Embed Size (px)
Citation preview

Genetic linkage analysisDotan SchreiberAccording to a series of presentations by M. Fishelson

OutLine
• Introduction.
• Basic concepts and some background.
• Motivation for linkage analysis.
• Linkage analysis: main approaches.
• Latest developments.

“Genetic linkage analysis is a statistical method that is used to associate functionality of genes to their location on chromosomes.“
http://bioinfo.cs.technion.ac.il/superlink/

The Main Idea/usage:
Neighboring genes on the chromosome have a tendency to stick together when passed on to offsprings.
Therefore, if some disease is often passed to offsprings along with specific marker-genes , then it can be concluded that the gene(s) which are responsible for the disease are located close on the chromosome to these markers.
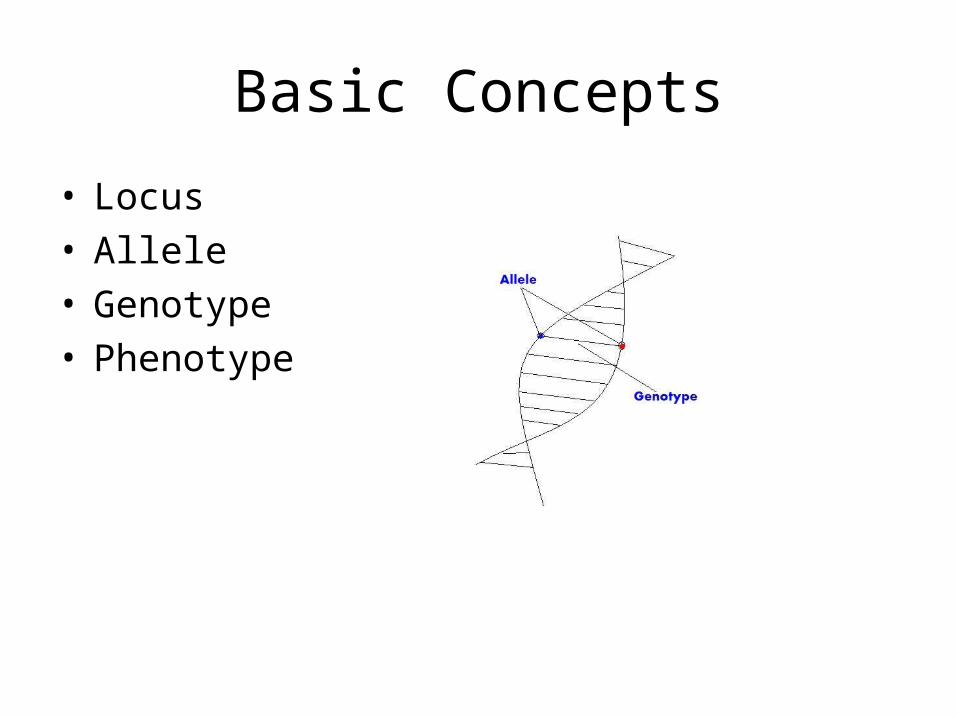
Basic Concepts
• Locus• Allele• Genotype• Phenotype

Dominant Vs. Recessive Allele
דוגמא קלאסית: צבע עיניים
homozygoteheterozygote

)se(X-Linked Allele
Most human cells contain 46 chromosomes:• 2 sex chromosomes (X,Y):
XY – in males.
XX – in females.• 22 pairs of chromosomes named autosomes.
Around 1000 human alleles are found only on the X chromosome.

“…the Y chromosome essentially is reproduced via cloning from one generation to the next. This prevents mutant Y chromosome genes from being eliminated from male genetic lines. Subsequently, most of the human Y chromosome now contains genetic junk rather than genes.”
http://anthro.palomar.edu/biobasis/bio_3b.htm

Medical Perspective
When studying rare disorders, 4 general patterns
of inheritance are observed:• Autosomal recessive (e.g., cystic fibrosis).
– Appears in both male and female children of unaffected parents.
• Autosomal dominant (e.g., Huntington disease).– Affected males and females appear in each generation of the
pedigree.– Affected parent transmits the phenotype to both male and
female children.

Continued..
• X-linked recessive (e.g., hemophilia).– Many more males than females show the disorder.– All daughters of an affected male are “carriers”.– None of the sons of an affected male show the disorder or are
carriers.
• X-linked dominant.– Affected males pass the disorder to all daughters but to none of
their sons.– Affected heterozygous females married to unaffected males
pass the condition to half their sons and daughters.
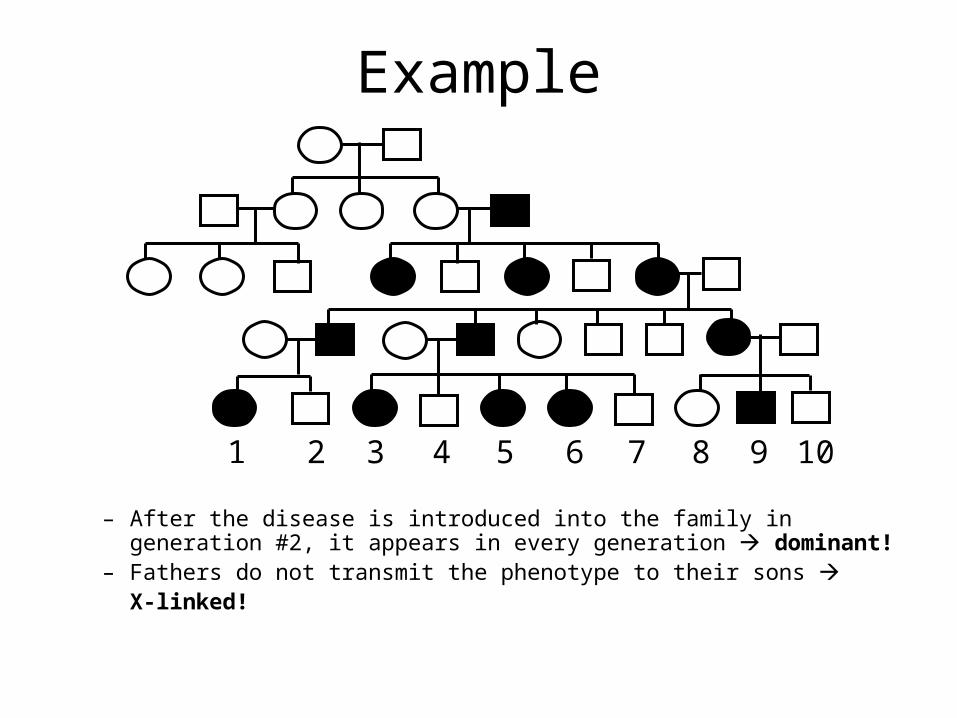
Example
– After the disease is introduced into the family in generation #2, it appears in every generation dominant!
– Fathers do not transmit the phenotype to their sons X-linked!
1 2 3 4 5 6 7 8 9 10

Crossing Over
Sometimes in meiosis, homologous chromosomes exchange parts in a process called crossing-over, or recombination.

Recombination Fraction
The probability for a recombination between two genes is a monotone, non-linear function of the physical distance between their loci on the chromosome.
Linkage) No(5.0)ionRecombinat(0)Linkage( P

Linkage
The further apart two genes on the same chromosome are, the more it is likely that a recombination between them will occur.
Two genes are called linked if the recombination fraction between them is small (<< 50% chance)

Linkage related Concepts
• Interference - A crossover in one region usually decreases the probability of a crossover in an adjacent region.
• CentiMorgan (cM) - 1 cM is the distance between genes for which the recombination frequency is 1%.
• Lod Score - a method to calculate linkage distances (to determine the distance between genes).

Ultimate Goal: Linkage Mapping
With the following few minor problems:– It’s impossible to make controlled crosses in
humans.– Human progenies are rather small.– The human genome is immense. The
distances between genes are large on average.

Possible Solutions
• Make general assumptions:Hardy-Weinberg Equilibrium – assumes certain probability
for a certain individual to have a certain genotype.
Linkage Equilibrium – assumes two alleles at different loci are independent of each other.
• Incorporate those assumptions into possible solutions:
Elston-Stewart method.
Lander-Green method.

Elston-Stewart method
• Input: A simple pedigree + phenotype information about some of the people. These people are called typed.
• Simple pedigree – no cycles, single pair of founders.
founder
leaf1/2

..Continued
• Output: the probability of the observed data, given some probability model for the transmission of alleles. Composed of:
founder probabilities - Hardy-Weinberg equilibrium
penetrance probabilities -The probability of the phenotype, given the genotype
transmission probabilities -the probability of a child having a certain genotype given the parents’
genotypes

..Continued
• Bottom-Up: sum conditioned probabilities over all possible genotypes of the children and only then on the possible genotypes for the parents.
• Linear in the number of people.
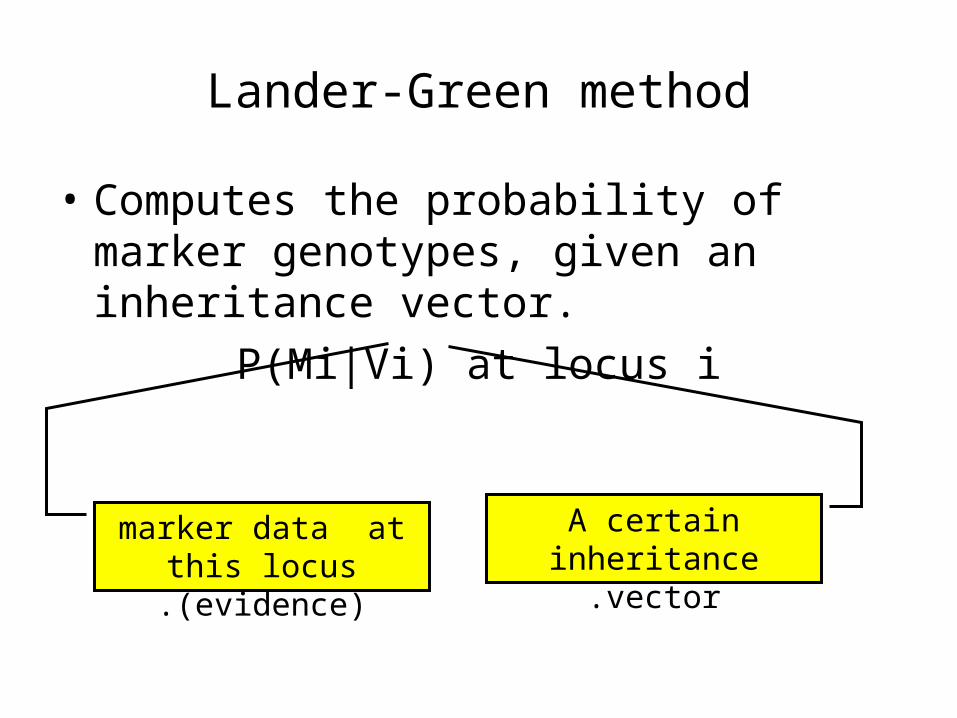
Lander-Green method
• Computes the probability of marker genotypes, given an inheritance vector.
P(Mi|Vi) at locus i
marker data at this locus (evidence).
A certain inheritance vector.

Main Idea• Let a = (a1,…,a2f) be a vector of alleles assigned to
founders of the pedigree (f is the number of founders).
• We want a graph representation of the restrictions imposed by the observed marker genotypes on the vector a that can be assigned to the founder genes.
• The algorithm extracts only vectors a compatible with the marker data.
• Pr[m|v] is obtained via a sum over all compatible vectors a.

Example – marker data on a pedigree
1 2
1211
a/ba/b
21
13
22
14
a/ba/b
23 24
b/da/c
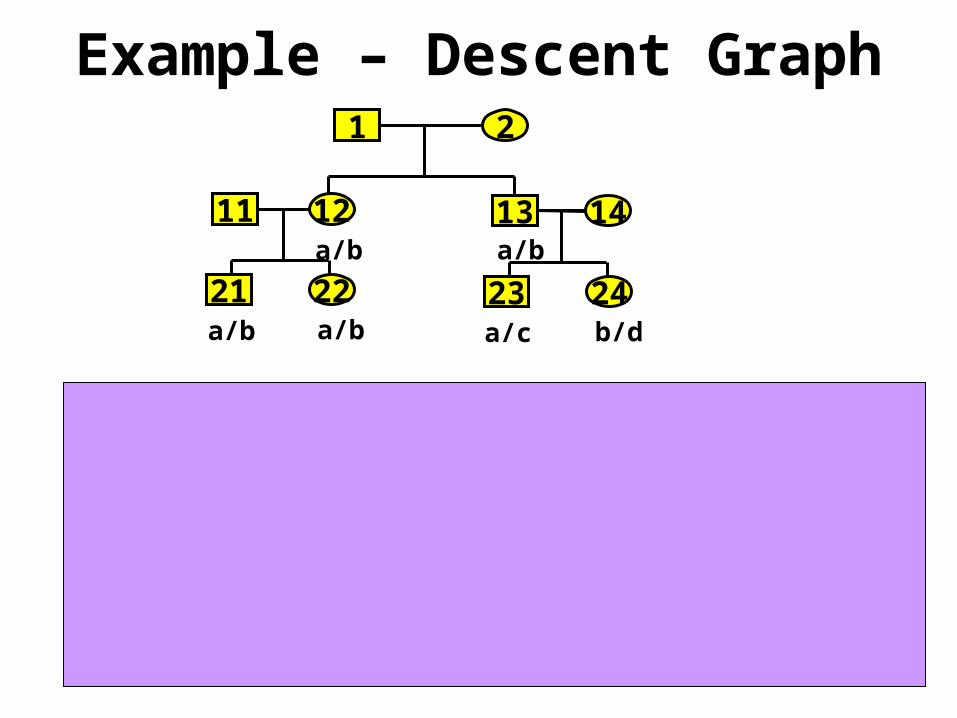
Example – Descent Graph1 2
1211a/ba/b
21
13
22
14
a/ba/b23 24
b/da/c
3 4 5 6
1 2 7 8)a,b(
)a,c( )b,d()a,b(
)a,b(
)a,b(
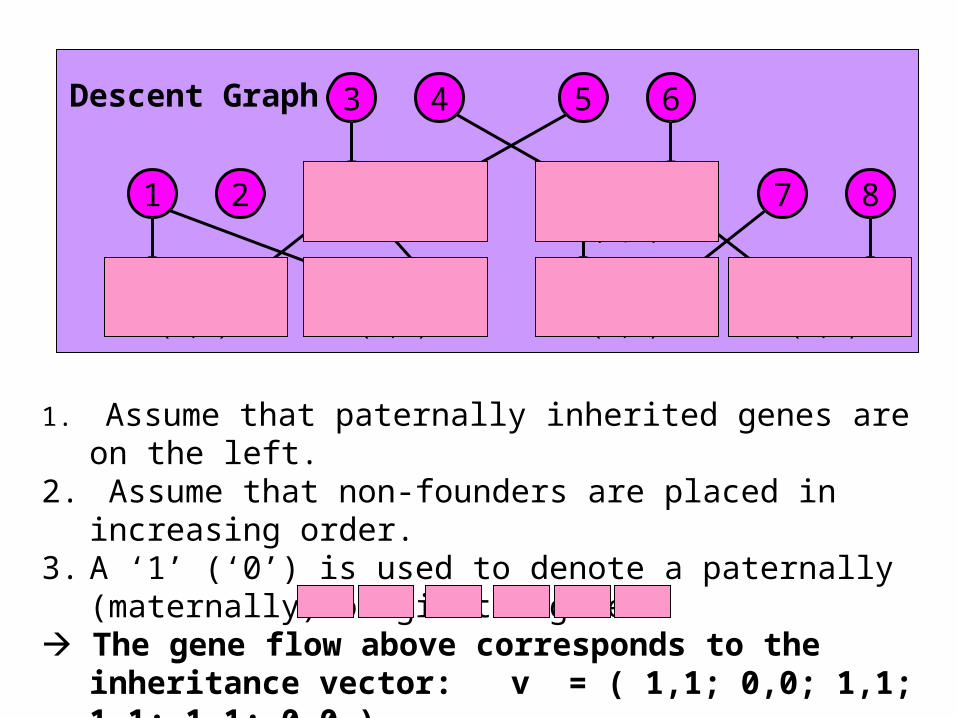
Descent Graph
3 4 5 6
1 2 7 8)a,b(
)a,c( )b,d()a,b(
)a,b(
)a,b(
Descent Graph
1. Assume that paternally inherited genes are on the left. 2. Assume that non-founders are placed in increasing order.3. A ‘1’ (‘0’) is used to denote a paternally (maternally)
originated gene. The gene flow above corresponds to the inheritance
vector: v = ( 1,1; 0,0; 1,1; 1,1; 1,1; 0,0 )

Example – Founder Graph
5 3
2 1
6 4
8 7
)b,d(
)a,b(
)a,b( )a,c(
)a,b(
Founder Graph
3 4 5 6
1 2 7 8)a,b(
)a,c( )b,d()a,b(
)a,b(
)a,b(
Descent Graph
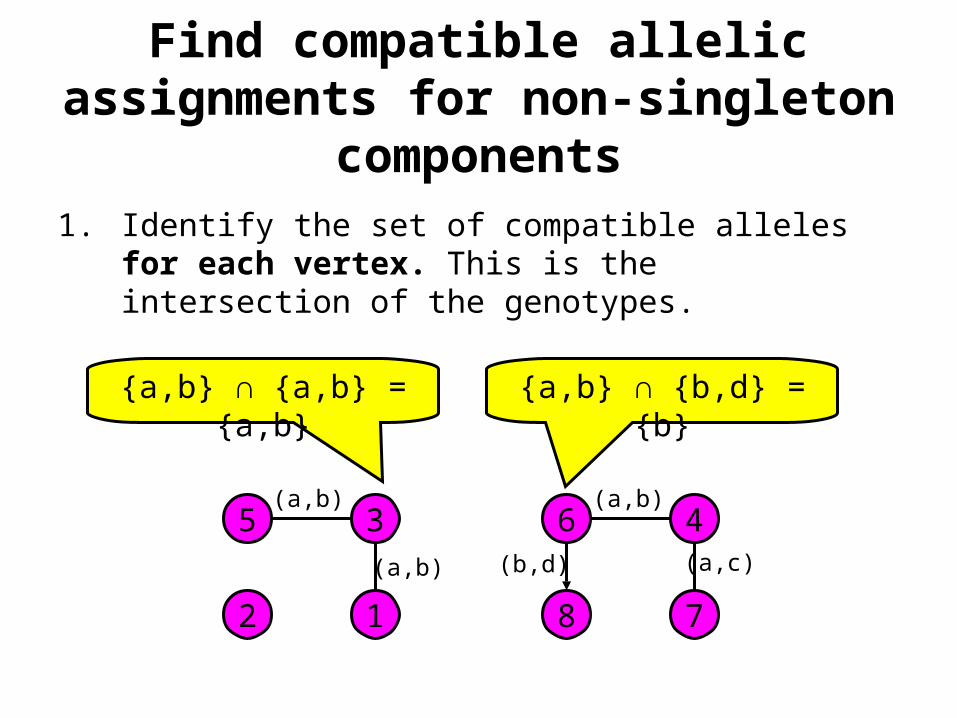
Find compatible allelic assignments for non-singleton components
1. Identify the set of compatible alleles for each vertex. This is the intersection of the genotypes.
5 3
2 1
6 4
8 7
)b,d(
)a,b(
)a,b( )a,c(
)a,b(
{a,b} ∩ {a,b} = {a,b} {a,b} ∩ {b,d} = {b}

Possible Allelic Assignments
5 3
2 1
6 4
8 7
)b,d(
)a,b(
)a,b( )a,c(
)a,b(}a,b{ }a,b{
}a,b{ }a,c{
}a{}b{
}b,d{}a,b,c,d{
Graph ComponentAllelic Assignments
(2)(a), (b), (c), (d)
(1,3,5)(a,b,a), (b,a,b)
(4,6,7,8)(a,b,c,d)

Computing P(m|v)
• If for some component there are no possible allelic assignments, then P(m|v) = 0.
• The probability of singleton components is 1 we can ignore them.
• Let ahi be an element of a vector of alleles assigned to the vertices of component Ci.
]Pr[]Pr[}:{
iCjj
jhi aa
]Pr[]Pr[}:{
ihi Aah
hii aC
]Pr[]|Pr[1
m
iiCvm
over 2f elements
2 terms at most
Linear in the number of founders

Latest News: SuperLink
• Combines the covered approaches in one unified program.
• Has other built-in abilities that increase its computations efficiency.
• Claimed to be more capable and faster than other related programs (by its own makers).
• http://bioinfo.cs.technion.ac.il/superlink/

The End





























