Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 83, pp. 4389-4393, June 1986Genetics
Genetic evidence suggests that homosporous ferns with highchromosome numbers are diploid
(Pteridophyta/diploidy/polyploidy/electrophoresis/isozymes)
CHRISTOPHER H. HAUFLER* AND DOUGLAS E. SOLTISt*Department of Botany, University of Kansas, Lawrence, KS 66045; and tDepartment of Botany, Washington State University, Pullman, WA 99163
Communicated by Peter H. Raven, February 10, 1986
ABSTRACT Homosporous ferns have usually been con-sidered highly polyploid because they have high chromosomenumbers (average n = 57.05). In angiosperms, species withchromosome numbers higher than n = 14 generally have moreisozymes than those with lower numbers, consistent with theirpolyploidy. By extrapolation, homosporous ferns would beexpected to have many isozymes. However, ongoing surveysindicate that within fern genera, species having the lowestchromosome numbers have the number of isozymes consideredtypical of diploid seed plants. Only species above these lowestnumbers have additional isozymes. Therefore, homosporousferns either have gone through repeated cycles of polyploidyand gene silencing or were initiated with relatively highchromosome numbers. The latter possibility represents aradical departure from currently advocated hypotheses of fernevolution and suggests that there may be fundamental differ-ences between the genomes of homosporous ferns and those ofhigher plants. These hypotheses can be tested by genetic,karyological, and molecular techniques.
The average haploid chromosome number for homosporousferns, 57.05 (1), is so high that up to 95% of all fern specieshave been considered polyploid (2). However, since theessence of polyploidy is the number of component genomesand not the number ofchromosomes, the genetic significanceof such high polyploidy estimates may be questioned. Asreviewed by Wagner and Wagner (3), defining absolute levelsof polyploidy in homosporous ferns has never been a simplematter. While accepting that these taxa reached their extantchromosome numbers through ancient polyploid events,Wagner and Wagner also discussed the alternative hypoth-esis "that the original vascular land plants may have had highchromosome numbers" (p. 201 in ref. 3). To avoid proposingoriginal basic chromosome numbers in ferns, Manton andVida (4) developed a practical, working definition for hap-loidy in homosporous ferns as "the lowest actual gameticnumber for which there is direct evidence in any group" (p.365 in ref. 4). Vida (5) subsequently used this definition incalculating that only 43.5% of homosporous ferns arepolyploid. Because this method of determining ploidal levelssimplifies the presentation of data, we have adopted it in thepresent paper (see Table 1).Chromosomal data may be open to a wide range of
interpretation when estimating ploidy. It is also possible toestimate ploidal levels by counting the number of genesspecifying enzymes (6). In the present communication, weintroduce an independent data set from electrophoreticanalyses for addressing the question: "Are homosporousferns truly polyploid?"
Electrophoresis of constituent enzymes has been used toidentify isozymic forms (those encoded by different loci) as
discrete bands on a starch or acrylamide gel. Gottlieb (7)demonstrated that, because the genes in the contributinggenomes are usually expressed, polyploid angiosperm spe-cies (defined as those having chromosome numbers greaterthan n = 14) generally exhibit isozymic multiplicity-i.e.,they have more isozymes per enzyme than do diploids. Suchextra isozymes are most easily detected as fixed heterozygote(nonsegregating) bands in progeny arrays. However, caremust be exercised when interpreting multiple-banded en-zyme patterns because they can result from a variety ofsources. For example, extra bands that appear to representthe polyploid accumulation of loci can actually represent thecompartmentalization of enzymes within plant cells (6, 8). Inseed plants, the glycolytic enzymes typically are found as apair of isozymes, one in the chloroplasts and the other in thecytosol (8). Gastony and Darrow (9) demonstrated thatenzymes in the fern Athyrium filix-femina show a similarpattern of compartmentalization.
Determining the genetic basis of enzyme banding patternsis essential, since diploids may appear to have additionalisozymes merely because they are heterozygotes. Analysis ofmeiotic segregation patterns can be used to identify bandsthat are allelic variants (allozymes) at a single locus. Inhomosporous ferns, this analysis is relatively straightforwardbecause gametophytes are multicellular, mitotic derivativesof single, meiotically produced spores. These gametophytescan be analyzed individually by electrophoresis similar to thecase for pollen of angiosperms (10), and studies have beencarried out in several fern species (11, 12). Using these andother (13) established procedures, we examined a repre-sentative set of homosporous ferns (Table 1).
RESULTS AND DISCUSSIONAs introduced by Gastony and Darrow (9), if ferns with highchromosome numbers are polyploid, they should have manyisozymes. However, patterns of isozyme expression in alldiploid fern species surveyed are the same as those for diploidangiosperms (8), even though the average number of chro-mosomes in the two groups is remarkably different (2). Wefound only a few instances of isozyme multiplicity.Phosphoglucomutase, for example, has a duplicated locus inBommeria ehrenbergiana (14), Asplenium montanum (17),and Polypodium virginianum (16). Such occasional depar-tures from diploid expression may be considered insignifi-cant; they are also seen, for example, in diploid angiosperms.The complex banding patterns of triose-phosphate isomerasedetected in most homosporous ferns stand out as exceptionsto the general simplicity of other enzymes. In diploid seedplants, triose-phosphate isomerase typically displays twoisozymes, one in the chloroplasts and the other in the cytosol(18). Nearly all diploid species surveyed from the Polypo-diaceae (sensu lato) have an additional triose-phosphateisomerase isozyme; in Athyrium fliix-femina (9), Dryopterisexpansa (D.E.S., P. S. Soltis, and L. H. Rieseberg, unpub-lished data), and Pteridium aquilinum (P. G. Wolf, personal
4389
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 83 (1986)
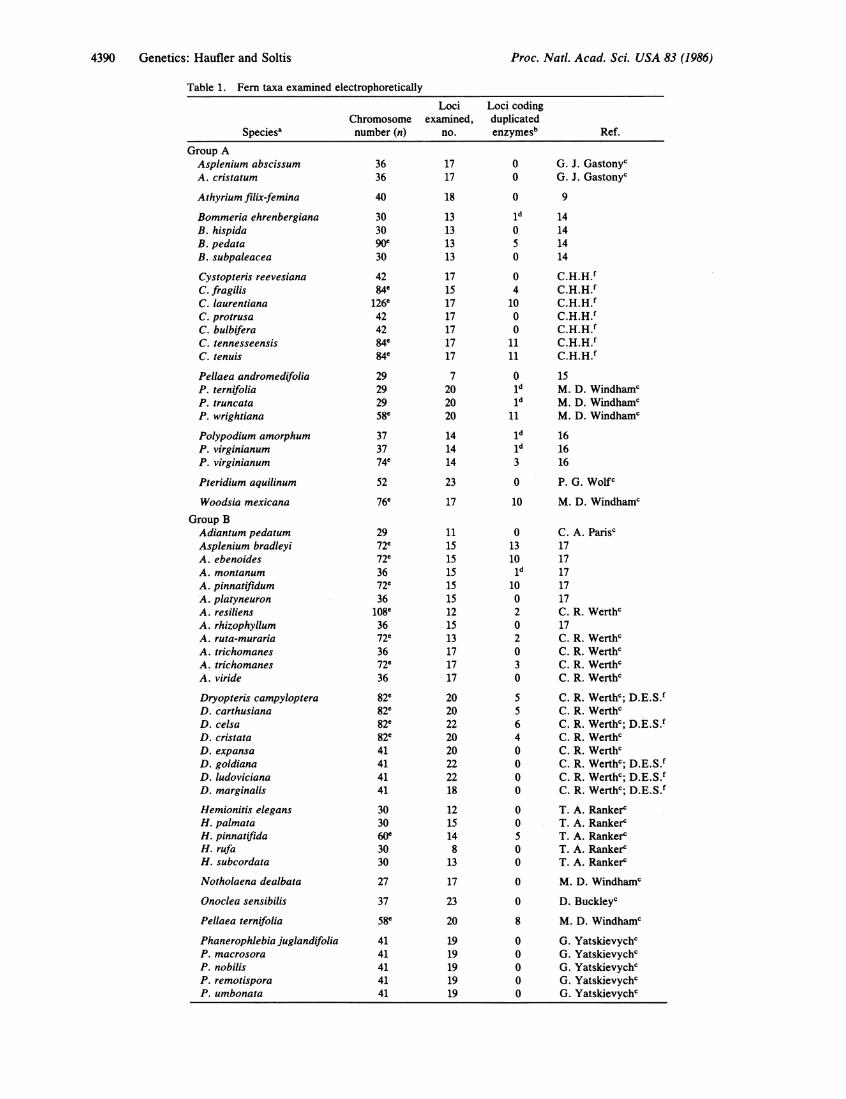
Table 1. Fern taxa examined electrophoretically
Loci Loci codingChromosome examined, duplicated
Speciesa number (n) no. enzymesb Ref.
Group AAsplenium abscissumA. cristatum
Athyrium filix-femina
Bommeria ehrenbergianaB. hispidaB. pedataB. subpaleacea
Cystopteris reevesianaC. fragilisC. laurentianaC. protrusaC. bulbiferaC. tennesseensisC. tenuis
Pellaea andromedifoliaP. ternifoliaP. truncataP. wrightiana
Polypodium amorphumP. virginianumP. virginianum
Pteridium aquilinum
Woodsia mexicana
Group BAdiantum pedatumAsplenium bradleyiA. ebenoidesA. montanumA. pinnatifidumA. platyneuronA. resiliensA. rhizophyllumA. ruta-murariaA. trichomanesA. trichomanesA. viride
Dryopteris campylopteraD. carthusianaD. celsaD. cristataD. expansaD. goldianaD. ludovicianaD. marginalis
Hemionitis elegansH. palmataH. pinnatifidaH. rufaH. subcordata
Notholaena dealbata
Onoclea sensibilis
Pellaea ternifolia
Phanerophlebia juglandifoliaP. macrosoraP. nobilisP. remotisporaP. umbonata
17 017 0
18 0
13 ld
13 013 513 0
17 015 417 1017 017 017 1117 11
7 020 1d20 ld
20 11
14 ld
14 ld
14 3
23 0
17 10
11 015 1315 1015 ld
15 1015 012 215 013 217 017 317 0
20 520 522 620 420 022 022 018 0
12 015 014 58 0
13 0
17 0
23 0
20 8
19 019 019 019 019 0
G. J. GastonycG. J. Gastonyc
9
14141414
C.H.H.fC.H.H.fC.H.H.fC.H.H.fC.H.H.fC.H.H.fC.H.H.f
15M. D. WindhamcM. D. WindhamcM. D. Windhamc
161616
P. G. Wolfc
M. D. Windhamc
C. A. Parisc1717171717C. R. Werthc17C. R. WerthcC. R. WerthcC. R. WerthcC. R. Werthc
C. R. Werthc; D.E.S.fC. R. WerthcC. R. Werthc; D.E.S.fC. R. WerthcC. R. WerthcC. R. Werthc; D.E.S.fC. R. Werthc; D.E.S.fC. R. Werthc; D.E.S.f
T. A. RankercT. A. RankercT. A. RankercT. A. RankercT. A. Rankerc
M. D. Windhamc
D. Buckleyc
M. D. Windhamc
G. YatskievychcG. YatskievychcG. YatskievychcG. YatskievychcG. Yatskievychc
3636
40
3030goe30
4284c
126e424284e84e
29292958c
373774C
52
76C
2972C72c3672e36
108e3672e3672e36
82e82c82e82C41414141
303060e3030
27
37
58e
4141414141
4390 Genetics: Haufler and Soltis

Proc. Natl. Acad. Sci. USA 83 (1986) 4391
Table 1. Continued
Loci Loci codingChromosome examined, duplicated
Speciesa number (n) no. enzymesb Ref.Polypodium hesperium 74e 11 6 M. D. Windhamc
Polystichum acrostichoides 41 19 0 G. Yatskievychc;D. S. Barringtonc
P. braunii 82' 9 3 D. S. BarringtoncP. imbricans ssp. cortum 41 19 0 G. YatskievychcP. munitum 41 19 0 G. Yatskievychc; D.E.S.,
and P. S. Soltis'P. talamancanum 82e 11 5 D. S. BarringtoncWoodsia obtusa 76e 15 4 M. D. WindhamcW. oregana 76' 15 6 M. D. Windhamc
aGroup A comprises taxa for which the genetic basis of band patterns has been confirmed by analysisof gametophytic progeny arrays. Group B comprises taxa for which the genetic basis of band patternshas been inferred from sporophytic population samples.bDuplicated loci are here defined as those that usually exhibit fixed heterozygote phenotypes-i.e.,have several bands per locus that fail to segregate in population samples or among gametophyticprogeny. Triose-phosphate isomerase appeared to have an extra isozyme in all species tested. Sinceit seems likely that this complex banding pattern resulted from either a tandem duplication orpost-translational modification, it has not been included as evidence of polyploid isozyme expression.See text for more discussion.Personal communication.dDiploid taxa that appear to have extra isozymes.ePolyploid taxa, following the definition of Manton and Vida (4).'Unpublished data.
communication), it is located in the cytosol. The bandingpattern expressed for these cytosolic enzymes appears toreflect a fixed heterozygotic genotype that could have result-ed from the polyploid addition of chromosomes. Interesting-ly, this duplication is not seen in species surveyed from theeusporangiate genus Botrychium (19).
Consideration of triose-phosphate isomerase expression inCystopteris protrusa helps to elucidate the genetic constitu-tion of the complex polypodiaceous pattern. Sporophytes ofthis species have either three rapidly migrating enzymes (Fig.la, lanes A), three slowly migrating ones (Fig. la, lanes B),or a five-banded pattern (Fig. la, lanes C). The five-bandedpattern appears additive of the two three-banded patterns,with the lower band of the rapid triad nearly overlapping theupper band of the slower set. If the three-banded patternsresulted from the polyploid addition of a second genome,then the genes should be on separate chromosomes, and fourdifferent genotypes should be seen in gametophytic progenyarrays from the five-banded, presumably heterozygoussporophyte (Fig. lb). In the more than 300 sporophytesexamined from geographically diverse population samplesand more than 150 individual gametophytic progeny, onlytwo of these genotypes are evident (Fig. ic). These findingsindicate that the apparent duplications do not segregateindependently and are not, therefore, on separate chromo-somes. The complex banding patterns may represent either atightly linked, tandem duplication or post-translational mod-ification of the gene products, but in neither case could thepattern seen in this diploid species be attributed to genomemultiplication through polyploidy.
If additional studies demonstrate that this genetic analysisof triose-phosphate isomerase is typical, then there is noconsistent evidence that diploid homosporous ferns haveadditional isozymes resulting from polyploidy. While thesedata do not fully resolve the "question of whether the highchromosome numbers of homosporous pteridophytes areprimitive or derived" (p. 202 in ref. 3), they certainlycontradict the current hypothesis that ferns have selectivelyretained genetic polyploidy to maintain variability (21).
On the other hand, extant polyploid species do exhibitincreases in isozyme number. C. tennesseensis, for example,is a putative allotetraploid resulting from polyploidy afterhybridization between two diploids, C. protrusa and C.bulbifera (22, 23). Isozymic patterns of the diploids aredistinct from each other, and those of the allotetraploid areadditive of the two putative parents (Fig. ld). Analysis ofgametophytic progeny demonstrates that these complexpatterns are fixed and, therefore, have resulted from theallopolyploid amalgamation of the diploid genomes. Similar-ly, complex isozyme patterns have been found in otherpolyploid fern species (14, 17).
CONCLUSIONSIn this paper, data from published and on-going analyses of39 diploid and 25 polyploid species are reported. In thediploids, only 6 of the 652 loci may be interpreted as havingduplicated expression that could have arisen throughpolyploidy. Among the polyploids, 162 of the 394 loci hadfixed heterozygotic enzyme phenotypes that probably result-ed from the polyploid amalgamation of divergent genomes.Thus, among extant species, the practical determination ofploidal levels by Manton and Vida (4) appears geneticallysound. A similar pattern has been reported for the fern allyEquisetum (24). Species of this genus have uniformly highchromosome numbers of 2n = 216 and, like the homosporousferns, have been considered ancient polyploids, althoughisozymically they too are diploid.Three alternative evolutionary hypotheses may be drawn
from these observations. First, the high chromosome numn-bers of extant diploid ferns could have resulted from ancient,repeated episodes of allopolyploidy. The resulting fixedheterozygotic expression typically seen in allopolyploidsmust then have been reduced to the pattern characteristic, ofdiploid species by silencing nearly all of the duplicated genes.Although no studies have been made to document genesilencing in polyploid ferns, loss of duplicate isozyme expres-sion has occurred in recently originated, polyploid cropplants (25, 26).
Genetics: Haufler and Soltis

4392 Genetics: Haufler and Soltis
Heterozygou sparent
b
Progeny
- - -
FIG. 1. (a) Zymogram showing variability among individuals fortriose-phosphate isomerase in a population of C. protrusa sporo-phytes. The lowest (most cathodal) isozyme is invariant for thisspecies. The more anodal bands occur as three rapidly migratingbands (lanes A) or as three slowly migrating bands (lanes B) or as afive-banded pattern (lanes C) that combines the two three-bandedpatterns. The most anodal bands stain faintly. (b) Diagrammaticrepresentation of expected banding patterns among gametophyticprogeny ofa five-banded C. protrusa sporophyte. These hypotheticalpatterns assume that the sporophyte is heterozygous at two loci thatsegregate independently. (c) Zymogram showing a gametophyticprogeny array from a five-banded C. protrusa sporophyte. A spo-rophyte (S) was run in the left-hand lane. All other lanes haveindividual gametophytes grown from spores of that sporophyte. Onlytwo of the four patterns diagrammed in b, progeny a and b, arepresent. (d) Zymogram of triose-phosphate isomerase expression insporophytes of the allopolyploid Cystopteris tennesseensis. Lanes:1, C. protrusa; 2-8, C. tennesseensis from a variety of geographicalareas in North America (2, Donaphan Co., KS; 3, Richardson Co.,NB; 4, Franklin Co., MO; 5, Jackson Co., IL; 6, Ottawa Co., OK;7, Pike Co., IL; 8, Carter Co., KY); 9, Cystopteris bulbifera. Thechromosome number of all individuals shown here has been verifiedby meiotic chromosome squashes. Variability in tetraploid C. tennes-seensis electromorphs parallels that in the two diploid progenitorsand indicates that the tetraploid has multiple origins (cf. ref. 20).
The second hypothesis suggests that the high chromosomenumbers in homosporous fern species may actually representthe original diploid condition. Supporting this hypothesis isthe improbability that wholesale silencing of all duplicatedloci occurred in so many different evolutionary lineages.
There certainly is no evidence of such extensive diploidiza-tion in the angiosperms (8).A third, less likely possibility is that autopolyploidy is
common in ferns and of recent origin so that isozyme lociwere duplicated without divergence. If this were true, thegenetic consequences of autopolyploidy, specifically tetra-somic inheritance, should be evident. However, all analysesof segregation in diploid species have revealed only disomicpatterns. Thus, there is good evidence that recent autopoly-ploidy cannot be implicated in explaining diploid isozymeexpression in homosporous ferns.Although data on the biochemical consequences of
polyploid evolution in plants are scanty, consideration of themore extensive fish literature can provide insights for inter-preting fern isozyme patterns. The catostomid fishes presentthe best studied example of ancient allopolyploidy (27). Inthis group, it has been suggested that the event initiating theallopolyploid lineage occurred 50 million years before pres-ent. Since that event, an average of 50% of the duplicatedgenes in different lineages have been silenced (28). Availabledata indicate that the lineages of extant homosporous ferngenera (the level at which diploidy is inferred in this paper)are at least 100 million years old (T. Delevoryas and J. Skog,personal communication). Thus, by analogy with thecatostomid fish data, there has been sufficient time to silencemost, if not all, of the duplicated genes. However, thecatostomid data also suggest that species in allopolyploidlineages should exhibit some pattern in the loss of duplicategene expression. In such fish lineages, although an average of50% of the duplicated loci have been silenced during theestimated 50 million years, the level of silencing varies amongthe species. The less modified (more primitive) species showonly about 35% silencing, while the highly derived (moreadvanced) species have lost expression in 65% of theirduplicates. As discussed above, there are occasional instanc-es of duplicate gene expression among the diploid fernssurveyed. However, these duplications are not extensiveenough to draw any phylogenetic inferences.The two most likely hypotheses may be evaluated through
a series of genetic, cytological, and molecular studies. First,detailed comparison of isozymic expression in allopolyploidsand their diploid progenitors could provide insight intowhether gene silencing has occurred. Second, critical infor-mation could be obtained through linkage analyses, since thedata discussed here predict that ferns have fewer loci perchromosome than do angiosperms. Third, meiotic andkaryotypic analyses may help to identify chromosomal ho-mologies within and between genomes. As recently reviewed(3), only univalents or bivalents are typically encountered,even in artificial hybrids between closely related pterido-phyte species. However, current studies (29) demonstratethat multivalents can be seen clearly in natural autotriploids.Further work with such taxa may help to establish genomichomologies and/or genetic control ofchromosomal pairing inpteridophytes. In addition, karyotypic studies could indicatewhether there are multiple (homoeologous) copies of chro-mosomes with similar morphologies. Although some fernkaryotypes have been described, critical evaluation demon-strates that there are no significant data to support theexistence of multiple sets of chromosomes in the ferns hereconsidered as diploids (30). In fact, Walker (31) has recentlyrefined standard karyotypic analyses for work with ferns andcomments that there are no data to support the proposal oflow base chromosome numbers in the ferns. Additionalstudies using chromosome banding techniques may generatemarkers that can determine whether there are multiple sets ofdistinctive chromosomal morphologies. Finally, by usingmolecular hybridization techniques, it may be possible toidentify silenced genes that no longer code enzyme products.Tests of the two alternative hypotheses should lead to an
Proc. Natl. Acad. Sci. USA 83 (1986)

Proc. Natl. Acad. Sci. USA 83 (1986) 4393
improved understanding of the genetic and chromosomalsystems of homosporous ferns.
We are indebted to David S. Barrington, Donald Buckley, GeraldJ. Gastony, Cathy A. Paris, Thomas A. Ranker, Pamela S. Soltis,Charles R. Werth, Michael D. Windham, Paul G. Wolf, and GeorgeYatskievych for supplying us with their unpublished results. Wethank Theodore Delevoryas and Judy Skog for offering their opinionsof the age of homosporous fern genera. Finally we are grateful toDavid Barrington, Daniel Crawford, Leslie Gottlieb, Yvonne van derMeer, Cathy Paris, Tom Ranker, Pam Soltis, and Paul Wolf forproviding valuable ideas and constructive criticisms of this manu-script. This research was supported in part by National ScienceFoundation Grant BSR85-15842 to C.H.H.
1. Klekowski, E. J. & Baker, H. G. (1966) Science 153, 305-307.2. Grant, V. (1981) Plant Speciation (Columbia Univ. Press, New
York).3. Wagner, W. H. & Wagner, F. S. (1980) in Polyploidy: Biolog-
ical Relevance, ed. Lewis, W. H. (Plenum, New York), pp.199-214.
4. Manton, I. & Vida, G. (1968) Proc. R. Soc. London Ser. B 170,361-379.
5. Vida, G. (1976) in Evolutionary Biology, eds. Novak, V. J. A.& Pacltova', B. (Czechoslovak Acad. Sci., Prague), pp.267-294.
6. Gottlieb, L. D. (1981) Proc. Natl. Acad. Sci. USA 78,3726-3729.
7. Gottlieb, L. D. (1981) Prog. Phytochem. 7, 1-46.8. Gottlieb, L. D. (1982) Science 216, 373-380.9. Gastony, G. J. & Darrow, D. C. (1983) Am. J. Bot. 70,
1409-1415.10. Weeden, N. F. & Gottlieb, L. D. (1979) Biochem. Genet. 17,
287-296.
11. Gastony, G. J. & Gottlieb, L. D. (1982) Am. J. Bot. 69, 634-637.12. Haufler, C. H. & Soltis, D. E. (1984) Am. J. Bot. 71, 878-881.13. Soltis, D. E., Haufler, C. H., Darrow, D. C. & Gastony, G. J.
(1983) Am. Fern J. 73, 9-27.14. Haufler, C. H. (1985) Syst. Bot. 10, 92-104.15. Gastony, G. J. & Gottlieb, L. D. (1985) Am. J. Bot. 72,
257-267.16. Bryan, F. A. & Soltis, D. E. (1984) Am. J. Bot. 71, 159.17. Werth, C. R., Guttman, S. I. & Eshbaugh, W. H. (1985) Syst.
Bot. 10, 184-192.18. Pichersky, E. & Gottlieb, L. D. (1984) Plant Physiol. 74,
340-347.19. Soltis, D. E. & Soltis, P. S. (1986) Am. J. Bot. 73, 588-592.20. Werth, C. R., Guttman, S. I. & Eshbaugh, W. H. (1985)
Science 228, 731-733.21. Klekowski, E. J. (1979) in The Experimental Biology ofFerns,
ed. Dyer, A. (Academic, London), pp. 133-170.22. Haufler, C. H. (1985) Proc. R. Soc. Edinburgh 86, 315-323.23. Shaver, J. M. (1954) Ferns of the Southeastern United States
with Special Reference to Tennessee (General Publishing Co.,Toronto).
24. Soltis, D. E. (1986) Am. J. Bot. 73, 908-913.25. Hart, G. E. (1983) in Isozymes: Current Topics in Biological
and Medical Research (Liss, New York), Vol. 10, pp. 365-380.26. Wilson, H. D., Barber, S. C. & Walters, T. (1983) Biochem.
Syst. Ecol. 11, 7-13.27. Ferris, S. D. (1984) in Evolutionary Genetics of Fishes, ed.
Turner, B. J. (Plenum, New York), pp. 55-93.28. Ferris, S. D. & Whitt, G. S. (1977) Nature (London) 265,
258-260.29. Haufler, C. H., Windham, M. D., Britton, D. M. & Robinson,
S. J. (1985) Can. J. Bot. 10, 1855-1863.30. Duncan, T. & Smith, A. R. (1978) Syst. Bot. 3, 105-114.31. Walker, T. G. (1985) Bull. Br. Mus. (Nat. Hist.) Bot. 13,
149-249.
Genetics: Haufler and Soltis





























