Embed Size (px)
Citation preview

11/30/2017
1
Perikles Simon
Johannes Gutenberg-University Mainz
Faculty of Social Science, Media and Sport
Department of Sports Medicine, Disease Prevention and Rehabilitation
www.sportmedizin.uni-mainz.de
Free circulating DNA as a quantitative and qualitative marker in exercise physiology
Exercise induces a highly dynamic and variable aseptic inflammation
Modified from: Walsh et al. Exercise Immunology Reviews 2010
Metabolism linkedto
Inflamation linkedto
Innate andadaptive Immunity
IL-6
TP1
TP2
Non-Linearity
Associationswith and without
Correlations

11/30/2017
2
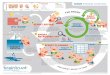
Input
Output
gene expression
protein profile
Black
Box
Body
Setting Athlete Key-Technologies
Transcriptomics
Proteomics
metabolite profile Metabolomics
NGS Appsgenome
Histone Modifications
DNA MethylationLiquid biopsy
cell-free DNA (cfDNA)
epi-
cfDNA
DNAse I
Real-time Physiome
164 bp
Information Technology
None- Wearable
Wearable
Information Technology
The Dawning of Big Data Viru, A. A., & Viru, M. (2001). Biochemical monitoring of sport
training. Human Kinetics.
Following Moore’s law
The dawning of Big Data: “The” technological innovation

11/30/2017
3
Beating Moore’s law
The dawning of even Bigger Data: Another technological innovation
By Aaron Krol
CfDNA a „versatile“ marker or no marker at all?
Injury
Exercise
Trauma
Down-Syndrome
Cancer
Sepsis
Chronicdiseases
Elevated cfDNA levels

11/30/2017
4
Exercise: an aseptic inflammation associated with cfDNA increases
Walsh et al. Exercise Immunology Reviews 2010
Metabolism linkedto
Inflamation and Innate andAdaptive Immunity
cfDNA
Beiter et al. Clinical Chemistry 2011
Lactate
cfDNA and Lactate during a 40 min aerobic endurance run at 9.6 km/h
10.90.8
0.7
0.6
2
3
10 20 30 40 +15 +30
time (min)
Lacta
te (
fold
-chnge
tore
st)
***
*
1
109876
5
4
3
2
cfD
NA
(fo
ld-c
hnge to r
est)
10 20 30 40 +15 +30
time (min)
***
*
******
**
1
Lacta
te (
mm
ol/l)
10
cfD
NA
(ng/m
l)
2.5
5
20
40
2
4
8
0.610 20 30 40 +15 +30
time (min)010 20 30 40 +15 +30
time (min)0
Haller et al. IJSPP 2016

11/30/2017
5
Limit of Detection
0,03 pg DNA 1/200 Cell-DNA-content
or 1,2 ng/ml plasma
Breitbach et al. J.of Appl.Physiol. 2014
Breitbach et al. PLoS ONE 2014
Establishing ultra-sensitiv detection procedures for cfDNA
Detection of amplicons in L1PA2 repeats (14000 copies/cell)
Detection of amplicons in LTR5 repeats (ca. 1000) copies/cell)
Helmig et al. Europ. J. Appl. Physiol. 2015
Detection of amplicons in ALU repeats
Direct PCR with native plasma
Multilocus primers for highly conserved repeat regions
Breitbach et al. PLoS ONE 2014
Routine silica-based DNA isolation looses > 60% of the cfDNA

11/30/2017
6
Breitbach et al. J Appl Physiol 2014 Mehdorn et al. IJSPP 2016
A similar study applying cycling revealed roughly 50% reducedincreases for the same O2-consumption
Subjects:
10 male triathletes
Inclusion criteria:
Complete 10 km in less than 40 min..
Preliminary investigation:
Pulmonary function test
Body impedance analysis (BIA)
Measurement of height [m] and weight [kg]
Intervention:
60% of VO2max downhill (-3.5%)
60% of VO2max uphill (6.5%)
80% of VO2max downhill (-3.5%)
80% of VO2max uphill (6.5%)
Mean SD min max
age [years] 36,9 7,03 27 48
height [m] 1,83 0,05 1,75 1,94
weight [kg] 80,18 4,49 75,1 88,2
BMI [kg/m^2] 23,97 1,49 21,94 26,92
fatty mass [%] 13,97 3,55 8,5 18,8
VC [l] 6,58 0,77 5,45 8,19
FEV1 [l] 4,98 0,65 4,01 6,38
VO2-max [ml/min/kg] 56,54 3,40 51 62,8
IAT [km/h] 13,15 1,01 12,3 15,4
V at 4mmol/l lactate [km/h] 14,03 1,19 12,6 16,4
Tab. 1: Characteristics of triathletes (N= 10).
Uphill and downhill running at same relative VO2

11/30/2017
7
Lacta
te m
mol/l (
log)
cfD
NA
ng/m
l (log)
HR
beats
/min
RE
R (
log)
At 80% VO2max downhill running ins increasing cfDNA more than uphill running
Increases in cfDNA following five repetitive sprints over 40m
9 participants (age: 23.6 ± 1.7 years; height: 179.6 ± 7.9 cm; weight: 80.9 ± 9.9 kg) 15 minute warm-up Random assignment in group A and B
5 * 40m sprint exercise(1 min break)
5 * 40m sprint exercise(5 min break)
5 * 40m sprint exercise(5 min break)
5 * 40m sprint exercise(1 min break)
75 min break 75 min break
blood sampling capillary cfDNA at baseline, after warming up, after every sprint (finger tip) venous cfDNA at baseline, after series 1 and 2 lactate at baseline, after warming up, after every sprint (ear lobe)
laboratory analysis (qPCR) statistical analysis

11/30/2017
8
Results
Folie Nr. 15Datum: 30.11.2017
venous cfDNA increased on average 2.4-fold after the short pause sprints
long breaks led to a signticantly lower 1.9-fold increase after long pauses (p<0.05)
lactate increased 3.4 fold after long pause exercise and 4.9-fold after 1 minute pauses (n.s.; p≥0.258).
venous cfDNA correlated significantly with lactate (r=0.69; p<0.05).
lactate(mmol/l)
venous cfDNA(ng/ml)
*average
cfDNA long
cfDNA short
average
lactate long
lactate short
Acute and chronic cfDNA alterations following strength training
Acute:16 strength exercises; 4-15 REPs Chronic effects (values before training)
Tug et al. PLoS ONE 2017

11/30/2017
9
50
100
150
200
HR
(beats
/min
)
pre 0 1 2 3 4 5 6_post
Heart rate
6x400m, 2‘pause
6x400m, 5‘pause
6x1000m, 2‘pause
1
1,5
2
0 1 2 3 4 5 6_post
Gold Standard: Trainings Impuls (Trimp)
Trim
p
(lo
g s
cale
)6x400m, 2‘pause at 18km/h
6x400m, 5‘pause at 18km/h
6x1000m, 2‘pause at maximal lactate steady state (MLSS)
Mean velocity: 14,4 km/h; range 12,8 – 16,5 km/h
cfDNA as a biomarker for exercise load?
EE median: 636 kcal; range: 484 – 794 kcal
EE median 310 kcal; range: 227 – 379 kcal
EE median 294 kcal; range: 248 – 382 kcal
1
1,5
2
2,5
pre
cfDNA (90bp)
cfD
NA
ng/m
l
(log s
cale
)
6x400m, 2‘pause at 18km/h
6x400m, 5‘pause at 18km/h
6x1000m, 2‘pause at maximal lactate steady state (MLSS)
Mean velocity: 14,4 km/h; range 12,8 – 16,5 km/h
cfDNA as a marker for carbohydrate energy expenditure
EE median: 636 kcal; range: 484 – 794 kcal
EE median 310 kcal; range: 227 – 379 kcal
EE median 294 kcal; range: 248 – 382 kcal
0
0,5
1
p3
p20
Lactate
Lacta
te m
mol/l
(lo
g s
cale
)
0 1 2 3 4 5
6_post
pre
p3
p20
0 1 2 3 4 5
6_post 1,6
1,8
2,0
2,2
2,4
2,6
2,8
1 2
log cfDNA (ng/ml)
Lo
g C
HO
(kca
l)
r =0,87; p<0.0001
”Metabolic” DNA ?

11/30/2017
10
Improved rehydration strategy for all out cycling time trial lowers cfDNA
Dehydration 16h before cycling under extrem environmental consditions in a heat chamber (air temperature 32°C, relative humidity, RH, 46%) until participants lost ca. 4 % of body mass.
Rehydration:Citrate (CIT) vs. Placebo (PLC)
Trial conducted by Martin Mooses and Vahur Oopik
Increases in cfDNA correlated with total distance covered
during game (Spearman‘s r=0.87)
Lactate vs. cfDNA during a week at baseline and following a soccer game
Lactate cfDNA

11/30/2017
11
Rykova et al. Expert Opinion on Biological Therapy 2012
Rapid release of cfDNA during exercise: Type, Origin and Function?
Beiter et al. Sports Medicine 2014
Active vesicular secretionMicroparticles /-vesiclesExosomesApoptotic bodies
Passive releaseNecrosisDetachment from cell surfaces
NetosisCell free chromatinActive release by other immune cells
Virtosomes / Metabolic DNA?
Free mitochondrial DNA ->DAMP
OriginFunction
Par
ticu
lom
ics
?
Trophic, oncogenic,immune response
Immune responseApoptosis/Metastasis
„waste disposal“
Magnetic Bead Isolation
Size ExclusionChromatography
OptiPrep densitygradient centrifugation
Extracellular vesicle and particle isolation and characterization
Alexandra Brahmer

11/30/2017
12
Extracellular Vesicles are released in parallel with cfDNA during exercise
Frühbeis et al. Journal of Extracellular Vesicles 2015
Eur J Appl Physiol
1 3
after subsequent centrifugation at 1600× g, 10,000× g and
100,000× g was nearly similar, demonstrating that centrifu-
gation and filtration had no critical influence on the cfDNA
levels in supernatants. Thus, the major part of the released
nuclear DNA exists in a non-vesicle-bound form in blood
plasma. Moreover, considering the low amounts of nuclear
DNA in 10,000× g and 100,000× g pellets, the contribu-
tion of EV-associated DNA to the total nuclear cfDNA pool
is marginal in healthy individuals. These findings are not
restricted to eccentric running exercise, since concentric
bicycle exercise revealed similar results (Supplemental Fig-
ure S1).
In contrast to nuclear cfDNA, we could not determine
a significant increase in mitochondrial DNA in any frac-
tion (Fig. 2b; Table 3). The majority [mean (± SD) 48.39
(21.25) %] of mitochondrial DNA was observed in the
10,000× g pellet, which is expected to include mito-
chondrial fragments pelleting together with MVs. Trace
amounts of mitochondrial DNA were identified in the
100,000× g supernatants and pellets. We detected only
(mean ± SD) 0.12 (0.11) % of total mitochondrial DNA in
the untreated 100,000× g pellets and 0.04 (0.03) % in the
DNase-treated 100,000× g pellets. In summary, most of the
cfDNA liberated in the blood stream during physical exer-
cise is released independent of EVs into the soluble plasma
fraction.
The results also permit some conclusions concerning
pre-analytical considerations. It was recommended to per-
form a first centrifugation step at 1200–1600× g and a sec-
ond step at 16,000× g to remove cellular debris which could
influence cfDNA quantification (El Messaoudi et al. 2013).
Our results indicate that this second step is not required
Fig. 2 Kinetics of mean (a)
nuclear and (b) mitochondrial
cfDNA concentrations in five
healthy subjects before and after
an incremental treadmill exer-
cise. LOQ indicates the lower
limit for the quantification of
DNA concentrations. Error bars
indicate ± SD. SN supernatant.
Total plasma, 1600× g superna-
tant; 10,000× g SN, 10,000× g
supernatant; 100,000× g SN,
100,000× g supernatant. For fur-
ther details please see Table 3
Author's personal copy
Eur J Appl Physiol
1 3
after subsequent centrifugation at 1600× g, 10,000× g and
100,000× g was nearly similar, demonstrating that centrifu-
gation and filtration had no critical influence on the cfDNA
levels in supernatants. Thus, the major part of the released
nuclear DNA exists in a non-vesicle-bound form in blood
plasma. Moreover, considering the low amounts of nuclear
DNA in 10,000× g and 100,000× g pellets, the contribu-
tion of EV-associated DNA to the total nuclear cfDNA pool
is marginal in healthy individuals. These findings are not
restricted to eccentric running exercise, since concentric
bicycle exercise revealed similar results (Supplemental Fig-
ure S1).
In contrast to nuclear cfDNA, we could not determine
a significant increase in mitochondrial DNA in any frac-
tion (Fig. 2b; Table 3). The majority [mean (± SD) 48.39
(21.25) %] of mitochondrial DNA was observed in the
10,000× g pellet, which is expected to include mito-
chondrial fragments pelleting together with MVs. Trace
amounts of mitochondrial DNA were identified in the
100,000× g supernatants and pellets. We detected only
(mean ± SD) 0.12 (0.11) % of total mitochondrial DNA in
the untreated 100,000× g pellets and 0.04 (0.03) % in the
DNase-treated 100,000× g pellets. In summary, most of the
cfDNA liberated in the blood stream during physical exer-
cise is released independent of EVs into the soluble plasma
fraction.
The results also permit some conclusions concerning
pre-analytical considerations. It was recommended to per-
form a first centrifugation step at 1200–1600× g and a sec-
ond step at 16,000× g to remove cellular debris which could
influence cfDNA quantification (El Messaoudi et al. 2013).
Our results indicate that this second step is not required
Fig. 2 Kinetics of mean (a)
nuclear and (b) mitochondrial
cfDNA concentrations in five
healthy subjects before and after
an incremental treadmill exer-
cise. LOQ indicates the lower
limit for the quantification of
DNA concentrations. Error bars
indicate ± SD. SN supernatant.
Total plasma, 1600× g superna-
tant; 10,000× g SN, 10,000× g
supernatant; 100,000× g SN,
100,000× g supernatant. For fur-
ther details please see Table 3
Author's personal copy
cfDNA / vesicDNA / vesic_surfDNA Mitochondria / cirmtDNA / DAMP
cirmtDNA
vesicDNA
vesic_surfDNA
Independent release of cfDNA and extracellular vesicles during exercise
Helmig et al. Europ. J. Appl. Physiol. 2015

11/30/2017
13
Muscle-specific hypo-methylation pattern of the MyoD core enhancer
Methylation level of MyoD core enhancer CpG2
skeletal muscle: <10% nonmuscle tissue: >50%
Brunk et al., Dev Biol. 1996
skeletalmuscle
wholeblood cells
pre post
Ø 93% (+/- 6.9)(n=14)
Ø 90% (+/- 8.1)(n=14)
23% 91%
Beiter and Simon unpublished
pre post 90+Chimerism
Blood
Chimerismus
Bone marrow
HSCT1 41 42 47 87 -
HSCT2 54 77 54 - 100
HSCT3 32 33 32 - 73
HSCT4 43 60 51 100 -
HSCT5 53 43 36 - 100
HSCT6 90+ 96+ 88+ 100 100
HSCT7 89+ 93+ 90+ 100 100
Mittelwert 57,5 63,5 56,9
SD 23,1 25,4 23,5
59,3 (±23)
X, Y X, X
Liver
HämatopoeticStem cells
X, Y
Y-chromosomal DNA: DYZ1, 77bp Botezatu et al., Clin Chem 2000
Predominant haematopoetic origin of cfDNA during exercise
Sex-missmatched transplant model
Tug et al. Exercise Immunology Reviews; 2015

11/30/2017
14
60
70
80
90
100
He
art
ra
te
(be
ats
/min
)
+8
d
36
36,4
36,8
37,2
37,6
38
Te
mp
era
ture
(°C
)
4000
6000
8000
10000
12000
-2,5
-1,5
-0,5
0,5
1,5
0,5
1
1,5
2
2,5
2,6
2,8
3
3,2
3,4
***
**
***
***
*** *
L1PA2 qPCR
PicoGreen Kitn.s.
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
Le
uko
cyte
s
(n/µ
l)
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1cfD
NA
ng
/ml
(lo
g s
ca
le)
hsC
RP
ng
/ml
(lo
g s
ca
le)
cfD
NA
ng
/ml
(lo
g s
ca
le)
CfDNA kinetics following cardiovascular surgery
21 Patients were monitored peri-operatively at the University Hospital in Tübingen, Germany14 extracorporal circulation (ECC)
7 without extracorporal circulation (no ECC)
Samples provided by S. Stoppelkamp and HP Wendel
-2,5
-1,5
-0,5
0,5
1,5 ECC
No ECC
hsC
RP
ng
/ml
(lo
g s
ca
le)
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
0,5
1
1,5
2
2,5
3
cfD
NA
ng
/ml
(lo
g s
ca
le)
2,6
2,8
3
3,2
3,4
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
+8
d
+5
d
+3
d
+2
d
+1
d
Po
st
Sta
rt
-d
1
**
n.s.
cfD
NA
ng
/ml
(lo
g s
ca
le) PicoGreen Kit
L1PA2 qPCR
Establishing ultra-sensitiv detection procedures for cfDNA and ctDNA
Detection of cfDNA with a KRAS mutationAs a liquid biopsy for cancer
The ideaMonitoring tumor DNA for optimizingtreatment options and sports therapy
Neuberger et al. Gene Therapy 2017
Ehlert et al. Sci. Reports 2017
Quintruple Pre-amplification followed by pooling
Transgene detection
Oncogene detection

11/30/2017
15
vv
v
v
cfDNA concentrations over the course of surgery in colon carcinoma patients
Ehlert et al. Sci. Reports 2017
cfDNA (ng/ml)
KRAS+DNA (copies/ml)
DNAse I activityreduction (%)

11/30/2017
16
cfDNA concentrations over the course of exercise of colon carcinoma patients
cfDNA (ng/ml)
% KRAS+DNA (%)
1
1,5
0
25
50
75
100
12
5
po
st3
po
st5
post2
0
post3
0
Power (W)
cfDNA
0
0,5
0
25
50
75
10
0
12
5
post3
po
st5
po
st2
0
po
st3
0
Power (W)
Lactate
cfD
NA
ng/m
l
(log s
cale
)L
acta
te m
mo
l/l
(lo
g s
ca
le)
Development of lactate and cfDNA values in patients with stable coronary heart disease
Michal et al. Trials 2015
74 coronary heart disease patients: cfDNA90bp 20.1 (23.8) ng/ml; range 5.1–183.0 ng/ml 74 healthy controls: cfDNA90bp 9.7 (4.2) ng/ml; range 1.6–23.7 ng/ml
Breitbach et al. PLoS ONE 2014
cfDNA
Lactate

11/30/2017
17
1
1,5
0
25
50
75
10
0
12
5
po
st3
po
st5
po
st2
0
po
st3
0
Power (W)
cfDNA
0
0,5
0
25
50
75
100
12
5
po
st3
post5
po
st2
0
po
st3
0
Power (W)
Lactate
cfD
NA
ng/m
l
(log s
cale
)L
acta
te m
mo
l/l
(lo
g s
ca
le)
Development of lactate and cfDNA values in patients with stable coronary heart disease
Michal et al. Trials 2015
74 coronary heart disease patients: cfDNA90bp 20.1 (23.8) ng/ml; range 5.1–183.0 ng/ml 74 healthy controls: cfDNA90bp 9.7 (4.2) ng/ml; range 1.6–23.7 ng/ml
Breitbach et al. PLoS ONE 2014
1
1,2
1,4
1,6
1,8
pre
0
du
rin
g3
du
rin
g6
du
rin
g9
du
rin
g1
2
du
rin
g1
5
po
st3
po
st5
po
st2
0
po
st3
0
0
0,2
0,4
0,6
0,8
log
lacta
te
KQ
-Mitte
lwe
rte
pre
0
du
rin
g3
du
rin
g6
du
rin
g9
du
rin
g1
2
du
rin
g1
5
po
st3
po
st5
po
st2
0
po
st3
0
cfD
NA
ng
/ml
(log s
cale
)
No CHFCHF
Haematocrit (%)
Erythrocytes (T/l)
Thrombocytes (G/l)
Immature Neutrophiles (G/l)
Mature Neutrophiles (G/l)
Eosinophiles (G/l)
Basophiles (G/l)
Monocytes (G/l)
Lymphocytes (G/l)
0,33
0,31
0,48
0,10
0,45
0,11
0,08
0,14
0,65
0,0019*
0,0040*
<,0001*
0,3937
<,0001*
0,3370
0,4574
0,1844
<,0001*
Spearman r p-Value
0,25 0,0236*
0,21 0,0609
0,41 0,0001*
0,09 0,4404
0,40 0,0002*
0,19 0,0800
0,10 0,3726
0,16 0,1424
0,62 <,0001*
Spearman r p-Value
Correlation with Capillary cfDNA Venous cfDNA
Netosis ?
Metabolic
DNAAnker et al. (1975) Spontaneous release of DNA by human blood lymphocytes as shown in an in vitro system. Cancer Research 35(9):2375–2382
Association of cfDNA with cellular populations during exercise

11/30/2017
18
Rykova et al. Expert Opinion on Biological Therapy 2012
Conclusion: We are proud to say; we can measure something
Beiter et al. Sports Medicine 2014
Active vesicular secretionExosomesMicroparticles /-vesiclesApoptotic bodies
Passive releaseNecrosisDetachment from cell surfaces
NetosisCell free chromatin
Virtosomes / Metabolic DNAFree mitochondrial DNA ->DAMP
OriginFunction
Trophic, Oncogenic,Immune response
Immune response
Metastasis!???
? Myogenic ?
( )( )
cfDNA increases withCHO Energy expenditureEccentricity of exerciseProlonged duration
• Anaerobic exercise induces cell-free DNA up to 20-fold within minutes
• Cell-free DNA as well as microvesicles but not mitochondrial DNA respond to exercise in a dose-response relationship
• Cell-free DNA is a marker for carbohydrate expenditure enabling objective determination of exercise load in elite intermittent sports
• Exercise is a valuable model to optimize conditions, pre-analyticalprocessing procedures and timing for liquid biopsies
• Liquid biopsies addressing disease specific sequences and cell- andtissue-type specific sequences / methylation pattern could be a valuable tool in personalized medicine
The take home messages

11/30/2017
19
Input
Output
gene expression
protein profile
Black
Box
Body
Setting Athlete Key-Technologies
Transcriptomics
Proteomics
metabolite profile Metabolomics
NGSgenome
Histone Modifications
DNA Methylation
Establishing cfDNA as marker for personalizedsports medicine optimizing training andintervention prevention
epi-
Radiomics
Real-time Omics for everyone
Visit our Department and turn your body into a hunting gepard!

11/30/2017
20
Thank You for Your Attention!
Dr. Elmo Neuberger
Dr. Tobias Ehlert
Dr. Susanne HelmigDipl biol. Thomas Beiter
Acknowledgement
Dr. Sarah Breitbach
Dr. Suzan Tug
Dr. Ella Lachtermann
Barlo HillenDavid Ochmann
Alexandra BrahmerNils Haller
From our team
Alumini involved
Daniel Pfirrmann
Cooperation
Dr. Martin Mooses (Tartu, Estonia) Prof. Dr. Vahur Oopik (Tartu, Estonia) PD. Dr. Eva Maria Krämer-Albers (Mainz, Germany)Prof. Ines Gockel (Leipzig; Germany)Prof. Mauro Giacca (Triest; Italy)Dr. Nägele (Optoprecision; Bremen, Germany)Prof. Richard Snyder (Florida; USA)Prof. Philippe Moullier (Nantes; France)Prof. Hauke Lang (Mainz; Germany)Prof. Hans-Peter Wendel (Tübingen; Gemrany)
Funding
Optoprecision
World Anti-Doping Agency
Röttger-FoundationKalkhoff-Rose-FoundationRhine-Main-University NetworkLift-Off Fund
Boehringer-Ingelheim Fund
Federal Institute of Sports Science