Embed Size (px)
Citation preview

Comparative genomics of the methionine metabolismin Gram-positive bacteria a variety of regulatorysystemsDmitry A Rodionov12 Alexey G Vitreschak1 Andrey A Mironov23 and
Mikhail S Gelfand123
1Institute for Information Transmission Problems Russian Academy of Sciences Bolshoi Karetny pereulok 19Moscow 127994 Russia 2State Scientific Center GosNIIGenetika Moscow 117545 Russia and 3Department ofBioengineering and Bioinformatics Moscow State University Moscow 119992 Russia
Received February 9 2004 Revised March 22 2004 Accepted June 2 2004
ABSTRACT
Regulation of the methionine biosynthesis and trans-port genes in bacteria is rather diverse and involvestwo RNA-level regulatory systems and at least threeDNA-level systems In particular the methioninemetabolism in Gram-positive bacteria was known tobe controlled by the S-box and T-box mechanismsboth acting on the level of premature termination oftranscription Using comparative analysis of genesoperons and regulatory elements we described themethionine metabolic pathway and the methionineregulons in available genomes of Gram-positivebacteria A large number of methionine-specificRNA elements were identified S-boxes were shownto be widely distributed in Bacillales and Clostridiawhereas methionine-specific T-boxes occurredmostly in Lactobacillales A candidate binding signal(MET-box) for a hypothetical methionine regulatorpossibly MtaR was identified in Streptococcaceaethe only family in the BacillusClostridium group ofGram-positive bacteria having neither S-boxes normethionine-specific T-boxes Positional analysis ofmethionine-specific regulatory sites complementedby genome context analysis lead to identification ofnew members of the methionine regulon bothenzymes and transporters and reconstructionof the methionine metabolism in various bacterialgenomes In particular we found candidate trans-porters for methionine (MetT) and methylthioribose(MtnABC) as well as new enzymes forming the S-adenosylmethionine recycling pathway Methioninebiosynthetic enzymes in various bacterial speciesare quite variable In particular Oceanobacillusiheyensis possibly uses a homolog of the betainendashhomocysteine methyltransferase bhmT gene fromvertebrates to substitute missing bacterial-typemethionine synthases
INTRODUCTION
Sulfur-containing amino acid methionine is synthesizedde novo by most microorganisms and plants after the initialsteps of inorganic sulfate assimilation and synthesis ofcysteine or homocysteine (1) The fact that methionine isthe universal N-terminal amino acid of proteins as well asthe use of its derivative S-adenosylmethionine (SAM) in avariety of methyltransferase reactions argue for the impor-tance of methionine in the cellular metabolism There aretwo alternative pathways of methionine synthesis in micro-organisms The transsulfuration pathway of Escherichia coliinvolves cystathionine as an intermediate and utilizes cysteineas the sulfur source (2) In contrast the direct sulfhydrylationpathway found in Saccharomyces cerevisiae (3) Leptospirameyeri (4) and Corynebacterium glutamicum (5) bypassescystathionine and uses inorganic sulfur instead While mostmicroorganisms synthesize methionine via either one of thesepathways Cglutamicum utilizes both pathways (6)
Biosynthesis of methionine starts at homoserine which isthe common precursor for amino acids of the aspartate familyisoleucine threonine and methionine (Figure 1) Homoserineis derived from aspartate semialdehyde by the hom geneproduct Acylation of homoserine is catalyzed by homoserineacetyltransferase MetB in Bacillus subtilis (7) and byhomoserine succinyltransferase MetA in Ecoli (8) andthese two enzymes are homologous Lmeyeri uses homo-serine acetyltransferase MetX not related to the aboveenzymes (9) In the transsulfuration pathway homocysteineis formed from O-acetylhomoserine or O-succinylhomoserineand cysteine in two steps catalyzed by cystathionineg-synthase MetI and cystathionine b-lyase MetC in Bsubtilis(10) In the alternative pathway O-acetylhomoserine isdirectly converted to homocysteine by O-acetylhomoserinesulfhydrylase MetY utilizing sulfide as the sulfur donor (11)
Unlike the transsulfurationsulfhydrylation enzymes thatare present only in organisms with de novo methioninesynthesis methionine synthase is required by all organismsto ensure regeneration of the methyl group of SAM (1) Twotypes of methionine synthases can perform this function inEcoli Reaction catalyzed by B12-dependent protein MetH
To whom correspondence should be addressed at Institute for Information Transmission Problems Russian Academy of Sciences Bolshoi Karetny pereulok 19Moscow 127994 Russia Tel +7 095 3150156 Fax +7 095 1356080 Email rodionovgenetikaru
Nucleic Acids Research Vol 32 No 11 ordf Oxford University Press 2004 all rights reserved
3340ndash3353 Nucleic Acids Research 2004 Vol 32 No 11DOI 101093nargkh659
Published online June 23 2004
with coenzyme B12 as a cofactor is more than 100-fold fasterthan the reaction catalyzed by B12-independent isoenzymeMetE (8) In enterobacteria the methyl group of methionineis donated by methyl-tetrahydrofolate (methyl-THF) Thelatter is formed by reduction of methylene-THF in a reactioncatalyzed by the metF gene product Bsubtilis has only theB12-independent methionine synthase (formerly named MetC)and an ortholog of MetF encoded by yitJ that contains anextra N-terminal domain highly similar to the homocysteine-binding domain of MetH (12)
Ecoli is able to uptake methionine by a high-affinity trans-port system encoded by the metD locus (8) Recently it wasshown that this methionine transporter and its orthologs invarious bacteria (yusCBA in Bsubtlis renamed metNPQ) con-stitute a new family of the ABC superfamily (13ndash16) No othermethionine transporters have been identified in bacteria so far
S-adenosylmethionine synthase encoded by the metK geneis responsible for formation of SAM from methionine andATP SAM is essential for a large number of methylationprocesses and is also used for modification of rRNA nucleo-tides and polyamine synthesis (17) The main product of thetransmethylation reaction is S-adenosylhomocysteine (SAH)In Ecoli this molecule seems to be recycled to homocysteinein two steps a nucleosidase encoded by the pfs gene (mtn inBsubtilis) hydrolyzes SAH into adenine and S-ribosylhomo-cysteine which is then cleaved by a specific hydrolase to formhomocysteine (1819) Homocysteine can then be metabolizedto synthesize cysteine and methionine Notably some
organisms are able to synthesize cysteine from homocysteinevia the reverse transsulfuration pathway (320) The cystathio-nine b-synthase and g-lyase activities required for thispathway are possibly encoded by the yrhA and yrhB genesin Bsubtilis (1221) Methylthioadenosine is produced bythe polyamine synthesis pathway and then hydrolyzed by anucleosidase (encoded by mtn in Bsubtilis) yieldingmethylthioribose (19) The latter is efficiently recycled inBsubtilis via the methionine salvage pathway encoded bythe mtnKSUVWXYZ gene cluster (21ndash23)
Not methionine but SAM a major constituent of theintermediary metabolism is the regulatory molecule for themethionine biosynthesis both in Ecoli and BsubtilisHowever the mechanisms of SAM-dependent regula-tion differ in these bacteria SAM-responsive repressorMetJ binds to tandemly repeated MET box sequences(50-AGACGTCT-30) and represses transcription of most metgenes in Ecoli (8) In Bsubtilis these genes are regulated byattenuation of transcription using a highly conserved regula-tory leader sequence the S-box (2425) SAM directlyand specifically binds to this RNA structural element causingformation of the downstream terminator hairpin andsubsequent premature termination of transcription (26ndash28)In the absence of effector molecules formation of a moreenergetically favorable antiterminator which is alternativeboth to the S-box domain and the terminator structureleads to transcription read-through The S-box domainand the terminator hairpin fold independently while the
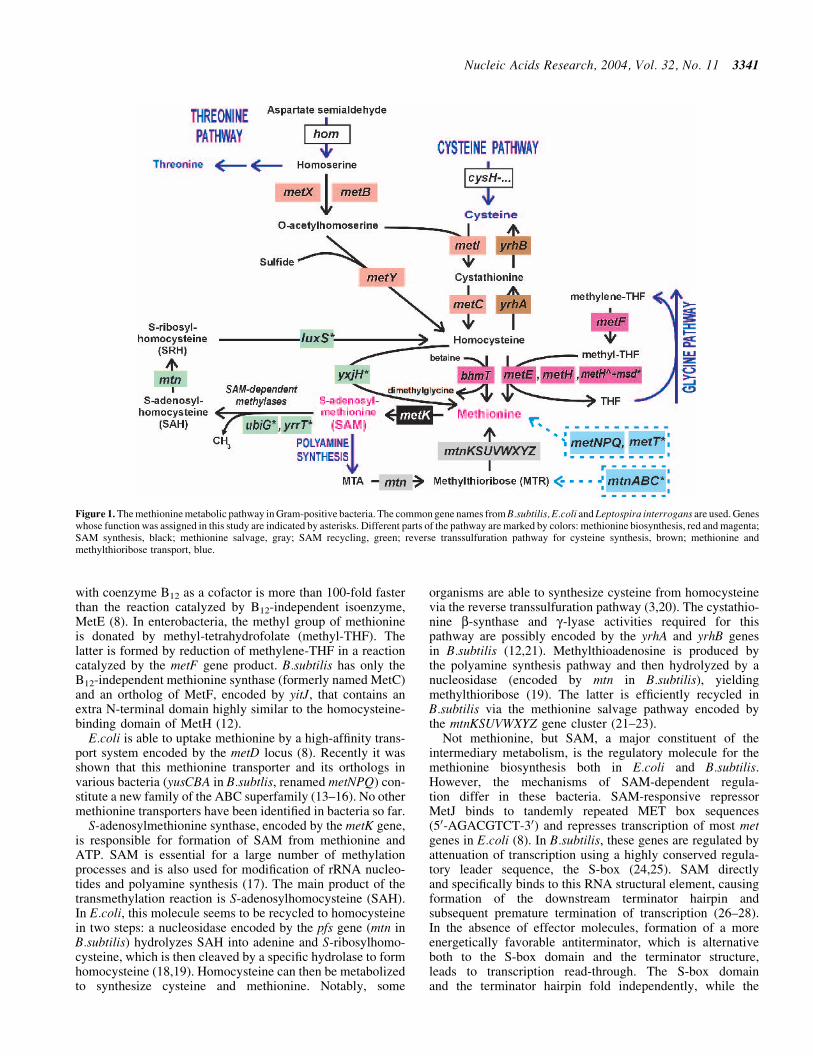
Figure 1 The methionine metabolic pathway in Gram-positive bacteria The common gene names from Bsubtilis Ecoli and Leptospira interrogans are used Geneswhose function was assigned in this study are indicated by asterisks Different parts of the pathway are marked by colors methionine biosynthesis red and magentaSAM synthesis black methionine salvage gray SAM recycling green reverse transsulfuration pathway for cysteine synthesis brown methionine andmethylthioribose transport blue
Nucleic Acids Research 2004 Vol 32 No 11 3341
antiterminator structure consists of several conserved helicesof the S-box and a new stemndashloop which is alternative to theterminator and the base stem of the S-box Moreover Gram-positive bacteria have another control system involvingpremature transcription termination (T-box) which regulatesexpression of various aminoacyl-tRNA synthetases and genesinvolved in the amino acid biosynthesis (252930) The T-boxsequence is able to bind uncharged tRNA promoting forma-tion of the antiterminator The major role in regulation isplayed by the T-box lsquospecifier codonrsquo which interacts withthe anticodon of an uncharged tRNA As the position of thisregulatory codon in the T-box structure is fixed one can pre-dict the amino acid specificity of the regulatory signal (31) Forexample the presence of a T-box with an isoleucine specifiercodon upstream of ileS in Thermoanaerobacter tengcongensisargues against a recently published theory that this gene ismis-annotated and encodes a methionyl-tRNA synthase (23)
Comparative genomics is a powerful technique for recon-struction of metabolic pathways and their DNA- or RNA-levelregulation in bacteria (32ndash37) We analyzed the methioninepathway and its regulation in various Gram-positive bacteriaWe extended the S-box and methionine-specific T-box regu-lons that appear to be widely distributed in bacilli clostridiaand lactobacilli identified a new regulatory signal for methio-nine biosynthetic and transport genes in streptococci recon-structed the likely evolutionary scenario for methionineregulons of Gram-positive bacteria and described the possiblemechanism of dual regulation of the reverse transsulfurationpathway by cysteine and SAM in Clostridium acetobutylicumvia RNA regulatory structures and antisense RNA Afterreconstruction of the methionine metabolic pathways and reg-ulatory interactions in Gram-positive bacteria we identifiedseveral new enzymes and transporters involved in the methio-nine metabolism In particular we identified a set of candidategenes forming the complete SAM-recycling pathway
MATERIALS AND METHODS
Complete and partial sequences of bacterial genomes weredownloaded from GenBank (38) Preliminary sequence datawere obtained also from the websites of The Institutefor Genomic Research (httpwwwtigrorg) University ofOklahomarsquos Advanced Center for Genome Technology(httpwwwgenomeouedu) the Sanger Centre (httpwww sangeracuk) the DOE Joint Genome Institute (httpwww jgidoegov) and the ERGO Database IntegratedGenomics Inc (39)
The conserved secondary structure of the S-box and T-boxleaders was derived using the RNAMultAln program(A A Mironov in preparation) This heuristic program simul-taneously creates a multiple alignment and a conserved sec-ondary structure for a set of RNA sequences using positionalrelationship of conserved sequence boxes and paired regionsof candidate helices This program is not based on the energyminimization but rather uses simple heuristic rules of basepairing and analysis of complementary substitutions TheRNA-PATTERN program (40) was used to search for newS-boxes and methionine-specific T-boxes in bacterial gen-omes The input RNA pattern described the RNA secondarystructure and the sequence consensus motifs as a set of the
following parameters the number of helices the length ofeach helix the loop lengths and description of the topologyof helix pairs The latter is defined by coordinates of helicesFor instance two helices may be either independent or embed-ded helices or else they could form a pseudoknot structureThis definition is similar to an approach implemented in thePalingol algorithm (41) Free energy of the S-box structureswere calculated using Zukerrsquos algorithm of free energy mini-mization (4243) implemented in the Mfold program (httpbioinfomathrpiedu~mfoldrna) RNA secondary structureof the antiterminatorantisequestor conformation which in-cludes parts of the S-box and overlaps with the terminatorsequester hairpin (Figure 7A) was predicted using Mfold withthe input including the S-box sequence extended downstreamas far as the center of the terminatorsequestor hairpin Thisstructure is more energetically favorable than S-box in theabsence of ligand To model the effect of ligand bindingthat stabilizes the S-box formation of the terminatorsequesterhairpins was modeled with input sequence starting immedi-ately downstream of S-box
A simple iterative procedure implemented in the programSignalX was used for construction of the MET-box profilefrom a set of upstream gene fragments (44) Weak palin-dromes were selected in each region Each palindrome wascompared to all other palindromes and the palindromes mostsimilar to the initial one were used to make a profile Thepositional nucleotide weights in this profile were defined as
W bketh THORN = log N bketh THORN + 05frac12 025X
ifrac14ACGT
log N iketh THORN + 05frac12
where N(b k) was the count of nucleotide b in position k (45)The candidate site score was defined as the sum of the respect-ive positional nucleotide weights
Z b1 bLeth THORN =X
Kfrac141 L
W bk keth THORN
where k was the length of the site Z-score can be used to assessthe significance of an individual site
These profiles are used to scan the set of palindromes againand the procedure was iterated until convergence Thus a set ofprofiles is constructed The quality of a profile was defined asits information content (46)
I =X
Kfrac141 L
X
ifrac14ACGT
f i keth THORNlog f i keth THORN=025frac12
where f(i k) is the frequency of nucleotide i in position k ofsites generating the profile The best profile is used as therecognition rule
Each genome was scanned with the profile and genes withcandidate regulatory sites in the upstream regions (in positions325 to +25 relative to the translation start) were selected Thethreshold for the site search was defined as the lowest scoreobserved in the training set
Protein similarity search was done using the SmithndashWaterman algorithm implemented in the GenomeExplorerprogram (47) Orthologous proteins were initially definedby the best bidirectional hit criterion (48) and if necessaryconfirmed by construction of phylogenetic trees for the
3342 Nucleic Acids Research 2004 Vol 32 No 11
corresponding protein families The phylogenetic trees of themethionine biosynthesis and transport proteins were con-structed by the maximum likelihood method implementedin PHYLIP (49) Multiple sequence alignments were doneusing CLUSTALX (50) Transmembrane segments were pre-dicted using TMpred (httpwwwchembnetorgsoftwareTMPRED_formhtml) The COG (48) InterPro (51) andPFAM (52) databases were used to verify the protein func-tional and structural annotation
Alignments of S-box and T-box sequences mentioned in thispaper as well as sequences of proteins whose functions wereassigned here are available as supplementary Figures 1 2 and 3respectively
RESULTS
Reconstruction of methionine regulons
Initially orthologs of known methionine biosynthetic andtransport genes (MET) were identified by similarity searchin the genomes of all available Gram-positive bacteria(Table 1) For further analysis positional clusters (includingpossible operons) of the MET genes were also described in thistable The hom gene for homoserine dehydrogenase shared bythe methionine and threonine biosynthesis was considered onlyif it was co-localized or co-regulated with other MET genes
S-box regulon Then we constructed the pattern of the S-boxmotif using the training set of 18 S-box leader regions (24) andscanned available genomic sequences using the RNA-PAT-TERN program Multiple alignment of 100 S-box leaders from23 bacterial genomes confirms high degree of conservation ofthe S-box primary and secondary structure (see supplementaryFigure 1) Similar to other metabolite-binding RNA elements(32) the S-box motif has a set of helices (P2 to P5) closed by asingle base stem (P1) and numerous highly conserved regions(Figure 2) In fact the regions of sequence conservation covermost of the S-box sequence and therefore may be involved inSAM binding or tertiary interactions Only two stemndashloops atthe ends of the fourth and fifth helices are not conserved on thesequence level and have variable length The end loop of helixP3 (50-CnGG-30) possibly forms a pseudoknot structure withthe interior loop between helices P4 and P5 (50-CCnG-30) Thispossible pseudoknot interaction is confirmed by several com-pensatory substitutions and could be required for the formationof stable S-box tertiary structure For instance the S-box struc-ture on the Bacillus cereus mtnW gene contains a possiblepseudoknot (50-CGAG-30 50-CTCG-30) with complementarysubstitutions in the conserved positions of both arms
Among Gram-positive bacteria the S-box motif is widelydistributed in the orders Bacillales and Clostridia but it hasnot been found in Lactobacillales including EnterococcusStreptococcus and Lactococcus species (Table 1) Positionalanalysis of genes possessing this regulatory motif has showedthat the S-box regulon in Gram-positive bacteria contains mostgenesof themethioninebiosynthesis and transport aswellas theSAM synthase gene metK The S-box regulon is most extensivein three bacilli Bsubtilis Bcereus and Oiheyensis where itcontains 11 16 and 13 regulatory elements respectively andincludes additional genes for the cysteine biosynthesis andmethionine salvage pathways as well as hypothetical genesThe detailed phylogenetic and positional analysis of the
S-box-regulated genes in Gram-positive bacteria is given inthe next section
The S-box motif is not restricted to the BacillusClostridiumgroup of bacteria In two actinobacteria Streptomyces coeli-color and Thermobifida fusca S-box precedes the hypotheticalgene SCD95A26 In addition single S-box motif was foundupstream of the metY-metX operon in the genomes of Chlor-obium tepidum Chloroflexus aurantiacus and Cytophagahutchisonii as well as upstream of the metNPQ operon encod-ing methionine transport system in Petrotoga miotherma Amore relaxed search in (28) identified candidate S-boxes alsoin the genomes of Fusobacterium nucleatum (upstream of themetK and metN genes) Deinococcus radiodurans (metH andmetN) Xanthomonas campestris (metX) and Geobacter sul-furreducens (metB and metX) All other bacterial taxonomicgroups with available genomic sequences seem to lack theS-box regulatory system
Recent experiments (26ndash28) demonstrated that SAM-dependent regulation of the methionine biosynthesis genesof Bsubtilis involves attenuation of transcription using for-mation of alternative secondary structures in the S-box regionHere we tested whether the same regulatory mechanism couldoperate for all found S-boxes Downstream of all S-boxes fromthe BacillusClostridium group of bacteria we identifiedadditional hairpins that are followed by runs of thymidinesand therefore are candidate terminators of transcriptionIn addition we observed complementary RNA regions thatpartially overlap both the proposed terminator and the basestem (helix P1) of the S-box Thus the same termination-antitermination S-box mechanism possibly operates in allGram-positive bacteria
Analysis of the upstream regions of the S-box-regulatedgenes from two actinobacteria reveals another possiblemode of regulation In this case the S-box motif directly over-laps the ribosome-binding site of the SCD95A26 gene pos-sibly acting as a sequestor We predict that in these bacteriaSAM-stabilized S-box structure directly represses initiation oftranslation Similar mode of regulation was previously sug-gested for other metabolite-responsive riboswitches in actino-bacteria (32)
Methionine-specific T-box regulon Using a training set ofexperimentally known T-box structures from Bsubtilis andother Gram-positive bacteria (31) we constructed a patternfor the T-box motif and scanned bacterial genomes using theRNA-PATTERN program The pattern was absolutely specificsince candidate T-boxes were mostly found upstream of aminoacid metabolism-related genes (data not shown) Then weselected 38 T-box sequences with AUG (methionine) specifiercodons (see supplementary Figure 2 for multiple alignment)Although the T-box system is widely used in bacteria from theBacillusClostridium group (where more than 200T-boxeswere identified A G Vitreschak unpublished results) methio-nine-specific T-boxes occur only in a limited number of species
In the genomes of Bacillus halodurans and six clostridiathere is only one methionine-specific T-box that precedes themetS gene encoding methionyl-tRNA synthetase (not includedin Table 1) The methionine T-box regulation seems tobe extensively used only in the Lactobacillales group(30 Met-T-boxes) where it exclusively controls methioninebiosynthesis and transport genes (Table 1) This phylogenetic
Nucleic Acids Research 2004 Vol 32 No 11 3343
Ta
ble
1
Met
hio
nin
ebio
synth
esis
and
tran
sport
gen
esan
dm
ethio
nin
e-sp
ecif
icre
gula
tory
elem
ents
inG
ram
-posi
tive
bac
teri
a
Gen
eso
fth
eu
pp
erp
arto
fth
em
eth
ion
ine
pat
hw
ayin
vo
lved
inth
eh
om
ocy
stei
ne
syn
thes
isar
esh
ow
nin
red
met
hio
nin
esy
nth
ases
and
met
hy
len
e-T
HF
red
uct
ases
in
mag
enta
Gen
esen
codin
gtr
ansp
ort
pro
tein
sar
esh
ow
nin
blu
eG
enes
from
the
SA
Mre
cycl
ing
and
rever
setr
anss
ulf
ura
tion
pat
hw
ays
are
ingre
enan
dbro
wnr
espec
tivel
yG
enes
form
ing
one
candid
ate
op
ero
n(w
ith
spac
erle
ssth
anlt1
00
bp
)ar
ese
par
ated
by
das
hes
(lsquo
rsquo)D
irec
tio
no
ftr
ansc
rip
tio
nin
div
erg
on
sis
sho
wn
by
ang
leb
rack
ets
Pre
dic
ted
S-b
ox
es(lsquo
Srsquo)
met
hio
nin
e-sp
ecif
icT
-bo
xes
(lsquoT
rsquo)M
ET
-bo
xes
(lsquoM
rsquo)an
dB
12-e
lem
ents
(lsquoB
rsquo)ar
esh
ow
nin
yel
low
gre
enr
eda
nd
blu
ere
spec
tivel
yC
yst
eine-
spec
ific
T-b
ox
(lsquoT
crsquo)
isal
sosh
ow
nin
gre
enC
onti
gen
ds
are
mar
ked
by
squar
ebra
cket
sG
enom
eab
bre
via
tions
are
giv
enin
colu
mn
lsquoAB
rsquow
ith
un
fin
ish
edg
eno
mes
mar
ked
by
ah
ash
3344 Nucleic Acids Research 2004 Vol 32 No 11
distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353
with coenzyme B12 as a cofactor is more than 100-fold fasterthan the reaction catalyzed by B12-independent isoenzymeMetE (8) In enterobacteria the methyl group of methionineis donated by methyl-tetrahydrofolate (methyl-THF) Thelatter is formed by reduction of methylene-THF in a reactioncatalyzed by the metF gene product Bsubtilis has only theB12-independent methionine synthase (formerly named MetC)and an ortholog of MetF encoded by yitJ that contains anextra N-terminal domain highly similar to the homocysteine-binding domain of MetH (12)
Ecoli is able to uptake methionine by a high-affinity trans-port system encoded by the metD locus (8) Recently it wasshown that this methionine transporter and its orthologs invarious bacteria (yusCBA in Bsubtlis renamed metNPQ) con-stitute a new family of the ABC superfamily (13ndash16) No othermethionine transporters have been identified in bacteria so far
S-adenosylmethionine synthase encoded by the metK geneis responsible for formation of SAM from methionine andATP SAM is essential for a large number of methylationprocesses and is also used for modification of rRNA nucleo-tides and polyamine synthesis (17) The main product of thetransmethylation reaction is S-adenosylhomocysteine (SAH)In Ecoli this molecule seems to be recycled to homocysteinein two steps a nucleosidase encoded by the pfs gene (mtn inBsubtilis) hydrolyzes SAH into adenine and S-ribosylhomo-cysteine which is then cleaved by a specific hydrolase to formhomocysteine (1819) Homocysteine can then be metabolizedto synthesize cysteine and methionine Notably some
organisms are able to synthesize cysteine from homocysteinevia the reverse transsulfuration pathway (320) The cystathio-nine b-synthase and g-lyase activities required for thispathway are possibly encoded by the yrhA and yrhB genesin Bsubtilis (1221) Methylthioadenosine is produced bythe polyamine synthesis pathway and then hydrolyzed by anucleosidase (encoded by mtn in Bsubtilis) yieldingmethylthioribose (19) The latter is efficiently recycled inBsubtilis via the methionine salvage pathway encoded bythe mtnKSUVWXYZ gene cluster (21ndash23)
Not methionine but SAM a major constituent of theintermediary metabolism is the regulatory molecule for themethionine biosynthesis both in Ecoli and BsubtilisHowever the mechanisms of SAM-dependent regula-tion differ in these bacteria SAM-responsive repressorMetJ binds to tandemly repeated MET box sequences(50-AGACGTCT-30) and represses transcription of most metgenes in Ecoli (8) In Bsubtilis these genes are regulated byattenuation of transcription using a highly conserved regula-tory leader sequence the S-box (2425) SAM directlyand specifically binds to this RNA structural element causingformation of the downstream terminator hairpin andsubsequent premature termination of transcription (26ndash28)In the absence of effector molecules formation of a moreenergetically favorable antiterminator which is alternativeboth to the S-box domain and the terminator structureleads to transcription read-through The S-box domainand the terminator hairpin fold independently while the
Figure 1 The methionine metabolic pathway in Gram-positive bacteria The common gene names from Bsubtilis Ecoli and Leptospira interrogans are used Geneswhose function was assigned in this study are indicated by asterisks Different parts of the pathway are marked by colors methionine biosynthesis red and magentaSAM synthesis black methionine salvage gray SAM recycling green reverse transsulfuration pathway for cysteine synthesis brown methionine andmethylthioribose transport blue
Nucleic Acids Research 2004 Vol 32 No 11 3341
antiterminator structure consists of several conserved helicesof the S-box and a new stemndashloop which is alternative to theterminator and the base stem of the S-box Moreover Gram-positive bacteria have another control system involvingpremature transcription termination (T-box) which regulatesexpression of various aminoacyl-tRNA synthetases and genesinvolved in the amino acid biosynthesis (252930) The T-boxsequence is able to bind uncharged tRNA promoting forma-tion of the antiterminator The major role in regulation isplayed by the T-box lsquospecifier codonrsquo which interacts withthe anticodon of an uncharged tRNA As the position of thisregulatory codon in the T-box structure is fixed one can pre-dict the amino acid specificity of the regulatory signal (31) Forexample the presence of a T-box with an isoleucine specifiercodon upstream of ileS in Thermoanaerobacter tengcongensisargues against a recently published theory that this gene ismis-annotated and encodes a methionyl-tRNA synthase (23)
Comparative genomics is a powerful technique for recon-struction of metabolic pathways and their DNA- or RNA-levelregulation in bacteria (32ndash37) We analyzed the methioninepathway and its regulation in various Gram-positive bacteriaWe extended the S-box and methionine-specific T-box regu-lons that appear to be widely distributed in bacilli clostridiaand lactobacilli identified a new regulatory signal for methio-nine biosynthetic and transport genes in streptococci recon-structed the likely evolutionary scenario for methionineregulons of Gram-positive bacteria and described the possiblemechanism of dual regulation of the reverse transsulfurationpathway by cysteine and SAM in Clostridium acetobutylicumvia RNA regulatory structures and antisense RNA Afterreconstruction of the methionine metabolic pathways and reg-ulatory interactions in Gram-positive bacteria we identifiedseveral new enzymes and transporters involved in the methio-nine metabolism In particular we identified a set of candidategenes forming the complete SAM-recycling pathway
MATERIALS AND METHODS
Complete and partial sequences of bacterial genomes weredownloaded from GenBank (38) Preliminary sequence datawere obtained also from the websites of The Institutefor Genomic Research (httpwwwtigrorg) University ofOklahomarsquos Advanced Center for Genome Technology(httpwwwgenomeouedu) the Sanger Centre (httpwww sangeracuk) the DOE Joint Genome Institute (httpwww jgidoegov) and the ERGO Database IntegratedGenomics Inc (39)
The conserved secondary structure of the S-box and T-boxleaders was derived using the RNAMultAln program(A A Mironov in preparation) This heuristic program simul-taneously creates a multiple alignment and a conserved sec-ondary structure for a set of RNA sequences using positionalrelationship of conserved sequence boxes and paired regionsof candidate helices This program is not based on the energyminimization but rather uses simple heuristic rules of basepairing and analysis of complementary substitutions TheRNA-PATTERN program (40) was used to search for newS-boxes and methionine-specific T-boxes in bacterial gen-omes The input RNA pattern described the RNA secondarystructure and the sequence consensus motifs as a set of the
following parameters the number of helices the length ofeach helix the loop lengths and description of the topologyof helix pairs The latter is defined by coordinates of helicesFor instance two helices may be either independent or embed-ded helices or else they could form a pseudoknot structureThis definition is similar to an approach implemented in thePalingol algorithm (41) Free energy of the S-box structureswere calculated using Zukerrsquos algorithm of free energy mini-mization (4243) implemented in the Mfold program (httpbioinfomathrpiedu~mfoldrna) RNA secondary structureof the antiterminatorantisequestor conformation which in-cludes parts of the S-box and overlaps with the terminatorsequester hairpin (Figure 7A) was predicted using Mfold withthe input including the S-box sequence extended downstreamas far as the center of the terminatorsequestor hairpin Thisstructure is more energetically favorable than S-box in theabsence of ligand To model the effect of ligand bindingthat stabilizes the S-box formation of the terminatorsequesterhairpins was modeled with input sequence starting immedi-ately downstream of S-box
A simple iterative procedure implemented in the programSignalX was used for construction of the MET-box profilefrom a set of upstream gene fragments (44) Weak palin-dromes were selected in each region Each palindrome wascompared to all other palindromes and the palindromes mostsimilar to the initial one were used to make a profile Thepositional nucleotide weights in this profile were defined as
W bketh THORN = log N bketh THORN + 05frac12 025X
ifrac14ACGT
log N iketh THORN + 05frac12
where N(b k) was the count of nucleotide b in position k (45)The candidate site score was defined as the sum of the respect-ive positional nucleotide weights
Z b1 bLeth THORN =X
Kfrac141 L
W bk keth THORN
where k was the length of the site Z-score can be used to assessthe significance of an individual site
These profiles are used to scan the set of palindromes againand the procedure was iterated until convergence Thus a set ofprofiles is constructed The quality of a profile was defined asits information content (46)
I =X
Kfrac141 L
X
ifrac14ACGT
f i keth THORNlog f i keth THORN=025frac12
where f(i k) is the frequency of nucleotide i in position k ofsites generating the profile The best profile is used as therecognition rule
Each genome was scanned with the profile and genes withcandidate regulatory sites in the upstream regions (in positions325 to +25 relative to the translation start) were selected Thethreshold for the site search was defined as the lowest scoreobserved in the training set
Protein similarity search was done using the SmithndashWaterman algorithm implemented in the GenomeExplorerprogram (47) Orthologous proteins were initially definedby the best bidirectional hit criterion (48) and if necessaryconfirmed by construction of phylogenetic trees for the
3342 Nucleic Acids Research 2004 Vol 32 No 11
corresponding protein families The phylogenetic trees of themethionine biosynthesis and transport proteins were con-structed by the maximum likelihood method implementedin PHYLIP (49) Multiple sequence alignments were doneusing CLUSTALX (50) Transmembrane segments were pre-dicted using TMpred (httpwwwchembnetorgsoftwareTMPRED_formhtml) The COG (48) InterPro (51) andPFAM (52) databases were used to verify the protein func-tional and structural annotation
Alignments of S-box and T-box sequences mentioned in thispaper as well as sequences of proteins whose functions wereassigned here are available as supplementary Figures 1 2 and 3respectively
RESULTS
Reconstruction of methionine regulons
Initially orthologs of known methionine biosynthetic andtransport genes (MET) were identified by similarity searchin the genomes of all available Gram-positive bacteria(Table 1) For further analysis positional clusters (includingpossible operons) of the MET genes were also described in thistable The hom gene for homoserine dehydrogenase shared bythe methionine and threonine biosynthesis was considered onlyif it was co-localized or co-regulated with other MET genes
S-box regulon Then we constructed the pattern of the S-boxmotif using the training set of 18 S-box leader regions (24) andscanned available genomic sequences using the RNA-PAT-TERN program Multiple alignment of 100 S-box leaders from23 bacterial genomes confirms high degree of conservation ofthe S-box primary and secondary structure (see supplementaryFigure 1) Similar to other metabolite-binding RNA elements(32) the S-box motif has a set of helices (P2 to P5) closed by asingle base stem (P1) and numerous highly conserved regions(Figure 2) In fact the regions of sequence conservation covermost of the S-box sequence and therefore may be involved inSAM binding or tertiary interactions Only two stemndashloops atthe ends of the fourth and fifth helices are not conserved on thesequence level and have variable length The end loop of helixP3 (50-CnGG-30) possibly forms a pseudoknot structure withthe interior loop between helices P4 and P5 (50-CCnG-30) Thispossible pseudoknot interaction is confirmed by several com-pensatory substitutions and could be required for the formationof stable S-box tertiary structure For instance the S-box struc-ture on the Bacillus cereus mtnW gene contains a possiblepseudoknot (50-CGAG-30 50-CTCG-30) with complementarysubstitutions in the conserved positions of both arms
Among Gram-positive bacteria the S-box motif is widelydistributed in the orders Bacillales and Clostridia but it hasnot been found in Lactobacillales including EnterococcusStreptococcus and Lactococcus species (Table 1) Positionalanalysis of genes possessing this regulatory motif has showedthat the S-box regulon in Gram-positive bacteria contains mostgenesof themethioninebiosynthesis and transport aswellas theSAM synthase gene metK The S-box regulon is most extensivein three bacilli Bsubtilis Bcereus and Oiheyensis where itcontains 11 16 and 13 regulatory elements respectively andincludes additional genes for the cysteine biosynthesis andmethionine salvage pathways as well as hypothetical genesThe detailed phylogenetic and positional analysis of the
S-box-regulated genes in Gram-positive bacteria is given inthe next section
The S-box motif is not restricted to the BacillusClostridiumgroup of bacteria In two actinobacteria Streptomyces coeli-color and Thermobifida fusca S-box precedes the hypotheticalgene SCD95A26 In addition single S-box motif was foundupstream of the metY-metX operon in the genomes of Chlor-obium tepidum Chloroflexus aurantiacus and Cytophagahutchisonii as well as upstream of the metNPQ operon encod-ing methionine transport system in Petrotoga miotherma Amore relaxed search in (28) identified candidate S-boxes alsoin the genomes of Fusobacterium nucleatum (upstream of themetK and metN genes) Deinococcus radiodurans (metH andmetN) Xanthomonas campestris (metX) and Geobacter sul-furreducens (metB and metX) All other bacterial taxonomicgroups with available genomic sequences seem to lack theS-box regulatory system
Recent experiments (26ndash28) demonstrated that SAM-dependent regulation of the methionine biosynthesis genesof Bsubtilis involves attenuation of transcription using for-mation of alternative secondary structures in the S-box regionHere we tested whether the same regulatory mechanism couldoperate for all found S-boxes Downstream of all S-boxes fromthe BacillusClostridium group of bacteria we identifiedadditional hairpins that are followed by runs of thymidinesand therefore are candidate terminators of transcriptionIn addition we observed complementary RNA regions thatpartially overlap both the proposed terminator and the basestem (helix P1) of the S-box Thus the same termination-antitermination S-box mechanism possibly operates in allGram-positive bacteria
Analysis of the upstream regions of the S-box-regulatedgenes from two actinobacteria reveals another possiblemode of regulation In this case the S-box motif directly over-laps the ribosome-binding site of the SCD95A26 gene pos-sibly acting as a sequestor We predict that in these bacteriaSAM-stabilized S-box structure directly represses initiation oftranslation Similar mode of regulation was previously sug-gested for other metabolite-responsive riboswitches in actino-bacteria (32)
Methionine-specific T-box regulon Using a training set ofexperimentally known T-box structures from Bsubtilis andother Gram-positive bacteria (31) we constructed a patternfor the T-box motif and scanned bacterial genomes using theRNA-PATTERN program The pattern was absolutely specificsince candidate T-boxes were mostly found upstream of aminoacid metabolism-related genes (data not shown) Then weselected 38 T-box sequences with AUG (methionine) specifiercodons (see supplementary Figure 2 for multiple alignment)Although the T-box system is widely used in bacteria from theBacillusClostridium group (where more than 200T-boxeswere identified A G Vitreschak unpublished results) methio-nine-specific T-boxes occur only in a limited number of species
In the genomes of Bacillus halodurans and six clostridiathere is only one methionine-specific T-box that precedes themetS gene encoding methionyl-tRNA synthetase (not includedin Table 1) The methionine T-box regulation seems tobe extensively used only in the Lactobacillales group(30 Met-T-boxes) where it exclusively controls methioninebiosynthesis and transport genes (Table 1) This phylogenetic
Nucleic Acids Research 2004 Vol 32 No 11 3343
Ta
ble
1
Met
hio
nin
ebio
synth
esis
and
tran
sport
gen
esan
dm
ethio
nin
e-sp
ecif
icre
gula
tory
elem
ents
inG
ram
-posi
tive
bac
teri
a
Gen
eso
fth
eu
pp
erp
arto
fth
em
eth
ion
ine
pat
hw
ayin
vo
lved
inth
eh
om
ocy
stei
ne
syn
thes
isar
esh
ow
nin
red
met
hio
nin
esy
nth
ases
and
met
hy
len
e-T
HF
red
uct
ases
in
mag
enta
Gen
esen
codin
gtr
ansp
ort
pro
tein
sar
esh
ow
nin
blu
eG
enes
from
the
SA
Mre
cycl
ing
and
rever
setr
anss
ulf
ura
tion
pat
hw
ays
are
ingre
enan
dbro
wnr
espec
tivel
yG
enes
form
ing
one
candid
ate
op
ero
n(w
ith
spac
erle
ssth
anlt1
00
bp
)ar
ese
par
ated
by
das
hes
(lsquo
rsquo)D
irec
tio
no
ftr
ansc
rip
tio
nin
div
erg
on
sis
sho
wn
by
ang
leb
rack
ets
Pre
dic
ted
S-b
ox
es(lsquo
Srsquo)
met
hio
nin
e-sp
ecif
icT
-bo
xes
(lsquoT
rsquo)M
ET
-bo
xes
(lsquoM
rsquo)an
dB
12-e
lem
ents
(lsquoB
rsquo)ar
esh
ow
nin
yel
low
gre
enr
eda
nd
blu
ere
spec
tivel
yC
yst
eine-
spec
ific
T-b
ox
(lsquoT
crsquo)
isal
sosh
ow
nin
gre
enC
onti
gen
ds
are
mar
ked
by
squar
ebra
cket
sG
enom
eab
bre
via
tions
are
giv
enin
colu
mn
lsquoAB
rsquow
ith
un
fin
ish
edg
eno
mes
mar
ked
by
ah
ash
3344 Nucleic Acids Research 2004 Vol 32 No 11
distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

antiterminator structure consists of several conserved helicesof the S-box and a new stemndashloop which is alternative to theterminator and the base stem of the S-box Moreover Gram-positive bacteria have another control system involvingpremature transcription termination (T-box) which regulatesexpression of various aminoacyl-tRNA synthetases and genesinvolved in the amino acid biosynthesis (252930) The T-boxsequence is able to bind uncharged tRNA promoting forma-tion of the antiterminator The major role in regulation isplayed by the T-box lsquospecifier codonrsquo which interacts withthe anticodon of an uncharged tRNA As the position of thisregulatory codon in the T-box structure is fixed one can pre-dict the amino acid specificity of the regulatory signal (31) Forexample the presence of a T-box with an isoleucine specifiercodon upstream of ileS in Thermoanaerobacter tengcongensisargues against a recently published theory that this gene ismis-annotated and encodes a methionyl-tRNA synthase (23)
Comparative genomics is a powerful technique for recon-struction of metabolic pathways and their DNA- or RNA-levelregulation in bacteria (32ndash37) We analyzed the methioninepathway and its regulation in various Gram-positive bacteriaWe extended the S-box and methionine-specific T-box regu-lons that appear to be widely distributed in bacilli clostridiaand lactobacilli identified a new regulatory signal for methio-nine biosynthetic and transport genes in streptococci recon-structed the likely evolutionary scenario for methionineregulons of Gram-positive bacteria and described the possiblemechanism of dual regulation of the reverse transsulfurationpathway by cysteine and SAM in Clostridium acetobutylicumvia RNA regulatory structures and antisense RNA Afterreconstruction of the methionine metabolic pathways and reg-ulatory interactions in Gram-positive bacteria we identifiedseveral new enzymes and transporters involved in the methio-nine metabolism In particular we identified a set of candidategenes forming the complete SAM-recycling pathway
MATERIALS AND METHODS
Complete and partial sequences of bacterial genomes weredownloaded from GenBank (38) Preliminary sequence datawere obtained also from the websites of The Institutefor Genomic Research (httpwwwtigrorg) University ofOklahomarsquos Advanced Center for Genome Technology(httpwwwgenomeouedu) the Sanger Centre (httpwww sangeracuk) the DOE Joint Genome Institute (httpwww jgidoegov) and the ERGO Database IntegratedGenomics Inc (39)
The conserved secondary structure of the S-box and T-boxleaders was derived using the RNAMultAln program(A A Mironov in preparation) This heuristic program simul-taneously creates a multiple alignment and a conserved sec-ondary structure for a set of RNA sequences using positionalrelationship of conserved sequence boxes and paired regionsof candidate helices This program is not based on the energyminimization but rather uses simple heuristic rules of basepairing and analysis of complementary substitutions TheRNA-PATTERN program (40) was used to search for newS-boxes and methionine-specific T-boxes in bacterial gen-omes The input RNA pattern described the RNA secondarystructure and the sequence consensus motifs as a set of the
following parameters the number of helices the length ofeach helix the loop lengths and description of the topologyof helix pairs The latter is defined by coordinates of helicesFor instance two helices may be either independent or embed-ded helices or else they could form a pseudoknot structureThis definition is similar to an approach implemented in thePalingol algorithm (41) Free energy of the S-box structureswere calculated using Zukerrsquos algorithm of free energy mini-mization (4243) implemented in the Mfold program (httpbioinfomathrpiedu~mfoldrna) RNA secondary structureof the antiterminatorantisequestor conformation which in-cludes parts of the S-box and overlaps with the terminatorsequester hairpin (Figure 7A) was predicted using Mfold withthe input including the S-box sequence extended downstreamas far as the center of the terminatorsequestor hairpin Thisstructure is more energetically favorable than S-box in theabsence of ligand To model the effect of ligand bindingthat stabilizes the S-box formation of the terminatorsequesterhairpins was modeled with input sequence starting immedi-ately downstream of S-box
A simple iterative procedure implemented in the programSignalX was used for construction of the MET-box profilefrom a set of upstream gene fragments (44) Weak palin-dromes were selected in each region Each palindrome wascompared to all other palindromes and the palindromes mostsimilar to the initial one were used to make a profile Thepositional nucleotide weights in this profile were defined as
W bketh THORN = log N bketh THORN + 05frac12 025X
ifrac14ACGT
log N iketh THORN + 05frac12
where N(b k) was the count of nucleotide b in position k (45)The candidate site score was defined as the sum of the respect-ive positional nucleotide weights
Z b1 bLeth THORN =X
Kfrac141 L
W bk keth THORN
where k was the length of the site Z-score can be used to assessthe significance of an individual site
These profiles are used to scan the set of palindromes againand the procedure was iterated until convergence Thus a set ofprofiles is constructed The quality of a profile was defined asits information content (46)
I =X
Kfrac141 L
X
ifrac14ACGT
f i keth THORNlog f i keth THORN=025frac12
where f(i k) is the frequency of nucleotide i in position k ofsites generating the profile The best profile is used as therecognition rule
Each genome was scanned with the profile and genes withcandidate regulatory sites in the upstream regions (in positions325 to +25 relative to the translation start) were selected Thethreshold for the site search was defined as the lowest scoreobserved in the training set
Protein similarity search was done using the SmithndashWaterman algorithm implemented in the GenomeExplorerprogram (47) Orthologous proteins were initially definedby the best bidirectional hit criterion (48) and if necessaryconfirmed by construction of phylogenetic trees for the
3342 Nucleic Acids Research 2004 Vol 32 No 11
corresponding protein families The phylogenetic trees of themethionine biosynthesis and transport proteins were con-structed by the maximum likelihood method implementedin PHYLIP (49) Multiple sequence alignments were doneusing CLUSTALX (50) Transmembrane segments were pre-dicted using TMpred (httpwwwchembnetorgsoftwareTMPRED_formhtml) The COG (48) InterPro (51) andPFAM (52) databases were used to verify the protein func-tional and structural annotation
Alignments of S-box and T-box sequences mentioned in thispaper as well as sequences of proteins whose functions wereassigned here are available as supplementary Figures 1 2 and 3respectively
RESULTS
Reconstruction of methionine regulons
Initially orthologs of known methionine biosynthetic andtransport genes (MET) were identified by similarity searchin the genomes of all available Gram-positive bacteria(Table 1) For further analysis positional clusters (includingpossible operons) of the MET genes were also described in thistable The hom gene for homoserine dehydrogenase shared bythe methionine and threonine biosynthesis was considered onlyif it was co-localized or co-regulated with other MET genes
S-box regulon Then we constructed the pattern of the S-boxmotif using the training set of 18 S-box leader regions (24) andscanned available genomic sequences using the RNA-PAT-TERN program Multiple alignment of 100 S-box leaders from23 bacterial genomes confirms high degree of conservation ofthe S-box primary and secondary structure (see supplementaryFigure 1) Similar to other metabolite-binding RNA elements(32) the S-box motif has a set of helices (P2 to P5) closed by asingle base stem (P1) and numerous highly conserved regions(Figure 2) In fact the regions of sequence conservation covermost of the S-box sequence and therefore may be involved inSAM binding or tertiary interactions Only two stemndashloops atthe ends of the fourth and fifth helices are not conserved on thesequence level and have variable length The end loop of helixP3 (50-CnGG-30) possibly forms a pseudoknot structure withthe interior loop between helices P4 and P5 (50-CCnG-30) Thispossible pseudoknot interaction is confirmed by several com-pensatory substitutions and could be required for the formationof stable S-box tertiary structure For instance the S-box struc-ture on the Bacillus cereus mtnW gene contains a possiblepseudoknot (50-CGAG-30 50-CTCG-30) with complementarysubstitutions in the conserved positions of both arms
Among Gram-positive bacteria the S-box motif is widelydistributed in the orders Bacillales and Clostridia but it hasnot been found in Lactobacillales including EnterococcusStreptococcus and Lactococcus species (Table 1) Positionalanalysis of genes possessing this regulatory motif has showedthat the S-box regulon in Gram-positive bacteria contains mostgenesof themethioninebiosynthesis and transport aswellas theSAM synthase gene metK The S-box regulon is most extensivein three bacilli Bsubtilis Bcereus and Oiheyensis where itcontains 11 16 and 13 regulatory elements respectively andincludes additional genes for the cysteine biosynthesis andmethionine salvage pathways as well as hypothetical genesThe detailed phylogenetic and positional analysis of the
S-box-regulated genes in Gram-positive bacteria is given inthe next section
The S-box motif is not restricted to the BacillusClostridiumgroup of bacteria In two actinobacteria Streptomyces coeli-color and Thermobifida fusca S-box precedes the hypotheticalgene SCD95A26 In addition single S-box motif was foundupstream of the metY-metX operon in the genomes of Chlor-obium tepidum Chloroflexus aurantiacus and Cytophagahutchisonii as well as upstream of the metNPQ operon encod-ing methionine transport system in Petrotoga miotherma Amore relaxed search in (28) identified candidate S-boxes alsoin the genomes of Fusobacterium nucleatum (upstream of themetK and metN genes) Deinococcus radiodurans (metH andmetN) Xanthomonas campestris (metX) and Geobacter sul-furreducens (metB and metX) All other bacterial taxonomicgroups with available genomic sequences seem to lack theS-box regulatory system
Recent experiments (26ndash28) demonstrated that SAM-dependent regulation of the methionine biosynthesis genesof Bsubtilis involves attenuation of transcription using for-mation of alternative secondary structures in the S-box regionHere we tested whether the same regulatory mechanism couldoperate for all found S-boxes Downstream of all S-boxes fromthe BacillusClostridium group of bacteria we identifiedadditional hairpins that are followed by runs of thymidinesand therefore are candidate terminators of transcriptionIn addition we observed complementary RNA regions thatpartially overlap both the proposed terminator and the basestem (helix P1) of the S-box Thus the same termination-antitermination S-box mechanism possibly operates in allGram-positive bacteria
Analysis of the upstream regions of the S-box-regulatedgenes from two actinobacteria reveals another possiblemode of regulation In this case the S-box motif directly over-laps the ribosome-binding site of the SCD95A26 gene pos-sibly acting as a sequestor We predict that in these bacteriaSAM-stabilized S-box structure directly represses initiation oftranslation Similar mode of regulation was previously sug-gested for other metabolite-responsive riboswitches in actino-bacteria (32)
Methionine-specific T-box regulon Using a training set ofexperimentally known T-box structures from Bsubtilis andother Gram-positive bacteria (31) we constructed a patternfor the T-box motif and scanned bacterial genomes using theRNA-PATTERN program The pattern was absolutely specificsince candidate T-boxes were mostly found upstream of aminoacid metabolism-related genes (data not shown) Then weselected 38 T-box sequences with AUG (methionine) specifiercodons (see supplementary Figure 2 for multiple alignment)Although the T-box system is widely used in bacteria from theBacillusClostridium group (where more than 200T-boxeswere identified A G Vitreschak unpublished results) methio-nine-specific T-boxes occur only in a limited number of species
In the genomes of Bacillus halodurans and six clostridiathere is only one methionine-specific T-box that precedes themetS gene encoding methionyl-tRNA synthetase (not includedin Table 1) The methionine T-box regulation seems tobe extensively used only in the Lactobacillales group(30 Met-T-boxes) where it exclusively controls methioninebiosynthesis and transport genes (Table 1) This phylogenetic
Nucleic Acids Research 2004 Vol 32 No 11 3343
Ta
ble
1
Met
hio
nin
ebio
synth
esis
and
tran
sport
gen
esan
dm
ethio
nin
e-sp
ecif
icre
gula
tory
elem
ents
inG
ram
-posi
tive
bac
teri
a
Gen
eso
fth
eu
pp
erp
arto
fth
em
eth
ion
ine
pat
hw
ayin
vo
lved
inth
eh
om
ocy
stei
ne
syn
thes
isar
esh
ow
nin
red
met
hio
nin
esy
nth
ases
and
met
hy
len
e-T
HF
red
uct
ases
in
mag
enta
Gen
esen
codin
gtr
ansp
ort
pro
tein
sar
esh
ow
nin
blu
eG
enes
from
the
SA
Mre
cycl
ing
and
rever
setr
anss
ulf
ura
tion
pat
hw
ays
are
ingre
enan
dbro
wnr
espec
tivel
yG
enes
form
ing
one
candid
ate
op
ero
n(w
ith
spac
erle
ssth
anlt1
00
bp
)ar
ese
par
ated
by
das
hes
(lsquo
rsquo)D
irec
tio
no
ftr
ansc
rip
tio
nin
div
erg
on
sis
sho
wn
by
ang
leb
rack
ets
Pre
dic
ted
S-b
ox
es(lsquo
Srsquo)
met
hio
nin
e-sp
ecif
icT
-bo
xes
(lsquoT
rsquo)M
ET
-bo
xes
(lsquoM
rsquo)an
dB
12-e
lem
ents
(lsquoB
rsquo)ar
esh
ow
nin
yel
low
gre
enr
eda
nd
blu
ere
spec
tivel
yC
yst
eine-
spec
ific
T-b
ox
(lsquoT
crsquo)
isal
sosh
ow
nin
gre
enC
onti
gen
ds
are
mar
ked
by
squar
ebra
cket
sG
enom
eab
bre
via
tions
are
giv
enin
colu
mn
lsquoAB
rsquow
ith
un
fin
ish
edg
eno
mes
mar
ked
by
ah
ash
3344 Nucleic Acids Research 2004 Vol 32 No 11
distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

corresponding protein families The phylogenetic trees of themethionine biosynthesis and transport proteins were con-structed by the maximum likelihood method implementedin PHYLIP (49) Multiple sequence alignments were doneusing CLUSTALX (50) Transmembrane segments were pre-dicted using TMpred (httpwwwchembnetorgsoftwareTMPRED_formhtml) The COG (48) InterPro (51) andPFAM (52) databases were used to verify the protein func-tional and structural annotation
Alignments of S-box and T-box sequences mentioned in thispaper as well as sequences of proteins whose functions wereassigned here are available as supplementary Figures 1 2 and 3respectively
RESULTS
Reconstruction of methionine regulons
Initially orthologs of known methionine biosynthetic andtransport genes (MET) were identified by similarity searchin the genomes of all available Gram-positive bacteria(Table 1) For further analysis positional clusters (includingpossible operons) of the MET genes were also described in thistable The hom gene for homoserine dehydrogenase shared bythe methionine and threonine biosynthesis was considered onlyif it was co-localized or co-regulated with other MET genes
S-box regulon Then we constructed the pattern of the S-boxmotif using the training set of 18 S-box leader regions (24) andscanned available genomic sequences using the RNA-PAT-TERN program Multiple alignment of 100 S-box leaders from23 bacterial genomes confirms high degree of conservation ofthe S-box primary and secondary structure (see supplementaryFigure 1) Similar to other metabolite-binding RNA elements(32) the S-box motif has a set of helices (P2 to P5) closed by asingle base stem (P1) and numerous highly conserved regions(Figure 2) In fact the regions of sequence conservation covermost of the S-box sequence and therefore may be involved inSAM binding or tertiary interactions Only two stemndashloops atthe ends of the fourth and fifth helices are not conserved on thesequence level and have variable length The end loop of helixP3 (50-CnGG-30) possibly forms a pseudoknot structure withthe interior loop between helices P4 and P5 (50-CCnG-30) Thispossible pseudoknot interaction is confirmed by several com-pensatory substitutions and could be required for the formationof stable S-box tertiary structure For instance the S-box struc-ture on the Bacillus cereus mtnW gene contains a possiblepseudoknot (50-CGAG-30 50-CTCG-30) with complementarysubstitutions in the conserved positions of both arms
Among Gram-positive bacteria the S-box motif is widelydistributed in the orders Bacillales and Clostridia but it hasnot been found in Lactobacillales including EnterococcusStreptococcus and Lactococcus species (Table 1) Positionalanalysis of genes possessing this regulatory motif has showedthat the S-box regulon in Gram-positive bacteria contains mostgenesof themethioninebiosynthesis and transport aswellas theSAM synthase gene metK The S-box regulon is most extensivein three bacilli Bsubtilis Bcereus and Oiheyensis where itcontains 11 16 and 13 regulatory elements respectively andincludes additional genes for the cysteine biosynthesis andmethionine salvage pathways as well as hypothetical genesThe detailed phylogenetic and positional analysis of the
S-box-regulated genes in Gram-positive bacteria is given inthe next section
The S-box motif is not restricted to the BacillusClostridiumgroup of bacteria In two actinobacteria Streptomyces coeli-color and Thermobifida fusca S-box precedes the hypotheticalgene SCD95A26 In addition single S-box motif was foundupstream of the metY-metX operon in the genomes of Chlor-obium tepidum Chloroflexus aurantiacus and Cytophagahutchisonii as well as upstream of the metNPQ operon encod-ing methionine transport system in Petrotoga miotherma Amore relaxed search in (28) identified candidate S-boxes alsoin the genomes of Fusobacterium nucleatum (upstream of themetK and metN genes) Deinococcus radiodurans (metH andmetN) Xanthomonas campestris (metX) and Geobacter sul-furreducens (metB and metX) All other bacterial taxonomicgroups with available genomic sequences seem to lack theS-box regulatory system
Recent experiments (26ndash28) demonstrated that SAM-dependent regulation of the methionine biosynthesis genesof Bsubtilis involves attenuation of transcription using for-mation of alternative secondary structures in the S-box regionHere we tested whether the same regulatory mechanism couldoperate for all found S-boxes Downstream of all S-boxes fromthe BacillusClostridium group of bacteria we identifiedadditional hairpins that are followed by runs of thymidinesand therefore are candidate terminators of transcriptionIn addition we observed complementary RNA regions thatpartially overlap both the proposed terminator and the basestem (helix P1) of the S-box Thus the same termination-antitermination S-box mechanism possibly operates in allGram-positive bacteria
Analysis of the upstream regions of the S-box-regulatedgenes from two actinobacteria reveals another possiblemode of regulation In this case the S-box motif directly over-laps the ribosome-binding site of the SCD95A26 gene pos-sibly acting as a sequestor We predict that in these bacteriaSAM-stabilized S-box structure directly represses initiation oftranslation Similar mode of regulation was previously sug-gested for other metabolite-responsive riboswitches in actino-bacteria (32)
Methionine-specific T-box regulon Using a training set ofexperimentally known T-box structures from Bsubtilis andother Gram-positive bacteria (31) we constructed a patternfor the T-box motif and scanned bacterial genomes using theRNA-PATTERN program The pattern was absolutely specificsince candidate T-boxes were mostly found upstream of aminoacid metabolism-related genes (data not shown) Then weselected 38 T-box sequences with AUG (methionine) specifiercodons (see supplementary Figure 2 for multiple alignment)Although the T-box system is widely used in bacteria from theBacillusClostridium group (where more than 200T-boxeswere identified A G Vitreschak unpublished results) methio-nine-specific T-boxes occur only in a limited number of species
In the genomes of Bacillus halodurans and six clostridiathere is only one methionine-specific T-box that precedes themetS gene encoding methionyl-tRNA synthetase (not includedin Table 1) The methionine T-box regulation seems tobe extensively used only in the Lactobacillales group(30 Met-T-boxes) where it exclusively controls methioninebiosynthesis and transport genes (Table 1) This phylogenetic
Nucleic Acids Research 2004 Vol 32 No 11 3343
Ta
ble
1
Met
hio
nin
ebio
synth
esis
and
tran
sport
gen
esan
dm
ethio
nin
e-sp
ecif
icre
gula
tory
elem
ents
inG
ram
-posi
tive
bac
teri
a
Gen
eso
fth
eu
pp
erp
arto
fth
em
eth
ion
ine
pat
hw
ayin
vo
lved
inth
eh
om
ocy
stei
ne
syn
thes
isar
esh
ow
nin
red
met
hio
nin
esy
nth
ases
and
met
hy
len
e-T
HF
red
uct
ases
in
mag
enta
Gen
esen
codin
gtr
ansp
ort
pro
tein
sar
esh
ow
nin
blu
eG
enes
from
the
SA
Mre
cycl
ing
and
rever
setr
anss
ulf
ura
tion
pat
hw
ays
are
ingre
enan
dbro
wnr
espec
tivel
yG
enes
form
ing
one
candid
ate
op
ero
n(w
ith
spac
erle
ssth
anlt1
00
bp
)ar
ese
par
ated
by
das
hes
(lsquo
rsquo)D
irec
tio
no
ftr
ansc
rip
tio
nin
div
erg
on
sis
sho
wn
by
ang
leb
rack
ets
Pre
dic
ted
S-b
ox
es(lsquo
Srsquo)
met
hio
nin
e-sp
ecif
icT
-bo
xes
(lsquoT
rsquo)M
ET
-bo
xes
(lsquoM
rsquo)an
dB
12-e
lem
ents
(lsquoB
rsquo)ar
esh
ow
nin
yel
low
gre
enr
eda
nd
blu
ere
spec
tivel
yC
yst
eine-
spec
ific
T-b
ox
(lsquoT
crsquo)
isal
sosh
ow
nin
gre
enC
onti
gen
ds
are
mar
ked
by
squar
ebra
cket
sG
enom
eab
bre
via
tions
are
giv
enin
colu
mn
lsquoAB
rsquow
ith
un
fin
ish
edg
eno
mes
mar
ked
by
ah
ash
3344 Nucleic Acids Research 2004 Vol 32 No 11
distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353
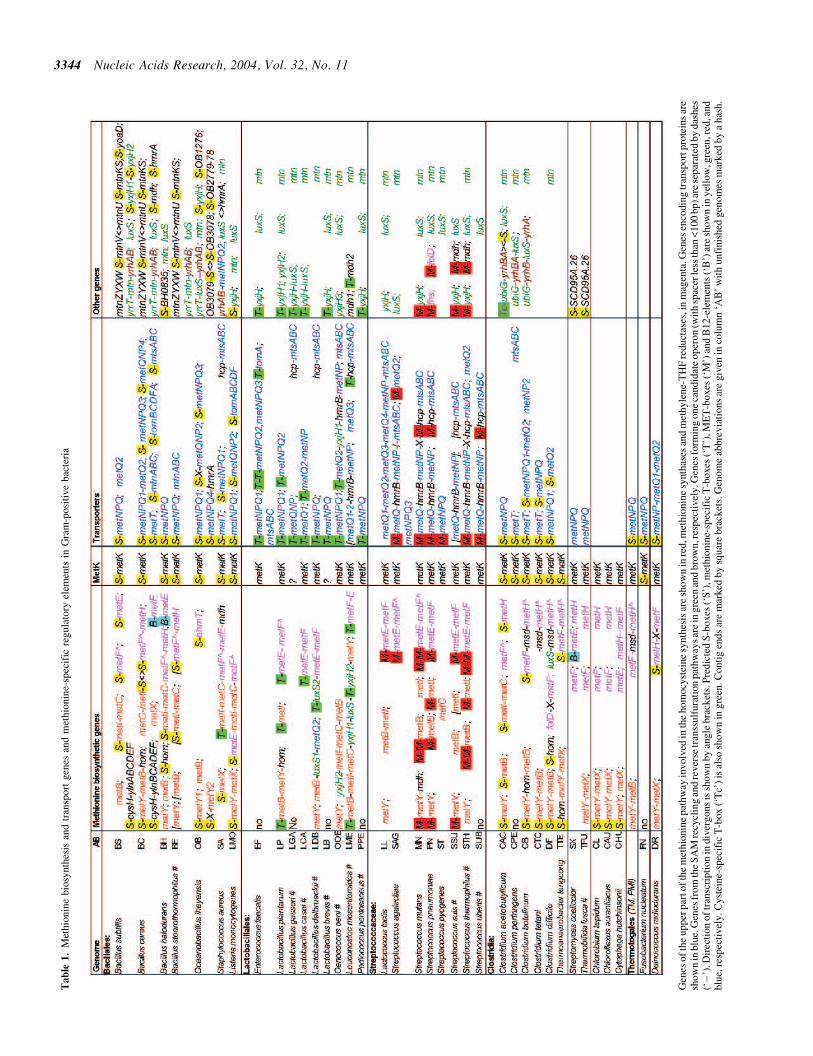
Ta
ble
1
Met
hio
nin
ebio
synth
esis
and
tran
sport
gen
esan
dm
ethio
nin
e-sp
ecif
icre
gula
tory
elem
ents
inG
ram
-posi
tive
bac
teri
a
Gen
eso
fth
eu
pp
erp
arto
fth
em
eth
ion
ine
pat
hw
ayin
vo
lved
inth
eh
om
ocy
stei
ne
syn
thes
isar
esh
ow
nin
red
met
hio
nin
esy
nth
ases
and
met
hy
len
e-T
HF
red
uct
ases
in
mag
enta
Gen
esen
codin
gtr
ansp
ort
pro
tein
sar
esh
ow
nin
blu
eG
enes
from
the
SA
Mre
cycl
ing
and
rever
setr
anss
ulf
ura
tion
pat
hw
ays
are
ingre
enan
dbro
wnr
espec
tivel
yG
enes
form
ing
one
candid
ate
op
ero
n(w
ith
spac
erle
ssth
anlt1
00
bp
)ar
ese
par
ated
by
das
hes
(lsquo
rsquo)D
irec
tio
no
ftr
ansc
rip
tio
nin
div
erg
on
sis
sho
wn
by
ang
leb
rack
ets
Pre
dic
ted
S-b
ox
es(lsquo
Srsquo)
met
hio
nin
e-sp
ecif
icT
-bo
xes
(lsquoT
rsquo)M
ET
-bo
xes
(lsquoM
rsquo)an
dB
12-e
lem
ents
(lsquoB
rsquo)ar
esh
ow
nin
yel
low
gre
enr
eda
nd
blu
ere
spec
tivel
yC
yst
eine-
spec
ific
T-b
ox
(lsquoT
crsquo)
isal
sosh
ow
nin
gre
enC
onti
gen
ds
are
mar
ked
by
squar
ebra
cket
sG
enom
eab
bre
via
tions
are
giv
enin
colu
mn
lsquoAB
rsquow
ith
un
fin
ish
edg
eno
mes
mar
ked
by
ah
ash
3344 Nucleic Acids Research 2004 Vol 32 No 11
distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

distribution is consistent with the absence of the S-boxregulatory system in all available genomes of Lactobacillales(see Discussion) Among all other available bacterial geno-mes we have found only one additional methionine T-box Itprecedes the metICFE-mdh operon in Staphylococcus aureusa bacterium that mainly uses the S-box system
Methionine regulation in streptococci Then we attempted toanalyze potential methionine regulons in Streptococcaceae alarge family of Gram-positive bacteria which have neither S-boxes nor methionine-specific T-boxes For this aim we col-lected upstream regions of all MET genes from Lactococcuslactis and Streptococcus species and applied the signaldetection procedure A highly conserved 17 bp palindromicsequence (named MET-box) with consensus 50-TATAGTTt-naAACTATA-30 was identified in all streptococci (Table 2)To find new members of the candidate regulon the derivedprofile for the MET-box signal was used to scan the genomesIn various Streptococcus species the MET regulon appears toinclude most methionine biosynthesis and transport genes aswell as several hypothetical genes including yxjH mdh fhsand folD (Table 1) The predicted MET regulon in Llactiscontains only one transcriptional unit the metEF operonGiven the palindromic structure of the derived signal typicalto binding signals of transcriptional regulators we proposethat the MET-box signal plays a role in transcriptional regula-tion of the methionine metabolism in Streptococcus speciesand Llactis although the responsible regulatory protein
remains to be identified Interestingly in a recent study itwas shown that the LysR-type transcriptional regulatorMtaR is necessary for the efficient methionine uptake in Strep-tococcus agalactiae (53) MtaR has orthologs in other studiedStreptococcus species as well as in Llactis Since we havefound MET-boxes upstream of the predicted methionine trans-port operons in Sagalactiae we suppose that this MET-box isthe DNA binding signal of MtaR Moreover the MET-boxsignal is similar to signals recognized by LysR-family regu-lators in length and palindromic symmetry
Reconstruction of the methionine pathway
Positional analysis of a large number of regulatory elements(S- T- and MET-boxes) in Gram-positive bacteria allowed usto identify new genes possibly involved in the methionine
Figure 2 Conserved S-box structure Capitals indicate invariant positionsLower case letters indicate strongly conserved positions Degeneratepositions R = A or G Y = C or U D = A G or U H = A C or U N = anynucleotide Conserved helices are numbered P1 to P5 Dashes and dots indicateobligatory and facultative base pairs respectively Stemndashloops of variablelengths are shown by broken lines Possible tertiary interaction between theend loop of helix P3 and the interior loop is shown
Table 2 Candidate methionine-specific DNA signals (MET-boxes) in
Streptococcaceae
Gene Pos Score Sites
consensus TATAGTTtnaAACTATA
Lactococcus lactismetE 90 651 TATAGTTTAAAACTATAStreptococcus agalactiaemetQ1 98 651 TATAGTTTAAAACTATAmetE 131 651 TATAGTTTAAAACTATAmetQ2 116 587 TATAGTTaAAAgCTATAStreptococcus mutansyxjH 95 651 TATAGTTTAAAACTATAmetY 65 627 TATAGTTTAtAACTATAmetE 156 611 TATAGcTTAAAACTATA
187 573 TATAGaTgAAAACTATAmetQ 99 587 TATAGTTTTtAgCTATAmetB 4 581 TATAtTTgAtAACTATA
38 547 TATAGTTggAAcCTATAhcp 109 543 TATAGgTaAtAACTATAStreptococcus pneumoniaemetQ 100 651 TATAGTTTTAAACTATAmetE 100 625 TATAGTTTcAAACTATAfhs 155 625 TATAGTTTcAAACTATAmetI 92 625 TATAGTTTgAAACTATAfolD 99 611 TATAGTTTAAAgCTATAmetB 108 595 TATAaTTgAAAACTATAmetY 208 565 gATAtTTgTAAACTATAhcp 65 492 TATAaggTAAAACTATAStreptococcus pyogenesmetQ 104 591 TATAGTTTAAAACTATgStreptococcus suismetY 93 609 TATAGTTgTtAACTATAyxjH 99 567 TATAGTTTTtAACTATgmdh 80 526 TATAGTTaAAAgCaATAmetE 100 488 TATAGTcaAAAgtTATAStreptococcus thermophilusmetE 98 651 TATAGTTTAAAACTATA
170 563 TATAGTTaTtAgCTATAmetQ 104 627 TATAGTTaAAAACTATAyxjH 318 590 TATAGTcTAAAACTATAmetB 101 573 TATAGTTTAAAgtTATA
67 520 TATAtTTgAtAACaATAmdh 141 571 TATAGcTTAAAgCTATAmetI 128 566 TATAGTaaTAAACTATAStreptococcus uberismetQ 102 611 TATAGTTTTAAgCTATAhcp 86 590 TATAGTTTTAgACTATA
Site positions in lsquoPosrsquo column are given relative to the translation start site
Nucleic Acids Research 2004 Vol 32 No 11 3345
metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

metabolism In addition we analyzed the candidate MetJ bind-ing sites upstream of some methionine synthesis and transportgenes in gamma proteobacteria (data not shown) The detailedanalysis of new members of the methionine regulons andreconstruction of the metabolic pathways in various organismsis presented below
Methionine biosynthesis The pathway of methionine bio-synthesis via the transsulfuration or direct sulfhydrylationroute is conserved in most Gram-positive bacteria butsome steps vary Only complete genomes of FnucleatumClostridium perfringens Enterococccus faecalis Sagalactiaeand Streptococcus pyogenes as well as unfinished genomes ofseveral lactobacilli and Streptococcus uberis lack the de novosynthesis pathway but possess the SAM synthase gene metK(Table 1) We suggest that this metabolic gap could be filled bymethionine-specific transport systems detected in these gen-omes (metNPQ or metT see below)
We observed several cases of expansion of the methionineregulon when the upstream reactions preceding the methio-nine biosynthetic pathway become methionine regulated Oneexample is homoserine dehydrogenase (the hom gene productin Bsubtilis) which is shared by the threonine and methioninepathways (Figure 1) In most Gram-positive bacteria the homgene is co-localized with the threonine biosynthesis genesthrBC (data not shown) However in the genomes of twobacilli three clostridia and Lactobacillus plantarum wehave found a second hom paralog belonging to the methionineregulon either S-box or T-box (Table 1) Another example isthe cysteine biosynthesis gene cluster (cysH-ylnABCDEF)which is a member of the methionine S-box regulon inBsubtilis and Bcereus but not in other Bacillales Interest-ingly even in these two related bacteria the in vitro affinitiesof candidate cysH S-box motifs to the effector molecule(SAM) vary by two orders of magnitude (28) Furthermorethe expression of the Bsubtilis cysH operon is not repressed bymethionine in vivo indicating disfunction of the cysH S-boxelement (54) Indeed this S-box contains a unique CAsubstitution destabilizing helix 3 Moreover upstream regionsof cysH genes of other closely related genomes (BhaloduransBacillus stearothermophilus Oihyensis) demonstrate residualsimilarity to upstream regions of Bsubtilis and BcereuscysH but cannot fold into the S-box structure (data notshown) In any case in Bcereus this functional regulatoryinteraction seems to be rational since the bacillary transsulfura-tion pathway of the methionine biosynthesis uses cysteine as asulfur donor
The first step of the methionine biosynthesis is catalyzed byone of two non-homologous homoserine O-acetyltransferasesMetB or MetX The main difference of these two isoenzymesis that in contrast to MetX from L meyeri the MetB enzymefrom Bsubtilis is feedback inhibited by SAM (79) The MetBisoenzyme detected in most Gram-positive bacteria possessingthe methionine pathway is replaced by MetX in Staphylococ-cus Listeria and Ttengcongensis whereas the Bcereusgenome encodes both proteins (Table 1) Phylogenetic distri-bution of the MetB and MetX isoenzymes differs significantlythe former prevails in enterobacteria firmicutes and cyano-bacteria whereas the latter is common in other proteobacteriaactinobacteria various early branching bacteria and infungi Notably the SAM-inhibited isoenzyme MetB is not
methionine regulated in many cases at least not using S-T- or MET-boxes (eg in Bsubtilis) In contrast the MetXsynthesis is S-box controlled with the exception of Bcereuswhere both enzymes are present
Then we have analyzed distribution of the direct sulfhydry-lation and transsulfuration pathways of the methionine bio-synthesis which are catalyzed by the MetY and MetI-MetCrespectively Among Gram-positive bacteria the formerprevails over the latter twelve species have only MetYtwo species have MetI-MetC whereas seven remainingmethionine-producing bacteria possess both pathways(Table 1) The role of single metI genes (not accompaniedby metC) in several genomes containing the metY gene isstill not clear Thus in contrast to Bsubtilis and Saureusother methionine-producing firmicutes potentially use thedirect sulfhydrylation pathway and therefore they do notrequire cysteine for the methionine synthesis
Two non-homologous methionine synthases coenzymeB12-dependent MetH and B12-independent MetE are knownin bacteria The metE gene was detected in all methionine-producing firmicutes except clostridia and two bacilli thathave the metH gene Bcereus and Bhalodurans have boththe B12-dependent methionine synthase MetH which belongsto the S-box regulon and the B12-independent isoenzymeMetE that is likely regulated by coezyme-B12 via the B12-element riboswitch (34) The only exception is Oiheyensiswhich has neither metE nor metH Analysis of candidateS-box signals allowed us to identify a new member of themethionine regulon in this bacterium OB0691 which is simi-lar to the betainendashhomocysteine methyltransferase BhmT frommammals (55) BhmT catalyzes conversion of homocysteineto methionine like the bacterial MetE and MetH isoenzymesbut it uses another methyl donor betaine instead of methyl-THF Thus we predict that Oiheyensis uses a eukaryotic-typemethionine synthase and does not require the methylene-THFreductase MetF which is absent in this bacterium
B12-dependent methionine synthases from clostridia andThermotogales lack the C-terminal domain which is involvedin reactivation of spontaneously oxidized coenzyme B12and therefore is required for the catalysis (denoted metH^in Table 1) In all these bacteria except Ttengcongensiswe found a hypothetical gene located immediately upstreamof the metH^ gene (see Thermotoga maritima TM0269 as arepresentative of this gene family) This gene named msdhas no homologs in other genomes However we identifieda conserved sequence motif (hhhThG-28-hEhhh[DE]--RxxxGY-32-Pxx[SA][TV]x[GA]hh where lsquohrsquo denotes anyhydrophobic amino acid) which is common to all full-lengthMetH proteins and to the Msd proteins As shown by (56) theRxxxGY motif is critical for binding of SAM to the C-terminalreactivation domain of methionine synthase MetH Thus wetentatively assign the missing function of reactivation of theB12-dependent methionine synthase to the product of the msdgene The three-dimensional structure of the TM0269 proteinwas recently resolved in Joint Center for Structural Genomics(httpwwwjcsgorg) but this did not lead to assignment of acellular role for this protein
The metF gene encoding methylene-THF reductase hasbeen identified in most Gram-positive bacteria that havemethionine synthases (MetH or MetE) an exclusionbeing Clostridium tetani The MetF proteins from Bacillales
3346 Nucleic Acids Research 2004 Vol 32 No 11
Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

Cacetobutylicum Lplantarum and two streptococci have anadditional N-terminal domain highly similar to the homocys-teine-binding domain of MetH (denoted metF^ in Table 1)Previously it was proposed that this domain could be involvedin positive allosteric regulation of MetF by homocysteine (12)
Recycling of methylene-THF from THF is connected withinterconversion of serine and glycine mediated by GlyAbelonging to the methionine regulon in Ecoli (57) TheglyA gene is present in all Gram-positive bacteria butnever in the methionine regulon The alternative pathway ofmethylene-THF recycling which proceeds in two steps andrequires ATP and NADP is mediated by FolD and Fhs Thecorresponding genes are candidate members of the MET-boxregulon in Streptococcus pneumoniae whereas the folD andmetF genes form one possible operon in Clostridium difficile(Table 1) On the other hand the folD gene is a candidatemember of the purine regulon in some g-proteobacteria (58)that is also rational since methylene-THF is required for thepurine biosynthesis These facts once again demonstrategenome-specific regulon expansions and in particularindicate considerable variability in regulation of the methy-lene-THF synthesis in bacteria
Methionine transport The only known transport system formethionine is the ABC transporter MetNIQ of enteric bacteriawhich belongs to the methionine uptake transporter (MUT)family (13ndash15) An ortholog of this system in Bsubtilisencoded by the metNPQ operon is regulated by the S-boxsystem (12) Recently it was shown that metNPQ encodes anABC permease transporting methionine sulfoxide D- andL-methionine (16) To describe candidate methionine transpor-ters in Gram-positive bacteria we combined similarity searchfor the metNPQ orthologs with identification of methionine-specific regulatory sites and with positional analysis of genes
Candidate methionine transporters metNPQ are widelydistributed in Gram-positive bacteria they are absent onlyin two clostridia species (Table 1) In most cases compo-nents of methionine transporters are encoded by clusters ofco-localized genes which are preceded by S-boxes(in Bacillales and clostridia) methionine-specific T-boxes(in lactobacilli) or MET-boxes (in streptococci) To analyzethe possible origin of a large number of metNPQ paralogs inGram-positive bacteria we constructed the phylogenetic treefor the substrate-binding components of MUT-family trans-porters (Figure 3) Though in some cases eg in LlactisCampylobacter jejuni metQ paralogs possibly result fromrecent genome-specific duplications in most other casesthey have diverged early Since almost all large branchesof the tree contain members of various methionine regulonsincluding MetJ-regulated orthologs from enterobacteria (15)we believe that the methionine specificity has been retainedby all members of the MUT family However we cannotexclude the possibility that some members of the MUTfamily have a more broad specificity
A new type of candidate methionine transporters namedMetT was identified in some bacteria from the BacillusClostridium group and g-proteobacteria Among Gram-positivebacteria only Bcereus Saureus and three clostridia specieshave the metT genes which are regulated by upstream S-boxesin all cases (see Table 1 and Figure 4 for genomic identifiers)Among g-proteobacteria only Vibrio and Shewanella species
have metT genes and they are preceded by candidateMetJ sites (data not shown) MetT proteins contain elevenpredicted transmembrane segments and are similar to proteinsfrom the NhaC Na+H+ antiporter superfamily Existence oflikely S-adenosylmethionine-regulated metT in the completegenome of Cperfringens that has no methionine biosynthesisgenes suggests that MetT is a methionine transporter
The phylogenetic tree of transporters from the NhaC super-family consists of four deeply diverged branches (Figure 4)The first branch comprises predicted lysine transporters LysWmost of which are preceded by regulatory LYS-elements (35)The second branch contains predicted methionine transportersMetT preceded by either candidate S-boxes (in Gram-positivebacteria) or MetJ binding sites (in g-proteobacteria) The thirdbranch includes predicted tyrosine transporters TyrT most ofwhich are members of the tyrosine T-box regulon (59) Thefourth branch includes orthologs of malate lactate antiporterMleN from Bsubtilis (60) Thus analysis of candidate regu-latory signals allows us to tentatively assign specificities to theremaining three large sub-groups of transporters from theNhaC superfamily
Analysis of the methionine-specific regulatory signalsallowed us to identify two more hypothetical methionine-related ABC transport systems in Gram-positive bacteriaThe first one named mtsABC is present in all streptococcisome lactobacilli and Cperfringens Saureus and Bcereus(Table 1) MtsA (SMU1935c in Streptococcus mutans) hasfive predicted transmembrane segments and is not similar toany known protein The MtsB and MtsC components(SMU1934c and SMU1933c) are similar to typical ATP-binding (CbiO) and transmembrane (CbiQ) components ofvarious ABC transporters respectively In most cases mtsABCgenes are clustered with the hcp gene (SMU1936c) thatencodes a hypothetical cytosolic protein
The second methionine-related ABC transport systemnamed tom (for Transporter for Oligopeptides or Methionine)belongs to a large family of oligopeptide ABC transporters InBcereus and Listeria monocytogenes the tom gene clusters(BC0207-BC0208-BC0209-BC0210-BC0211 and LMO2196-LMO2195-LMO2194-LMO2193-LMO2192 respectively) arepreceded by S-boxes Moreover Efaecalis has a single tomAgene (EF3081) for a substrate-binding component of transpor-ter which is preceded by a methionine-specific T-box Theavailable data were insufficient to assign specificities of thesetwo methionine-regulated transporters by the genome contextanalysis and metabolic reconstruction One possibility is thatthey are involved in the uptake of some methionine precursorsor oligopeptides
Methionine salvage and SAM recycling The methionine salv-age genes mtnKSUVWXYZ involved in the methylthioriboseutilization in Bsubtlis (21ndash23) were identified only in threeBacillus species always as members of the S-box regulon(Table 1) In Bcereus one S-box motif was found upstreamof the BC0768-767-766 gene cluster (named mtnABC) encod-ing a hypothetical ABC transport system An ortholog of thissystem exists in the genome of Bstearothermophilus but notin other studied bacterial genomes Based on S-box regulatorysite similarity to the ribose ABC transporter and phylogeneticco-occurrence with the mtnKSUSVWXYZ genes we tentativelyassign the methylthioribose specificity to the MtnABC
Nucleic Acids Research 2004 Vol 32 No 11 3347
transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

transport system The absence of MtnABC in Bsubtilis whichalso can grow on methylthioribose (219) suggests existenceof other specific transport systems
In addition to the autotrophic pathway of cysteine biosynth-esis some bacterial species (including Bsubtilis) can synthes-ize this amino acid through the reverse transsulfurationpathway using methionine as a precursor (61) In this pathwaymethionine is first converted to homocysteine via the SAMrecycling pathway (Figure 1) In an attempt to discover miss-ing genes of this pathway we started with positional analysisof the Bsubtilis yrhA and yrhB genes apparently encoding thecystathionine b-synthase and g-lyase respectively (12)Orthologs of these genes were found in all Bacillus speciesSaureus and three clostridia and are always clustered formingone candidate operon (Table 1) Moreover the yrhAB genesform possible operons with the SAH nucleosidase gene mtn inbacilli and with orthologs of the Bsubtilis luxS gene inOiheyensis Cperfringens and Clostridium botulinum The
functional role of the Bsubtilis LuxS protein is notknown though its structure has been recently determined(62) The autoinducer-2 production protein LuxS from pro-teobacteria catalyzes transformation of S-ribosylhomocysteineto homocysteine (63) We assign the previously missingribosylhomocysteinase function of the SAM recycling path-way to the luxS gene product Orthologs of the luxS andmtn genes were identified in most Gram-positive bacteriacorroborating the existence of SAM recycling pathways inthese organisms
Two other genes co-localized in positional clusters with theyrhAB genes (yrrT from bacilli and ubiG from clostridia) bothcontain SAM-binding motifs and are similar to variousSAM-dependent methyltranferases (Table 1) It was knownthat SAH is synthesized from SAM as a by-product ofnumerous methylation reactions in the cell (2) Based onco-localization with the yrhAB genes we propose that thepathway of reverse synthesis of cysteine from methionine
Figure 3 Phylogenetic tree of the substrate-binding components MetQ of bacterial methionine transporters from the MUT family Proteins are denoted by genomeabbreviations (listed in Table 1) Multiple paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted andbroken lines respectively Genes known and predicted to be regulated by the methionine repressor MetJ (data not shown) are circled Additional genomeabbreviations are Escherichia coli (EC) Salmonella typhi (TY) Yersinia pestis (YP) Yersinia enterocolitica (YE) Erwinia carotovora (EO) Vibrio cholerae(VC) Haemophilus influenzae (HI) Fusobacterium nucleatum (FN) Helicobacter pylori (HP) Deinococcus radiodurans (DR) Treponema pallidum (TP)Caulobacter crescentus (CO) and Pseudomonas aeruginosa (PA)
3348 Nucleic Acids Research 2004 Vol 32 No 11
could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

could require specific SAM-dependent methylases and assignthis role to YrrT in bacilli and UbiG in clostridia
The hypothesis that the ubiG-yrhAB operon of Cacetobu-tylicum is involved in the cysteine synthesis is further sup-ported by observation of a cysteine-specific T-box upstream ofthis operon likely mediating repression by cysteine (12)Moreover a backward-directed S-box motif located immedi-ately downstream of this operon could regulate formation of anantisense transcript for this locus assuming activation of theubiG-yrhAB operon by methionine Thus we predict that genesfor the reverse transsulfuration pathway in Cacetobutylicumare apparently expressed only in the conditions of methionineexcess and cysteine deficiency (see Figures 1 and 5) Someother mode of regulation by interference of transcription fromthe complementary strand also could be involved but it wouldnot change the main conclusion
Two highly similar genes of unknown function yxjH andyxjG are members of the S-box regulon in Bsubtilis (24) Theyencode proteins with moderate similarity to the C-terminal
part of the B12-independent methionine synthase MetEMutation analysis has showed that these genes are not requiredfor the de novo methionine synthesis in Bsubtlis (12) Ortho-logs of these genes (denoted yxjH) were identified in variousGram-positive bacteria several proteobacteria and archaea(see the phylogenetic tree in Figure 6) Similar to Bsubtilisthe yxjH genes in Oiheyensis and Lmonocytogenes are S-boxregulated Moreover most yxjH orthologs from Lactobacil-lales and Streptococcus species belong to the methionineT-box and MET-box regulons respectively (Table 1)
Negative regulation of the B12-independent methioninesynthase isozymes by B12-element and vitamin B12 is com-mon in bacteria possessing both B12-dependent and B12-independent isozymes (34) At that vitamin B12-responsiveregulatory elements were detected upstream of the yxjH ortho-logs in Rhodopseudomonas palustris and Bacteroides fragilistwo bacteria possessing B12-dependent methionine synthases(34) It indicates that these two B12-regulated yxjH orthologslikely function as B12-independent methionine synthases
Figure 5 Predicted regulation of the Cacetobutylicum ubiG-yrhBA operon by S-adenosylmethionine-specific S-box and cysteine-specific T-box regulatory signals
Figure 4 Phylogenetic tree of the NhaC Na+H+ antiporter superfamily including predicted methionine- lysine- and tyrosine-specific transporters Gene identifiersare shown for annotated complete genome sequences Genes predicted to be regulated by the methionine-specific S-box motif and MetJ repressor are boxed andcircled respectively Genes predicted to be regulated by LYS elements (29) are denoted by filled circles Genes predicted to be regulated by tyrosine-specific T-boxes(48) are denoted by black squares
Nucleic Acids Research 2004 Vol 32 No 11 3349
Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

Further based on results of positional analysis and phylo-genetic profiling we suggest that YxjH could function as analternative methionine synthase enzyme mainly involved inthe SAM recycling pathway Indeed we identified severalcases of co-localization of the yxjH genes with the candidateSAM recycling gene luxS (in Lactobacillus gasseri Lactoba-cillus casei and Leuconostoc mesenteroides) and with themethionine biosynthesis genes metICB (in Oenococcusoeni) Notably the latter genome lacks both methioninesynthase genes (metE and metH) In addition we foundmethionine-regulated yxjH genes in some species from theorder Lactobacillales which lack genes for the de novomethionine synthesis but possess genes for both methioninetransport and SAM recycling However our assignment of themethionine synthase role to YxjH contradicts the observationthat the metE Bsubtilis mutant is a methionine auxotroph thussuggesting that MetE is the only methionine synthase inthis species (1264) Additional experiments are required toprove this tentative assignment of yxjH to the SAM recyclingpathway
Other candidate members of methionine regulons Candidatemethionine transporter operons in Streptococcus OenococcusLeuconostoc and Oceanobacillus species include a hypothe-tical gene (hmrA or hmrB) from the M20 family of zinc metal-lopeptidases Known members of this family catalyzethe release of an N-terminal amino acid usually neutral or
hydrophobic from a polypeptide (65) Single hmrA gene ofBcereus (BC3176) is a member of the S-box regulon Inaddition methionine regulons of Bcereus Saureus Lmesen-teroides and three Streptococcus species contain a putativemetal-dependent hydrolase gene named mdh (genesBC0395 SA0343 and SMU1172 in complete genomes andCOG entry 1878) In other Gram-positive bacteria the mdhgenes are not regulated by methionine at least not by knownsystems The role of the hmrA hmrB and mdh gene products inthe methionine metabolism is not clear
In Scoelicolor and Tfusca S-box precedes the SCD95A26gene encoding a hypothetical pyridoxal-phosphate dependentenzyme (PFAM entry PF00291) Enzymes of this class cata-lyze various reactions in the metabolism of amino acids (66)SCD95A26 is a distant homolog of the threonine synthasegene thrC However actinobacteria have a proper thrCgene located within the threonine biosynthetic gene clusterSince the complete genome of Scoelicolor lacks orthologs ofknown genes for the synthesis of homocysteine the directprecursor of methionine we tentatively suggest thatSCD95A26 could be involved in the homocysteine synthesisin actinobacteria
DISCUSSION
Methionine biosynthetic and transport genes are regulated bydifferent mechanisms in various microbial species (Figure 7)The S-box system in Bacillales and Clostridia orders and themethionine-specific T-box system in Lactobacillales are RNA-dependent regulatory systems that both control transcriptiontermination although in a different manner The S-box RNAstructure is stabilized by direct binding of an effector mole-cule S-adenosylmethionine whereas the T-box senses lack ofamino acid via the presence of uncharged tRNAs Bacteriafrom the Streptococcaceae family lacking both theseRNA-dependent systems are predicted to have a classicalDNA-dependent system for regulation of the methioninemetabolism which includes numerous MET-box sites andthe MtaR regulatory protein Regulation of the methioninebiosynthesis and transport genes in Gram-negative entero-bacterium Ecoli is also DNA-dependent and involves twoknown transcription factors activator MetR and repressorMetJ that bind operators absolutely different from MET-box (8) The most intriguing mode of regulation was foundfor the Cacetobutylicum ubiG-yrhAB operon which seems tobe regulated by both S- and T-box systems (Figure 5)
Inspite of a variety of regulatory systems the cores of themethionine regulons in Gram-negative and Gram-positive spe-cies almost completely coincide They include most genesrequired for the de novo methionine synthesis and transportIn contrast SAM synthase gene metK belongs to the methio-nine regulons only in bacilliclostridia and g-proteobacteriavia the S-box and MetJ regulatory systems respectively Inlactobacilli and streptococci this gene is not regulated by eitherthe T-box or MET-box systems One possible explanation forthis could be the use of different effector molecules for themethionine regulons in these taxonomic groups SAM is aknown effector of the S-box and MetJ regulatory systemswhereas methionine itself is involved in the regulation bythe T-box system via methionine-specific tRNA Apparently
Figure 6 Phylogenetic tree of bacterial orthologs of the Bsubtilis yxjH geneProteins are denoted by the genome abbreviations (listed in Table 1) Multiplegene paralogs are numbered Genes predicted to be regulated by S-boxes T-boxes and MET-boxes are boxed by solid dotted and broken lines respectivelyGenes predicted to be regulated by the vitamin B12-specific RNA regulatoryelement (28) are circled by broken lines Additional genome abbreviations areErwinia carotovora (EO) Klebsiella pneumoniae (KP) Rhodopseudomonaspalustris (RPA) Mannheimia haemolytica (PQ) Bfragilis (BX) andChlamydophila pneumoniae (CPX)
3350 Nucleic Acids Research 2004 Vol 32 No 11
in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

in the latter case it is not necessary to regulate SAM synthesisby the methionine availability The effector molecule for thepredicted MET-box system in streptococci is not known In thisbacterial group MET-boxes precede most of the methioninebiosynthesis and transport genes but not the metK genes(Table 1) Using the same biochemical logic we tentativelyassign the role of the regulatory molecule in streptococci tomethionine itself
Compared to other riboswitches S-boxes demonstratesomewhat mosaic phylogenetic distribution They wereobserved in two major groups of firmicutes bacilli and clos-tridia and in a number of other taxons Although riboswitchesare subject to frequent horizontal transfer (32) and it cannot beruled out in this case especially for isolated proteobacterialgenomes (Xanthomonas and Geobacter) the existence ofS-boxes in a variety of genomes argues for their ancient originIn particular the most parsimonious evolutionary scenario forfirmicutes seems to be the following S-boxes were present inthe last common ancestor of bacilli and clostridia ie the lastcommon ancestor of all firmicutes and it was lost in theStreptococcaceae and Lactobacillales lineages In strepto-cocci the role of S-boxes in the regulation of methioninemetabolism was assumed by the transcriptional regulator
MtaR with the MET-box binding signal whereas in lactoba-cilli the S-box regulon was absorbed by the expanded Met-T-box regulon which initially included only aminoacyl-tRNAsynthetases One possible example of an early stage of suchexpansion could be the Met-T-box upstream of the metICFE-mdh operon in Saureus On the other hand degradation of aS-box is exemplified by the case of cysH genes of Bsubtilisand closely related genomes
The comparative analysis of regulation supplemented bygenome context and similarity search techniques allowed usto identify several new methionine-related transport systemsassign missing functions of the SAM recycling pathway anddemonstrate variability in the upper part of the methioninepathway in different species Furthermore we identifiedgenome-specific extensions of the methionine regulon thatinvolve genes shared with other biochemical pathwaysThis study not only demonstrates once again the power ofcomparative genomics in functional annotation of genesprediction of regulatory interactions and metabolicreconstruction but also provides one of the first exampleswhere the possible scenario of evolution of severalsystems regulating one metabolic pathway could be tentativelyreconstructed
Figure 7 Regulation of the methionine biosynthesis and transport genes in bacteria by S-box riboswitch (A) T-box structure (B) unknown transcriptional regulatorbinding the MET-box operator (C) activator MetR (D) and repressor MetJ (E) The mechanisms of the S-box and T-box regulation involve the formation ofalternative RNA structures SAM stabilizes the repressing conformation (blunt end) in (A) whereas in (B) the anti-repressing conformation (arrow) folds in thepresence of uncharged methionyl-tRNA The regions involved in alternative interactions are shown by bold lines or curves Poly-U tracts in terminators are shownabove the lsquoRNA linersquo Large arrows in (CndashE) denote regulated genes or operons
Nucleic Acids Research 2004 Vol 32 No 11 3351
SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

SUPPLEMENTARY MATERIAL
Supplementary Material is available at NAR Online
ACKNOWLEDGEMENTS
We are grateful to Vadim Brodyansky for the MetJ recognitionprofile and to Alexandra Rachmaninova Valerie de Crecy-Lagard and Andrei Osterman for discussions We are gratefulto Isabelle Martin-Verstraete for critical reading of the manu-script and many helpful suggestions This study was partiallysupported by grants from the Ludwig Institute for CancerResearch (CRDF RBO-1268) the Howard Hughes MedicalInstitute (55000309) and the Russian Fund of BasicResearch (04-04-49361-a)
REFERENCES
1 RavanelS GakiereB JobD and DouceR (1998) The specific featuresof methionine biosynthesis and metabolism in plants Proc NatlAcad Sci USA 95 7805ndash7812
2 SekowskaA KungHF and DanchinA (2000) Sulfur metabolism inEscherichia coli and related bacteria facts and fiction J Mol MicrobiolBiotechnol 2 145ndash177
3 ThomasD and Surdin-KerjanY (1997) Metabolism of sulfur aminoacids in Saccharomyces cerevisiae Microbiol Mol Biol Rev 61503ndash532
4 BelfaizaJ MartelA MargaritaD and Saint GironsI (1998) Directsulfhydrylation for methionine biosynthesis in Leptospira meyeriJ Bacteriol 180 250ndash255
5 KimJW KimHJ KimY LeeMS and LeeHS (2001) Properties ofthe Corynebacterium glutamicum metC gene encoding cystathionineb-lyase Mol Cells 11 220ndash225
6 LeeHS and HwangBJ (2003) Methionine biosynthesis and itsregulation in Corynebacterium glutamicum parallel pathways oftranssulfuration and direct sulfhydrylation Appl Microbiol Biotechnol101007s00253-003-1306-7
7 BrushA and PaulusH (1971) The enzymic formation of O-acetylhomoserine in Bacillus subtilis and its regulation by methionine andS-adenosylmethionine Biochem Biophys Res Commun 45 735ndash741
8 GreenRC (1994) Biosynthesis of methionine In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 542ndash561
9 BourhyP MartelA MargaritaD Saint GironsI and BelfaizaJ(1997) Homoserine O-acetyltransferase involved in the Leptospirameyeri methionine biosynthetic pathway is not feedback inhibitedJ Bacteriol 179 4396ndash4398
10 AugerS YuenWH DanchinA and Martin-VerstraeteI (2002)The metIC operon involved in methionine biosynthesis inBacillus subtilis is controlled by transcription antiterminationMicrobiology 148 507ndash518
11 HwangBJ YeomHJ KimY and LeeHS (2002) Corynebacteriumglutamicum utilizes both transsulfuration and direct sulfhydrylationpathways for methionine biosynthesis J Bacteriol 184 1277ndash1286
12 GrundyFJ and HenkinTM (2002) Synthesis of serine glycinecysteine and methionine In SonensheinAL HochJA and LosickR(eds) Bacillus subtilis and its Relatives From Genes to Cells AmericanSociety for Microbiology Washington DC pp 245ndash254
13 MerlinC GardinerG DurandS and MastersM (2002) TheEscherichia coli metD locus encodes an ABC transporter which includesAbc (MetN) YaeE (MetI) and YaeC (MetQ) J Bacteriol 1845513ndash5517
14 GalJ SzvetnikA SchnellR and KalmanM (2002) The metDD-methionine transporter locus of Escherichia coli is an ABC transportergene cluster J Bacteriol 184 4930ndash4932
15 ZhangZ FeigeJN ChangAB AndersonIJ BrodianskiVMVitreschakAG GelfandMS and SaierMHJr (2003) A transporter ofEscherichia coli specific for L- and D-methionine is the prototype
for a new family within the ABC superfamily Arch Microbiol180 88ndash100
16 HulloMF AugerS DassaE DanchinA and Martin-VerstraeteI(2004) The metNPQ operon of Bacillus subtilis encodes an ABCpermease transporting methionine sulfoxide D- and L-methionineRes Microbiol 155 80ndash86
17 SekowskaA BertinP and DanchinA (1998) Characterization ofpolyamine synthesis pathway in Bacillus subtilis 168 Mol Microbiol29 851ndash858
18 Della RagioneF PorcelliM Carteni-FarinaM ZappiaV andPeggAE (1985) Escherichia coli S-adenosylhomocysteine50-methylthioadenosine nucleosidase Purification substrate specificityand mechanism of action Biochem J 232 335ndash341
19 SekowskaA and DanchinA (1999) Identification of yrrU as themethylthioadenosine nucleosidase gene in Bacillus subtilis DNA Res6 255ndash264
20 ZhouD and WhiteRH (1991) Transsulfuration in archaebacteriaJ Bacteriol 173 3250ndash3251
21 MurphyBA GrundyFJ and HenkinTM (2002) Prediction of genefunction in methylthioadenosine recycling from regulatory signalsJ Bacteriol 184 2314ndash2318
22 SekowskaA and DanchinA (2002) The methionine salvage pathway inBacillus subtilis BMC Microbiol 2 8
23 SekowskaA DenervaudV AshidaH MichoudK HaasDYokotaA and DanchinA (2004) Bacterial variations on the methioninesalvage pathway BMC Microbiol 4 9
24 GrundyFJ and HenkinTM (1998) The S box regulon a new globaltranscription termination control system for methionine and cysteinebiosynthesis genes in gram-positive bacteria Mol Microbiol 30737ndash749
25 GrundyFJ and HenkinTM (2003) The T box and S box transcriptiontermination control systems Front Biosci 8 d20ndashd31
26 McDanielBA GrundyFJ ArtsimovitchI and HenkinTM (2003)Transcription termination control of the S box system directmeasurement of S-adenosylmethionine by the leader RNA Proc NatlAcad Sci USA 100 3083ndash3088
27 EpshteinV MironovAS and NudlerE (2003) The riboswitch-mediated control of sulfur metabolism in bacteria Proc Natl Acad SciUSA 100 5052ndash5056
28 WinklerWC NahviA SudarsanN BarrickJE and BreakerRR(2003) An mRNA structure that controls gene expression bybinding S-adenosylmethionine Nat Struct Biol 10 701ndash707
29 GrundyFJ MoirTR HaldemanMT and HenkinTM (2002)Sequence requirements for terminators and antiterminators in theT box transcription antitermination system disparity betweenconservation and functional requirements Nucleic Acids Res 301646ndash1655
30 PutzerH CondonC Brechemier-BaeyD BritoR and Grunberg-ManagoM (2002) Transfer RNA-mediated antitermination in vitroNucleic Acids Res 30 3026ndash3033
31 HenkinTM (1994) tRNA-directed transcription antiterminationMol Microbiol 13 381ndash387
32 VitreschakAG RodionovDA MironovAA and GelfandMS(2004) Riboswitches the oldest mechanism for the regulation of geneexpression Trends Genet 20 44ndash50
33 RodionovDA MironovAA and GelfandMS (2002) Conservation ofthe biotin regulon and the BirA regulatory signal in Eubacteria andArchaea Genome Res 12 1507ndash1516
34 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Comparative genomics of the vitamin B12 metabolismand regulation in prokaryotes J Biol Chem 278 41148ndash41159
35 RodionovDA VitreschakAG MironovAA and GelfandMS(2003) Regulation of lysine biosynthesis and transport genes inbacteria yet another RNA riboswitch Nucleic Acids Res 316748ndash6757
36 OstermanA and OverbeekR (2003) Missing genes in metabolicpathways a comparative genomics approach Curr Opin Chem Biol7 238ndash251
37 RivasE and EddySR (2001) Noncoding RNA gene detection usingcomparative sequence analysis BMC Bioinformatics 2 8
38 BensonDA Karsch-MizrachiI LipmanDJ OstellJ RappBA andWheelerDL (2000) GenBank Nucleic Acids Res 28 15ndash18
39 OverbeekR LarsenN WalunasT DrsquoSouzaM PuschGSelkovEJr LioliosK JoukovV KaznadzeyD AndersonIet al
3352 Nucleic Acids Research 2004 Vol 32 No 11
(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353

(2003) The ERGO genome analysis and discovery system Nucleic AcidsRes 31 164ndash171
40 VitreschakAG MironovAA and GelfandMS (2001) TheRNApattern program searching for RNA secondary structure by thepattern rule Proceedings of the 3rd International Conference onlsquoComplex Systems Control and Modeling Problemsrsquo September 492001 The Institute of Control of Complex Systems Samara Russiapp 623ndash625
41 BilloudB KonticM and ViariA (1996) Palingol a declarativeprogramming language to describe nucleic acidsrsquo secondary structuresand to scan sequence database Nucleic Acids Res 24 1395ndash1403
42 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nucleic Acids Res 31 3406ndash3415
43 MathewsDH SabinaJ ZukerM and TurnerDH (1999) Expandedsequence dependence of thermodynamic parameters improves predictionof RNA secondary structure J Mol Biol 288 911ndash940
44 GelfandMS KooninEV and MironovAA (2000) Prediction oftranscription regulatory sites in Archaea by a comparative genomicapproach Nucleic Acids Res 28 695ndash705
45 MironovAA KooninEV RoytbergMA and GelfandMS (1999)Computer analysis of transcription regulatory patterns in completelysequenced bacterial genomes Nucleic Acids Res 27 2981ndash2989
46 SchneiderTD StormoGD GoldL and EhrenfeuchtA (1986)Information content of binding sites on nucleotide sequencesJ Mol Biol 188 415ndash431
47 MironovAA VinokurovaNP and GelfandMS (2000)GenomeExplorer software for analysis of complete bacterial genomesMol Biol 34 222ndash231
48 TatusovRL NataleDA GarkavtsevIV TatusovaTAShankavaramUT RaoBS KiryutinB GalperinMYFedorovaND and KooninEV (2001) The COG database newdevelopments in phylogenetic classification of proteins from completegenomes Nucleic Acids Res 29 22ndash28
49 FelsensteinJ (1981) Evolutionary trees from DNA sequences amaximum likelihood approach J Mol Evol 17 368ndash376
50 ThompsonJD GibsonTJ PlewniakF JeanmouginF andHigginsDG (1997) The CLUSTAL_X windows interface flexiblestrategies for multiple sequence alignment aided by quality analysis toolsNucleic Acids Res 25 4876ndash4882
51 ApweilerR AttwoodTK BairochA BatemanA BirneyEBiswasM BucherP CeruttiL CorpetF CroningMD et al(2001) The InterPro database an integrated documentation resourcefor protein families domains and functional sites Nucleic Acids Res29 37ndash40
52 BatemanA BirneyE CerrutiL DurbinR EtwillerL EddySRGriffiths-JonesS HoweKL MarshallM and SonnhammerEL(2002) The Pfam protein families database Nucleic Acids Res 30276ndash280
53 ShelverD RajagopalL HarrisTO and RubensCE (2003) MtaR aregulator of methionine transport is critical for survival of group BStreptococcus in vivo J Bacteriol 185 6592ndash6599
54 MansillaMC AlbanesiD and de MendozaD (2000) Transcriptionalcontrol of the sulfur-regulated cysH operon containing genes involved inL-cysteine biosynthesis in Bacillus subtilis J Bacteriol 182 5885ndash5892
55 ChadwickLH McCandlessSE SilvermanGL SchwartzSWestawayD and NadeauJH (2000) Betainendashhomocysteinemethyltransferase-2 cDNA cloning gene sequence physical mappingand expression of the human and mouse genes Genomics 70 66ndash73
56 DixonMM HuangS MatthewsRG and LudwigM (1996) Thestructure of the C-terminal domain of methionine synthase presentingS-adenosylmethionine for reductive methylation of B12 Structure4 1263ndash1275
57 MatthewsRG (1994) One-carbon metabolism In NeidhardtFC (ed)Escherichia coli and Salmonella Cellular and Molecular BiologyAmerican Society for Microbiology Washington DC pp 600ndash612
58 RavcheevDA GelrsquofandMS MironovAA and RakhmaninovaAB(2002) Purine regulon of gamma-proteobacteria a detailed descriptionGenetika 38 1203ndash1214
59 PaninaEM VitreschakAG MironovAA and GelfandMS (2003)Regulation of biosynthesis and transport of aromatic amino acidsin low-GC Gram-positive bacteria FEMS Microbiol Lett 222211ndash220
60 WeiY GuffantiAA ItoM and KrulwichTA (2000) Bacillus subtilisYqkI is a novel malicNa+-lactate antiporter that enhances growthon malate at low protonmotive force J Biol Chem 27530287ndash30292
61 MarcosAT KosalkovaK CardozaRE FierroF GutierrezS andMartinJF (2001) Characterization of the reverse transsulfuration genemecB of Acremonium chrysogenum which encodes a functionalcystathionine-gamma-lyase Mol Gen Genet 264 746ndash754
62 HilgersMT and LudwigML (2001) Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site Proc Natl Acad SciUSA 98 11169ndash11174
63 SuretteMG MillerMB and BasslerBL (1999) Quorum sensing inEscherichia coli Salmonella typhimurium and Vibrio harveyi a newfamily of genes responsible for autoinducer production Proc Natl AcadSci USA 96 1639ndash1644
64 YocumRR PerkinsJB HowittCL and PeroJ (1996) Cloning andcharacterization of the metE gene encoding S-adenosylmethioninesynthetase from Bacillus subtilis J Bacteriol 178 4604ndash4610
65 BarrettAJ and RawlingsND (1995) Evolutionary families ofmetallopeptidases Meth Enzymol 248 183ndash228
66 AlexanderFW SandmeierE MehtaPK and ChristenP (1994)Evolutionary relationships among pyridoxal-50-phosphate-dependentenzymes Regio-specific alpha beta and gamma families Eur JBiochem 219 953ndash960
Nucleic Acids Research 2004 Vol 32 No 11 3353





























