Embed Size (px)
Citation preview

of January 31, 2019.This information is current as
CC Chemokine Receptor 5 (CCR5)Gene Organization and Promoter Function for
Philip M. MurphyFlorence Guignard, Christophe Combadiere, H. Lee Tiffany and
http://www.jimmunol.org/content/160/2/9851998; 160:985-992; ;J Immunol
Referenceshttp://www.jimmunol.org/content/160/2/985.full#ref-list-1
, 17 of which you can access for free at: cites 45 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1998 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 31, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

Gene Organization and Promoter Function for CC ChemokineReceptor 5 (CCR5)1,2
Florence Guignard, Christophe Combadiere, H. Lee Tiffany, and Philip M. Murphy3
CC chemokine receptor 5 (CCR5) functions physiologically as a receptor for the leukocyte chemoattractants macrophageinflammatory protein-1a, macrophage inflammatory protein-1b, and RANTES, and functions pathologically as a key cell entrycoreceptor for HIV-1. The factors that regulate CCR5 expression may be useful therapeutic targets for HIV-1 infection. Toidentify nuclear regulatory factors, we have located and functionally characterized the CCR5 gene promoter. The gene consistsof two exons separated by a 1.9-kb intron. Exon 1 contains 43 bp of the 5*-untranslated region; exon 2 contains 11 bp of the5*-untranslated region and the complete open reading frame. Primer extension analysis identified two adjacent transcriptionalstart points (tsp) that map to the first 2 bp found in the longest known CCR5 cDNA sequence. A TATA box is present 31 bpupstream from the first tsp. CCR5 mRNA was detected constitutively in both primary human myeloid and lymphoid cells byNorthern blot hybridization. Consistent with this, transcription of a chloramphenicol acetyltransferase reporter gene wasconstitutively activated in both transiently transfected myeloid and lymphoid cell lines by the 80-bp gene fragment locatedimmediately upstream of the tsp. Deletion analysis located a strong silencer element between nucleotides 2244 and 280, anda strong enhancer element between 2486 and 2244. These results suggest that the gene region between 2486 and 21 mayregulate the expression of CCR5 in monocyte/macrophages and T lymphocytes. The Journal of Immunology, 1998, 160:985–992.
Chemokines constitute a structurally related family of se-creted proteins, most of which chemoattract and activatespecific subsets of leukocytes in vitro. Chemokines are
classified into two major subfamilies depending on the position ofthe first two of four conserved cysteines, which are adjacent in thecase of CC chemokines and separated by a single amino acid in thecase of CXC chemokines (1). In vitro, CXC chemokines attractlymphocytes and neutrophils, whereas CC chemokines typicallydo not attract neutrophils, but instead attract monocytes, macro-phages, eosinophils, basophils, dendritic cells, and lymphocyteswith variable selectivity and potency. In vivo, chemokines appearto act as locally produced emergency signals that direct leukocytesto sites of infection and tissue injury, but they may also regulatebasal leukocyte trafficking, hemopoiesis, angiogenesis, and otherprocesses (2–5).
Chemokines activate leukocytes by binding to selective, seven-transmembrane domain, G protein-coupled receptors present onthe plasma membrane (6). To date, twelve functional human che-mokine receptors have been identified: four are specific for CXCchemokines, and eight are specific for CC chemokines. Most of thereceptors identified to date bind multiple chemokines; conversely,most chemokines tested to date bind to two or more receptor sub-
types. In addition to their suspected beneficial role in host defenseand tissue repair, several chemokine receptors (e.g., CCR2B,CCR3, CCR5, and CXCR4)4 are exploited pathologically byHIV-1, acting together with CD4 as cell entry coreceptors in vitro(7–14). The HIV-1 strain specificity of the coreceptors is complex.CCR5 is used preferentially by most primary isolates and not by Tcell line-adapted laboratory strains (7–15). CXCR4 is used pref-erentially by laboratory strains and by some primary isolates.CCR3 is used by both primary isolates and laboratory-adaptedstrains (11, 12, 16, 17). Only a few strains are able to use CCR2B(12). The importance of this for HIV-1 transmission in vivo hasbeen clarified for CCR5 by the discovery of a benign, inherited,nonfunctional mutant CCR5 allele that in homozygous individualsis associated with a high level of resistance to natural HIV-1 in-fection (18–22). Consistent with this, the specific agonists forCCR5, macrophage inflammatory protein-1a, macrophage inflam-matory protein-1b, and RANTES, are able to suppress infection ofCCR5-expressing cells by appropriate HIV-1 strains (8–12, 23).Also, HIV-11 individuals heterozygous for the mutant CCR5 al-lele appear to have slightly delayed progression to AIDS comparedwith individuals homozygous for the wild-type allele (20, 21). Thisimplies that measures designed to block CCR5 expression or func-tion could be used to block HIV-1 transmission and/or to treatestablished HIV-1 infection. In this regard, detailed knowledge ofthe factors regulating CCR5 expression is an important goal.
CCR5 mRNA has been detected in PBMCs and adherent mono-cytes (8, 24). Using a specific mAb, CCR5 protein has been de-tected in microglial cells of the central nervous system and mem-ory T cells (25, 26). CCR5 protein expression can be up-regulatedby treatment of T cells with IL-2 (26). In contrast, CCR5 RNA andHIV-1 coreceptor function can be down-regulated in CD41 T cells
Laboratory of Host Defenses, National Institute of Allergy and Infectious Dis-eases, National Institutes of Health, Bethesda, MD 20892
Received for publication June 13, 1997. Accepted for publication October6, 1997.
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by a grant from the Swiss National ScienceFoundation (to F.G.).2 The sequence of CCR5 has been deposited in GenBank (accession numberAF032132).3 Address correspondence and reprint requests to Dr. Philip M. Murphy, Build-ing 10, Room 11N113, National Institutes of Health, Bethesda, MD 20892. E-mail address: [email protected]
4 Abbreviations used in this paper: CCR, CC chemokine receptor; Til cells, tu-mor-infiltrating T lymphocytes; ORF, open reading frame; UTR, untranslated re-gion; CAT, chloramphenicol acetyltransferase; tsp, transcription start point;CXCR, CXC chemokine receptor.
Copyright © 1998 by The American Association of Immunologists 0022-1767/98/$02.00
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

by CD3/CD28 costimulation (27). In the present report we haveidentified the structural organization and sequence of the CCR5gene and have located a functional promoter with high resolutionthat may be responsible for constitutive expression of CCR5 inboth myeloid and lymphoid tissues.
Materials and MethodsCell culture
Derivation of human CD41 and CD81 tumor-infiltrating T lymphocytes(Til cells) has been previously described (28, 29). The cells were providedby J. Farber (National Institute of Allergy and Infectious Diseases, Na-tional Institutes of Health, Bethesda, MD). The histiocytic lymphoma cellline U937, the acute T cell leukemia cell line Jurkat, and the human em-bryonic kidney (HEK) cell line 293 were obtained from the American TypeCulture Collection (Rockville, MD). U937 and Jurkat cells were grown inRPMI 1640 (Biofluids, Rockville, MD) supplemented with 10% heat-in-activated FBS (HyClone (Logan, UT) or Life Technologies (Gaithersburg,MD)), 2 mM glutamine, 100 U/ml penicillin, and 100mg/ml streptomycin(Quality Biologics, Gaithersburg, MD). CD4 and CD8 Til cells were grownin AIM-V medium with IL-2 (500 U/ml), glutamine, streptomycin (50mg/ml), and gentamicin (10mg/ml; Life Technologies) supplemented with10% heat-inactivated FBS. HEK 293 cells were grown in DMEM (Bioflu-ids) supplemented with 10% FBS, 4.5 g/l glucose, 2 mM glutamine, 100U/ml penicillin, and 100mg/ml streptomycin. All cells were grown at 37°Cin 5% CO2 in a humidified incubator.
Northern blot analysis of RNA
Total RNA was isolated from cultured cell lines and primary leukocytesusing a kit (Qiagen (Chatsworth, CA) or Stratagene (La Jolla, CA)). PBMCswere purified from healthy human donors by Hypaque/Ficoll density gra-dient centrifugation, dextran sedimentation, and hypotonic lysis of residualerythrocytes. Lymphocytes and monocytes were separated by adherence ofthe mononuclear layer from the Hypaque/Ficoll gradient to tissue cultureplastic in RPMI 1640 with 10% FBS at 37°C in 5% CO2 for 18 h. Lym-phocytes were recovered in the nonadherent fraction of cells. Highly pu-rified human monocytes were obtained by elutriation performed by theDepartment of Transfusion Medicine, National Institutes of Health. Iso-lated RNA (10mg/lane) was electrophoresed in a 1% agarose gel contain-ing 2% formaldehyde in 3-morpholino-propanesulfonic acid (MOPS)buffer (pH 7.0) consisting of 10 mM MOPS, 5 mM sodium acetate, and 1mM EDTA. After migration, RNA was transferred overnight by capillaryaction onto Nytran membranes (Schleicher and Schuell, Keene, NH) andUV cross-linked using a Stratalinker (Stratagene). Blots were probed withthe total open reading frame (ORF) of CCR5 labeled with [a-32P]dCTPusing a random-primed DNA labeling kit (Boehringer Mannheim, India-napolis, IN) and purified on size exclusion columns (Stratagene). The blotswere prehybridized in a buffer containing 50% formamide, 63 SSPE, 0.5%SDS, and 50mg/ml denatured salmon sperm DNA for 90 min at 37°C. Theradiolabeled probe was added (13 106 cpm/ml), and the filters were hy-bridized overnight at 37°C. The filters were then washed with 13 SSPEand 0.1% SDS at 60°C for 30 min and autoradiographed with Kodak X-OMAT AR films (Eastman Kodak, Rochester, NY) between intensifyingscreens at280°C.
RNA analysis by primer extension
Poly(A)1 RNA was purified from CD4 Til cell total RNA using thePoly(A) Quik Kit (Stratagene), and 1mg was analyzed using a commercialprimer extension kit according to the instructions of the manufacturer (Pro-mega, Madison, WI). Briefly, an antisense CCR5-specific primer (10pmol), corresponding to nucleotides 78 to 58 (59-TGGACTTGACACTTGATAATC-39) of the clone 134 cDNA encoding CCR5, reported by Rap-ort et al. (30), was end-labeled with [g-32P]ATP and annealed to thepoly(A)1 RNA at 53°C for 20 min before reverse transcription using avianmyeloblastosis virus reverse transcriptase. A control reaction was per-formed in parallel using control RNA provided in the kit. The reactionproducts were separated on a 6% acrylamide gel containing 8 M urea alongwith known DNA sequence for size determination. After electrophoresis,the gel was dried and visualized by autoradiography.
Genomic DNA analysis
CCR5 genomic clones were isolated by plaque hybridization from a com-mercially available human library in the vectorl FIX (Stratagene) usingthe 63-2 cDNA encoding a portion of the CCR5 ORF as a probe (31)labeled with [a-32P]dCTP by the random primed DNA labeling kit. Clones
containing the 59 end of the gene were identified by hybridization with two59-UTR sense primers corresponding to nucleotides 1 to 21 (59-AGAAGAGCTGAGACATCCGTT-39) and nucleotides 18 to 42 (59-CGTTCCCCTACAAGAAACTCTCCC-39) of the clone 134 CCR5 cDNA (30).The same probes were then used for mapping restriction sites and to iden-tify appropriate restriction fragments for subcloning, sequencing, and func-tional analysis. DNA sequences were analyzed with software from theUniversity of Wisconsin Genetics Computer Group on a Cray supercom-puter maintained by the National Cancer Institute Advanced ScientificComputing Laboratory, Frederick Cancer Research and Development Cen-ter (Frederick, MD) (32).
Reporter gene constructs
The reporter gene used in these studies was bacterial chloramphenicolacetyltransferase (CAT) as found in the pCAT-basic expression vector(Promega). A 2.5-kbXbaI/EcoRI fragment containing the putative 59 endof the CCR5 gene was subcloned into Bluescript KS II. Portions of thisfragment were amplified by PCR usingPfu polymerase (Stratagene) andprimers containing 21 specific nucleotides with additional 59 nucleotidesencoding eitherPstI or XbaI sites to facilitate subcloning upstream of CAT.The PCR conditions were denaturation at 94°C for 90 s, annealing at 59°Cfor 2 min, and extension at 72°C for 2 min (25 cycles). All constructs wereconfirmed by DNA sequencing on both strands. The pCAT-basic plasmid,which contains the CAT ORF without a promoter or an enhancer, was usedas a negative control. pSV40 (pCAT-Promoter, Promega), which has theSV40 promoter cloned in the sense orientation upstream of CAT, was usedas a positive control.
CAT assay
The human cell lines Jurkat and U937 were grown in suspension as de-scribed above to a density of 0.5 to 1.03 106 cells/ml, and adherent HEK293 cells were grown to subconfluence. Cells were harvested and resus-pended at a density of 303 106 cells/ml in their respective completemedium. Uncut plasmid DNA (20mg; prepared with Qiagen Maxiprep kit)was used to electroporate 153 106 cells in 500ml of complete mediumwith a 0.4-cm gap electroporation cuvette (Bio-Rad Laboratories, Hercules,CA) at 960mF and 250 V using a Gene Pulser (Bio-Rad). The cells werethen chilled on ice, added to 30 ml of complete medium, and incubated for2 days at 37°C in 5% CO2 in a humidified incubator. In addition, cells werecotransfected with 10mg of pCMV (CMV)-b-galactosidase plasmid (Clon-tech, Palo Alto, CA) as a control for electroporation efficiency. The levelof b-galactosidase activity was determined spectrophotometrically using ab-galactosidase assay kit (Promega). In some experiments, plasmid DNAwas transferred to target cells by lipofection. Specifically, 3mg of testplasmid DNA plus 2mg of pCMV-b-galactosidase plasmid DNA weremixed with 20ml of Superfect (Qiagen), according to the instructions of themanufacturer, and then added to 5 million target cells in appropriate media.After 2 days of incubation, transfected cells were harvested by centrifuga-tion at 1,800 rpm for 5 min, washed twice with PBS, and resuspended in500ml of a buffer containing 40 mM Tris (pH 7.4), 1 mM EDTA, and 150mM NaCl. The cells were incubated for 5 min at room temperature, cen-trifuged at 14,000 rpm for 1 min, resuspended in 100ml of 250 mM Tris,pH 7.8, and disrupted by freezing/thawing four times using dry ice and a37°C water bath. Cell debris was removed by a 2-min centrifugation at14,000 rpm. A portion of the supernatant containing 50mg of protein wasincubated overnight with 250 mM Tris (pH 7.5), 9 mM acetyl coenzyme A(Pharmacia Biotech, Piscataway, NJ), and 0.25mCi of [14C]chlorampheni-col (DuPont-New England Nuclear, Boston, MA) in a final volume of 150ml at 37°C. The [14C]chloramphenicol and acetylated products were thenextracted with 1 ml of ethyl acetate by vortexing for 30 s. The ethyl acetatelayer was removed, lyophilized, resuspended in 30ml of ethyl acetate, andapplied to a TLC sheet (Baker-flex silica gel 1B, J. T. Baker, Inc., Phil-ipsburg, NJ). Separation of acetylated and unacetylated forms was per-formed in a chloroform/methanol (95/5) ascending mobile phase followedby autoradiography at room temperature using a phosphor screen (Molec-ular Dynamics, Sunnyvale, CA). The radioactivity of each spot was quan-titated with a PhosphorImager (Molecular Dynamics). All cell lines weretransfected and analyzed on the same day for each independent experiment.The relative CAT activity in each lysate was quantitated by the followingequation: ((Ac/(Ac 1 Uc)/(Ab/(Ab 1 Ub)), whereA andU refer to the vol-umes of the acetylated and unacetylated forms of chloramphenicol, respec-tively, andc andb refer to CAT constructs and the pBasic control plasmid,respectively. The data were normalized for transfection efficiency based onrelative b-galactosidase activity, which varied,10% among samples ineach experiment.
986 CCR5 GENE REGULATION
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
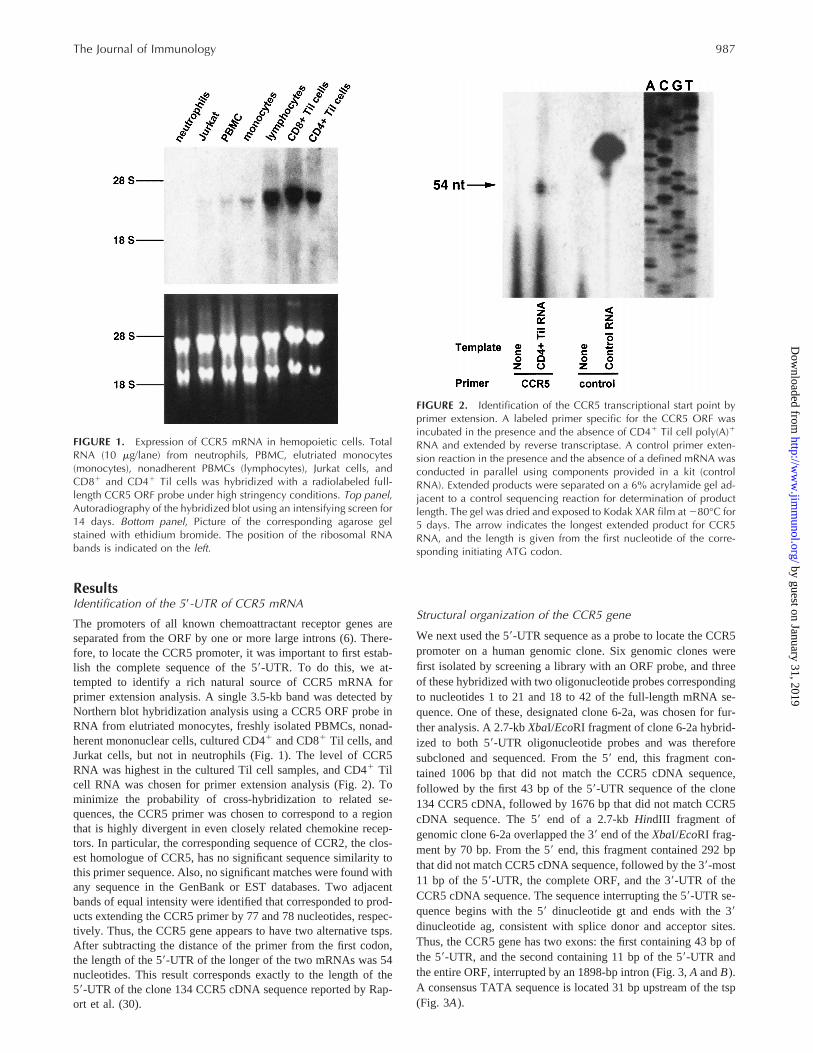
ResultsIdentification of the 59-UTR of CCR5 mRNA
The promoters of all known chemoattractant receptor genes areseparated from the ORF by one or more large introns (6). There-fore, to locate the CCR5 promoter, it was important to first estab-lish the complete sequence of the 59-UTR. To do this, we at-tempted to identify a rich natural source of CCR5 mRNA forprimer extension analysis. A single 3.5-kb band was detected byNorthern blot hybridization analysis using a CCR5 ORF probe inRNA from elutriated monocytes, freshly isolated PBMCs, nonad-herent mononuclear cells, cultured CD41 and CD81 Til cells, andJurkat cells, but not in neutrophils (Fig. 1). The level of CCR5RNA was highest in the cultured Til cell samples, and CD41 Tilcell RNA was chosen for primer extension analysis (Fig. 2). Tominimize the probability of cross-hybridization to related se-quences, the CCR5 primer was chosen to correspond to a regionthat is highly divergent in even closely related chemokine recep-tors. In particular, the corresponding sequence of CCR2, the clos-est homologue of CCR5, has no significant sequence similarity tothis primer sequence. Also, no significant matches were found withany sequence in the GenBank or EST databases. Two adjacentbands of equal intensity were identified that corresponded to prod-ucts extending the CCR5 primer by 77 and 78 nucleotides, respec-tively. Thus, the CCR5 gene appears to have two alternative tsps.After subtracting the distance of the primer from the first codon,the length of the 59-UTR of the longer of the two mRNAs was 54nucleotides. This result corresponds exactly to the length of the59-UTR of the clone 134 CCR5 cDNA sequence reported by Rap-ort et al. (30).
Structural organization of the CCR5 gene
We next used the 59-UTR sequence as a probe to locate the CCR5promoter on a human genomic clone. Six genomic clones werefirst isolated by screening a library with an ORF probe, and threeof these hybridized with two oligonucleotide probes correspondingto nucleotides 1 to 21 and 18 to 42 of the full-length mRNA se-quence. One of these, designated clone 6-2a, was chosen for fur-ther analysis. A 2.7-kbXbaI/EcoRI fragment of clone 6-2a hybrid-ized to both 59-UTR oligonucleotide probes and was thereforesubcloned and sequenced. From the 59 end, this fragment con-tained 1006 bp that did not match the CCR5 cDNA sequence,followed by the first 43 bp of the 59-UTR sequence of the clone134 CCR5 cDNA, followed by 1676 bp that did not match CCR5cDNA sequence. The 59end of a 2.7-kbHindIII fragment ofgenomic clone 6-2a overlapped the 39 end of theXbaI/EcoRI frag-ment by 70 bp. From the 59 end, this fragment contained 292 bpthat did not match CCR5 cDNA sequence, followed by the 39-most11 bp of the 59-UTR, the complete ORF, and the 39-UTR of theCCR5 cDNA sequence. The sequence interrupting the 59-UTR se-quence begins with the 59 dinucleotide gt and ends with the 39dinucleotide ag, consistent with splice donor and acceptor sites.Thus, the CCR5 gene has two exons: the first containing 43 bp ofthe 59-UTR, and the second containing 11 bp of the 59-UTR andthe entire ORF, interrupted by an 1898-bp intron (Fig. 3,A andB).A consensus TATA sequence is located 31 bp upstream of the tsp(Fig. 3A).
FIGURE 1. Expression of CCR5 mRNA in hemopoietic cells. TotalRNA (10 mg/lane) from neutrophils, PBMC, elutriated monocytes(monocytes), nonadherent PBMCs (lymphocytes), Jurkat cells, andCD81 and CD41 Til cells was hybridized with a radiolabeled full-length CCR5 ORF probe under high stringency conditions. Top panel,Autoradiography of the hybridized blot using an intensifying screen for14 days. Bottom panel, Picture of the corresponding agarose gelstained with ethidium bromide. The position of the ribosomal RNAbands is indicated on the left.
FIGURE 2. Identification of the CCR5 transcriptional start point byprimer extension. A labeled primer specific for the CCR5 ORF wasincubated in the presence and the absence of CD41 Til cell poly(A)1
RNA and extended by reverse transcriptase. A control primer exten-sion reaction in the presence and the absence of a defined mRNA wasconducted in parallel using components provided in a kit (controlRNA). Extended products were separated on a 6% acrylamide gel ad-jacent to a control sequencing reaction for determination of productlength. The gel was dried and exposed to Kodak XAR film at 280°C for5 days. The arrow indicates the longest extended product for CCR5RNA, and the length is given from the first nucleotide of the corre-sponding initiating ATG codon.
987The Journal of Immunology
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

Identification of the CCR5 promoter
This structural analysis strongly suggested that the gene regionupstream of exon 1 would contain a functional promoter for CCR5.To test this, we constructed a series of chimeric reporter genes inwhich portions of the 1006-bp fragment upstream from exon 1were subcloned upstream from the CAT ORF in the plasmidpCAT-basic. We measured CAT activity in the human lymphoidcell line Jurkat, the human myeloid cell line U937, and the humanembryonic kidney cell line 293 transiently transfected with each ofthese constructs.
The longest CCR5 gene region tested (construct 1) contained971 bp upstream from exon 1 cloned in the sense orientation up-
stream from CAT. This region stimulated reporter gene activity25- and 80-fold over that observed with the pCAT-basic vector inJurkat and U937 cells, respectively. In contrast, the same generegion tested in the reverse orientation relative to CAT (construct2) lacked activity in both cell lines (Fig. 4). Together, these resultsindicate that this 971-bp gene region contains a functional pro-moter element for this system.
To identify a minimal promoter, we tested a series of nestedfragments in the sense orientation relative to CAT containing thesame 39end as that in construct 1, but with variably truncated 59ends (constructs 3–6). Construct 6 was the shortest constructtested, only 80 bp in length. This construct retained high activity
FIGURE 3. DNA sequence and organization of the CCR5 gene. A, CCR5 gene sequence. The exon sequence is in capital letters, whereas theintron and the putative promoter region are in lower case. The first nucleotide of exon 1 was chosen as nucleotide 11. The putative TATA boxas well as several consensus transcription factor binding sites are indicated. The last three nucleotides are the first codon of the CCR5 ORF. XbaI,EcoRI, and HindIII restriction sites are also indicated. An Alu repeat in the intron is underlined. B, Genomic organization of CCR5. Shaded box,ORF; unshaded large box, transcribed but untranslated sequence; unshaded small box, Alu repeat; X, XbaI; H, HindIII; E, EcoRI. Note that the 39limit of exon 2 has not been defined. A length standard is indicated at the upper right. The sequence has been deposited in GenBank (accessionnumber AF032132).
988 CCR5 GENE REGULATION
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
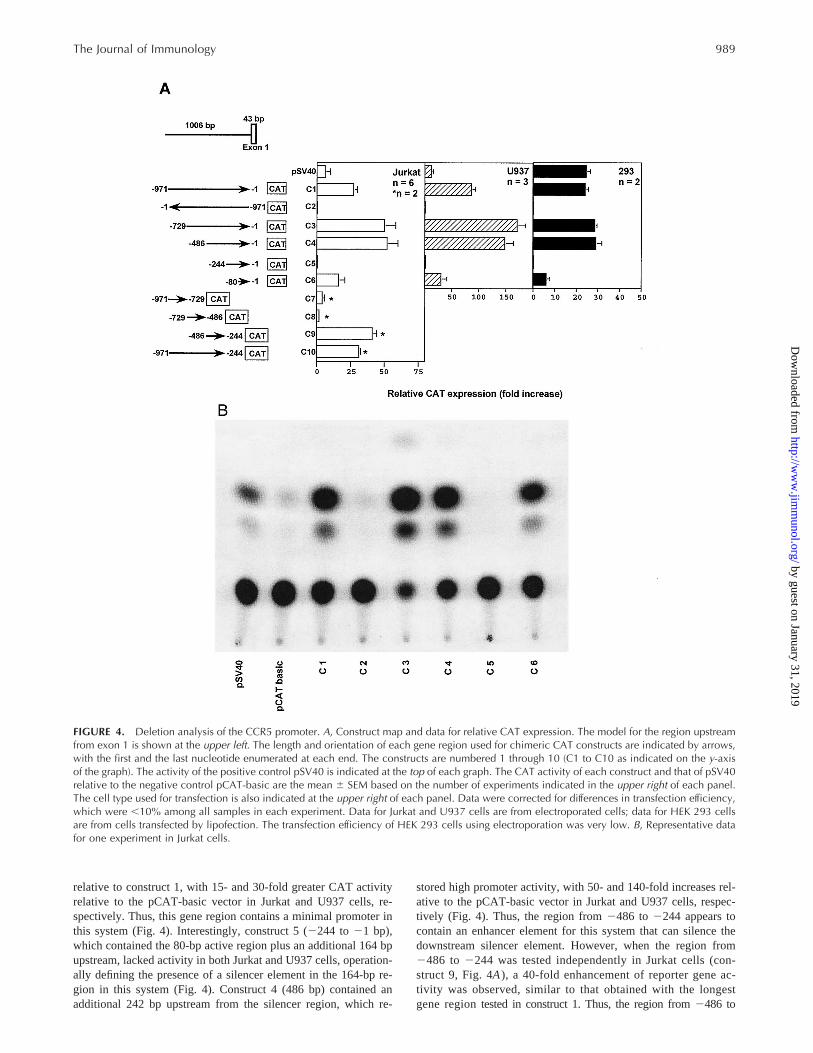
relative to construct 1, with 15- and 30-fold greater CAT activityrelative to the pCAT-basic vector in Jurkat and U937 cells, re-spectively. Thus, this gene region contains a minimal promoter inthis system (Fig. 4). Interestingly, construct 5 (2244 to21 bp),which contained the 80-bp active region plus an additional 164 bpupstream, lacked activity in both Jurkat and U937 cells, operation-ally defining the presence of a silencer element in the 164-bp re-gion in this system (Fig. 4). Construct 4 (486 bp) contained anadditional 242 bp upstream from the silencer region, which re-
stored high promoter activity, with 50- and 140-fold increases rel-ative to the pCAT-basic vector in Jurkat and U937 cells, respec-tively (Fig. 4). Thus, the region from2486 to 2244 appears tocontain an enhancer element for this system that can silence thedownstream silencer element. However, when the region from2486 to 2244 was tested independently in Jurkat cells (con-struct 9, Fig. 4A), a 40-fold enhancement of reporter gene ac-tivity was observed, similar to that obtained with the longestgene regiontested in construct 1. Thus, the region from2486 to
FIGURE 4. Deletion analysis of the CCR5 promoter. A, Construct map and data for relative CAT expression. The model for the region upstreamfrom exon 1 is shown at the upper left. The length and orientation of each gene region used for chimeric CAT constructs are indicated by arrows,with the first and the last nucleotide enumerated at each end. The constructs are numbered 1 through 10 (C1 to C10 as indicated on the y-axisof the graph). The activity of the positive control pSV40 is indicated at the top of each graph. The CAT activity of each construct and that of pSV40relative to the negative control pCAT-basic are the mean 6 SEM based on the number of experiments indicated in the upper right of each panel.The cell type used for transfection is also indicated at the upper right of each panel. Data were corrected for differences in transfection efficiency,which were ,10% among all samples in each experiment. Data for Jurkat and U937 cells are from electroporated cells; data for HEK 293 cellsare from cells transfected by lipofection. The transfection efficiency of HEK 293 cells using electroporation was very low. B, Representative datafor one experiment in Jurkat cells.
989The Journal of Immunology
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

2244 contains elements that can function independently as a pro-moter in this system.
Construct 3 (nucleotides2729 to 21 relative to the tsp) con-tains an additional 243-bp 59 of construct 4, but had activity similarto that of construct 4, suggesting that the unshared gene regionfrom 2729 to 2486 does not contain additional functional ele-ments affecting reporter gene expression. Consistent with this,when this region was tested independently (construct 8, Fig. 4A),no stimulation of CAT activity was observed. Finally, when wetested independently the 59-most 242-bp gene region upstreamfrom construct 8, we again observed no stimulation of CAT ac-tivity (construct 7 in Fig. 4A, nucleotides2971 to 2729). Con-struct 10, which tests the entire gene region tested separately inconstructs 7 to 9, had activity similar to that observed for the generegion from2486 to2244 tested separately in construct 9 (Fig.4A). Thus, the 59-most 485 bp of the parental 971-bp gene regiondo not affect the constitutive promoter activity found in region2486 to2244 or that found in region21 to 280.
The relative activity for all constructs was the same whether theDNA was delivered to Jurkat or U937 cells by electroporation(Fig. 4A) or lipofection (data not shown). In contrast, when wetested the constructs in human embryonic kidney epithelial 293cells (HEK 293 cells), a cell line lacking endogenous CCR5mRNA and CCR5 receptors, different transfection methods gavedifferent results. With electroporated HEK 293 cells, CAT-pro-moting activity significantly greater than that of pCAT-Basic wasnot observed for constructs C1 through C6. However, transfectionefficiency was very low, much lower than that for electroporatedJurkat and U937 cells, raising the possibility of false negative re-sults in this cell type (data not shown). We therefore repeated theexperiments using lipofection, which allowed high transfection ef-ficiency of the cells, as assessed byb-galactosidase activity. Sig-nificant promoter activity was then observed for constructs C1, C3,C4, and C6, but not for C2 or C5, compared with the pCAT-Basicplasmid control. This is the same pattern of activity as that ob-served for the same constructs tested in Jurkat and U937 cells.
Analysis of the CCR5 sequence
When the CCR5 sequence was compared with the GenBank da-tabase using the Blast algorithm, a completeAlu repeat was iden-tified from 11303 to11587 relative to the tsp, placing it towardthe end of the intron (Fig. 3). In the 1006-bp region upstream fromexon 1, only short stretches of limited sequence identity with otherhuman genes, including the CXCR1 and CXCR2 promoters, werefound. The CCR5 promoter region has several sequences similar toconsensus sequences for the transcription factors activating pro-tein-1, CCAAT-binding transcription factor/nuclear factor-1 (NF-1), NF-kB, NF-ATp, and IFN-stimulated response element bindingprotein (Fig. 3A).
DiscussionIn the present report we have established the structural organiza-tion of the gene for CCR5 and located its functional promoter. Likemost, but not all, G protein-coupled receptor genes, the CCR5ORF lacks introns. This is also the case for CXCR1, CXCR2,CCR1, and CCR3, as well as for receptor genes for the nonche-mokine classical chemoattractants fMet-Leu-Phe and platelet-ac-tivating factor (33–39). Exceptions to this pattern are CXCR3 andCXCR4, each of which has a large intron in the region encodingthe N-terminal segment before transmembrane domain 1 (40)(S. K. Ahuja and P. M. Murphy, unpublished observations);CCR2, which has an intron in the region encoding the C-terminalcytoplasmic tail that is used to make two receptors by alternative
splicing (41); and the C5a receptor, which has a large intron withinthe first codon (42).
Like genes for other chemokine receptors (e.g., CXCR1,CXCR2, and CCR1) and nonchemokine chemoattractant receptors(e.g., the fMet-Leu-Phe receptor and the platelet-activating factorreceptor) and unlike most other G protein-coupled receptor genes,the CCR5 gene has a large intron interrupting the 59-UTR se-quence, placing the promoter and transcription start point at a con-siderable genomic distance upstream from the translation initiationsite. CXCR2, CCR1, and the fMet-Leu-Phe receptor genes all havetwo or more introns in the 59-UTR, and alternative splicing givesrise to multiple mRNA species with the same ORF sequence butdifferent 59-UTR sequences (33, 36) (J.-L. Gao and P. M. Murphy,unpublished observations). This does not appear to be the casefor CCR5; variations in length, but not in sequence, have beenfound in the 59-UTR for the two cDNA sequences that havebeen reported (24, 30), and a single;3.5-kb band has beenconsistently identified by Northern blot analysis of CCR5-ex-pressing cells (24, 30, 43).
Although it is clear that the chemoattractant receptor ORFs havedescended from a common ancestor, a common origin for theirpromoters is not apparent from sequence comparisons. Thus, theboundaries of the replication unit for chemoattractant receptorgenes is not known. This is true even for CXCR1 and CXCR2,whose genes are clustered on human chromosome 2q35, expressedin neutrophils, and encode proteins with 78% amino acid identity,but have dissimilar promoter sequences (33, 44). It will be inter-esting to test whether this holds true for CCR5 and CCR2, whichare the most closely related known CC chemokine receptors (75%amino acid identity).
To define regions responsible for constitutive CCR5 expression,we tested the activity of a chimeric CAT reporter gene containing971 bp upstream from exon 1 in various cultured cell lines. Thisregion promoted a high level of constitutive CAT activity, but inthe sense orientation only, in both the T cell line Jurkat and thepromonocytic cell line U937. Consistent with this, we and othershave shown that endogenous CCR5 is constitutively expressed inboth primary CD41 and CD81 T cells and monocytes (24, 26, 27).Also, we have detected a low level of CCR5 mRNA in Jurkat cellsby Northern blot analysis (Fig. 1). Although this is consistent withthe promoter activity we have found, it is important to note that wedo not have evidence for expression of CCR5 protein in these cellsas assessed by staining with a polyclonal antiserum to CCR5 or bycalcium flux responses to CCR5 agonists (data not shown). It isalso important to note that we have not identified endogenousCCR5 mRNA in U937 or HEK 293 cell samples, although thesensitivity of the analysis was low (Northern blot analysis of 10mgof total RNA). Additional tissue-specific control sequences may belocated outside the gene regions we have tested that affect tissue-specific expression of endogenous CCR5.
When we analyzed a series of deletion mutants of the active971-bp region upstream from exon 1, we identified an 80-bp regionimmediately upstream from exon 1 that retained high constitutivepromoter activity in the CAT reporter gene system when tested inboth Jurkat and U937 cells. Since this region is near the tsp, con-tains a TATA sequence, and has promoter activity in the CATreporter gene system, it appears to contain a minimal promoter andmay be responsible for constitutive expression of endogenousCCR5 in myeloid and lymphoid cell types.
However, we also identified a second region, from2244 to2486 bp relative to the tsp, that also contained high constitutivepromoter activity in the CAT reporter gene system when expressedin Jurkat cells. In contrast, the upstream gene regions from2486to 2729 and from2729 to 2971 lacked independent promoter
990 CCR5 GENE REGULATION
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

activity and did not significantly affect the constitutive activity ofthe two downstream promoters. A strong suppressor element mustreside between nucleotides2244 to280, since no promoter ac-tivity was observed with a construct between2244 to21, whichcontains the 80-bp region that acts as a minimal promoter whentested independently. The activity of this silencer is overcome bythe upstream region from2244 to2486, restoring the activity tothe level found in the region from21 to 280 alone.
The transcription factors responsible for constitutive and regu-lated expression of leukocyte chemoattractant receptors have notyet been defined. The CCR5 promoter region contains several siteswith .80% identity with the consensus sequences for elementsthat bind the transcription factors CCAAT-binding transcriptionfactor/NF-1, activating protein-1, NF-kB, and NF-ATp. Also,three IFN-stimulated response elements were identified, suggest-ing a possible modulation of the promoter activity by IFN. CCR5expression has been reported to be slowly induced by IL-2 treat-ment of primary T cells and down-regulated by CD28 activation ofPBMCs. However, whether the mechanism involves transcrip-tional or post-transcriptional regulation, or both, has not yet beendetermined. To date, we have not observed changes in promoteractivity in our reporter gene system in response to the cytokinesIL-2 and IFN-g (data not shown).
The present study provides a foundation for future studies aimedat identifying protein factors and DNA sequences specifically re-sponsible for CCR5 transcription. In addition, the promoter se-quence we have described can now be used to screen cohorts ofindividuals who have been highly exposed to HIV-1 yet remainuninfected, to test whether additional inactivating CCR5 mutationsexist that could confer natural resistance to HIV-1. An analogousprecedent for this exists for another chemokine receptor known asthe Duffy Ag, which acts pathologically as an erythrocyte-specificcell entry factor for the malaria-causing protozoanPlasmodiumvivax(45). An inactivating mutation in a GATA-1 site is present inthe Duffy promoter of most Africans and is responsible for naturalresistance to vivax malaria (46). Finally, the CCR5 promoter se-quence is a potential target for gene therapy for HIV-1 throughtriplex DNA and gene-targeting strategies.
AcknowledgmentsWe thank J. Farber for generously providing the Til cell lines.
References1. Baggiolini, M., B. Dewald, and B. Moser. 1994. Interleukin-8 and related che-
motactic cytokines: CXC and CC chemokines.Adv. Immunol. 55:97.2. Broxmeyer, H. E., B. Sherry, S. Cooper, L. Lu, R. Maze, M. P. Beckmann,
A. Cerami, and P. Ralph. 1993. Comparative analysis of the human macrophageinflammatory protein family of cytokines (chemokines) on proliferation of humanmyeloid progenitor cells Interacting effects involving suppression, synergisticsuppression, and blocking of suppression.J. Immunol. 150:3448.
3. Strieter, R. M., P. J. Polverini, D. A. Arenberg, A. Walz, G. Opdenakker,J. Van Damme, and S. L. Kunkel. 1995. Role of C-X-C chemokines as regulatorsof angiogenesis in lung cancer.J. Leukocyte Biol. 57:752.
4. Springer, T. A. 1994. Traffic signals for lymphocyte recirculation and leukocyteemigration: the multistep paradigm.Cell 76:301.
5. Cook, D. N., M. A. Beck, T. M. Coffman, S. L. Kirby, J. F. Sheridan,I. B. Pragnell, and O. Smithies. 1995. Requirement of MIP-1a for an inflamma-tory response to viral infection.Science 269:1583.
6. Murphy, P. M. 1996. Chemokine receptors: structure, function and role in mi-crobial pathogenesis.Cytokine Growth Fact. Rev. 7:47.
7. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entryco-factor: functional cDNA cloning of a seven-transmembrane G-protein coupledreceptor.Science 272:872.
8. Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy,P. M. Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP-1a, MIP-1breceptor as a fusion cofactor for macrophage-tropic HIV-1.Science 272:1955.
9. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Nagashima,C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton. 1996.HIV-1 entry into CD41 cells is mediated by the chemokine receptor CC CKR-5.Nature 381:667.
10. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. DiMarzio,S. Marmon, R. E. Sutton, C. M. Hill, D. Littman, and N. R. Landau. 1996.Identification of a major co-receptor for primary isolates of HIV-1.Nature 381:661.
11. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. J. Rollins, P. D. Ponath, L. Wu,C. R. Mackay, G. LaRosa, W. Newman, N. P. Gerard, C. Gerard, and J. Sodroski.1996. The b-chemokine receptors CCR3 and CCR5 facilitate infection by pri-mary HIV isolates.Cell 85:1135.
12. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper,M. Parmentier, R. G. Collman, and R. W. Doms. 1996. A dual tropic primaryHIV-1 isolate that uses fusin and the b-chemokine receptors CKR-5, CKR-3, andCKR-2b as fusion co-factors.Cell 85:1149.
13. Bleul, C. C., M. Farzan, H. Choe, C. Parolin, I. Clark-Lewis, J. Sodroski, andT. A. Springer. 1996. The lymphocyte chemoattractant SDF-1 is a ligand forLESTR/fusin and blocks HIV-1 entry.Nature 382:829.
14. Oberlin, E., A. Amara, F. Bacherlerie, C. Bessia, J.-L. Virelizier,F. Arenzana-Seisdedos, O. Schwarta, J.-M. Heard, I. Clark-Lewis, D. F. Legler,M. Loetscher, M. Baggiolini, B. Moser. 1996. The CXC chemokine SDF-1 is theligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1.Nature 382:833.
15. Zhang, L., Y. Huang, T. He, Y. Cao, and D. D. Ho. 1996. HIV-1 subtype andsecond-receptor use.Nature 383:768.
16. Alkhatib, G., E. A. Berger, P. M. Murphy, and J. Pease. 1997. Determinants ofHIV-1 coreceptor function on CC chemokine receptor 3: importance of bothextracellular and transmembrane/cytoplasmic domains.J. Biol. Chem.272:20420.
17. Dittmar, M. T., A. McKnight, G. Simmons, P. R. Clapham, R. A. Weiss, andP. Simmonds. 1997. HIV-1 tropism and co-receptor use.Nature 385:495.
18. Liu, R., W. A. Paxton, S. Choe, D. Ceradini, S. R. Martin, R. Horuk,M. E. MacDonald, H. Stuhlman, R. A. Koup, and N. R. Landau. 1996. Homozy-gous defect in HIV-1 coreceptor accounts for resistance of some multiply-ex-posed individuals to HIV-1 infection.Cell 86:367.
19. Samson, M., F. Libert, B. J. Doranz, J. Rucker, C. Liesnard, C.-M. Farber,S. Saragosti, C. Lapoumeroulie, J. Cognaux, C. Forceille, G. Muyldermans,C. Verhofstede, G. Burtonbuy, M. Georges, T. Imai, S. Rana, Y. Yi, R. J. Smyth,R. G. Collman, R. W. Doms, G. Vassart, and M. Parmentier. 1996. Resistance toHIV-1 infection in Caucasian individuals bearing mutant alleles of the CCR-5chemokine receptor gene.Nature 382:722.
20. Dean, M., M. Carrington, C. Winkler, G. A. Huttley, M. W. Smith, R. Allikmets,J. J. Goedert, S. P. Buchbinder, E. Vittinghoff, E. Gomperts, S. Donfield,D. Vlahov, R. Kaslow, A. Saah, C. Rinaldo, R. Detels, Hemophilia Growth andDevelopment Study, Multicenter AIDS Cohort Study, Multicenter HemophiliaCohort Study, San Francisco City Cohort, ALIVE, and S. J. O’Brien. 1996. Ge-netic restriction of HIV-1 infection and progression to AIDS by a deletion alleleof the CKR5 structural gene.Science 273:1856.
21. Zimmerman, P. A., A. Buckler-White, G. Alkhatib, T. Spalding, J. Kubofcik,C. Combadiere, D. Weissman, O. Cohen, A. Rubbert, G. Lam, M. Vaccarezza,P. E. Kennedy, V. Kumaraswami, J. V. Giorgi, R. Detels, J. Hunter, M. Chopek,E. A. Berger, A. S. Fauci, T. B. Nutman, and P. M. Murphy. 1997. Inheritedresistance to HIV-1 conferred by an inactivating mutation in CC chemokinereceptor 5: studies in populations with contrasting clinical phenotypes, definedracial background and quantified risk.Mol. Med. 3:23.
22. Huang, Y., W. A. Paxton, S. M. Wolinsky, A. U. Neumann, L. Zhang, T. He,S. Kang, D. Ceradini, Z. Jin, K. Yazdanbakhsh, K. Kunstman, D. Erickson,E. Dragon, N. R. Landau, J. Phair, D. D. Ho, and R. A. Koup. 1996. The role ofa mutant CCR5 allele in HIV-1 transmission and disease progression.Nat. Med.2:1240.
23. Cocchi, F., A. L. DeVico, A. Garzino-Demo, S. K. Arya, R. C. Gallo, andP. Lusso. 1995. Identification of RANTES, MIP-1a, and MIP-1b as the majorHIV-suppressive factors produced by CD81 T cells.Science 270:1811.
24. Combadiere, C., S. K. Ahuja, H. L. Tiffany, and P. M. Murphy. 1996. Cloningand functional expression of CC CKR5, a human monocyte CC chemokine re-ceptor selective for MIP-1a, MIP-1b, and RANTES.J. Leukocyte Biol. 60:147.
25. He, J., Y. Chen, M. Farzan, H. Choe, A. Ohagen, S. Gartner, J. Susciglio,X. Yang, W. Hofmann, W. Newman, C. R. Mackay, J. Sodroski, and D. Gabuzda.1997. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia.Nature385:645.
26. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay. 1997. TheHIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated onhuman T lymphocytes.Proc. Natl. Acad. Sci. USA 94:1925.
27. Carroll, R. G., J. L. Riley, B. L. Levine, Y. Feng, S. Kaushal, D. W. Ritchey,W. Bernstein, O. S. Weislow, C. R. Brown, E. A. Berger, C. H. June, andD. C. St. Louis. 1997. Differential regulation of HIV-1 fusion cofactor expressionby CD28 costimulation of CD41 T cells.Science 276:273.
28. Yannelli, J. R. 1991. The preparation of effector cells for use in the adoptivecellular immunotherapy of human cancer.J. Immunol. Methods 139:1.
29. Liao, F., R. L. Rabin, J. R. Yannelli, L. G. Koniaris, P. Vanguri, and J. M. Farber.1995. Human Mig chemokine: biochemical and functional characterization.J. Exp. Med. 182:1301.
30. Raport, C. J., J. Gosling, V. L. Schweickart, P. W. Gray, and I. F. Charo. 1996.Molecular cloning and functional characterization of a novel human CC chemo-kine receptor (CCR5) for RANTES, MIP-1b, and MIP-1a. J. Biol. Chem. 271:17161.
31. Combadiere, C., S. K. Ahuja, and P. M. Murphy. 1995. Cloning, chromosomallocalization and RNA expression of a novel human b chemokine receptor-likegene.DNA Cell Biol. 14:673.
991The Journal of Immunology
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

32. Devereux, J., P. Haeberli, and O. Smithies. 1984. A comprehensive set of se-quence analysis programs for the VAX.Nucleic Acids Res. 12:389.
33. Ahuja, S. K., A. Shetty, H. L. Tiffany, and P. M. Murphy. 1994. Comparison ofthe genomic organization and promoter function for human interleukin-8 recep-tors A and B.J. Biol. Chem. 269:26381.
34. Lloyd, A. R., A. Biragyn, J. A. Johnston, D. D. Taub, L. Xu, D. Michiel,H. Sprenger, J. J. Oppenheim, and D. J. Kelvin. 1995. Granulocyte-colony stim-ulating factor and lipopolysaccharide regulate the expression of interleukin 8receptors on polymorphonuclear leukocytes.J. Biol. Chem. 270:28188.
35. Sprenger, H., A. R. Lloyd, R. G. Meyer, J. A. Johnston, and D. J. Kelvin. 1994.Genomic structure, characterization, and identification of the promoter of thehuman IL-8 receptor A gene.J. Immunol. 153:2524.
36. Murphy, P. M., H. L. Tiffany, D. McDermott, and S. K. Ahuja. 1993. Sequenceand organization of the humanN-formyl peptide receptor-encoding gene.Gene133:285.
37. Raport, C. J., V. L. Schweickart, D. Chantry, R. L. Eddy, Jr., T. B. Shows,R. Godiska, and P. W. Gray. 1996. New members of the chemokine receptor genefamily. J. Leukocyte Biol. 59:18.
38. Gao, J.-L., D. B. Kuhns, H. L. Tiffany, D. McDermott, X. Li, U. Francke, andP. M. Murphy. 1993. Structure and functional expression of the human macro-phage inflammatory protein-1a/RANTES receptor.J. Exp. Med. 177:1421.
39. Mutoh, H., H. Bito, M. Minami, M. Nakamura, Z. Honda, T. Izumi, R. Nakata,Y. Kurachi, A. Terano, and T. Shimizu. 1993. Two different promoters directexpression of two distinct forms of mRNAs of human platelet-activating factorreceptor.FEBS Lett. 322:129.
40. Marchese, A., M. Heiber, T. Nguyen, H. H. Q. Heng, V. R. Saldivia, R. Cheng,P. M. Murphy, L.-C. Tsui, X. Shi, P. Gregor, S. R. George, B. F. O’Dowd, andJ. M. Docherty. 1995. Cloning and chromosomal mapping of three novel genes,GPR9, GPR10, and GPR14, encoding receptors related to interleukin 8, neu-ropeptide Y, and somatostatin receptors.Genomics 29:335.
41. Charo, I., S. J. Myers, A. Herman, C. Franci, A. J. Connolly, and S. R. Coughlin.1994. Molecular cloning and functional expression of two monocyte chemoat-tractant protein 1 receptors reveals alternative splicing of the carboxyl-terminaltails. Proc. Natl. Acad. Sci. USA 91:2752.
42. Gerard, N. P., L. Bao, X.-P. He, R. L. Eddy, Jr., T. B. Shows, and C. Gerard.1993. Human chemotactic receptor genes cluster at 19q13.3/13.4: characteriza-tion of the human C5a receptor gene.Biochemistry 32:1243.
43. Samson, M., O. Labbe, C. Mollereau, G. Vassart, and M. Parmentier. 1996.Molecular cloning and functional expression of a new human CC chemokinereceptor gene.Biochemistry 35:3363.
44. Ahuja, S. K., T. Ozcelik, A. Milatovich, U. Francke, and P. M. Murphy. 1992.Molecular evolution of the interleukin-8 receptor gene cluster.Nat. Genet. 2:31.
45. Horuk, R., C. Chitnis, W. Darbonne, T. Colby, A. Rybicki, T. Hadley, andL. Miller. 1993. The erythrocyte chemokine receptor is a receptor for the malarialparasitePlasmodium vivax.Science 261:1182.
46. Tournamille, C., Y. Colin, J. P. Cartron, and C. Le Van Kim. 1995. Disruption ofa GATA motif in the Duffy gene promoter abolishes erythroid gene expression inDuffy-negative individuals.Nat. Genet. 10:224.
992 CCR5 GENE REGULATION
by guest on January 31, 2019http://w
ww
.jimm
unol.org/D
ownloaded from





























