTSD) working groups. The PsychiatricGenomics Consortium PTSD genome-wide associationstudy (GWAS) group has drastically expanded its scopesince its initiation in 2013. It currently includes workinggroups with emphases on complementary phenotypes(psychophysiology, physical health, and imaging),working groups contributing complementary “omics”data (copy number variants [CNV], epigenetics, tran-scriptome, and microbiome), and a systems biologygroup aiming at integration of the different types ofdata. Arrows indicate primary flow of data, but in-teractions among groups are expanding. EWAS,epigenome-wide association study; MRI, magneticresonance imaging; RNAseq, RNA sequencing; SNP,single nucleotide polymorphism.
Genomic Approaches to PTSDBiologicalPsychiatry
expression of transcripts involved in innate immunity, inter-feron signaling, and wound healing (42,43). These studies haveprovided valuable insights into the pathobiology of PTSD. Witha combined sample size of nearly 5000 samples, future pe-ripheral blood gene expression studies by the PGC-PTSDshould refine and extend these findings. As central nervoussystem tissue samples become increasingly available, geneexpression data from postmortem brains can be comparedand integrated with findings from peripheral blood.
PTSD EPIGENETICS
PTSD is unique among psychiatric disorders in that it requires atraumatic environmental event as part of its diagnosis. Amongthe different epigenetic modifications, DNA methylation hasreceived the most attention by researchers studying psycho-social stress, childhood trauma, and PTSD due to its relativestability and its ability to be assessed with microarrays thatfacilitate replication within and between studies (40,44–50). Im-mune dysregulation figured prominently among the biologicalpathways that are associated with PTSD and are replicablebetween studies (44,46). A recent study examined DNAmethylation along with microRNA, another epigenetic modifi-cation, in a small group of PTSD cases and control cases. Theauthors noted reductions in microRNA levels in PTSD cases andproposed that epigenetic changes may contribute to systemicinflammation in PTSD (51). These studies echo those of otherpsychiatric disorders that emphasize the cross-talk between theperipheral immune system and the brain (32,52).
Other studies suggest more widespread mechanisms forepigenetic dysregulation in PTSD. For example, Maddox et al.reported DNA methylation differences in HDAC4, a histonedeacetylase, in the blood of women with PTSD and went on toshow that variation in genetic and epigenetic predictors ofHDAC4 expression was associated with fear-potentiatedstartle response and functional connectivity differences in theamygdala (53). Similarly, lower expression of DICER1, which isrequired for processing mature microRNAs, is associated withPTSD cases with comorbid depression and increased amyg-dala activation in response to fearful stimuli (41), a neuralphenotype strongly associated with risk for PTSD even prior totrauma exposure [reviewed in (54)].
834 Biological Psychiatry May 15, 2018; 83:831–839 www.sobp.org/jo
In addition to these genome-scale approaches, epigeneticsummary measures may be particularly informative. The mostwidely used measure is the epigenetic clock (55,56) for assessingage acceleration, which is associated with psychosocial stressand higher mortality risk (56,57). The phenomenon describesmethylation-based prediction of age that exceeds chronologicalage. Of note, a relatively high proportion—nearly 25%—ofepigenetic clock-related CpG sites are located in glucocorticoidresponse elements, a genomic region in which methylation levelsvary in relation to trauma exposure (58) and dexamethasonesuppression (56). These environmentally sensitive genomic siteshave been explicitly linked to traumatic stress, neural integrity, andmortality (59), discussed in more detail below, further demon-strating the utility of using epigenetic summary measures as anindex of the biological impact of lived experience, including traumaexposure.
There are numerous challenges in conducting epigeneticstudies of PTSD. Similar to gene expression discussed above,DNA methylation patterns vary by tissue type, with the majorityof studies to date being conducted in blood, whereas the mostrelevant tissue is brain. As new postmortem samples andsingle-cell technologies become available, there will be sub-stantial advancement in identification of genes whose regula-tion is altered in those with PTSD. This may further support theidentification of peripheral biomarkers or may restrict the scopeof peripheral tissues. A second challenge is limited platforms,and thereby limited coverage, for DNA methylation or otherarrays that are widely used for population-scale studies.Epigenome-wide investigations require cost-effective andhighly reproducible methods to achieve the sample sizesrequired to detect associations that withstand multiple testingcorrection. The PGC-PTSD epigenome-wide association study(EWAS) group (Figure 2) has the goal to assemble such data toperform meta-analyses across cohorts with a common multisiteanalysis pipeline (60). In some ways, these platform limitationsincrease opportunities for replication but also complicate link-ing genome-wide discoveries with those based on sequencingor targeted assays.
PTSD IMAGING GENETICS
Elevated risk of psychopathology may be more powerfullyinvestigated with intermediate phenotypes (or endophenotypes)
urnal

Genomic Approaches to PTSDBiologicalPsychiatry
than clinical diagnoses. Brainmeasures frommagnetic resonanceimaging may have a simpler underlying genetic architectureinvolving fewer individual genes or pathways than the polygenicitydriving overall risk for psychopathology (61) and offer a moreprecise and reproducible phenotype than clinical diagnosticscales (62). A GWAS of continuous brain measures may be sta-tistically more powerful and more efficient than binary traits(diagnosis), whichmay disguise complexities such as comorbidityand syndromal heterogeneity (63). Furthermore, brain phenotypesmay provide common pathways for the combined effects ofenvironmental and genetic risk factors that may underlie multiplediagnoses (61). However, there are two important caveats, namelythat 1) the effect sizes for gene effects on neuroimaging pheno-types are unlikely to be greater than those for behavioral traits orpsychiatric disorders (64) and 2) genetics of brain phenotypesmayreveal commonmechanistic pathways for a number of psychiatricdisorders, resulting in a loss of specificity when moving frompsychiatric disorder to brain phenotype—different disorders maypossess nearly identical brain abnormalities (65). This loss ofspecificity may prove to be advantageous for drug developmentby facilitating the design of a target-specific intervention that iseffective for multiple neuropsychiatric disorders.
An international collaboration of investigators (17) within thePGC and Enhancing NeuroImaging Genetics through Meta-Analysis (ENIGMA) plans to investigate the genetic effects ofcomplex brain traits (65). The first major analysis of the PGC-ENIGMA PTSD working group with 1868 samples hasdemonstrated that PTSD is associated with smaller hippo-campus and amygdala volume (66). Exposure to childhoodtrauma was negatively associated with hippocampal andamygdala volume when adjusting for age, sex, and intracranialvolume (66). Both structures have ample a priori evidenceimplicating their role in PTSD, starting with the report ofreduced hippocampal volume in a small PTSD sample morethan 20 years ago (67). However, we confirmed this findingacross a large number of demographically and clinically het-erogeneous cohorts analyzed with a standardized segmenta-tion technique and a harmonized analysis protocol across allsites. Methodological consistency was promoted by using thesame statistical models across all samples, making this thelargest and most powerful study of subcortical volumes inPTSD to date. The analyses of 12 hippocampal subfield vol-umes, diffusion tensor imaging measures of 26 white mattertracts, and the cortical thickness of 78 regions of interest, aswell as whole-brain vertex-based analyses, are currently underway using ENIGMA pipelines, which can be downloaded athttps://pgc-ptsd.com/methods-tools/imaging-pipeline.
Forthcoming analyses of gene-by-environment GWASs ofrelevant structural brain phenotypes with childhood trauma asa major risk factor are planned with the long-term goal ofidentifying genetic modulators of brain structure that help earlyprediction and treatment for a range of psychiatric disorders,followed by deep sequencing in a subset of samples to identifypotential causal variants within the coding and/or regulatoryregions of implicated risk loci. More than 40 participating siteshave coalesced around the common goal to form the ENIGMA-PGC-PTSD, which has already received 4000 samples, amongwhich 3000 samples have been aggregated and analyzed.Nevertheless, the analyses are expected to be woefully un-derpowered with the large number of phenotypes available in
Biological
neuroimaging data. Two approaches address the shortcom-ings of previous neuroimaging-genetics studies that have beenplagued by small sample size due to the large expense ofmagnetic resonance imaging acquisition and the use ofcandidate genes that have been criticized for being susceptibleto population stratification and fueling information bottlenecks.First, no candidate gene analyses are planned. Second, repli-cation samples of neuroimaging data such as the U.K. Biobankand the Million Veteran Program will be leveraged. Third, wewill focus on polygenic risk score (PRS) calculation and PRS 3
environment interaction analyses. Discovery samples forcalculating PRS include 1) the GWAS of the PTSD diagnosisfrom the PGC from 80,000 samples, 2) the GWAS of subcor-tical volumetry performed from 31,000 normative neuro-imaging and genomic samples that provided several SNPassociations (68), and 3) the GWAS of cortical thickness andsurface area in which preliminary results from 30,000 sampleshave generated 120 SNPs that show robust genome-widesignificant associations after correction for 78 cortical struc-tures. The testing of PRSs that are calculated in discoverysamples, which have yielded robust genome-wide associa-tions, in our sample avoids the criticisms previously leveledagainst neuroimaging studies (69).
ADDRESSING THE INACCESSIBILITY OF BRAINTISSUE
To date, EWASs of PTSD have been limited to accessibleperipheral tissues, specifically whole blood (40,44,46). Whilepotentially informative as biomarkers of the disorder, the extentto which PTSD-associated DNA methylation patterns in bloodor other peripheral tissues reflect patterns that may exist in thebrain remains unknown. As the target organ of most interest tothe disorder, the brain remains a challenge to access in livingindividuals, and brain-based epigenetic predictors of PTSDhave yet to be identified.
Despite these challenges, recent work has attempted tobridge the link between brain and periphery by examiningperipherally derived epigenetic biomarkers of neuroimaging-based phenotypes and endophenotypes of PTSD. Much ofthis work to date has adopted a candidate gene approach[e.g., (70,71)]. A notable exception is a study conducted byWolf et al. (59), which tested the hypothesis that PTSD isassociated with accelerated cellular age and degraded neuralintegrity, as well as reduced performance on executive func-tion tasks, in a sample of U.S. veterans. Lifetime PTSD severitywas found to be positively associated with DNA methylation-derived age, and these age estimates were negatively asso-ciated with neural integrity in the genu of the corpus callosumand working memory performance. Of note, the mean age in thissample was w32 years, suggesting that the neurobiological ef-fects of traumatic stress may impair neuropsychological func-tioning of veteran populations. This genome-scale studyrepresents a genomic approach to PTSD with high public healthimpact because it holds the potential to identify individuals whomay be most in need of intervention by leveraging peripherallyderived, polygenic epigenetic measurements that are predictiveof neural integrity and memory performance. Cohorts within thePGC-PTSD that include both neuroimaging and EWAS data areoptimally positioned to combine efforts and pursue similar
Psychiatry May 15, 2018; 83:831–839 www.sobp.org/journal 835

Genomic Approaches to PTSDBiologicalPsychiatry
studies in the future, augmented by the meta-analytic frameworkcurrently being employed by both the EWAS and neuroimagingworking groups within the PGC-PTSD.
DATA INTEGRATION AND SYSTEMS BIOLOGYAPPROACHES
Based on current findings, the underlying etiology of PTSD islikely the result of a complex interplay between various mo-lecular systems (72), and to delineate this, a holistic (systemsbiology) approach that integrates different omics layers amongPTSD cases and trauma-exposed control cases may be thenext logical step. One of the strengths of the PGC-PTSD is thatmultiple forms of genomic data have been generated for manydatasets, which will allow cross-platform analyses to be per-formed (Figure 2). Several methods have been proposed formeta-analyses across platforms (73–76), as well as forimputing data to improve the ability to combine datasets(77–81). Such joint analyses of multiple omics datasets havebeen previously termed “genomic convergence” and havegreat potential to inform the genetic architecture of PTSD (82).
Through the coordination of the various PGC-PTSD workinggroups (Figure 2), there is potential to identify DNA methylationpatterns that may be giving rise to altered gene expression andsufficient power to conduct expression quantitative trait locus(QTL) and methylation QTL analyses, which assess whetherparticular genetic variants alter the levels of expression andmethylation of specific genes, respectively. For example, pre-vious studies for PTSD have shown that the risk variantrs363276, located within an intronic region of SLC18A2, is anexpression QTL significantly associated with decreasedexpression of the genes SLC18A2 and PDZD8 in the dorso-lateral prefrontal cortex of postmortem human brains (83). ThePTSD risk variant rs717947, located at chromosome 4p15,was shown to be a methylation QTL (84). These types ofanalysis provide clues on the etiology of PTSD; however,identifying definitive causal risk factors requires alternativemethods such as Mendelian randomization, which quantifiescausality between a risk factor and a disease outcome byusing SNP data as an instrumental variable (85). This techniquehas been applied to PTSD, whereby a causal relationship wasidentified between plasma dopamine beta-hydroxylase, anenzyme that catalyzes the synthesis of norepinephrine, andsymptoms of reexperiencing (86). Another study investigatedthe relationship between body mass index–adjusted weightcircumference and PTSD in women and found that an increasein body mass index–adjusted weight circumference results in arelative decrease in the risk of developing PTSD (87).
Current data integration efforts will be augmented throughthe increasing availability of publicly available data sets, suchas GTEx (88) and PsychENCODE (89), which can provideadditional annotation for the PTSD-specific findings. Forexample, recently developed methods such as PrediXcan usegenome-wide variation to predict or impute gene expression intest datasets based on tissue-dependent modeling performedon transcriptome data from reference databases such asGTEx. The imputed expression can be tested for association tothe phenotype of interest, enabling the identification of trait-associated loci (90). In addition, as technologies continue toevolve and become more widely available for single-cell RNA
836 Biological Psychiatry May 15, 2018; 83:831–839 www.sobp.org/jo
sequencing (91,92), induced pluripotent stem cell–derivedneural progenitor cells (93), and brain organoids (94–96),these technologies can also be integrated into the ongoingPGC-PTSD efforts, particularly with respect to understandingthe roles of specific genes and variants in PTSD risk.
Studies of this nature fill a significant gap in the availableliterature on the complex genetic mechanisms and pathwaysunderlying complex psychiatric disorders, including PTSD.Systems biology approaches will lay the groundwork for futuredevelopment of more accurate diagnostic methods, improvedmanagement, and the development of more suitable andindividualized treatment strategies for patients.
CONCLUSIONS
The promise of finding genetic determinants of psychiatric dis-orders is identifying etiologic pathways for targeted interventions.However, before this can become a reality, biological validationof genetic findings will be required. Making individual predictionsto support the emerging discipline of precision medicine holdsthe promise of personalized medical decisions driven by an in-dividual’s genetic makeup and environment, other risk factors,and large databases of genotype–phenotype relationships.
ACKNOWLEDGMENTS AND DISCLOSURESFinancial support was provided by the Stanley Center for Psychiatric Ge-netics at the Broad Institute, One Mind, Cohen Veterans Bioscience, and theNational Institute of Mental Health (NIMH) Grant Nos. R01-MH111671-01 (toRAM, AEA-K, and MAH) and by NIMH/U.S. Army Medical Research andMateriel Command Grant No. R01MH106595 (to CMN) and Grant No.R01MH108826 (to AKS, MU, and CMN). AEA-K and MAH were supportedby the Department of Veterans Affairs’ Mid-Atlantic Mental Illness Research,Education, and Clinical Center and the Research & Development and MentalHealth Services of the Durham Veterans Affairs Medical Center.
We thank the research groups contributing to the PGC-PTSD work-ing groups for sharing their data and time to make this work possible andespecially the more than 80,000 research participants worldwide whoshared personal experiences and biological samples with theseresearch studies.
The authors report no biomedical financial interests or potential conflictsof interest.
ARTICLE INFORMATIONFrom the Department of Psychiatry (CMN) and Department of FamilyMedicine and Public Health (CMN), University of California, San Diego, andCenter of Excellence for Stress and Mental Health (CMN), Veterans AffairsSan Diego Healthcare System, San Diego, California; Department of Medi-cine (AEA-K, MAH), Duke University Medical Center, Department of Psy-chiatry and Behavioral Sciences (RAM), Duke University School of Medicine,and Durham Veterans Affairs Medical Center (RAM), Durham, North Car-olina; Department of Gynecology and Obstetrics (AKS) and Department ofPsychiatry & Behavioral Sciences (AKS), Emory University, Atlanta, Georgia;Carl R. Woese Institute for Genomic Biology (MU) and Department of Psy-chology (MU), University of Illinois–Urbana-Champaign, Champaign, Illinois;and Department of Psychiatry and Mental Health (SD), University of CapeTown, Cape Town, South Africa.
Address correspondence to Caroline Nievergelt, Ph.D., Department ofPsychiatry, University of California, San Diego, 9500 Gilman Drive, La Jolla,CA 92093; E-mail: [email protected].
Received Aug 31, 2017; revised Dec 18, 2017; accepted Jan 18, 2018.
REFERENCES1. American Psychiatric Association (2013): Trauma- and Stressor-
Related Disorders: Diagnostic and Statistical Manual of Mental
urnal

Genomic Approaches to PTSDBiologicalPsychiatry
Disorders, 5th ed. Washington, DC: American PsychiatricAssociation.
2. Yehuda R, Hoge CW, McFarlane AC, Vermetten E, Lanius RA,Nievergelt CM, et al. (2015): Post-traumatic stress disorder. Nat RevDis Primers 1:15057.
3. Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB (1995):Posttraumatic stress disorder in the National Comorbidity Survey.Arch Gen Psychiatry 52:1048–1060.
4. True WJ, Rice J, Eisen SA, Heath AC, Goldberg J, Lyons MJ, et al.(1993): A twin study of genetic and environmental contributions to li-ability for posttraumatic stress symptoms. Arch Gen Psychiatry50:257–264.
5. Stein MB, Jang KJ, Taylor S, Vernon PA, Livesley WJ (2002): Geneticand environmental influences on trauma exposure and posttraumaticstress disorder: A twin study. Am J Psychiatry 159:1675–1681.
6. Sartor CE, Grant JD, Lynskey MT, McCutcheon VV, Waldron M,Statham DJ, et al. (2012): Common heritable contributions to low-risktrauma, high-risk trauma, posttraumatic stress disorder, and majordepression. Arch Gen Psychiatry 69:293–299.
7. Sartor CE, McCutcheon VV, Pommer NE, Nelson EC, Grant JD,Duncan AE, et al. (2011): Common genetic and environmental contri-butions to post-traumatic stress disorder and alcohol dependence inyoung women. Psychol Med 41:1497–1505.
8. Ioannidis JP, Tarone R, McLaughlin JK (2011): The false-positive tofalse-negative ratio in epidemiologic studies. Epidemiology 22:450–456.
9. Sullivan PF, Agrawal A, Bulik CM, Andreassen OA, Borglum AD,Breen G, et al. (2018): Psychiatric genomics: An update and anagenda. Am J Psychiatry 175:15–27.
10. Schizophrenia Working Group of the Psychiatric Genomics Con-sortium (2014): Biological insights from 108 schizophrenia-associatedgenetic loci. Nature 511:421–427.
11. Sklar P, Ripke S, Scott LJ, Andreassen OA, Cichon S, Craddock N,et al. (2011): Large-scale genome-wide association analysis of bipolardisorder identifies a new susceptibility locus near ODZ4. Nat Genet43:977–983.
12. Hyde CL, Nagle MW, Tian C, Chen X, Paciga SA, Wendland JR, et al.(2016): Identification of 15 genetic loci associated with risk of majordepression in individuals of European descent. Nat Genet 48:1031–1036.
13. Guffanti G, Galea S, Yan L, Roberts AL, Solovieff N, Aiello AE, et al.(2013): Genome-wide association study implicates a novel RNA gene, thelincRNA AC068718.1, as a risk factor for post-traumatic stress disorder inwomen. Psychoneuroendocrinology 38:3029–3038.
14. Logue MW, Baldwin C, Guffanti G, Melista E, Wolf EJ, Reardon AF,et al. (2013): A genome-wide association study of post-traumaticstress disorder identifies the retinoid-related orphan receptoralpha (RORA) gene as a significant risk locus. Mol Psychiatry18:937–942.
15. Xie P, Kranzler HR, Yang C, Zhao H, Farrer LA, Gelernter J (2013):Genome-wide association study identifies new susceptibility loci forposttraumatic stress disorder. Biol Psychiatry 74:656–663.
16. Nievergelt CM, Maihofer AX, Mustapic M, Yurgil KA, Schork NJ,Miller MW, et al. (2015): Genomic predictors of combat stressvulnerability and resilience in U.S. Marines: A genome-wide associa-tion study across multiple ancestries implicates PRTFDC1 as a po-tential PTSD gene. Psychoneuroendocrinology 51:459–471.
17. Logue MW, Amstadter AB, Baker DG, Duncan L, Koenen KC,Liberzon I, et al. (2015): The Psychiatric Genomics Consortium Post-traumatic Stress Disorder workgroup: Posttraumatic stress disorderenters the age of large-scale genomic collaboration. Neuro-psychopharmacology 40:2287–2297.
18. Solovieff N, Roberts AL, Ratanatharathorn A, Haloosim M, De Vivo I,King AP, et al. (2014): Genetic association analysis of 300 genesidentifies a risk haplotype in SLC18A2 for post-traumatic stress dis-order in two independent samples. Neuropsychopharmacology39:1872–1879.
19. Almli LM, Srivastava A, Fani N, Kerley K, Mercer KB, Feng H, et al.(2014): Follow-up and extension of a prior genome-wide associationstudy of posttraumatic stress disorder: Gene 3 Environment
Biological
associations and structural magnetic resonance imaging in a highlytraumatized African-American civilian population. Biol Psychiatry76:e3–e4.
20. Stein MB, Chen CY, Ursano RJ, Cai T, Gelernter J, Heeringa SG, et al.(2016): Genome-wide association studies of posttraumatic stress disor-der in 2 cohorts of US Army soldiers. JAMA Psychiatry 73:695–704.
21. Ashley-Koch AE, Garrett ME, Gibson J, Liu Y, Dennis MF, Kimbrel NA,et al. (2015): Genome-wide association study of posttraumatic stressdisorder in a cohort of Iraq–Afghanistan era veterans. J Affect Disord184:225–234.
22. Cross-Disorder Group of the Psychiatric Genomics Consortium (2013):Identification of risk loci with shared effects on five major psychiatricdisorders: A genome-wide analysis. Lancet 381:1371–1379.
23. Armour C, Mullerova J, Elhai JD (2016): A systematic literature reviewof PTSD’s latent structure in the Diagnostic and Statistical Manual ofMental Disorders: DSM-IV to DSM-5. Clin Psychol Rev 44:60–74.
24. Duncan LE, Ratanatharathorn A, Aiello AE, Almli LM, Amstadter AB,Ashley-Koch AE, et al. (2018): Largest GWAS of PTSD (N = 20 070)yields genetic overlap with schizophrenia and sex differences in heri-tability. Mol Psychiatry 23:666–673.
25. Zhou X, Stephens M (2014): Efficient multivariate linear mixed modelalgorithms for genome-wide association studies. Nat Methods11:407–409.
26. Gratten J, Wray NR, Keller MC, Visscher PM (2014): Large-scale ge-nomics unveils the genetic architecture of psychiatric disorders. NatNeurosci 17:782–790.
27. Otowa T, Hek K, Lee M, Byrne EM, Mirza SS, Nivard MG, et al. (2016):Meta-analysis of genome-wide association studies of anxiety disor-ders. Mol Psychiatry 21:1391–1399.
28. Sumner JA, Duncan LE, Wolf EJ, Amstadter AB, Baker DG,Beckham JC, et al. (2017): Letter to the Editor: Posttraumatic stressdisorder has genetic overlap with cardiometabolic traits. Psychol Med47:2036–2039.
29. Michopoulos V, Vester A, Neigh G (2016): Posttraumatic stressdisorder: A metabolic disorder in disguise? Exp Neurol 284:220–229.
30. Passos IC, Vasconcelos-Moreno MP, Costa LG, Kunz M, Brietzke E,Quevedo J, et al. (2015): Inflammatory markers in post-traumaticstress disorder: A systematic review, meta-analysis, and meta-regression. Lancet Psychiatry 2:1002–1012.
31. Mehta D, Binder EB (2012): Gene 3 Environment vulnerability factorsfor PTSD: The HPA-axis. Neuropharmacology 62:654–662.
32. Andrews JA, Neises KD (2012): Cells, biomarkers, and post-traumaticstress disorder: Evidence for peripheral involvement in a central dis-ease. J Neurochem 120:26–36.
33. Zieker J, Zieker D, Jatzko A, Dietzsch J, Nieselt K, Schmitt A, et al.(2007): Differential gene expression in peripheral blood of patientssuffering from post-traumatic stress disorder. Mol Psychiatry 12:116–118.
34. Segman R, Shefi N, Goltser-Dubner T, Friedman N, Kaminski N,Shalev A (2005): Peripheral blood mononuclear cell gene expressionprofiles identify emergent post-traumatic stress disorder amongtrauma survivors. Mol Psychiatry 10:500–513.
35. Sarapas C, Cai G, Bierer LM, Golier JA, Galea S, Ising M, et al. (2011):Genetic markers for PTSD risk and resilience among survivors of theWorld Trade Center attacks. Dis Markers 30:101–110.
36. Yehuda R, Cai G, Golier JA, Sarapas C, Galea S, Ising M, et al. (2009):Gene expression patterns associated with posttraumatic stress dis-order following exposure to the World Trade Center attacks. BiolPsychiatry 66:708–711.
37. O’Donovan A, Sun B, Cole S, Rempel H, Lenoci M, Pulliam L, et al.(2011): Transcriptional control of monocyte gene expression in post-traumatic stress disorder. Dis Markers 30:123–132.
38. Neylan TC, Sun B, Rempel H, Ross J, Lenoci M, O’Donovan A, et al.(2011): Suppressed monocyte gene expression profile in men versuswomen with PTSD. Brain Behav Immun 25:524–531.
39. Logue MW, Smith AK, Baldwin C, Wolf EJ, Guffanti G,Ratanatharathorn A, et al. (2015): An analysis of gene expression inPTSD implicates genes involved in the glucocorticoid receptor
Psychiatry May 15, 2018; 83:831–839 www.sobp.org/journal 837

Genomic Approaches to PTSDBiologicalPsychiatry
pathway and neural responses to stress. Psychoneuroendocrinology57:1–13.
40. Mehta D, Klengel T, Conneely KN, Smith AK, Altmann A, Pace TW,et al. (2013): Childhood maltreatment is associated with distinctgenomic and epigenetic profiles in posttraumatic stress disorder. ProcNatl Acad Sci U S A 110:8302–8307.
41. Wingo AP, Almli LM, Stevens JJ, Klengel T, Uddin M, Li Y, et al. (2015):DICER1 and microRNA regulation in post-traumatic stress disorderwith comorbid depression. Nat Commun 6:10106. [(2016): Corri-gendum, Nat Commun 7:10958.]
42. Breen MS, Maihofer AX, Glatt SJ, Tylee DS, Chandler SD, Tsuang MT,et al. (2015): Gene networks specific for innate immunity define post-traumatic stress disorder. Mol Psychiatry 20:1538–1545.
43. Breen MS, Tylee DS, Maihofer AX, Neylan TC, Mehta D, Binder E, et al.(2018): PTSD blood transcriptome mega-analysis: Shared inflamma-tory pathways across biological sex and modes of trauma. Neuro-psychopharmacology 43:469–481.
44. Smith AK, Conneely KN, Kilaru V, Mercer KB, Weiss TE, Bradley B,et al. (2011): Differential immune system DNA methylation and cyto-kine regulation in post-traumatic stress disorder. Am J Med Genet BNeuropsychiatr Genet 156:700–708.
45. Ressler KJ, Mercer KB, Bradley B, Jovanovic T, Mahan A, Kerley K,et al. (2011): Post-traumatic stress disorder is associated with PACAPand the PAC1 receptor. Nature 470:492–497.
46. Uddin M, Aiello AE, Wildman DE, Koenen KC, Pawelec G, de LosSantos R, et al. (2010): Epigenetic and immune function profilesassociated with posttraumatic stress disorder. Proc Natl Acad SciU S A 107:9470–9475.
47. Mehta D, Bruenig D, Carrillo-Roa T, Lawford B, Harvey W, Morris CP,et al. (2017): Genomewide DNA methylation analysis in combatveterans reveals a novel locus for PTSD. Acta Psychiatr Scand 136:493–505.
48. Hammamieh R, Chakraborty N, Gautam A, Muhie S, Yang R,Donohue D, et al. (2017): Whole-genome DNA methylation statusassociated with clinical PTSD measures of OIF/OEF veterans. TranslPsychiatry 7:e1169.
49. Kuan PF, Waszczuk MA, Kotov R, Marsit CJ, Guffanti G, Gonzalez A,et al. (2017): An epigenome-wide DNA methylation study of PTSD anddepression in World Trade Center responders. Transl Psychiatry 7:e1158.
50. Rutten BPF, Vermetten E, Vinkers CH, Ursini G, Daskalakis NP,Pishva E, et al. (2017): Longitudinal analyses of the DNA methylome indeployed military servicemen identify susceptibility loci for post-traumatic stress disorder [published online ahead of print Jun 20].Mol Psychiatry.
51. Bam M, Yang X, Zumbrun EE, Zhong Y, Zhou J, Ginsberg JP, et al.(2016): Dysregulated immune system networks in war veterans withPTSD is an outcome of altered miRNA expression and DNA methyl-ation. Sci Rep 6:31209.
52. Raison CL, Capuron L, Miller AH (2006): Cytokines sing the blues:Inflammation and the pathogenesis of depression. Trends Immunol27:24–31.
53. Maddox SA, Kilaru V, Shin J, Jovanovic T, Almli LM, Dias BG, et al.(2018): Estrogen-dependent association of HDAC4 with fear in femalemice and women with PTSD. Mol Psychiatry 23:658–665.
54. Admon R, Milad MR, Hendler T (2013): A causal model of post-traumatic stress disorder: Disentangling predisposed from acquiredneural abnormalities. Trends Cogn Sci 17:337–347.
55. Horvath S (2013): DNA methylation age of human tissues and celltypes. Genome Biol 14:R115.
56. Zannas AS, Arloth J, Carrillo-Roa T, Iurato S, Röh S, Ressler KJ,et al. (2015): Lifetime stress accelerates epigenetic aging in an ur-ban, African American cohort: Relevance of glucocorticoidsignaling. Genome Biol 16:266.
57. Marioni RE, Shah S, McRae AF, Chen BH, Colicino E, Harris SE, et al.(2015): DNA methylation age of blood predicts all-cause mortality inlater life. Genome Biol 16:25.
58. Klengel T, Mehta D, Anacker C, Rex-Haffner M, Pruessner JC,Pariante CM, et al. (2013): Allele-specific FKBP5 DNA
838 Biological Psychiatry May 15, 2018; 83:831–839 www.sobp.org/jo
demethylation mediates gene–childhood trauma interactions. NatNeurosci 16:33–41.
59. Wolf EJ, Logue MW, Hayes JP, Sadeh N, Schichman SA, Stone A,et al. (2016): Accelerated DNA methylation age: Associations withPTSD and neural integrity. Psychoneuroendocrinology 63:155–162.
60. Ratanatharathorn A, Boks MP, Maihofer AX, Aiello AE, Amstadter AB,Ashley-Koch AE, et al. (2017): Epigenome-wide association of PTSDfrom heterogeneous cohorts with a common multi-site analysis pipe-line. Am J Med Genet B Neuropsychiatr Genet 174:619–630.
61. Geschwind DH, Flint J (2015): Genetics and genomics of psychiatricdisease. Science 349:1489–1494.
62. Potkin S, Guffanti G, Lakatos A, Turner J, Kruggel F, Fallon J, et al.(2009): Hippocampal atrophy as a quantitative trait in a genome-wideassociation study identifying novel susceptibility genes for Alzheimer’sdisease. PLoS One 4:e6501.
63. Gottesman II, Gould TD (2003): The endophenotype concept in psy-chiatry: Etymology and strategic intentions. Am J Psychiatry 160:636–645.
64. Flint J, Timpson N, Munafò M (2014): Assessing the utility of inter-mediate phenotypes for genetic mapping of psychiatric disease.Trends Neurosci 37:733–741.
65. Thompson PM, Andreassen OA, Arias-Vasquez A, Bearden CE,Boedhoe PS, Brouwer RM, et al. (2017): ENIGMA and the individual:Predicting factors that affect the brain in 35 countries worldwide.NeuroImage 145(Pt B):389–408.
66. Logue MW, van Rooij SJH, Dennis EL, Davis SL, Hayes JP,Stevens JS, et al. (2018): Smaller hippocampal volume in post-traumatic stress disorder: A multisite ENIGMA-PGC study: Subcorticalvolumetry results from posttraumatic stress disorder consortia. BiolPsychiatry 83:244–253.
67. Bremner JD, Randall P, Scott TM, Bronen RA, Seibyl JP,Southwick SM, et al. (1995): MRI-based measurement of hippocampalvolume in patients with combat-related posttraumatic stress disorder.Am J Psychiatry 152:973–981.
68. Hibar DP, Stein JL, Renteria ME, Arias-Vasquez A, Desrivieres S,Jahanshad N, et al. (2015): Common genetic variants influence humansubcortical brain structures. Nature 520:224–229.
69. Hayes JP, Logue MW, Sadeh N, Spielberg JM, Verfaellie M,Hayes SM, et al. (2017): Mild traumatic brain injury is associated withreduced cortical thickness in those at risk for Alzheimer’s disease.Brain 140:813–825.
70. Vukojevic V, Kolassa IT, Fastenrath M, Gschwind L, Spalek K, Milnik A,et al. (2014): Epigenetic modification of the glucocorticoid receptorgene is linked to traumatic memory and post-traumatic stress disorderrisk in genocide survivors. J Neurosci 34:10274–10284.
71. Sadeh N, Spielberg JM, Logue MW, Wolf EJ, Smith AK, Lusk J, et al.(2016): SKA2 methylation is associated with decreased prefrontalcortical thickness and greater PTSD severity among trauma-exposedveterans. Mol Psychiatry 21:357–363.
72. Zhu L, Bhattacharyya A, Kurali E, Anderson A, Menius A, Lee K (2011):Review of integrative analysis challenges in systems biology. StatBiopharm Res 3:561–568.
73. Chang LC, Lin HM, Sibille E, Tseng GC (2013): Meta-analysis methodsfor combining multiple expression profiles: Comparisons, statisticalcharacterization and an application guideline. BMC Bioinformatics14:368.
74. Kim J, Patel K, Jung H, Kuo WP, Ohno-Machado L (2011): Any-Express: Integrated toolkit for analysis of cross-platform geneexpression data using a fast interval matching algorithm. BMC Bioin-formatics 12:75.
75. Campain A, Yang YH (2010): Comparison study of microarray meta-analysis methods. BMC Bioinformatics 11:408.
76. Ramasamy A, Mondry A, Holmes CC, Altman DG (2008): Key issues inconducting a meta-analysis of gene expression microarray datasets.PLoS Med 5:e184.
77. Chrétien S, Guyeux C, Conesa B, Delage-Mouroux R, Jouvenot M,Huetz P, et al. (2016): A Bregman-proximal point algorithm for robust non-negative matrix factorization with possible missing values and outliers—Application to gene expression analysis. BMC Bioinformatics 17:284.
urnal

Genomic Approaches to PTSDBiologicalPsychiatry
78. Wang J, Gamazon ER, Pierce BL, Stranger BE, Im HK, Gibbons RD,et al. (2016): Imputing gene expression in uncollected tissues withinand beyond GTEx. Am J Hum Genet 98:697–708.
79. Yang Y, Xu Z, Song D (2016): Missing value imputation for microRNAexpression data by using a GO-based similarity measure. BMC Bio-informatics 17(Suppl 1):10.
80. Li H, Zhao C, Shao F, Li GZ, Wang X (2015): A hybrid imputationapproach for microarray missing value estimation. BMC Genomics16(Suppl 9):S1.
81. Liew AW, Law NF, Yan H (2011): Missing value imputation for geneexpression data: Computational techniques to recover missing datafrom available information. Brief Bioinform 12:498–513.
82. Noureddine MA, Li YJ, van der Walt JM, Walters R, Jewett RM, Xu H,et al. (2005): Genomic convergence to identify candidate genes forParkinson disease: SAGE analysis of the substantia nigra. Mov Disord20:1299–1309.
83. Bharadwaj RA, Jaffe AE, Chen Q, Deep-Soboslay A, Goldman AL,Mighdoll MI, et al. (2018): Genetic risk mechanisms of posttraumaticstress disorder in the human brain. J Neurosci Res 96:21–30.
84. Almli LM, Stevens JS, Smith AK, Kilaru V, Meng Q, Flory J, et al.(2015): A genome-wide identified risk variant for PTSD is amethylation quantitative trait locus and confers decreased corticalactivation to fearful faces. Am J Med Genet B Neuropsychiatr Genet168:327–336.
85. Pasaniuc B, Price AL (2017): Dissecting the genetics of complex traitsusing summary association statistics. Nat Rev Genet 18:117–127.
86. Mustapic M, Maihofer AX, Mahata M, Chen Y, Baker DG,O’Connor DT, et al. (2014): The catecholamine biosynthetic enzymedopamine beta-hydroxylase (DBH): First genome-wide search posi-tions trait-determining variants acting additively in the proximal pro-moter. Hum Mol Genet 23:6375–6384.
87. Polimanti R, Amstadter AB, Stein MB, Almli LM, Baker DG, Bierut LJ,et al. (2017): A putative causal relationship between genetically
Biological
determined female body shape and posttraumatic stress disorder.Genome Med 9:99.
88. Consortium G (2015): The Genotype-Tissue Expression (GTEx) pilotanalysis: Multitissue gene regulation in humans. Science 348:648–660.
89. Akbarian S, Liu C, Knowles JA, Vaccarino FM, Farnham PJ,Crawford GE, et al. (2015): The PsychENCODE project. Nat Neurosci18:1707–1712.
90. Gamazon ER, Wheeler HE, Shah KP, Mozaffari SV, Aquino-Michaels K, Carroll RJ, et al. (2015): A gene-based association methodfor mapping traits using reference transcriptome data. Nat Genet47:1091–1098.
91. Cadwell CR, Scala F, Li S, Livrizzi G, Shen S, Sandberg R, et al.(2017): Multimodal profiling of single-cell morphology, electro-physiology, and gene expression using Patch-seq. Nat Protoc12:2531–2553.
92. Wu YE, Pan L, Zuo Y, Li X, Hong W (2017): Detecting activated cellpopulations using single-cell RNA-seq. Neuron 96:313–329.
93. Schwartzentruber J, Foskolou S, Kilpinen H, Rodrigues J, Alasoo K,Knights AJ, et al. (2018): Molecular and functional variation in iPSC-derived sensory neurons. Nat Genet 50:54–61.
94. Wang P, Mokhtari R, Pedrosa E, Kirschenbaum M, Bayrak C,Zheng D, et al. (2017): CRISPR/Cas9-mediated heterozygousknockout of the autism gene CHD8 and characterization of itstranscriptional networks in cerebral organoids derived from iPScells. Mol Autism 8:11.
95. Camp JG, Badsha F, Florio M, Kanton S, Gerber T, Wilsch-Brauninger M, et al. (2015): Human cerebral organoids recapitulategene expression programs of fetal neocortex development. Proc NatlAcad Sci U S A 112:15672–15677.
96. Jo J, Xiao Y, Sun AX, Cukuroglu E, Tran HD, Goke J, et al. (2016):Midbrain-like organoids from human pluripotent stem cells containfunctional dopaminergic and neuromelanin-producing neurons. CellStem Cell 19:248–257.
Psychiatry May 15, 2018; 83:831–839 www.sobp.org/journal 839





























