Embed Size (px)
Citation preview

Functional Expression of the Signaling Complex Sensory RhodopsinII/Transducer II from Halobacterium salinarum in Escherichia coli †
Young-Jun Kim1, Igor Chizhov2 and Martin Engelhard*1
1Max-Planck-Institut fur molekulare Physiology, Dortmund, Germany2Medizinische Hochschule Hannover, Institut fur Biophysik. Chemie, Hannover, Germany
Received 15 August 2008, accepted 3 September 2008, DOI: 10.1111 ⁄ j.1751-1097.2008.00470.x
ABSTRACT
Sensory rhodopsin II, a photoreceptor from Halobacterium
salinarum (HsSRII), in complex with its cognate transducer
protein (HsHtrII) triggers the photophobic response via a
cytoplasmic two-component signaling cascade. HsHtrII possess
in addition to the HsSRII binding and the cytoplasmic domains
an extracellular serine-receptor domain. Here we describe the
properties of HsSRII and HsHtrII and those of various
shortened transducer analogs, heterologously expressed in
Escherichia coli. HsSRII displays the photocycle typical of
archaeal photosensors with prolonged kinetics. Using an iso-
thermal titration calorimetric analysis for this complex a
dissociation constant of 1.1 lMM was obtained similar to that of
the corresponding transducer/receptor pair from Natronobacte-
rium pharaonis. A shortened transducer lacking the extracellular
and cytoplasmic domain is also sufficient to bind the receptor
with a slightly lower affinity. The dissociation constant of serine
binding to the extracellular domain was determined to be about
5 lMM. This result is in line with the proposal that the extracel-
lular domain indeed is a serine receptor.
INTRODUCTION
Phototaxis of the haloarchaeon Halobacterium salinarum ismediated by two sensory rhodopsins (HsSRI; HsSRII). These
photoreceptors belong to the large family of microbialrhodopsins (1,2) which enable the bacteria to seek conditionsfor optimal function of the two ion pumps bacteriorhodopsin
and halorhodopsin and to avoid photo-oxidative stress.Sensory rhodopsin I (HsSRI), in a one-photon reaction,triggers a photophilic response at wavelengths above 500 nm.
In a two-photon reaction additional blue light induces aphotophobic response (3). The other receptor sensory rho-dopsin II (HsSRII or phoborhodopsin) is responsible for thephotophobic response of H. salinarum near 490 nm. Microbial
rhodopsins such as HsSRI and HsSRII are seven-helixmembrane proteins with retinal as cofactor bound via aprotonated Schiff base to the �-amino group of a lysine
residue. Specific protein-chromophore interactions regulate the
color of the pigment (i.e. kmax = 580 nm for HsSRI and487 nm for HsSRII) (4). Sensors, in contrast to ion pumps, arebound to cognate transducers (HsHtrI, HsHtrII, respectively)
which receive the signal from their activated receptors andtransmit it to the cytoplasmic two-component signalingcascade. Other common features of microbial rhodopsins are
their photocycles initiated by the light-activated trans-13-cisisomerization of retinal. Generally, sensors have longer photo-cycle turnovers than ion pumps (seconds versus 10 ms).
Most knowledge about SRII-type sensors has been accumu-
lated for SRII from Natronobacterium pharaonis (NpSRII)(reviewed in refs. [5,6]) due to its higher biochemical stability (7)and the fact that it can be heterologously expressed in Escher-
ichia coli (8,9). The crystal structure ofNpSRII (10,11) as well asthat of the sensor-transducer (NpHtrII) complex (12) has beendetermined. The major difference between theH. salinarum and
theN. pharaonis systems is found in the domain organization ofthe transducers (13,14). Whereas NpHtrII consists of a trans-membrane sensor-binding domain and a long rod-shapedcytoplasmic domain, inHsHtrII an extracellular serine-receptor
domain is located between transmembrane helices 1 and2 (TM1,TM2) (14,15). Apparently, the HsSRII ⁄HsHtrII complex, likeHsSRI, possesses dual functionality. This observation is impor-
tant in relation to common mechanism of signal transfer inphototaxis and chemotaxis.
Recently, HsSRII has been cloned, heterologously
expressed in E. coli, and functionally characterized (16). Herewe describe the characterization of both HsSRII and HsHtrIIisolated and purified from E. coli membranes. The photocycle
properties of the sensor and the complex are quite similar andcomparable to that of NpSRII. The formation of the complexin detergents occurs with a stoichiometry of 1:1 and aKd = 1.1 lMM. The extracellular domain binds serine quite
tightly corroborating earlier physiological experiments (15).
MATERIALS AND METHODS
All buffers were prepared with reagent grade chemicals and distilledwater that was subsequently treated with a Milli Q (Millipore,Bedford, MA) water purification system. Purple membrane lipids(PML) were isolated from purple membrane using published proce-dures (17,18).
Cloning and expression. Escherichia coli XL1-Blue was used as hostfor DNA manipulation. The HssopII, HshtrII genes, and the HshtrIIanalogs were amplified by PCR from genomic DNA extraction ofH. salinarum. The 5¢-primer introduced an NcoI site including the startcodon. The 3¢-primer added a coding region for seven histidines
†This paper is part of the Proceedings of the 13th International Conference onRetinal Proteins, Barcelona, Spain, 15–19 June 2008.
*Corresponding author email: [email protected](Martin Engelhard)
� 2008TheAuthors. JournalCompilation.TheAmericanSociety ofPhotobiology 0031-8655/09
Photochemistry and Photobiology, 2009, 85: 521–528
521
followed by two stop codons and a HindIII site was introduced. TheNcoI ⁄HindIII PCR fragments (HssrII, HshtrII[1-765], HshtrII[1-398],HshtrII[1-398]D38-280, HshtrII[1-765]D38-280) were cloned into thepET27bmod vector. The gene sequence was confirmed by DNAsequencing. In the constructs lacking the serine binding domain thetwo transmembrane helices were connected by DNA sequence corre-sponding to residues 44-59 from NphtrII by the double-joint PCRmethod (19).
HsSRII andHsHtrII(1-398) and its analogs were expressed in E. coliBL21(DE3)Rosetta essentially as described (8,9).Cellswere grown inLBmedium containing 50 mg L)1 kanamycin and 30mg L)1 chloramphe-nicol at 37�C. Protein expression was induced by using isopropyl-b-DD-thiogalactopyranoside (IPTG). For production of HsSRII, 1 mMM IPTGand all-trans retinal (10 lMM; Sigma) were added. Cells were harvested bycentrifugation and lyzed in aMicrofluidizer (Microfluidics Corporation,Newton, MA). Membranes were sedimented at 100 000 g for 1.5 h at4�C and solubilized in buffer A (2% [wt ⁄ vol] n-dodecyl-b-DD-maltoside[DDM] (Calbiochem), 4 MM NaCl, 50 mMM MES, pH 6) for 16 h at 4�C.After centrifugation of the solubilized membrane protein fraction(100 000 g, 1.5 h, 4�C) the supernatant was purified using Ni-NTAchromatography. The Ni-NTA superflow (Qiagen, Hilden, Germany)was pre-equilibrated with buffer B (4 MM NaCl, 50 mMM MES, pH 6.0,0.05%DDM) for 1 h at 4�Cwhile slowly shaking the vessel. The loadedresin was filled onto a chromatography column and washed extensivelywith bufferC (buffer B + 60mMM imidazole). PurifiedHis-taggedproteinwas then eluted from the column with buffer D (buffer B + 250 mMM
imidazole). Thereafter, imidazole was removed by using an Econo-Pac10DG desalting column (Bio-Rad). HsSRII concentration was deter-mined by the optical density at 487 nm (� = 48 000 MM
)1 cm)1 [20]). Theyield of HsHtrII was calculated from the optical density at 280 nm(� = 24 000 MM
)1 cm)1).Laser flash photolysis and data analysis. The photocycle experiments
and the analysis of the data were carried out as described by Chizhovet al. (21,22). HsSRII reconstituted into PM lipids was immobilized ingels (16.5% acrylamide) to measure the pH dependency of theabsorption maximum and to determine the pKa of Asp73. Prior tothese experiments the gel slice was equilibrated with the appropriatebuffer (4 MM NaCl, 50 mMM MES, pH ranging from 6 to 1) for at least30 min. The transient absorption changes were recorded at 360, 490and 540 ⁄ 550 nm wavelength.
Isothermal titration calorimetry. The isothermal titration calorimet-ric (ITC) measurements were performed basically as described (23)using an AVP-ITC MicroCalorimeter (MicroCal). Protein sampleswere dialyzed at 4�C overnight against degassed buffer containing 4 MM
NaCl, 50 mMM MES, pH 6, 0.05% DDM (buffer A) prior to theexperiments. HsSRII and serine were injected into the reactionchamber (1.5 mL buffer A, 20 lMM transducer) in 12 lL increments(in 3 min intervals) at a concentration of between 300 and 500 lMM. Thesample was stirred at 290 rpm. Control experiments for heats ofmixing and dilution were performed under identical conditions andused for data correction in subsequent analysis. Titrations were carriedout at temperatures from 20 to 35�C. Data acquisition and subsequentnonlinear regression analysis were performed in terms of a simplebinding model, using ORIGIN software package.
RESULTS AND DISCUSSION
Expression and purification of HsSRII, HsHtrII and
HsHtrII analogs
In order to gain insight into the properties of sensoryrhodopsin II and its interaction with its cognate transducer,
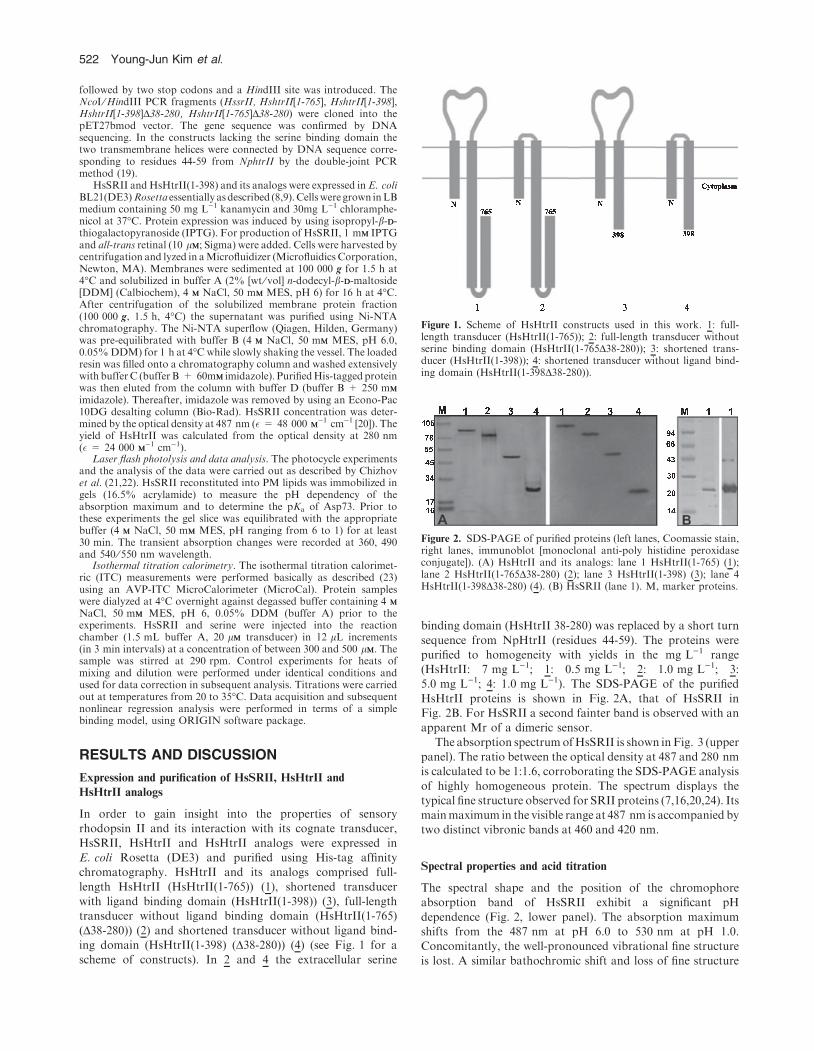
HsSRII, HsHtrII and HsHtrII analogs were expressed inE. coli Rosetta (DE3) and purified using His-tag affinitychromatography. HsHtrII and its analogs comprised full-
length HsHtrII (HsHtrII(1-765)) (1), shortened transducerwith ligand binding domain (HsHtrII(1-398)) (3), full-lengthtransducer without ligand binding domain (HsHtrII(1-765)
(D38-280)) (2) and shortened transducer without ligand bind-ing domain (HsHtrII(1-398) (D38-280)) (4) (see Fig. 1 for ascheme of constructs). In 2 and 4 the extracellular serine
binding domain (HsHtrII 38-280) was replaced by a short turn
sequence from NpHtrII (residues 44-59). The proteins werepurified to homogeneity with yields in the mg L)1 range(HsHtrII: 7 mg L)1; 1: 0.5 mg L)1; 2: 1.0 mg L)1; 3:5.0 mg L)1; 4: 1.0 mg L)1). The SDS-PAGE of the purified
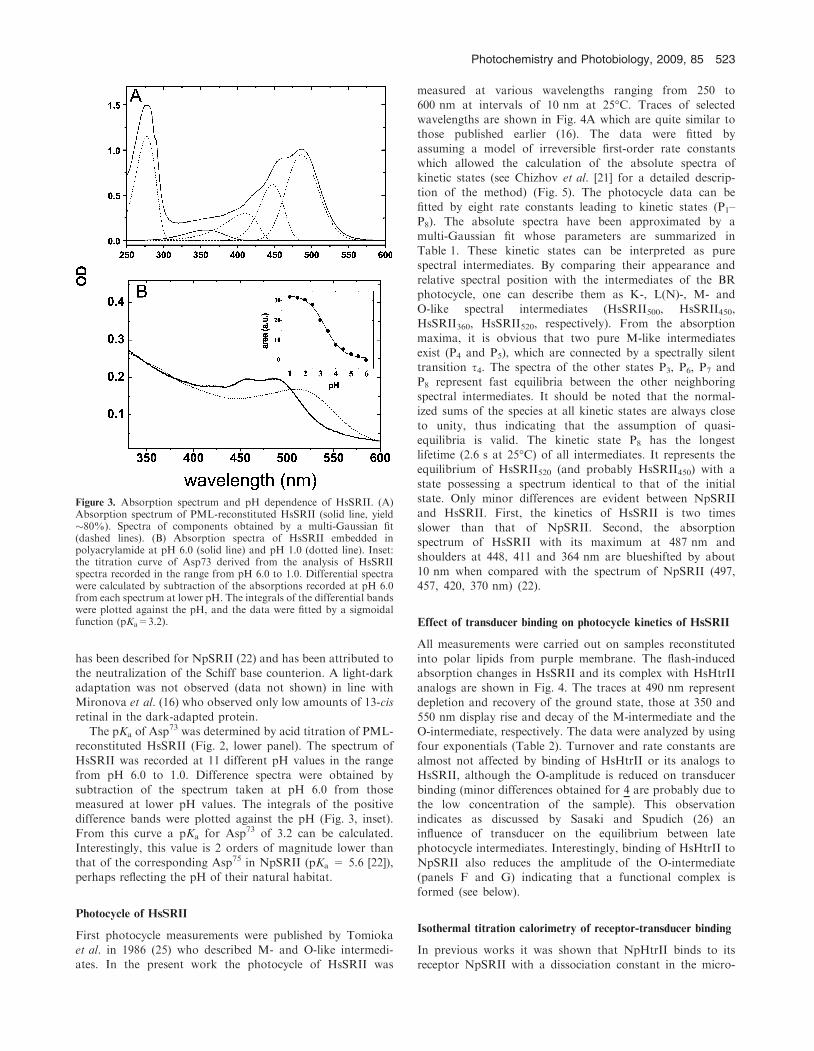
HsHtrII proteins is shown in Fig. 2A, that of HsSRII inFig. 2B. For HsSRII a second fainter band is observed with anapparent Mr of a dimeric sensor.
The absorption spectrumofHsSRII is shown inFig. 3 (upperpanel). The ratio between the optical density at 487 and 280 nmis calculated to be 1:1.6, corroborating the SDS-PAGE analysis
of highly homogeneous protein. The spectrum displays thetypical fine structure observed for SRII proteins (7,16,20,24). Itsmainmaximum in the visible range at 487 nm is accompanied bytwo distinct vibronic bands at 460 and 420 nm.
Spectral properties and acid titration
The spectral shape and the position of the chromophoreabsorption band of HsSRII exhibit a significant pH
dependence (Fig. 2, lower panel). The absorption maximumshifts from the 487 nm at pH 6.0 to 530 nm at pH 1.0.Concomitantly, the well-pronounced vibrational fine structure
is lost. A similar bathochromic shift and loss of fine structure
Figure 1. Scheme of HsHtrII constructs used in this work. 1: full-length transducer (HsHtrII(1-765)); 2: full-length transducer withoutserine binding domain (HsHtrII(1-765D38-280)); 3: shortened trans-ducer (HsHtrII(1-398)); 4: shortened transducer without ligand bind-ing domain (HsHtrII(1-398D38-280)).
Figure 2. SDS-PAGE of purified proteins (left lanes, Coomassie stain,right lanes, immunoblot [monoclonal anti-poly histidine peroxidaseconjugate]). (A) HsHtrII and its analogs: lane 1 HsHtrII(1-765) (1);lane 2 HsHtrII(1-765D38-280) (2); lane 3 HsHtrII(1-398) (3); lane 4HsHtrII(1-398D38-280) (4). (B) HsSRII (lane 1). M, marker proteins.
522 Young-Jun Kim et al.
has been described for NpSRII (22) and has been attributed tothe neutralization of the Schiff base counterion. A light-darkadaptation was not observed (data not shown) in line withMironova et al. (16) who observed only low amounts of 13-cis
retinal in the dark-adapted protein.The pKa of Asp73 was determined by acid titration of PML-
reconstituted HsSRII (Fig. 2, lower panel). The spectrum of
HsSRII was recorded at 11 different pH values in the rangefrom pH 6.0 to 1.0. Difference spectra were obtained bysubtraction of the spectrum taken at pH 6.0 from those
measured at lower pH values. The integrals of the positivedifference bands were plotted against the pH (Fig. 3, inset).From this curve a pKa for Asp73 of 3.2 can be calculated.
Interestingly, this value is 2 orders of magnitude lower thanthat of the corresponding Asp75 in NpSRII (pKa = 5.6 [22]),perhaps reflecting the pH of their natural habitat.
Photocycle of HsSRII
First photocycle measurements were published by Tomiokaet al. in 1986 (25) who described M- and O-like intermedi-ates. In the present work the photocycle of HsSRII was
measured at various wavelengths ranging from 250 to600 nm at intervals of 10 nm at 25�C. Traces of selectedwavelengths are shown in Fig. 4A which are quite similar tothose published earlier (16). The data were fitted by
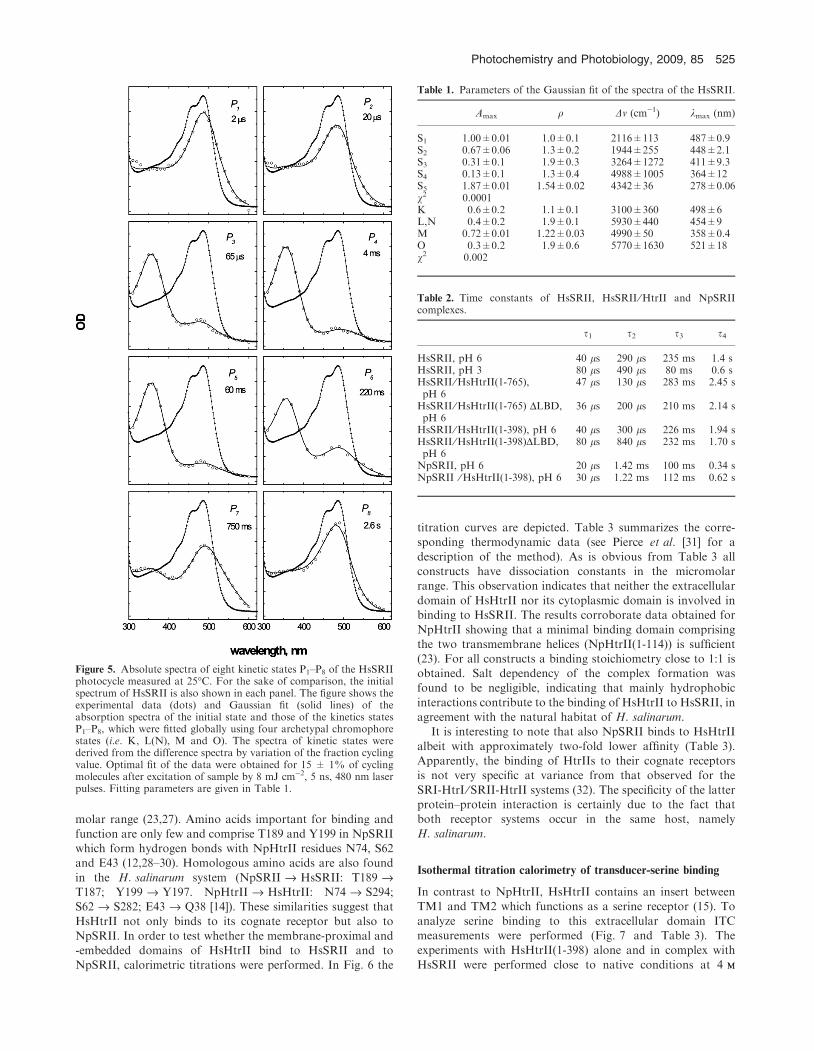
assuming a model of irreversible first-order rate constantswhich allowed the calculation of the absolute spectra ofkinetic states (see Chizhov et al. [21] for a detailed descrip-
tion of the method) (Fig. 5). The photocycle data can befitted by eight rate constants leading to kinetic states (P1–P8). The absolute spectra have been approximated by a
multi-Gaussian fit whose parameters are summarized inTable 1. These kinetic states can be interpreted as purespectral intermediates. By comparing their appearance and
relative spectral position with the intermediates of the BRphotocycle, one can describe them as K-, L(N)-, M- andO-like spectral intermediates (HsSRII500, HsSRII450,HsSRII360, HsSRII520, respectively). From the absorption
maxima, it is obvious that two pure M-like intermediatesexist (P4 and P5), which are connected by a spectrally silenttransition s4. The spectra of the other states P3, P6, P7 and
P8 represent fast equilibria between the other neighboringspectral intermediates. It should be noted that the normal-ized sums of the species at all kinetic states are always close
to unity, thus indicating that the assumption of quasi-equilibria is valid. The kinetic state P8 has the longestlifetime (2.6 s at 25�C) of all intermediates. It represents theequilibrium of HsSRII520 (and probably HsSRII450) with a
state possessing a spectrum identical to that of the initialstate. Only minor differences are evident between NpSRIIand HsSRII. First, the kinetics of HsSRII is two times
slower than that of NpSRII. Second, the absorptionspectrum of HsSRII with its maximum at 487 nm andshoulders at 448, 411 and 364 nm are blueshifted by about
10 nm when compared with the spectrum of NpSRII (497,457, 420, 370 nm) (22).
Effect of transducer binding on photocycle kinetics of HsSRII
All measurements were carried out on samples reconstitutedinto polar lipids from purple membrane. The flash-inducedabsorption changes in HsSRII and its complex with HsHtrIIanalogs are shown in Fig. 4. The traces at 490 nm represent
depletion and recovery of the ground state, those at 350 and550 nm display rise and decay of the M-intermediate and theO-intermediate, respectively. The data were analyzed by using
four exponentials (Table 2). Turnover and rate constants arealmost not affected by binding of HsHtrII or its analogs toHsSRII, although the O-amplitude is reduced on transducer
binding (minor differences obtained for 4 are probably due tothe low concentration of the sample). This observationindicates as discussed by Sasaki and Spudich (26) aninfluence of transducer on the equilibrium between late
photocycle intermediates. Interestingly, binding of HsHtrII toNpSRII also reduces the amplitude of the O-intermediate(panels F and G) indicating that a functional complex is
formed (see below).
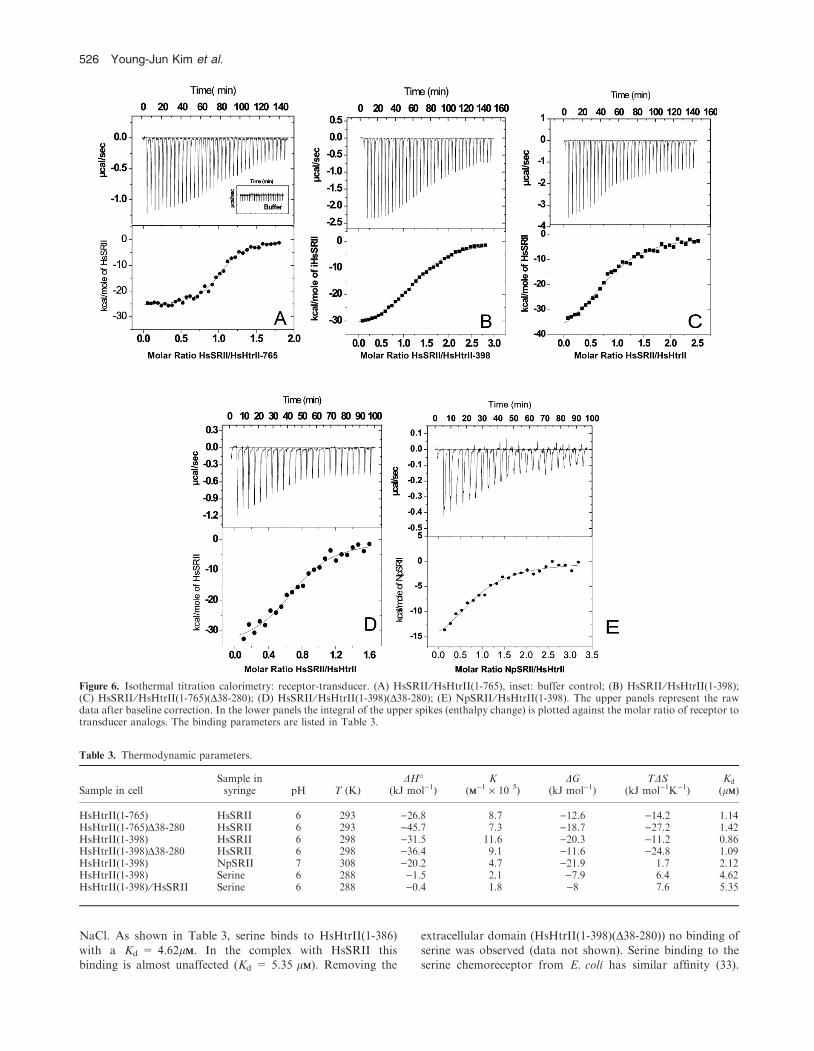
Isothermal titration calorimetry of receptor-transducer binding
In previous works it was shown that NpHtrII binds to its
receptor NpSRII with a dissociation constant in the micro-
Figure 3. Absorption spectrum and pH dependence of HsSRII. (A)Absorption spectrum of PML-reconstituted HsSRII (solid line, yield�80%). Spectra of components obtained by a multi-Gaussian fit(dashed lines). (B) Absorption spectra of HsSRII embedded inpolyacrylamide at pH 6.0 (solid line) and pH 1.0 (dotted line). Inset:the titration curve of Asp73 derived from the analysis of HsSRIIspectra recorded in the range from pH 6.0 to 1.0. Differential spectrawere calculated by subtraction of the absorptions recorded at pH 6.0from each spectrum at lower pH. The integrals of the differential bandswere plotted against the pH, and the data were fitted by a sigmoidalfunction (pKa=3.2).
Photochemistry and Photobiology, 2009, 85 523
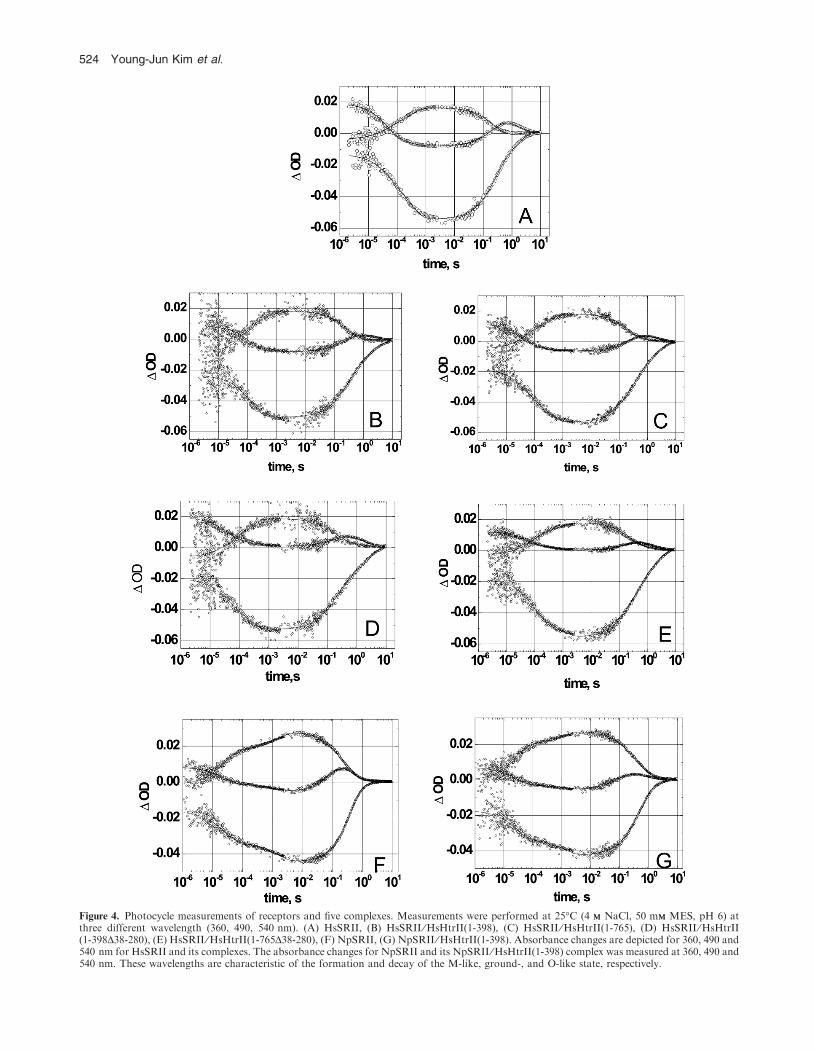
Figure 4. Photocycle measurements of receptors and five complexes. Measurements were performed at 25�C (4 MM NaCl, 50 mMM MES, pH 6) atthree different wavelength (360, 490, 540 nm). (A) HsSRII, (B) HsSRII ⁄HsHtrII(1-398), (C) HsSRII ⁄HsHtrII(1-765), (D) HsSRII ⁄HsHtrII(1-398D38-280), (E) HsSRII ⁄HsHtrII(1-765D38-280), (F) NpSRII, (G) NpSRII ⁄HsHtrII(1-398). Absorbance changes are depicted for 360, 490 and540 nm for HsSRII and its complexes. The absorbance changes for NpSRII and its NpSRII ⁄HsHtrII(1-398) complex was measured at 360, 490 and540 nm. These wavelengths are characteristic of the formation and decay of the M-like, ground-, and O-like state, respectively.
524 Young-Jun Kim et al.
molar range (23,27). Amino acids important for binding and
function are only few and comprise T189 and Y199 in NpSRIIwhich form hydrogen bonds with NpHtrII residues N74, S62and E43 (12,28–30). Homologous amino acids are also found
in the H. salinarum system (NpSRII fi HsSRII: T189 fiT187; Y199 fi Y197. NpHtrII fi HsHtrII: N74 fi S294;S62 fi S282; E43 fi Q38 [14]). These similarities suggest that
HsHtrII not only binds to its cognate receptor but also toNpSRII. In order to test whether the membrane-proximal and-embedded domains of HsHtrII bind to HsSRII and toNpSRII, calorimetric titrations were performed. In Fig. 6 the
titration curves are depicted. Table 3 summarizes the corre-sponding thermodynamic data (see Pierce et al. [31] for a
description of the method). As is obvious from Table 3 allconstructs have dissociation constants in the micromolarrange. This observation indicates that neither the extracellular
domain of HsHtrII nor its cytoplasmic domain is involved inbinding to HsSRII. The results corroborate data obtained forNpHtrII showing that a minimal binding domain comprising
the two transmembrane helices (NpHtrII(1-114)) is sufficient(23). For all constructs a binding stoichiometry close to 1:1 isobtained. Salt dependency of the complex formation wasfound to be negligible, indicating that mainly hydrophobic
interactions contribute to the binding of HsHtrII to HsSRII, inagreement with the natural habitat of H. salinarum.
It is interesting to note that also NpSRII binds to HsHtrII
albeit with approximately two-fold lower affinity (Table 3).Apparently, the binding of HtrIIs to their cognate receptorsis not very specific at variance from that observed for the
SRI-HtrI ⁄SRII-HtrII systems (32). The specificity of the latterprotein–protein interaction is certainly due to the fact thatboth receptor systems occur in the same host, namelyH. salinarum.
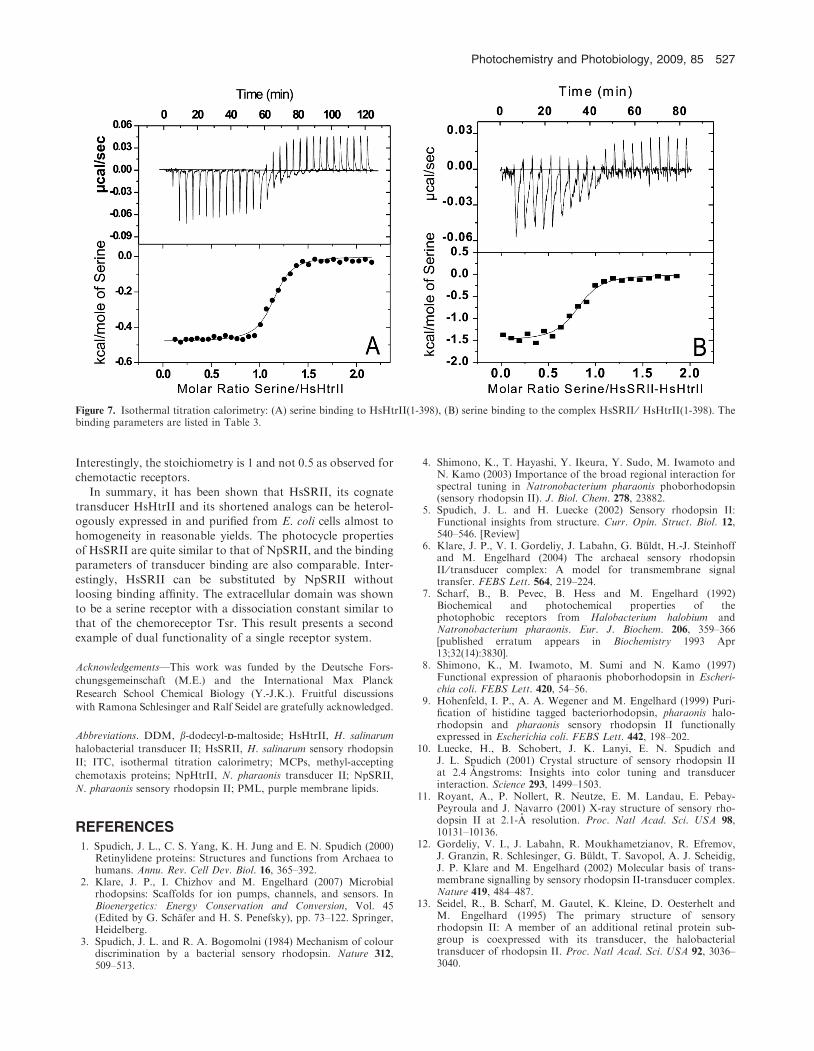
Isothermal titration calorimetry of transducer-serine binding
In contrast to NpHtrII, HsHtrII contains an insert betweenTM1 and TM2 which functions as a serine receptor (15). To
analyze serine binding to this extracellular domain ITCmeasurements were performed (Fig. 7 and Table 3). Theexperiments with HsHtrII(1-398) alone and in complex with
HsSRII were performed close to native conditions at 4 MM
Figure 5. Absolute spectra of eight kinetic states P1–P8 of the HsSRIIphotocycle measured at 25�C. For the sake of comparison, the initialspectrum of HsSRII is also shown in each panel. The figure shows theexperimental data (dots) and Gaussian fit (solid lines) of theabsorption spectra of the initial state and those of the kinetics statesP1–P8, which were fitted globally using four archetypal chromophorestates (i.e. K, L(N), M and O). The spectra of kinetic states werederived from the difference spectra by variation of the fraction cyclingvalue. Optimal fit of the data were obtained for 15 ± 1% of cyclingmolecules after excitation of sample by 8 mJ cm)2, 5 ns, 480 nm laserpulses. Fitting parameters are given in Table 1.
Table 1. Parameters of the Gaussian fit of the spectra of the HsSRII.
Amax q Dv (cm)1) kmax (nm)
S1 1.00±0.01 1.0±0.1 2116±113 487±0.9S2 0.67±0.06 1.3±0.2 1944±255 448±2.1S3 0.31±0.1 1.9±0.3 3264±1272 411±9.3S4 0.13±0.1 1.3±0.4 4988±1005 364±12S5 1.87±0.01 1.54±0.02 4342±36 278±0.06v2 0.0001K 0.6±0.2 1.1±0.1 3100±360 498±6L,N 0.4±0.2 1.9±0.1 5930±440 454±9M 0.72±0.01 1.22±0.03 4990±50 358±0.4O 0.3±0.2 1.9±0.6 5770±1630 521±18v2 0.002
Table 2. Time constants of HsSRII, HsSRII ⁄HtrII and NpSRIIcomplexes.
s1 s2 s3 s4
HsSRII, pH 6 40 ls 290 ls 235 ms 1.4 sHsSRII, pH 3 80 ls 490 ls 80 ms 0.6 sHsSRII ⁄HsHtrII(1-765),pH 6
47 ls 130 ls 283 ms 2.45 s
HsSRII ⁄HsHtrII(1-765) DLBD,pH 6
36 ls 200 ls 210 ms 2.14 s
HsSRII ⁄HsHtrII(1-398), pH 6 40 ls 300 ls 226 ms 1.94 sHsSRII ⁄HsHtrII(1-398)DLBD,pH 6
80 ls 840 ls 232 ms 1.70 s
NpSRII, pH 6 20 ls 1.42 ms 100 ms 0.34 sNpSRII ⁄HsHtrII(1-398), pH 6 30 ls 1.22 ms 112 ms 0.62 s
Photochemistry and Photobiology, 2009, 85 525
NaCl. As shown in Table 3, serine binds to HsHtrII(1-386)
with a Kd = 4.62lMM. In the complex with HsSRII thisbinding is almost unaffected (Kd = 5.35 lMM). Removing the
extracellular domain (HsHtrII(1-398)(D38-280)) no binding of
serine was observed (data not shown). Serine binding to theserine chemoreceptor from E. coli has similar affinity (33).
Figure 6. Isothermal titration calorimetry: receptor-transducer. (A) HsSRII ⁄HsHtrII(1-765), inset: buffer control; (B) HsSRII ⁄HsHtrII(1-398);(C) HsSRII ⁄HsHtrII(1-765)(D38-280); (D) HsSRII ⁄HsHtrII(1-398)(D38-280); (E) NpSRII ⁄HsHtrII(1-398). The upper panels represent the rawdata after baseline correction. In the lower panels the integral of the upper spikes (enthalpy change) is plotted against the molar ratio of receptor totransducer analogs. The binding parameters are listed in Table 3.
Table 3. Thermodynamic parameters.
Sample in cellSample insyringe pH T (K)
DH�(kJ mol)1)
K(MM)1 · 10 5)
DG(kJ mol)1)
TDS(kJ mol)1K)1)
Kd
(lMM)
HsHtrII(1-765) HsSRII 6 293 )26.8 8.7 )12.6 )14.2 1.14HsHtrII(1-765)D38-280 HsSRII 6 293 )45.7 7.3 )18.7 )27.2 1.42HsHtrII(1-398) HsSRII 6 298 )31.5 11.6 )20.3 )11.2 0.86HsHtrII(1-398)D38-280 HsSRII 6 298 )36.4 9.1 )11.6 )24.8 1.09HsHtrII(1-398) NpSRII 7 308 )20.2 4.7 )21.9 1.7 2.12HsHtrII(1-398) Serine 6 288 )1.5 2.1 )7.9 6.4 4.62HsHtrII(1-398) ⁄HsSRII Serine 6 288 )0.4 1.8 )8 7.6 5.35
526 Young-Jun Kim et al.
Interestingly, the stoichiometry is 1 and not 0.5 as observed forchemotactic receptors.
In summary, it has been shown that HsSRII, its cognatetransducer HsHtrII and its shortened analogs can be heterol-ogously expressed in and purified from E. coli cells almost to
homogeneity in reasonable yields. The photocycle propertiesof HsSRII are quite similar to that of NpSRII, and the bindingparameters of transducer binding are also comparable. Inter-
estingly, HsSRII can be substituted by NpSRII withoutloosing binding affinity. The extracellular domain was shownto be a serine receptor with a dissociation constant similar to
that of the chemoreceptor Tsr. This result presents a secondexample of dual functionality of a single receptor system.
Acknowledgements—This work was funded by the Deutsche Fors-
chungsgemeinschaft (M.E.) and the International Max Planck
Research School Chemical Biology (Y.-J.K.). Fruitful discussions
with Ramona Schlesinger and Ralf Seidel are gratefully acknowledged.
Abbreviations. DDM, b-dodecyl-DD-maltoside; HsHtrII, H. salinarum
halobacterial transducer II; HsSRII, H. salinarum sensory rhodopsin
II; ITC, isothermal titration calorimetry; MCPs, methyl-accepting
chemotaxis proteins; NpHtrII, N. pharaonis transducer II; NpSRII,
N. pharaonis sensory rhodopsin II; PML, purple membrane lipids.
REFERENCES1. Spudich, J. L., C. S. Yang, K. H. Jung and E. N. Spudich (2000)
Retinylidene proteins: Structures and functions from Archaea tohumans. Annu. Rev. Cell Dev. Biol. 16, 365–392.
2. Klare, J. P., I. Chizhov and M. Engelhard (2007) Microbialrhodopsins: Scaffolds for ion pumps, channels, and sensors. InBioenergetics: Energy Conservation and Conversion, Vol. 45(Edited by G. Schafer and H. S. Penefsky), pp. 73–122. Springer,Heidelberg.
3. Spudich, J. L. and R. A. Bogomolni (1984) Mechanism of colourdiscrimination by a bacterial sensory rhodopsin. Nature 312,509–513.
4. Shimono, K., T. Hayashi, Y. Ikeura, Y. Sudo, M. Iwamoto andN. Kamo (2003) Importance of the broad regional interaction forspectral tuning in Natronobacterium pharaonis phoborhodopsin(sensory rhodopsin II). J. Biol. Chem. 278, 23882.
5. Spudich, J. L. and H. Luecke (2002) Sensory rhodopsin II:Functional insights from structure. Curr. Opin. Struct. Biol. 12,540–546. [Review]
6. Klare, J. P., V. I. Gordeliy, J. Labahn, G. Buldt, H.-J. Steinhoffand M. Engelhard (2004) The archaeal sensory rhodopsinII ⁄ transducer complex: A model for transmembrane signaltransfer. FEBS Lett. 564, 219–224.
7. Scharf, B., B. Pevec, B. Hess and M. Engelhard (1992)Biochemical and photochemical properties of thephotophobic receptors from Halobacterium halobium andNatronobacterium pharaonis. Eur. J. Biochem. 206, 359–366[published erratum appears in Biochemistry 1993 Apr13;32(14):3830].
8. Shimono, K., M. Iwamoto, M. Sumi and N. Kamo (1997)Functional expression of pharaonis phoborhodopsin in Escheri-chia coli. FEBS Lett. 420, 54–56.
9. Hohenfeld, I. P., A. A. Wegener and M. Engelhard (1999) Puri-fication of histidine tagged bacteriorhodopsin, pharaonis halo-rhodopsin and pharaonis sensory rhodopsin II functionallyexpressed in Escherichia coli. FEBS Lett. 442, 198–202.
10. Luecke, H., B. Schobert, J. K. Lanyi, E. N. Spudich andJ. L. Spudich (2001) Crystal structure of sensory rhodopsin IIat 2.4 Angstroms: Insights into color tuning and transducerinteraction. Science 293, 1499–1503.
11. Royant, A., P. Nollert, R. Neutze, E. M. Landau, E. Pebay-Peyroula and J. Navarro (2001) X-ray structure of sensory rho-dopsin II at 2.1-A resolution. Proc. Natl Acad. Sci. USA 98,10131–10136.
12. Gordeliy, V. I., J. Labahn, R. Moukhametzianov, R. Efremov,J. Granzin, R. Schlesinger, G. Buldt, T. Savopol, A. J. Scheidig,J. P. Klare and M. Engelhard (2002) Molecular basis of trans-membrane signalling by sensory rhodopsin II-transducer complex.Nature 419, 484–487.
13. Seidel, R., B. Scharf, M. Gautel, K. Kleine, D. Oesterhelt andM. Engelhard (1995) The primary structure of sensoryrhodopsin II: A member of an additional retinal protein sub-group is coexpressed with its transducer, the halobacterialtransducer of rhodopsin II. Proc. Natl Acad. Sci. USA 92, 3036–3040.
Figure 7. Isothermal titration calorimetry: (A) serine binding to HsHtrII(1-398), (B) serine binding to the complex HsSRII ⁄ HsHtrII(1-398). Thebinding parameters are listed in Table 3.
Photochemistry and Photobiology, 2009, 85 527

14. Zhang, W. S., A. Brooun, M. M. Mueller and M. Alam (1996)The primary structures of the archaeon Halobacterium salinariumblue light receptor sensory rhodopsin II and its transducer, amethyl-accepting protein. Proc. Natl Acad. Sci. USA 93, 8230–8235.
15. Hou, S. B., A. Brooun, H. S. Yu, T. Freitas and M. Alam (1998)Sensory rhodopsin II transducer HtrII is also responsiblefor serine chemotaxis in the archaeon Halobacterium salinarum.J. Bacteriol. 180, 1600–1602.
16. Mironova, O. S., R. G. Efremov, B. Person, J. Heberle, I. L.Budyak, G. Buldt and R. Schlesinger (2005) Functional charac-terization of sensory rhodopsin II from Halobacterium salinarumexpressed in Escherichia coli. FEBS Lett. 579, 3147–3151.
17. Kates, M., S. C. Kushwaha and G. D. Sprott (1982) Lipids ofpurple membrane from extreme halophiles and of methanogenicbacteria. Meth. Enzymol. 88, 98–111.
18. Wegener, A. A., J. P. Klare, M. Engelhard and H. J. Steinhoff(2001) Structural insights into the early steps of receptor-trans-ducer signal transfer in archaeal phototaxis. EMBO J. 20, 5312–5319.
19. Yu, J. H., Z. Hamari, K. H. Han, J. A. Seo, Y. Reyes-Dominguezand C. Scazzocchio (2004) Double-joint PCR: A PCR-basedmolecular tool for gene manipulations in filamentous fungi. Fun-gal Genet. Biol. 41, 973–981.
20. Scharf, B., B. Hess and M. Engelhard (1992) Chromophore ofsensory rhodopsin II from Halobacterium halobium. Biochemistry31, 12486–12492.
21. Chizhov, I., D. S. Chernavskii, M. Engelhard, K. H. Muller,B. V. Zubov and B. Hess (1996) Spectrally silent transitions in thebacteriorhodopsin photocycle. Biophys. J. 71, 2329–2345.
22. Chizhov, I., G. Schmies, R. Seidel, J. R. Sydor, B. Luttenberg andM. Engelhard (1998) The photophobic receptor from Natrono-bacterium pharaonis—Temperature and pH dependencies of thephotocycle of sensory rhodopsin II. Biophys. J. 75, 999–1009.
23. Hippler-Mreyen, S., J. P. Klare, A. A. Wegener, R. Seidel,C. Herrmann, G. Schmies, G. Nagel, E. Bamberg and M. Engel-hard (2003) Probing the sensory rhodopsin II binding domain of
its cognate transducer by calorimetry and electrophysiology.J. Mol. Biol. 330, 1203–1213.
24. Takahashi, T., B. Yan, P. Mazur, F. Derguini, K. Nakanishi andJ. L. Spudich (1990) Color regulation in the archaebacterialphototaxis receptor phoborhodopsin (sensory rhodopsin-Ii). Bio-chemistry 29, 8467–8474.
25. Tomioka, H., T. Takahashi, N. Kamo and Y. Kobatake (1986)Flash spectrometric identification of a fourth rhodopsin-like pig-ment inHalobacterium halobium. Biochem. Biophys. Res. Commun.139, 389–395.
26. Sasaki, J. and J. L. Spudich (1998) The transducer protein HtrIImodulates the lifetimes of sensory rhodopsin II photointermedi-ates. Biophys. J. 75, 2435–2440.
27. Sudo, Y., M. Iwamoto, K. Shimono and N. Kamo (2001)Pharaonis phoborhodopsin binds to its cognate truncated trans-ducer even in the presence of a detergent with a 1:1 stoichiometry.Photochem. Photobiol. 74, 489–494.
28. Sudo, Y., M. Iwamoto, K. Shimono and N. Kamo (2002) Tyr-199and charged residues of pharaonis phoborhodopsin are importantfor the interaction with its transducer. Biophys. J. 83, 427–432.
29. Sudo, Y. and J. L. Spudich (2006) Three strategically placedhydrogen-bonding residues convert a proton pump into a sensoryreceptor. Proc. Natl Acad. Sci. USA 103, 16129–16134.
30. Sudo, Y., M. Yamabi, S. Kato, C. Hasegawa, M. Iwamoto,K. Shimono and N. Kamo (2006) Importance of specific hydrogenbonds of archaeal rhodopsins for the binding to the transducerprotein. J. Mol. Biol. 357, 1274–1282.
31. Pierce, M. M., C. S. Raman and B. T. Nall (1999) Isothermaltitration calorimetry of protein-protein interactions. Meth.—AComp. Meth. Enzym. 19, 213–221.
32. Zhang, X. N., J. Zhu and J. L. Spudich (1999) The specificity ofinteraction of archaeal transducers with their cognate sensoryrhodopsins is determined by their transmembrane helices. Proc.Natl Acad. Sci. USA 96, 857–862.
33. Lin, L. N., J. Li, J. F. Brandts and R. M. Weis (1994) The serinereceptor of bacterial chemotaxis exhibits half-site saturation forserine binding. Biochemistry 33, 6564–6570.
528 Young-Jun Kim et al.






![Rhodopsin Dimers: Molecular Dynamics Simulations Using ... · membranes, support a molecular model of rhodopsin monomers orga-nized into two dimensional arrays of dimers [3]. Specifically,](https://img.pdfslide.us/doc/110x75/61455a0534130627ed50ebd3/rhodopsin-dimers-molecular-dynamics-simulations-using-membranes-support-a.jpg)






















