Embed Size (px)
Citation preview

Developmental Biology 344 (2010) 1088–1099
Contents lists available at ScienceDirect
Developmental Biology
j ourna l homepage: www.e lsev ie r.com/deve lopmenta lb io logy
Genomes & Developmental Control
Genetic interactions of eyes absent, twin of eyeless and orthodenticle regulate sineoculis expression during ocellar development in Drosophila
Jorge Blanco a,c,⁎, Tobias Pauli a, Makiko Seimiya b, Gerald Udolph c, Walter J. Gehring a
a Biozentrum, University of Basel, Klingelbergstrasse 70, CH-4056 Basel, Switzerlandb Department of Biosystems Science and Engineering, Mattenstrasse 26, CH-4058 Basel, Switzerlandc Institute of Medical Biology, 8A Biomedical Grove, Singapore 138648, Singapore
⁎ Corresponding author. Institute of Medical Biology, 8138648, Singapore. Fax: +65 64642048.
E-mail address: [email protected] (J. B
0012-1606/$ – see front matter © 2010 Elsevier Inc. Adoi:10.1016/j.ydbio.2010.05.494
a b s t r a c t
a r t i c l e i n f oArticle history:Received for publication 21 April 2010Revised 14 May 2010Accepted 15 May 2010Available online 24 May 2010
Keywords:DrosophilaEyes absentEye/ocellus enhancerOcelli developmentOrthodenticleSine oculisTwin of eyeless
The homeobox gene sine oculis (so) is required for the development of the entire visual system in Drosophila,which includes the compound eyes, the ocelli, the optic lobe of the brain and the Bolwig's organ. Duringocelli development, so expression labels, together with eyes absent (eya), the emergence of the ocellarprecursor cells in the third instar eye-antennal disc. Footprinting and misexpression studies have led to theproposal that the Pax6 homologue twin of eyeless (toy) directly regulates the initiation of so expression inocellar precursor cells. However, so expression in a toy loss-of-function mutant background has not been yetanalyzed due to the lack of eye-antennal disc development in strong toy mutant alleles. Using an embryoniceye primordium-specific enhancer of toy, we have rescued the developmental defect of a strong toy mutantallele and analyzed so expression in the ocelli primordium of toy loss-of-function eye-antennal discs duringthird instar larva. The results show that so expression is only marginally affected in the absence of Toytranscriptional activity and that the toy positive effect on so expression is largely eya-mediated. These resultssuggest that eya is the main factor controlling both initiation and maintenance of so expression in ocellarprecursor cells. In addition, we present the characterization of a new minimal eye/ocellus-specific enhancerof the so gene.
A Biomedical Grove, Singapore
lanco).
ll rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Introduction
The Drosophila adult visual system provides with an interestingexample of functionally homologous organs. It consists of thestructurally complex compound eye and three simple light-sensitiveocelli located at the top of the head. Both organs arise from specificprogenitor cells present in the eye-antennal imaginal disc during thirdinstar larva and their development can be traced back to lateembryogenesis (stage 15), when a subset of cells from the anteriorectoderm is set aside to constitute the embryonic eye primordium, e.g.the precursor of the larval eye-antennal disc (Green et al., 1993;Younossi-Hartenstein et al., 1993). These cells divide and grow duringtwo consecutive larval instar stages to give rise to a mature thirdinstar eye-antennal imaginal disc containing two different morpho-genetic fields (eye and antenna), each one harboring severalprimordia of adult head structures (reviewed by Domìnguez andCasares, 2005; Haynie and Bryant, 1986). The compound eye derivesfrom progenitor cells present in the central part of the eye morpho-
genetic field, whereas the primordium of the ocelli arises as twoclusters of cells in the anterior dorsal part of the eye field (Royet andFinkelstein, 1996).
Behind their morphological disparity, profound homologies existin the genetic programs that control the development of both visualorgans. Extensive analysis of the genes and regulatory circuitsinvolved in the determination of the compound eye and ocelliprimordia has defined a group of evolutionarily conserved genescollectively known as retinal determination (RD) genes. They areinitially organized in a sequential and hierarchical genetic cascade,which later turns into a network of genetic interactions (for reviewsee Gehring, 2005; Kumar, 2009; Pappu and Mardon, 2004; Silver andRebay, 2005). The Pax6 homologues eyeless (ey) and twin of eyeless(toy) are at the top of the genetic cascade (Gehring and Ikeo, 1999;Gehring, 2004). They form, together with the downstream genes eyesabsent (eya) (Bonini et al., 1993) and sine oculis (so) (Cheyette et al.,1994), the core of the RD network. ey and toy are expressed first, incombination with the Pax6-like genes eyegone (eyg) and twin ofeyegone (toe), in the eye primordium during late embryogenesis(Czerny et al., 1999; Jun et al., 1998; Quiring et al., 1994; Yao et al.,2008). Whereas mutations negatively affecting ey function inhibitcompound eye development (Punzo et al., 2002; Gehring and Seimiya,2010), toy or eyg loss-of-function mutations impair the developmentof the whole eye-antennal disc and give rise to flies lacking all the

1089J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
structures derive from this imaginal disc (headless flies) (Gehring andSeimiya, 2010; Jang et al., 2003; Kronhamn et al., 2002). Moreover, toyand ey expression at this stage shows a clear epistatic relationship, asey transcription depends on toy activity (Czerny et al., 1999). Apartfrom promoting cell proliferation in the early eye disc, eyg also actscooperatively with ey during compound eye development (Jang et al.,2003). Thus, double toy and ey loss-of-function headless mutantsshow remnants of the compound eyes (two “eyeballs” of facets) in theanterior thorax, which are only removed upon introduction of a thirdloss-of-function mutation in the eyg gene (Gehring and Seimiya,2010).
During first instar larva, both ey and toy are expressed in the entireeye-antennal disc. Later, their expression becomes restricted to theposterior part of the disc, determining its division into two majormorphogenetic fields: eye field (Ey and Toy positive) and antennalfield (Ey and Toy negative) (Kenyon et al., 2003). The downstreamretinal genes eya and so are expressed in two different domains of theeyemorphogenetic field, labeling the emergence of the compound eyeand ocelli primordia. eya and so interact at the protein level forming acomplex that works as a transcriptional activator (Pignoni et al., 1997)and mutations that negatively affect their function impair bothcompound eye and ocelli development. Moreover, Eya has beenshown to have protein phosphatase activity (Li et al., 2003; Tootleet al., 2003), which is important for cytoplasmic signaling eventsrelevant to eye development (Xiong et al., 2009).
The regulation of eya and so expression in the compound eye andocelli primordia is differential and depends on the unequal distribu-tion of Ey and Toy in the eye morphogenetic field during third instarlarvae. Ey is present in the compound eye primordium, where itactivates eya and so expression (Halder et al., 1998; Niimi et al., 1999;Zimmerman et al., 2000), but is absent from the ocelli primordium.Therefore, ey loss-of-functions mutations impair compound eyeformation but not ocelli development (Punzo et al., 2002; Gehringand Seimiya, 2010). On the contrary, Toy is present in both thecompound eye and ocelli primordia. Whereas toy function in thecompound eye primordium seems to be negligible (Punzo et al.,2002), it has been proposed that so expression in the ocelliprimordium is initiated by toy (Punzo et al., 2002; Pauli et al.,2005). Indeed, the eye/ocellus-specific enhancer of the so genecontains Toy-specific binding sites, whose mutations abolish enhanc-er activity in the ocelli primordium (Punzo et al., 2002). Later, the Soprotein itself seems to be necessary for proper so expression throughits binding to an autoregulatory element (soAE) (Pauli et al., 2005). Onthe contrary, eya expression in the ocelli primordium appears to betoy-independent, but indirectly controlled by the homeodomaincontaining gene orthodenticle (otd) (Blanco et al., 2009). In summary,the current model supports the view of two independent regulatorypathways (one controlled by toy and the other by otd) regulating(respectively) so and eya expression in the ocelli primordium, withtoy controlling just the initiation of so expression. In order to test thismodel, two predictions can be made. First, so expression in the ocelliprimordium of toy loss-of-functions mutants should be abolished,whereas eya expression should remain unaffected. Second, in toy loss-of-function mutants, ocelli formation, but not compound eyedevelopment, should be impaired.
So far, toyhdl is the unique toy mutant allele which has beenmolecularly characterized. It is considered a null with respect to eyeand brain development and homozygous toyhdl flies die as pharatesshowing a headless phenotype (Kronhamn et al., 2002; Furukubo-Tokunaga et al., 2009). This phenotype is due to the absence of Toytranscriptional activity in the embryonic eye primordium, whichblocks subsequent development of the eye-antennal disc during larvalstages (Kronhamn et al., 2002). Therefore, in order to analyze so andeya expression in the third instar eye-antennal disc of a toy loss-of-function mutant, we need first to rescue the developmental defect ofthe eye primordium in toyhdl embryos.
In this paper, we have isolated the embryonic eye primordium-specific enhancer of the toy gene. Using this enhancer, we haverescued the developmental defect of the eye primordium in toyhdl
embryos and tested the predictions described above regarding theexpression of so and eya in a toy loss-of-function eye-antennal discduring third instar larva. In addition, we have further characterizedthe eye/ocellus-specific enhancer of the so gene and defined a newminimal eye/ocellus-specific enhancer.
Materials and methods
Fly strains and clonal analysis
Flies were reared on standard medium at 25 °C. The followingtransgene and reporter lines were used: dppblink-Gal4 (Staehling-Hampton and Hoffmann, 1994), so7-Gal4 (Punzo et al., 2002), UAS-otd (J. Blanco, unpublished), UAS-ey (Halder et al., 1995), UAS-toy(Czerny et al., 1999), UAS-eya (Bonini et al., 1997). Mutant allelesused in this study: ocγa1, otdYH13, eyacli-IID, FRT42D so3 (Pignoni et al.,1997), so1 (Cheyette et al., 1994), toyhdl (Kronhamn et al., 2002).Mutant alleles without reference are described in Flybase (http://flybase.bio.indiana.edu).
Mitotic clones were generated and positively labeled (withmembrane tethered CD8::GFP) according to the MARCM technique(Lee and Luo, 1999), using transgenes and recombinant chromosomesobtained from the Bloomington Stock Center. Recombination wasinduced 60± 12 h after egg laying by a 1 h heat shock at 37 °C and thelarvae were dissected 48 h later. Genotypes of the analyzed larvaewere as follows:
- eyacli-IID clones: y w hs-FLP/+; eyacli-IID FRT40A/tubP-GAL80LL10
FRT40A; tubP-GAL4LL7 UAS-mCD8::GFPLL6/+* (UAS-ey, UAS-toy orUAS-otd instead of+* for the misexpression of ey, toy or otd,respectively, in eyacli-IID clones).
- so3 clones: y w hs-FLP/+; FRT42D so3/FRT42D tubP-GAL80; tubP-GAL4LL7 UAS-mCD8::GFPLL6
- wild-type clones: y w hs-FLP/+; FRT40A/tubP-GAL80LL10 FRT40A;tubP-GAL4LL7 UAS-mCD8::GFPLL6/+* (UAS-ey, UAS-toy, UAS-otd orUAS-eya instead of+* for the misexpression of ey, toy, otd or eya,respectively, in wild-type clones).
DNA constructs and transgenic flies
Enhancer activity was monitored in transgenic flies containingreporter constructs. Briefly, the different DNA fragments wereobtained by PCR and cloned into the pcβ plasmid (Niimi et al.,1999) upstream of the lacZ gene. The EEP enhancer was also clonedinto the pPhsp-Gal4 vector (Punzo et al., 2002) upstream of the gal4gene, generating the EEP-Gal4 driver line. For the so1 rescueexperiment, the new minimal eye/ocellus-specific enhancer of theso gene (so11-soAE) was cloned upstream of the so cDNA using thepUAST vector as a backbone (as described in Pauli et al., 2005). TheGal4 driver line, the so1 rescue construct and the lacZ reporterconstructs were injected into Drosophila w1118 embryos followingstandard P-element transformation protocols. Three to five indepen-dent transgenic lines were established for each construct.
β-Galactosidase staining and immunohistochemistry
To detect β-galactosidase activity, third instar larval imaginal discswere fixed and subjected to a standard X-gal color reaction at 37 °C.Antibody staining on discs was performed according to Halder et al.(1998). Primary antibodies were as follows: rabbit anti-β-galactosi-dase (β-gal 1:200; Cappel), mouse anti- β-gal (1:500; Promega),rabbit anti-Otd (1:250; Hirth et al., 2003), mouse anti-Eya (1:10;
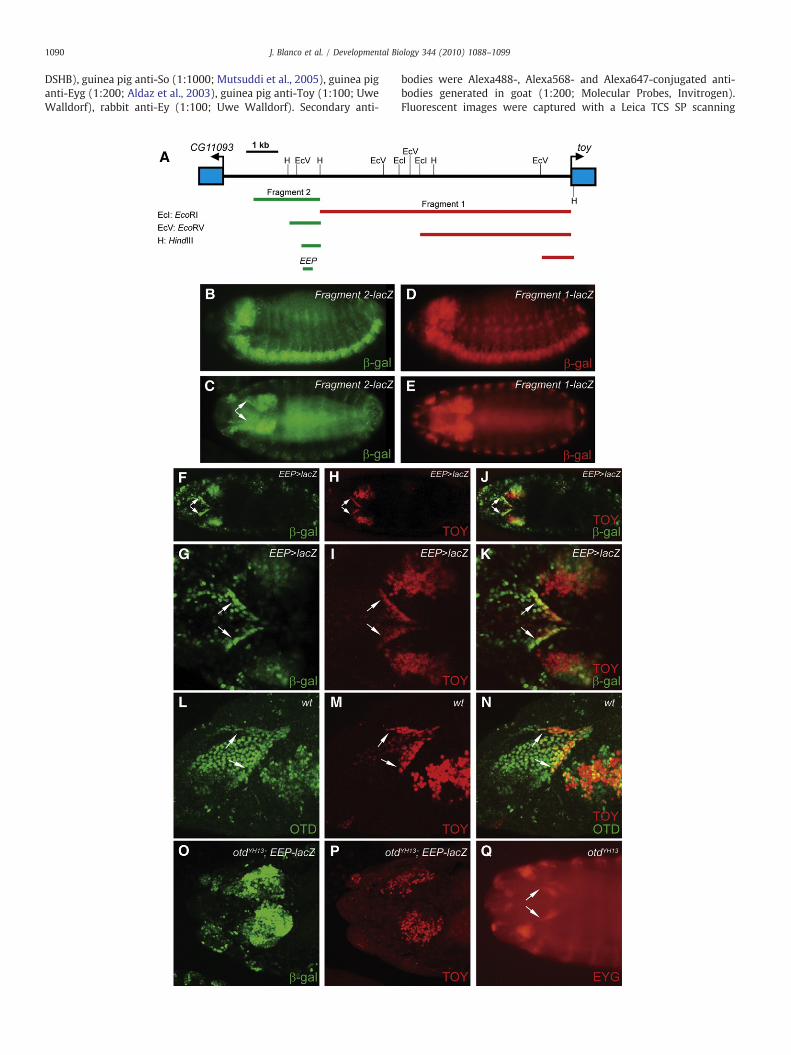
1090 J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
DSHB), guinea pig anti-So (1:1000; Mutsuddi et al., 2005), guinea piganti-Eyg (1:200; Aldaz et al., 2003), guinea pig anti-Toy (1:100; UweWalldorf), rabbit anti-Ey (1:100; Uwe Walldorf). Secondary anti-
bodies were Alexa488-, Alexa568- and Alexa647-conjugated anti-bodies generated in goat (1:200; Molecular Probes, Invitrogen).Fluorescent images were captured with a Leica TCS SP scanning

1091J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
confocal microscope. Unless indicated, pictures correspond to singleoptical sections (0.5 μm thick).
Results
Embryonic eye primordium-specific enhancer of the toy gene
The Toyhdl allele codes for a truncated Toy protein that lacks part ofthe homeodomain and the C-terminal transactivation domain(Kronhamn et al., 2002). It is considered a null with respect to eye andbrain development and homozygous toyhdl flies die as pharates showinga headless phenotype (Kronhamn et al., 2002; Furukubo-Tokunaga et al.,2009). This phenotype is due to the absence of Toy transcriptionalactivity in the embryonic eye primordium (the precursor of the eye-antennal disc) and the subsequent blockage in the development of thisimaginal disc during larval stages (Kronhamn et al., 2002).
In order to analyze so expression in the third instar eye-antennaldisc of a toy loss-of-function mutant, we needed first to rescue thedevelopmental defect of the embryonic eye primordium in toyhdl flies.Ideally, the rescue experiment would involve the expression of the toycDNA under the control of the embryonic eye primordium-specificenhancer of the toy gene. In order to find this enhancer, we analyzedin transgenic reporter constructs the 5′ genomic region of toy andsearched for cis-regulatory sequences active during embryogenesis.We found two DNA fragments which showed similar enhanceractivity in the brain and central nerve cord during middle and latestages of embryogenesis (Fig. 1A–E), consistent with the endogenoustoy expression pattern. Fragment 1 (red bar in Fig. 1A) was furtherreduced to a 1 kb long DNA sequence lying just upstream of the toytranscription initiation site and which maintained the originalenhancer activity (Fig. 1D and E). Recently, similar results havebeen reported by others (Jacobsson et al., 2009), indicating that thegenomic region surrounding the transcription start site is importantfor toy expression during embryogenesis. However, contrary to theirresults, no enhancer activity was detected in the embryonic eyeprimordium either using the 1 kb DNA fragment or bigger fragmentsuncovering more upstream sequences. On the other hand, fragment2 (green bar in Fig. 1A), which lies 8.2 kb upstream of the toytranscription initiation site, did show enhancer activity in theembryonic eye primordium (Fig. 1C). Deletion analysis allowed thereduction of this fragment to a 300 bp long DNA sequence(Supplementary Fig. 2) that retained enhancer activity in the eyeprimordium and, therefore, was named embryonic eye primordium-specific enhancer of the toy gene (or EEP enhancer).
To further investigate the role of the EEP enhancer in embryoniceye primordium development, a Gal4 driver line containing the EEPenhancer (EEP-gal4) was generated (Fig. 1F–K) and used in acomplementation assay to rescue the toyhdl mutation. Expression ofthe toy cDNA under the control of the EEP-gal4 driver line rescuedboth the embryonic eye primordium developmental defect and thepupal lethality phenotype associated with toyhdl (Fig. 2P).
Apart from ey, toy and the Pax6-like homologues eyg and toe, theselector gene orthodenticle (otd) is also expressed in the embryoniceye primordium (arrows in Fig. 1L–N). otd encodes for a home-odomain protein required during embryogenesis for the correctdevelopment of the head and the ventral midline (Finkelstein et al.,1990; Royet and Finkelstein, 1995). As mentioned in the Introduction,
Fig. 1. Isolation and characterization of the EEP enhancer. (A) Map of the 5′ genomic regiopresence of embryonic enhancer activity. (B–E) Enhancer activity of fragment 1 (D and E) andtype embryos using anti-β-galactosidase (β-gal) immunocytochemistry. B and D show laprimordium. (F–K) Comparison of EEP enhancer activity with toy expression pattern in a sembryonic eye primordium (arrows). F–J show dorsal views of the entire embryo, whereas Gstage 15 wild-type embryo showing Otd and Toy co-localization in the embryonic eye prishowing β-gal (O) and Toy (P) protein distribution. (Q) Eyg is present in the eye primordprojections of individual confocal optical sections.
ey expression in the embryonic eye primordium depends on toytranscriptional activity (Czerny et al., 1999; Kronhamn et al., 2002).However, how toy expression is regulated in this embryonic structureis so far unknown. Thus, we checked whether otd could be involved inthis process. The results showed that both toy expression and EEPenhancer activity were abolished in the embryonic eye primordium ofembryos homozygous for a null otd allele (Fig. 1O and P). On thecontrary, eyg expression was unaffected in this mutant background(Fig. 1Q). These results indicate that the embryonic eye primordium ispresent in otd mutant embryos and that toy expression in thisstructure is otd-dependent.
Analysis of so and eya expression in toyhdl third instar eye-antennalimaginal discs
The emergence of the ocelli primordium in the anterior dorsal partof the third instar eye-antennal disc is labeled by eya and socoexpression (arrows in Fig. 2A–C). The two genes are differentlyregulated in ocellar precursor cells: otd, through the maintenance ofHedgehog (Hh) signaling, controls eya expression (Blanco et al., 2009),whereas eya is necessary to maintain so expression through a positiveautoregulatory loop (Pauli et al., 2005). As a consequence, in otdmutants not only eya, but also so, failed to be expressed in ocelliprecursor cells (arrow in Fig. 2F). Although maintenance of soexpression in the ocelli primordium needs eya, initial so expressionwas postulated to be dependent on toy transcriptional activity (Punzoet al., 2002; Pauli et al., 2005). To check this hypothesis, wemadeuse ofthe EEP-gal4 driver line. The EEP enhancer is not active in the thirdinstar eye-antennal disc. Hence, the expression of toy under thecontrol of this enhancer permits the development of the embryoniceye primordium in toyhdl embryos, but gives rise in third instar larva toa mature eye-antennal disc deprived of Toy transcriptional activity(Fig. 2G). In this context, so expression in ocellar precursor cells shouldbe abolished (initiation of so expression is blocked), whereas eyaexpression should remain unaffected. However, in 60% of the cases(n=46) the expression of so and eya in the ocelli primordium wasnormal and gave rise to adult flies withwild-type ocelli (Fig. 2M). Onlyin 4% of the cases so and eya expressionwas greatly or totally abolished(arrow in Fig. 2I), giving rise to complete ocelliless adult flies (Fig. 2N).In the rest of the cases, the expression of both so and eyawas reduced,although eya showed a broader, yet not wild-type, expression patternthan so in ocellar precursor cells (arrows in Fig. 2J–L). Accordingly, thecorresponding adult flies displayed variable ocelliless phenotypes(Fig. 2O). In any case, compound eye formation was normal,supporting the role of ey+ as the Drosophila Pax6 homologuecommitted with the development of this structure.
Why is so expression inocellarprecursor cells not totally abolished inthe absence of Toy transcriptional activity? And why is eya expressionaffected at all? To answer these questions we first analyzed if theexpression of otd (a positive eya regulator) was also affected in thiscontext. No change in otd expression was detected in the ocelliprimordium of third instar eye-antennal discs deprived of Toytranscriptional activity (Fig. 2H). We also checked if eya expressioncould be affected by the lack of So, by inducing somutant cell clones inthe ocellar region of third instar eye-antennal discs. Depletion of So inthese cell clones (Fig. 3A–C) did not negatively affect eya expression. Onthe other hand, so expression was lost in a cell autonomous manner in
n of the toy gene showing the fragments analyzed in lacZ reporter constructs for thefragment 2 (B and C) was analyzed in a stage 13 (B and D) and stage 15 (C and E) wild-teral views and C and E dorsal views. The arrows in C point to the embryonic eyetage 15 wild-type embryo. EEP enhancer activity and toy expression co-localize in the–H show amagnification of the embryonic eye primordium. (L–N) Dorsolateral view of amordium (arrows). (O and P) Dorsolateral view of a stage 15 otdYH13 mutant embryoium (arrows) of a stage 15 otdYH13 mutant embryo. All the pictures correspond to Z

1092 J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
eyamutant cell clones (Fig. 3D–F), despite of the fact that toy (and otd)expression remained unaffected in these mutant cells (Fig. 3G–L).
In summary, we can conclude that the absence of Toy transcrip-tional activity must be directly or indirectly responsible for the down-
regulation of eya expression in ocellar precursor cells of toyhdl thirdinstar eye-antennal discs. In addition, the results also imply thateya is the main factor controlling so activation in ocellar precursorcells.
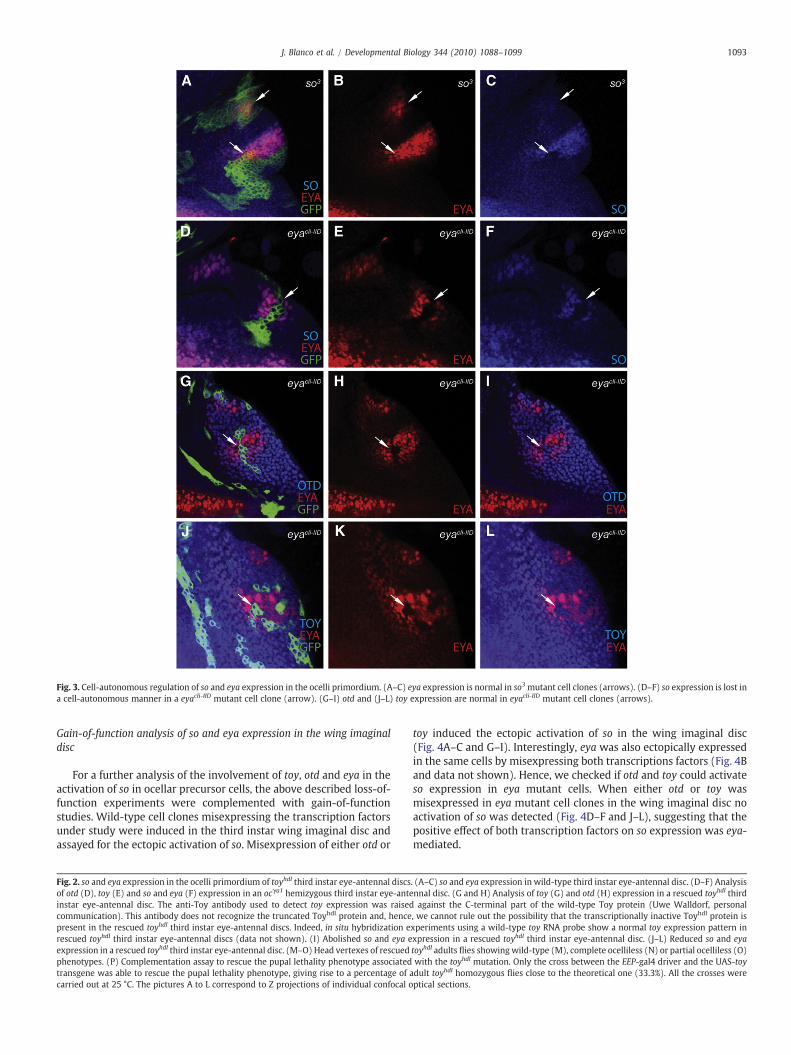
Fig. 3. Cell-autonomous regulation of so and eya expression in the ocelli primordium. (A–C) eya expression is normal in so3 mutant cell clones (arrows). (D–F) so expression is lost ina cell-autonomous manner in a eyacli-IID mutant cell clone (arrow). (G–I) otd and (J–L) toy expression are normal in eyacli-IID mutant cell clones (arrows).
1093J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
Gain-of-function analysis of so and eya expression in the wing imaginaldisc
For a further analysis of the involvement of toy, otd and eya in theactivation of so in ocellar precursor cells, the above described loss-of-function experiments were complemented with gain-of-functionstudies. Wild-type cell clones misexpressing the transcription factorsunder study were induced in the third instar wing imaginal disc andassayed for the ectopic activation of so. Misexpression of either otd or
Fig. 2. so and eya expression in the ocelli primordium of toyhdl third instar eye-antennal discsof otd (D), toy (E) and so and eya (F) expression in an ocγa1 hemizygous third instar eye-anteinstar eye-antennal disc. The anti-Toy antibody used to detect toy expression was raisedcommunication). This antibody does not recognize the truncated Toyhdl protein and, hencepresent in the rescued toyhdl third instar eye-antennal discs. Indeed, in situ hybridization erescued toyhdl third instar eye-antennal discs (data not shown). (I) Abolished so and eya eexpression in a rescued toyhdl third instar eye-antennal disc. (M–O) Head vertexes of rescuedphenotypes. (P) Complementation assay to rescue the pupal lethality phenotype associatedtransgene was able to rescue the pupal lethality phenotype, giving rise to a percentage of acarried out at 25 °C. The pictures A to L correspond to Z projections of individual confocal o
toy induced the ectopic activation of so in the wing imaginal disc(Fig. 4A–C and G–I). Interestingly, eya was also ectopically expressedin the same cells by misexpressing both transcriptions factors (Fig. 4Band data not shown). Hence, we checked if otd and toy could activateso expression in eya mutant cells. When either otd or toy wasmisexpressed in eya mutant cell clones in the wing imaginal disc noactivation of so was detected (Fig. 4D–F and J–L), suggesting that thepositive effect of both transcription factors on so expression was eya-mediated.
. (A–C) so and eya expression in wild-type third instar eye-antennal disc. (D–F) Analysisnnal disc. (G and H) Analysis of toy (G) and otd (H) expression in a rescued toyhdl thirdagainst the C-terminal part of the wild-type Toy protein (Uwe Walldorf, personal
, we cannot rule out the possibility that the transcriptionally inactive Toyhdl protein isxperiments using a wild-type toy RNA probe show a normal toy expression pattern inxpression in a rescued toyhdl third instar eye-antennal disc. (J–L) Reduced so and eyatoyhdl adults flies showingwild-type (M), complete ocelliless (N) or partial ocelliless (O)with the toyhdl mutation. Only the cross between the EEP-gal4 driver and the UAS-toydult toyhdl homozygous flies close to the theoretical one (33.3%). All the crosses wereptical sections.

Fig. 4. Gain-of-function analysis of so and eya expression in the third instar wingimaginal disc. (A–C) Misexpression of toy in the wing disc activates eya and so cell-autonomously. (D–F) In the absence of Eya transcriptional activity, misexpression of toyis not able to ectopically activate so. (G–I) Misexpression of otd is also able to ectopicallyactivate so (arrows), but not in eya mutant cells (J–L). Misexpression of ey ectopicallyactivates so both in wild-type cell clones (M–O) and in eya mutant cell clones (P–R).Misexpression of eya ectopically activates so (S–U) in an ey-independentmanner (V–X).All the pictures correspond to Z projections of individual confocal optical sections.
1094 J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
These results are consistent with previous data indicating that otdplays a positive role in the regulation of eya in ocellar precursor cells(Blanco et al., 2009). However, they do not agree with the proposedrole of toy as a direct activator of so in the ocelli primordium throughthe binding to the Toy-specific Pax6 binding sites present in the eye/ocellus-specific enhancer of the so gene (Punzo et al., 2002). As acomparison, we studied whether ey misexpression was also able toectopically activate so and whether this activation was eya-depen-dent. ey has also been characterized as a direct activator of so in thecompound eye region of the third instar eye-antennal-disc, throughthe binding to the Ey-specific Pax6 binding sites present in the eye/ocellus-specific enhancer of the so gene (Niimi et al., 1999; Punzo etal., 2002). Misexpression of ey in the wing imaginal disc was able toactivate so not only in wild-type cell clones (Fig. 4M–O), but also ineyamutant cell clones (arrows in Fig. 4P–R). These results support therole of ey as a direct regulator of so in the compound eye region of thethird instar eye-antennal disc. They also indicate that the involvementof toy in the activation of so expression in ocellar precursor cells islargely eya-mediated.
Sinceotd and toy ability to ectopically activate sowas eya-dependent,we checked if eya alone could also activate so.We inducedwild-type cellclones in the wing imaginal disc misexpressing eya and assayed for soactivation. As shown in Fig. 4S–U, expression of eya sufficed toectopically activate so in the wing imaginal disc. Feedback regulatoryloops between ey and downstream genes like eya and so have beendescribed during compound eye development (Bonini et al., 1997).Hence, we checked whether the ectopic activation of so in eyaexpressing cells was ey-mediated. However, no ectopic activation ofeywas detected in wild-type cell clones misexpressing eya (Fig. 4V–X).
Taken together, the above results place eya as the key factor in theregulation of so expression in ocellar precursor cells.
A new minimal eye/ocellus-specific enhancer of the so gene
The eye/ocellus-specific enhancer of the so gene has beenpreviously characterized in our laboratory as a DNA fragmentconsisting of two spatially separated cis-regulatory elements: a428 bp element (known as the so10 enhancer) and a 27 bp auto-regulatory element (soAE) located 334 bp downstream of so10(Fig. 5A, number 1) (Punzo et al., 2002; Pauli et al., 2005). Footprintinganalysis has identified five paired domain-specific Pax6 binding siteswithin the so10 enhancer. Three of them (sites 1, 2 and 5) are equallybound by Ey and Toy and necessary for enhancer activity in thecompound eye territory of the third instar eye-antennal disc. Sites 3and 4 are Toy-specific and required for enhancer activity in the ocelliterritory (Fig. 5A and Punzo et al., 2002). The so10 element (Fig. 5A,number 2) displays enhancer activity in the compound eye part of thethird instar eye-antennal imaginal disc (Fig. 5C), whereas the soAE isnecessary for enhancer activity in the ocelli territory. Both elements,when put together in cis, recapitulate the so expression pattern in thethird instar eye imaginal disc (Fig. 5B) and rescue the eyeless andocelliless phenotype of the so1 allele in a UAS/Gal4 based in vivo assay(Punzo et al., 2002: Pauli et al., 2005).
In order to further characterize the eye/ocellus-specific soenhancer, we compared its DNA sequence with homologoussequences present in different Drosophila species (SupplementaryFig. 1). Interestingly, only two out of the five Pax6 binding sites (sites4 and 5) seemed to be evolutionarily conserved. Accordingly, a DNAfragment containing the first three Pax6 binding sites (Fig. 5A,number 3) showed no enhancer activity in the eye-antennal discduring third instar larva (Fig. 5D). On the contrary, a 200 bp DNAfragment (named so11 enhancer; Fig. 5A, number 4) containing thePax6 binding sites 4 and 5 displayed enhancer activity comparable tothe so10 element (compare Fig. 5C and E). Furthermore, whencombined with the soAE (Fig. 5A, number 5), the so11 fragment alsoshowed enhancer activity in the ocelli territory (Fig. 5F), although not
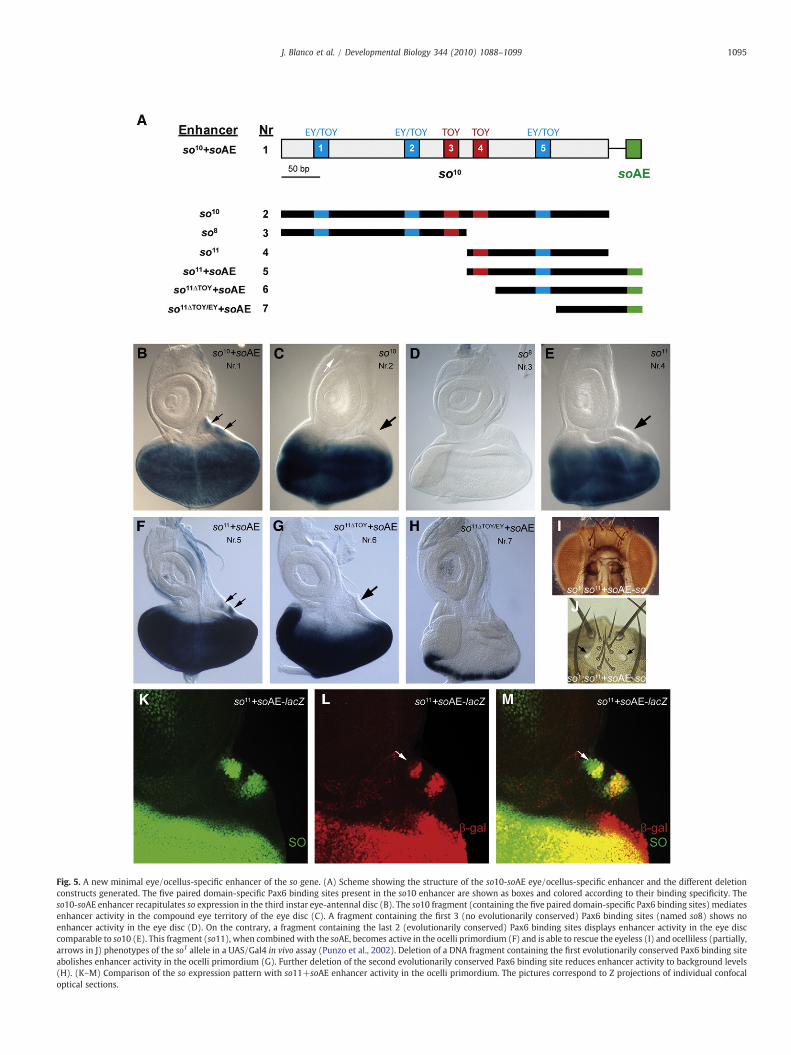
Fig. 5. A new minimal eye/ocellus-specific enhancer of the so gene. (A) Scheme showing the structure of the so10-soAE eye/ocellus-specific enhancer and the different deletionconstructs generated. The five paired domain-specific Pax6 binding sites present in the so10 enhancer are shown as boxes and colored according to their binding specificity. Theso10-soAE enhancer recapitulates so expression in the third instar eye-antennal disc (B). The so10 fragment (containing the five paired domain-specific Pax6 binding sites) mediatesenhancer activity in the compound eye territory of the eye disc (C). A fragment containing the first 3 (no evolutionarily conserved) Pax6 binding sites (named so8) shows noenhancer activity in the eye disc (D). On the contrary, a fragment containing the last 2 (evolutionarily conserved) Pax6 binding sites displays enhancer activity in the eye disccomparable to so10 (E). This fragment (so11), when combined with the soAE, becomes active in the ocelli primordium (F) and is able to rescue the eyeless (I) and ocelliless (partially,arrows in J) phenotypes of the so1 allele in a UAS/Gal4 in vivo assay (Punzo et al., 2002). Deletion of a DNA fragment containing the first evolutionarily conserved Pax6 binding siteabolishes enhancer activity in the ocelli primordium (G). Further deletion of the second evolutionarily conserved Pax6 binding site reduces enhancer activity to background levels(H). (K–M) Comparison of the so expression pattern with so11+soAE enhancer activity in the ocelli primordium. The pictures correspond to Z projections of individual confocaloptical sections.
1095J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
1096 J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
in all the so expressing cells (arrows in Fig. 5K–M). Deletion of a DNAfragment containing the first evolutionarily conserved Pax6 bindingsite (site 4) abolished enhancer activity in the ocelli primordium(Fig. 5G), but not in the compound eye primordium, suggesting thatthe remaining DNA fragment (which contains the Pax6 binding site 5.Fig. 5A, number 6) suffices for enhancer activity in the compound eyeprimordium (Fig. 5G). Further deletion of a DNA fragment containingthe second evolutionarily conserved Pax6 binding site (Fig. 5A,number 7) reduced enhancer activity to background levels (Fig. 5H).Although we cannot exclude the possibility that additional unchar-acterized regulatory sequences present in the deleted fragmentsmightaccount for or contribute to the lack of enhancer activity inthe so11ΔTOY/EY+soAE enhancer, the results agree with the previouslydone functional characterization of the so10 enhancer through thetargeted mutagenesis of the Toy-specific and Ey/Toy-specific Pax6binding sites (Punzo et al., 2002). Finally, so expression driven by thecombined so11-soAE elements was able to rescue the eyeless andpartially the ocelliless phenotype of the so1 allele (Fig. 5I and J). Insummary, we have characterized a new minimal eye/ocellus-specificenhancer of the so gene consisting of the so11 element (which containsthe two evolutionarily conserved Pax6 binding sites) and the soAE.
To functionally validate the two evolutionarily conserved Pax6binding sites present in the so11 element, we checked whether
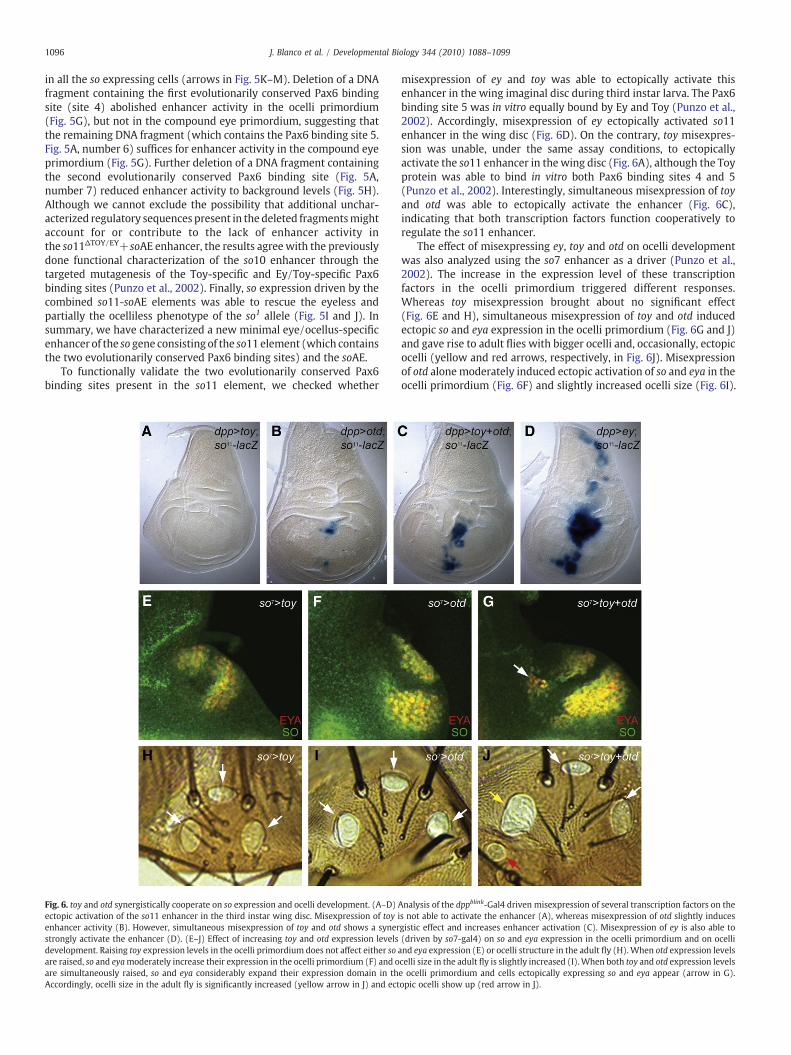
Fig. 6. toy and otd synergistically cooperate on so expression and ocelli development. (A–D)ectopic activation of the so11 enhancer in the third instar wing disc. Misexpression of toy ienhancer activity (B). However, simultaneous misexpression of toy and otd shows a synerstrongly activate the enhancer (D). (E–J) Effect of increasing toy and otd expression levelsdevelopment. Raising toy expression levels in the ocelli primordium does not affect either soare raised, so and eyamoderately increase their expression in the ocelli primordium (F) and oare simultaneously raised, so and eya considerably expand their expression domain in thAccordingly, ocelli size in the adult fly is significantly increased (yellow arrow in J) and ect
misexpression of ey and toy was able to ectopically activate thisenhancer in the wing imaginal disc during third instar larva. The Pax6binding site 5 was in vitro equally bound by Ey and Toy (Punzo et al.,2002). Accordingly, misexpression of ey ectopically activated so11enhancer in the wing disc (Fig. 6D). On the contrary, toy misexpres-sion was unable, under the same assay conditions, to ectopicallyactivate the so11 enhancer in the wing disc (Fig. 6A), although the Toyprotein was able to bind in vitro both Pax6 binding sites 4 and 5(Punzo et al., 2002). Interestingly, simultaneous misexpression of toyand otd was able to ectopically activate the enhancer (Fig. 6C),indicating that both transcription factors function cooperatively toregulate the so11 enhancer.
The effect of misexpressing ey, toy and otd on ocelli developmentwas also analyzed using the so7 enhancer as a driver (Punzo et al.,2002). The increase in the expression level of these transcriptionfactors in the ocelli primordium triggered different responses.Whereas toy misexpression brought about no significant effect(Fig. 6E and H), simultaneous misexpression of toy and otd inducedectopic so and eya expression in the ocelli primordium (Fig. 6G and J)and gave rise to adult flies with bigger ocelli and, occasionally, ectopicocelli (yellow and red arrows, respectively, in Fig. 6J). Misexpressionof otd alonemoderately induced ectopic activation of so and eya in theocelli primordium (Fig. 6F) and slightly increased ocelli size (Fig. 6I).
Analysis of the dppblink-Gal4 driven misexpression of several transcription factors on thes not able to activate the enhancer (A), whereas misexpression of otd slightly inducesgistic effect and increases enhancer activation (C). Misexpression of ey is also able to(driven by so7-gal4) on so and eya expression in the ocelli primordium and on ocelliand eya expression (E) or ocelli structure in the adult fly (H).When otd expression levelscelli size in the adult fly is slightly increased (I).When both toy and otd expression levelse ocelli primordium and cells ectopically expressing so and eya appear (arrow in G).opic ocelli show up (red arrow in J).

1097J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
Discussion
Characterization of the embryonic eye primordium-specific enhancer(EEP) of the toy gene
Our analysis of the 5′ genomic region of toy yielded theidentification of two DNA fragments containing cis-regulatorysequences active in the embryonic brain and ventral nerve cord,resembling the toy expression pattern during mid-late stages ofembryogenesis. One of the fragments (fragment 1) spans thepromoter region and was recently characterized as important forcorrect toy expression in the embryonic brain and eye primordium(Jacobsson et al., 2009). However, the reported embryonic eyeprimordium-specific enhancer activity was rather weak and whenused in a complementation assay to rescue the toyhdl mutant allele, nosignificant rescue at 25 °C was observed (Jacobsson et al., 2009).Despite of these results, we could not detect enhancer activity in theembryonic eye primordium using fragment 1. On the contrary, strongembryonic eye primordium-specific enhancer activity was detected infragment 2, a fragment not subjected to analysis by Jacobsson et al.This fragment lies 8.2 kb upstream of the toy transcription initiationsite and, when used to drive expression of a toy cDNA transgene, itefficiently rescued the pupal lethality phenotype associated with thetoyhdl mutation at 25 °C (Fig. 2P). We, therefore, designated thisfragment as embryonic eye primordium-specific enhancer (EEP) ofthe toy gene.
Apart from toy, three other transcription factors have been shownto be expressed in the eye primordium during late embryogenesis,namely ey and the two Pax6-like homologues eyg and toe, yet none ofthem has been involved in the regulation of toy expression. We haveobserved that the homeodomain containing gene otd is also expressedin this structure and, therefore, we checked whether it could beinvolved in toy regulation. Epistasis analysis supported this hypoth-esis, as toy expression and EEP enhancer activity were abolished in theeye primordium of embryos homozygous for an otd null mutant allele.Interestingly, eyg expression was not affected in this mutantbackground, supporting the view of an independent regulation ofPax6 (ey and toy) and Pax6-like (eyg and toe) homologues in theembryonic eye primordium (Jang et al., 2003). Moreover, it alsoindicated that the embryonic eye primordium was specified in an otdand toy loss-of-function mutant background. We still do not know ifthe otd-dependent regulation of toy expression is direct or indirect.Preliminary results have shown that a recombinant protein contain-ing the Otd homeodomain was able to bind to the EEP enhancer inEMSA experiments (data not shown), and putative Otd binding siteshave been found in the EEP DNA sequence, suggesting that a directregulation is conceivable.
Otd belongs to the Otd/Otx protein family of evolutionarilyconserved transcription factors, which have been proposed asancestral key players in anterior determination. The members ofthis protein family play important roles in cephalization, headsegmentation, brain patterning and photoreceptor differentiation inboth vertebrates and invertebrates and they can functionallysubstitute for each other to a great extend in crossphylum rescueexperiments (Acampora et al., 2005). In vertebrates, Otx genes havealso been shown to be required for proper differentiation of the retinalpigmented epithelium (Martinez-Morales et al., 2001) and severalOTX2 alleles are linked to congenital eye diseases in humans (Ragge etal., 2005). In the vertebrate embryo, Otx2 shows an initial widespreadexpression pattern that becomes progressively restricted to theanterior part, covering most of the forebrain and midbrain neuroe-pithelium, including the eye domain (Simeone et al., 1993).Interestingly, Drosophila otd shows a comparable expression patternduring embryogenesis. It is expressed as a cephalic gap gene duringthe syncitial blastoderm stage, covering the anteriormost part of theembryo, and later becomes restricted to the anterior neuroectoderm
from which the eye-antennal disc primordium arises by the end ofembryogenesis (Finkelstein et al., 1990; Gao et al., 1996). Here, wehave shown that otd is also expressed in the embryonic eyeprimordium and significantly contributes to the development of thisstructure through the positive regulation of toy.
toy control of so expression in the ocelli primordium
According to the current model, so expression in the ocelliprimordium is initiated by toy and subsequently maintained by apositive eya-dependent autoregulatory feedback loop (Pauli et al.,2005; Blanco et al., 2009). Whereas the involvement of so and eya inthe maintenance phase is well documented (Pauli et al., 2005), therole of toy as initiator of so expression in ocellar precursor cells has notbeen examined in detail, mainly due to the absence of eye-antennaldisc development in strong toy mutant alleles (like toyhdl). The EEPenhancer-mediated rescue of toyhdl overcomes this problem, makinggene expression analysis of third instar eye-antennal discs feasible ina toy loss-of-function mutant background.
The current model predicts that in the absence of Toy transcrip-tional activity (1) so expression should be lost and (2) eya expressionshould not be affected. On the contrary, the results presented in thispaper show that so expression was reduced in only 40% of the casesand that this reduction was always accompanied by a similarreduction in eya expression. Since eya expression in ocellar precursorcells is so-independent (Fig. 3A–C), these results indicate that toydirect or indirectly regulates eya in the ocelli primordium.
Conversely, in a toy gain-of-function situation, the model predictsthat (3) so, but not eya, should be ectopically activated. However,misexpression of toy in the third instar wing imaginal disc ectopicallyactivated not only so, but also eya (Fig. 4A–C) Interestingly, ectopicactivation of so was abolished when toy misexpression was carriedout in eya mutant cells (Fig. 4D–F). These results suggest that toyfunction as initiator of so expression in ocellar precursor cells is likelyindirect and largely eya-mediated. Indeed, eya misexpression in thewing disc sufficed for the ectopic activation of so (Fig. 4S–X). Thisresult is intriguing because Eya lacks a recognizable DNA bindingdomain and forms a complex with So that works as a transcriptionalactivator during compound eye and ocelli development (Pignoni et al.,1997). The fact that so was ectopically activated in only a smallpercentage of the wild-type clones misexpressing eya (or othertranscription factors like toy and otd) indicates that additionalelements (transcription factors and/or signaling molecules) must beinvolved in so expression.
We have recently shown that eya expression in ocellar precursorcells is indirectly controlled by otd (Blanco et al., 2009). Since otdexpression is normal in toy loss-of-function eye discs (Fig. 2H), toyinvolvement in eya expression in ocellar precursor cells is likely otd-independent. Hence, the model that arises supports the view of twoindependent regulatory pathways (one controlled by toy and theother by otd) synergistically regulating eya expression in the ocelliprimordium (Fig. 7). Eya would in turn trigger, by an unknownmechanism, so expression in ocellar precursor cells and latercontribute, together with So, to the maintenance of so expression.According to this model, eya expression levels and/or transcriptionalactivity in ocellar precursor cells would be reduced in a toy loss-of-function mutant background, but otherwise would suffice to activateso to wild-type levels in most of the cases, giving rise to normal flies.However, in a few cases, the reduction in eya expression and/ortranscriptional activity would go over a certain threshold, impairingso expression in ocellar precursor cells and giving rise to ocellilessflies. In an otd loss-of-function mutant background, the situationwould be more drastic, as wingless (a known eya and so repressor)would be ectopically activated, preventing eya and so expression inthe ocelli primordium and giving rise to ocelliless flies (Baonza and

Fig. 7.Model for the genetic control of so expression during ocelli development inDrosophila.Two independent regulatory pathways (one controlled by toy and the other by otd)synergistically regulate eya expression/activity in the ocelli primordium. eya is necessary notonly tomaintain, but also to activate, togetherwith additional factors, so expression in ocellarprecursor cells.
1098 J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
Freeman, 2002; Blanco et al., 2009; Hazelett et al., 1998; Lee andTreisman, 2001; Royet and Finkelstein, 1997).
Recently, we have isolated an ocelli-specific mutant allele of toy(toyD3.3). Homozygous toyD3.3 flies are viable, but show a very strongocelliless phenotype (stronger than the rescued toyhdl homozygousflies). Interestingly, so expression in the ocelli primordium duringthird instar larva is almost completely abolished (data not shown). Injust a few cases small patches of so expressing cells are still detected.eya expression is also affected, although to a lesser extent. Theseresults support the role of toy as a direct or indirect regulator of eyaexpression in the ocelli primordium. The molecular characterizationof this new toy allele will certainly provide an insight into themechanisms through which toy is involved in ocelli development.
A new minimal eye/ocellus-specific enhancer of so
Based on sequence conservation among several Drosophila species,we have defined a new minimal eye/ocellus-specific enhancer of theso gene (so11+soAE), which retains enhancer activity in the thirdinstar eye-antennal disc similar to the endogenous so expressionpattern. This new minimal enhancer contains just two of the fiveformerly characterized Pax6 binding sites in Drosophila melanogaster,e.g. a Toy-specific binding site (site 4) and a Toy/Ey binding site (site5). Deletion of a DNA fragment containing the Toy-specific bindingsite (Fig. 5G) or mutation of its sequence (Punzo et al., 2002)abolished enhancer activity in the ocelli primordium. Thus, ourprevious model suggested that toy alone directly regulated soexpression and, specifically, was necessary for the initiation of soexpression in the ocelli primordium (Pauli et al., 2002). However, thegenetic data presented in this paper (loss-of-function and gain-of-function analyses) suggest that toy largely exerts its action on sothrough the regulation of eya expression and/or activity in ocellarprecursor cells. Indeed, misexpression of toy in the wing imaginal discusing the dpp-gal4 driver was not able to ectopically activate the so11enhancer, whereas misexpression of ey (a well characterized directregulator of so in the compound eye primordium) succeeded in theectopic activation of this enhancer. Interestingly, simultaneous
misexpression of toy and otd also activated the so11 enhancer,supporting the view of both transcription factors cooperating in theregulation of so in ocellar precursor cells. The nature of thiscooperation is unknown. otd is necessary in the ocelli primordiumto maintain hedgehog expression (Fig. 7), which in turn activates eyain ocellar precursor cells (Blanco et al., 2009). toy might directly orindirectly contribute to eya expression in ocellar precursor cells and/or cooperate with eya in the activation of so in those cells (dashed linein Fig. 7). In this sense, Toy and Eya could form a protein complex andfunction together as a transcriptional activator of so in ocellarprecursor cells. The toyhdl allele codes for a truncated Toy protein(Toyhdl) that lacks part of the homoedomain and the C-terminal part,including the transactivation domain (Kronhamn et al., 2002). As aconsequence, Toyhdl is transcriptionally inactive, but it putativelyretains DNA binding capability through the paired domain. In theseconditions the transcriptional output of the putative Toyhdl-Eyaprotein complex (namely, so activation in ocellar precursor cells)would largely depend on the Eya transactivation domain and,therefore, would be very sensitive to the eya expression levels.Thus, lowering these levels over a certain threshold (as it occasionallyhappens in toyhdl mutants) would impair so expression in ocellarprecursor cells. Several proteins (including the retinal determinantsEy and So) have been shown to interact and putatively form proteincomplexes with Eya (Giot et al., 2003), yet Toy was not among them.Hence, further work is needed to find out if Toy and Eya are able toform a protein complex and if this complex can account for thesynergistic effect of otd and toy on so expression.
Conclusions
In summary, several conclusions can be drawn from the resultspresented in this paper: (1) eya expression in ocellar precursor cells ispositively regulated by both otd and toy. (2) The regulation of soexpression by toy in ocellar precursor cells is largely eya-mediated. (3)The Toy-specific Pax6 binding site 4 present in the minimal eye/ocellus-specific so enhancer is necessary both for enhancer activity inthe ocelli primordium during the emergence of this primordium(initiation phase) and also later during its subsequent development.(4) The binding of Toy to the Pax6 binding site 4 does not alone sufficeto trigger so expression in ocellar precursor cells, yet the contributionof additional factors (possibly Eya) is required.
Acknowledgments
We acknowledge N. Azpiazu, B. Bello, P. Callaerts, C. Desplan, F.Pignoni, I. Rebay, U. Walldorf, the Bloomington Stock Center and theDevelopmental Studies Hybridoma Bank for kindly providing flystrains and reagents. This work was supported by the Kantons ofBasel-Stadt and Basel-Land and the Swiss National Fonds.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ydbio.2010.05.494.
References
Acampora, D., Annino, A., Tuorto, F., Puelles, E., Lucchesi, W., Papalia, A., Simeone, A.,2005. Otx genes in the evolution of the vertebrate brain. Brain Res. Bull. 66,410–420.
Aldaz, S., Morata, G., Azpiazu, N., 2003. The Pax-homeobox gene eyegone is involved inthe subdivision of the thorax of Drosophila. Development 130, 4473–4482.
Baonza, A., Freeman, M., 2002. Control of Drosophila eye specification by Winglesssignalling. Development 129, 5313–5322.
Blanco, J., Seimiya, M., Pauli, T., Reichert, H., Gehring, W.J., 2009. Wingless andHedgehog signaling pathways regulate orthodenticle and eyes absent during ocellidevelopment in Drosophila. Dev. Biol. 329, 104–115.
Bonini, N.M., Leiserson, W.M., Benzer, S., 1993. The eyes absent gene: genetic control ofcell survival and differentiation in the developing Drosophila eye. Cell 72, 379–395.

1099J. Blanco et al. / Developmental Biology 344 (2010) 1088–1099
Bonini, N.M., Bui, Q.T., Gray-Board, G.L., Warrick, J.M., 1997. The Drosophila eyes absentgene directs ectopic eye formation in a pathway conserved between flies andvertebrates. Development 124, 4819–4826.
Cheyette, B.N., Green, P.J., Martin, K., Garren, H., Hartenstein, V., Zipursky, S.L., 1994. TheDrosophila sine oculis locus encodes a homeodomain-containing protein requiredfor the development of the entire visual system. Neuron 12, 996–997.
Czerny, T., Halder, G., Kloter, U., Souabni, A., Gehring, W.J., Busslinger, M., 1999. Twin ofeyeless, a second Pax-6 gene ofDrosophila, acts upstream of eyeless in the control ofeye development. Mol. Cell 3, 297–307.
Domìnguez, M., Casares, F., 2005. Organ specification-growth control connection: newin-sights from the Drosophila eye-antennal disc. Dev. Dyn. 232, 673–684.
Finkelstein, R., Smouse, D., Capaci, T.M., Spradling, A.C., Perrimon, N., 1990. Theorthodenticle gene encodes a novel homeo domain protein involved in thedevelopment of the Drosophila nervous system and ocellar visual structures. GenesDev. 4, 1516–1527.
Furukubo-Tokunaga, K., Adachi, Y., Kurusu, M., Walldorf, U., 2009. Brain patterningdefects caused bymutations of the twin of eyeless gene inDrosophilamelanogaster.Fly 3 (4), 263–269.
Gao, Q., Wang, Y., Finkelstein, R., 1996. orthodenticle regulation during embryonic headdevelopment in Drosophila. Mech. Dev. 56, 3–15.
Gehring, W.J., 2004. Historical perspective on the development and evolution of eyesand photoreceptors. Int. J. Dev. Biol. 48, 707–717.
Gehring, W.J., 2005. New perspectives on eye development and the evolution of eyesand photoreceptors. J. Hered. 96, 171–184.
Gehring, W.J., Ikeo, K., 1999. Pax-6 mastering eye morphogenesis and eye evolution.Trends Genet. 15, 371–377.
Gehring, W.J., Seimiya, M., 2010. Eye evolution and the origin of Darwin's eyeprototype. Ital. J. Zool. 77, 124–136.
Giot, L., Bader, J.S., Brouwer, C., Chaudhuri, A., Kuang, B., Li, Y., Hao, Y.L., Ooi, C.E.,Godwin, B., Vitols, E., et al., 2003. A protein interaction map of Drosophilamelanogaster. Science 302, 1727–1736.
Green, P., Hartenstein, A.Y., Hartenstein, V., 1993. The embryonic development of theDrosophila visual system. Cell Tissue Res. 273, 583–598.
Halder, G., Callaerts, P., Gehring, W.J., 1995. Induction of ectopic eyes by targetedexpression of the eyeless gene in Drosophila. Science 267, 1788–1792.
Halder, G., Callaerts, P., Flister, S., Walldorf, U., Kloter, U., Gehring, W.J., 1998. Eyelessinitiates the expression of both sine oculis and eyes absent during Drosophilacompound eye development. Development 125, 2181–2191.
Haynie, J.L., Bryant, P.J., 1986. Development of the eye-antenna imaginal disc andmorphogenesis of the adult head in Drosophila melanogaster. J. Exp. Zool. 237,293–308.
Hazelett, D.J., Bourouis, M., Walldorf, U., Treisman, J.E., 1998. Decapentaplegic andwingless are regulated by eyes absent and eyegone and interact to direct thepattern of retinal differentiation in the eye disc. Development 125, 3741–3751.
Hirth, F., Kammermeier, L., Frei, E., Walldorf, U., Noll, M., Reichert, H., 2003. Anurbilaterian origin of the tripartite brain: developmental genetic insights fromDrosophila. Development 130, 2365–2373.
Jacobsson, L., Kronhamn, J., Rasmuson-Lestander, A., 2009. The Drosophila Pax6paralogs have different functions in head development but can partially substitutefor each other. Mol. Genet. Genomics 282, 217–231.
Jang, C.C., Chao, J.L., Jones, N., Yao, L.C., Bessarab, D.A., Kuo, Y.M., Jun, S., Desplan, C.,Beckendorf, S.K., Sun, Y.H., 2003. Two Pax genes, eye gone and eyeless, actcooperatively in promoting Drosophila eye development. Development 130,2939–2951.
Jun, S., Wallen, R.V., Goriely, A., Kalionis, B., Desplan, C., 1998. Lune/eye gone, a Pax-likeprotein, uses a partial paired domain and a homeodomain for DNA recognition.Proc. Natl Acad. Sci. U. S. A. 95, 13720–13725.
Kenyon, K.L., Ranade, S.S., Curtiss, J., Mlodzik, M., Pignoni, F., 2003. Coordinatingproliferation and tissue specification to promote regional identity in the Drosophilahead. Dev. Cell 5, 403–414.
Kronhamn, J., Frei, E., Daube, M., Jiao, R., Shi, Y., Noll, M., Rasmuson-Lestander, A., 2002.Headless flies produced by mutations in the paralogous Pax6 genes eyeless andtwin of eyeless. Development 129, 1015–1026.
Kumar, J.P., 2009. The molecular circuitry governing retinal determination. Biochim.Biophys. Acta 1789, 306–314.
Lee, T., Luo, L., 1999. Mosaic analysis with a repressible cell marker for studies of genefunction in neuronal morphogenesis. Neuron 22, 451–461.
Lee, J.D., Treisman, J.E., 2001. The role of Wingless signaling in establishing theanteroposterior and dorsoventral axes of the eye disc. Development 128,1519–1529.
Li, X., Oghi, K.A., Zhang, J., Krones, A., Bush, K.T., Glass, C.K., Nigam, S.K., Aggarwal, A.K.,Maas, R., Rose, D.W., et al., 2003. Eya protein phosphatase activity regulates Six-Dach-Eya transcriptional effects in mammalian organogenesis. Nature 426,247–254.
Martinez-Morales, J.R., Signore, M., Acampora, D., Simeone, A., Bovolenta, P., 2001. Otxgenes are required for tissue specification in the developing eye. Development 128,2019–2030.
Mutsuddi, M., Chaffee, B., Cassidy, J., Silver, S.J., Tootle, T.L., Rebay, I., 2005. UsingDrosophila to decipher how mutations associated with human branchio-oto-renalsyndrome and optical defects compromise the protein tyrosine phosphatase andtranscriptional functions of eyes absent. Genetics 170, 687–695.
Niimi, T., Seimiya, M., Kloter, U., Flister, S., Gehring, W.J., 1999. Direct regulatoryinteraction of the eyeless protein with an eye-specific enhancer in the sine oculisgene during eye induction in Drosophila. Development 126, 2253–2260.
Pappu, K.S., Mardon, G., 2004. Genetic control of retinal specification and determinationin Drosophila. Int. J. Dev. Biol. 48, 913–924.
Pauli, T., Seimiya, M., Blanco, J., Gehring, W.J., 2005. Identification of functional sineoculis motifs in the autoregulatory element of its own gene, in the eyeless enhancerand in the signalling gene hedgehog. Development 132, 2771–2782.
Pignoni, F., Hu, B., Zavitz, K.H., Xiao, J., Garrity, P.A., Zipursky, S.L., 1997. The eye-specification proteins So and Eya form a complex and regulate multiple steps inDrosophila eye development. Cell 91, 881–891.
Punzo, C., Seimiya, M., Flister, S., Gehring, W.J., Plaza, S., 2002. Differential interactionsof eyeless and twin of eyeless with the sine oculis enhancer. Development 129,625–634.
Quiring, R., Walldorf, U., Kloter, U., Gehring, W.J., 1994. Homology of the eyeless gene ofDrosophila to the Small eye gene in mice and Aniridia in humans. Science 265,785–789.
Ragge, N.K., Brown, A.G., Poloschek, C.M., Lorenz, B., Henderson, R.A., Clarke, M.P.,Russell-Eggitt, I., Fielder, A., Gerrelli, D., Martinez-Barbera, J.P., et al., 2005.Heterozygous mutations of OTX2 cause severe ocular malformations. Am. J. Hum.Genet. 76, 1008–1022.
Royet, J., Finkelstein, R., 1995. Pattern formation in Drosophila head development: therole of the orthodenticle homeobox gene. Development 121, 3561–3572.
Royet, J., Finkelstein, R., 1996. hedgehog, wingless and orthodenticle specify adult headdevelopment in Drosophila. Development 122, 1849–1858.
Royet, J., Finkelstein, R., 1997. Establishing primordia in the Drosophila eye-antennalimaginal disc: the roles of decapentaplegic, wingless and hedgehog. Development124, 4793–4800.
Silver, S.J., Rebay, I., 2005. Signaling circuitries in development: insights from the retinaldetermination gene network. Development 132, 3–13.
Simeone, A., Acampora, D., Mallamaci, A., Stornaiuolo, A., D'Apice, M.R., Nigro, V.,Boncinelli, E., 1993. A vertebrate gene related to orthodenticle contains ahomeodomain of the bicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryo. EMBO J. 12, 2735–2747.
Staehling-Hampton, K., Hoffmann, F.M., 1994. Ectopic decapentaplegic in theDrosophila midgut alters the expression of five homeotic genes, dpp, and wingless,causing specific morphological defects. Dev. Biol. 164, 502–512.
Tootle, T.L., Silver, S.J., Davies, E.L., Newman, V., Latek, R.R., Mills, I.A., Selengut, J.D.,Parlikar, B.E., Rebay, I., 2003. The transcription factor Eyes absent is a proteintyrosine phosphatase. Nature 426, 299–302.
Xiong, W., Dabbouseh, N.M., Rebay, I., 2009. Interactions with the Abelson tyrosinekinase reveal compartmentalization of Eyes Absent function between nucleus andcytoplasm. Dev. Cell 16, 271–279.
Yao, J.G., Weasner, B.M., Wang, L.H., Jang, C.C., Weasner, B., Tang, C.Y., Salzer, C.L., Chen,C.H., Hay, B., Sun, Y.H., Kumar, J.P., 2008. Differential requirements for the Pax6(5a)genes eyegone and twin of eyegone during eye development in Drosophila. Dev.Biol. 315, 535–551.
Younossi-Hartenstein, A., Tepass, U., Hartenstein, V., 1993. Embryonic origin of theimaginal discs of the head of Drosophila melanogaster. Roux's Arch. Dev. Biol. 203,60–73.
Zimmerman, J.E., Bui, Q.T., Liu, H., Bonini, N.M., 2000. Molecular genetic analysis ofDrosophila eyes absent mutants reveals an eye enhancer element. Genetics 154,237–246.





























