Embed Size (px)
Citation preview

216
Biochirnica el Biophysica Acta, 392 (1975) 216--222 © Elsevier Scientific Publishing Company, Amsterdam - Printed in The Netherlands
BBA 27663
FORMATION OF PHAGOLYSOSOMES CONTAINING DEXTRAN AND TRITON WR 1339 IN MOUSE LIVER
PI-KWANG TSUNG, MASAMITSU FUTAI, SHOJI OHKUMA and DEN'ICHI MIZUNO
Faculty of Pharmaceutical Sciences, University of 7bkyo, Hongo, Bunkyo-ku, Tokyo (Japan)
(Received November 25th, 1974)
Summary
After injection of Triton WR 1339 and dextran into mice, phagolyso- somes containing both compounds were obtained from the liver regardless of the order of injection of these materials. This suggests that phagolysosomes containing either Trition WR 1339 or dextran can fuse with phagosomes con- taining the other material.
The recoveries of various lysosomal enzymes differed in phagolysosomes after injection o f Triton WR 1339 with or without dextran: recoveries of 13-glucuronidase, ~-N-acetylglucosaminidase and arylsulfatase were high, and that of acid phosphatase was low.
Introduct ion
Morphological studies have shown that in the endocytic process lysosomal particles fuse with phagosomes to form phagolysosomes [1--3]. Although it is still difficult to define phagolysosomes, it seems interesting to study the prop- erties and fate of phagolysosomes biochemically.
Phagolysosomes formed in rat liver after injection of Triton WR 1339 show decreased density so that they can be separated from other cellular orga- nelles [4--6]. Making use of this change, we examined the following questions: (A) Whether phagolysosomes cohtaining dextran can still incorporate Triton WR 1339 and change to particles of lower density. (B) Whether phagolyso- somes formed after incorporation of Triton WR 1339 can still incorporate dextran. Phagolysosomes containing Triton WR 1339 have been shown to fuse with phagosomes containing trypan blue [7], invertase [8] or ceruloplasmin [9], but not with phagosomes containing peroxidase [10] or albumin [11]. Thus, it seemed necessary to examine these questions using the non-digestible, high molecular weight compound, dextran, as the second material. The results obtained in this study suggest that phagolysosomes containing one foreign

217
material can still participate in the endocytic process and incorporate a second material.
It also seemed interesting to study the lysosomal enzymes in lower density phagolysosomes, as liver lysosomes have been suggested to be heterogeneous [12] *. The recoveries of different marker enzymes in the lower density phago- lysosome fraction differed, implying the heterogeneity of lysosomes; this type of phagolysosome appeared to be rich in ~-N-acetylglucosaminidase, ~-glucuro- nidase and arylsulfatase.
Materials and Methods
Animals. Male ddY strain mice, weighing 25--30 g, were used. Experimental design. Animals were divided into three groups (A, B and C)
and their injection schedules are summarized below. Group A: (a) Animals were injected intraperitoneally with 70 mg of Tri-
ton WR 1339 1 day after intravenous injection of 30 mg of dextran T70 with 6 pCi of dextran [14 C] carboxyl in 0.5 ml saline. They were killed 2 days after the second injection. (b) Animals were injected intravenously with dextran as in (a) and were killed 3 days later. (c) Animals were injected intraperitoneally with Triton WR 1339 and killed 2 days later. The doses and routes of injection of dextran and Triton WR 1339 in Groups B and C were the same as in Group A, unless otherwise specified.
Group B: (a) Animals were injected with dextran 1 day after injection of Triton WR 1339 and were killed 3 days after the first injection. (b) Animals were killed 2 days after the injection of dextran. (c) Animals were killed 3 days after injection of Triton WR 1339.
Group C: (a) Animals were killed 3 days after intravenous injection of a mixture of dextran and Triton WR 1339.
Tissue fractionation and enzyme assay. Liver homogenates were frac- t ionated essentially as described by Trouet [4]. Liver was perfused and homo- genized as described previously [12] except that 3 vol. of 0.25 M sucrose con- taining 1 mM EDTA were used for homogenization. Nuclei and unbroken cells were separated by centrifugation at 800 × g for 10 min. The supernatant was then centrifuged at 15 000 X g for 20 min. The pellet termed the "large granu- lar fraction" in this paper was resuspended in 45% (w/w) sucrose to a final volume of 1 ml/g of liver and 5 ml of this suspension were layered on the bot tom of a discontinuous gradient containing 8 ml of 34.5% sucrose, 8 ml of 14.3% sucrose and 4 ml of 8.6% of sucrose. The gradient itself was placed on a cushion of 4 ml of 60% sucrose solution. Centrifugation was carried out at 65 000 X g for 2 h in an SW-25 rotor.
Fractions were collected from the bot tom of the tube. The distribution of radioactivity [13,14], enzyme activity [12] and protein [15] were determined by published procedures. Usually the livers of several mice were pooled for each experiment.
* In th is p a p e r t h e t e r m p h a g o l y s o s o m e s is used fo r the l y s o s o m a l f r a c t i o n i n c o r p o r a t i n g T r i t o n WR 1339 a n d / o r d e x t r a r a

218
Chemicals. p-Nitrocatechol sulfate and p-nitrophenyl ~-N-acetylglu- cosaminide were from Sigma Chemical Co., dextran [14 C] carboxyl (molecular weight, 60 000--90 000; specific activity, 1 mCi/g), from New England Nuclear Corp., and dextran T70 from Pharmacia Fine Chemicals Co. Other chemicals used were as described previously [12] or were reagent grade materials from commercial sources.
Results
Recovery of ~ 4 C-labeled dextran in the large granular fraction First ~ 4 C-labeled dextran only was injected into mice and its recovery in
the large granular fraction from the liver was studied. About 5% of the i 4 C-la- beled dextran injected was recovered in the large granular fraction 24 h after the injection with almost complete loss of radioactivity from the supernatant fraction. About 4% of the injected material still remained in the large granular fraction 96 h after the injection. No non-specific binding of dextran to the large granular fraction could be detected. Namely, when dextran was incubated with the large granular fraction for 30 min at 0 or 37°C and then the mixture was subjected to Ficoll gradient centrifugation, the dextran stayed in the top fraction [12].
These preliminary results showed that, if Triton WR 1339 is injected 24 h after dextran, it is possible to test whether phagolysosomes containing dextran still incorporate Triton WR 1339 to form lighter phagolysosomes. Details of the design of the experiments are given in Materials and Methods.
Sucrose gradient centrifugation of the large granular fraction from mouse liver after injection o f ~ 4 C-labeled dextran and Triton WR 1339
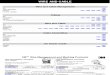
Phagolysosomes containing Triton WR 1339 were separated from other organelles using sucrose gradient centrifugation as described previously [4]. The large granular fraction obtained from mouse liver after injections of 14 C-labeled dextran and Triton WR 1339 together was fractionated by this procedure. As shown in Fig. 1, the activities of lysosomal marker enzymes and the radioactivity of dextran separated into two peaks: Peak 1 (Fractions Nos 1--5) and Peak 2 (Fractions Nos 6--10).
When Triton WR ].339 was not injected, no radioactivity or enzyme activ- ity was recovered in Peak 2, confirming that this peak contains phagolysosomes which have incorporated Triton WR 1339. The presence of radioactive Triton WR 1339 in the Peak 2 fraction has already been shown by previous workers [5]. Thus, the observation that lighter phagolysosomes contain dextran sug- gests that lysosomes can take up two foreign materials at the same time to form phagolysosomes.
The separation method used depends on the buoyancy of organelles during gradient centrifugation, and only the fraction of lower density (Peak 2) can be regarded as phagolysosomes. Phagolysosomes may be damaged when the large granular fraction is suspended in 45% sucrose solution. It was found that damaged phagolysosomes or lysosomes were recovered in the fraction of higher density (Peak 1). Namely, when the large granular fraction was subjected to the same gradient centrifugation after freezing and thawing or sonication,

2 1 9
u P
>
o
2
°4~t 14c- Dextr~.
20k
t Acid phosphatase
0 ~- GIucuronidase
1 5 10 15
Protein
Arylsulfatase
#- N-Acetyl glucosamimdase
5 10 15
Froction number Fig. 1. F r a c t i o n a t i o n o f the large granular fract ion f r o m m o u s e liver b y sucrose gradient e en tr i fuga t ion . Liver was r e m o v e d f r o m a m o u s e i n j e c t e d s i m u l t a n e o u s l y w i t h l 4 C-labeled d e x t r a n and T r i t o n WR 1 3 3 9 ( G r o u p C d e s c r i b e d u n d e r E x p e r i m e n t a l des ign) . T h e large granular f rac t ion s u s p e n d e d in 4 5 % sucrose was layered on 60% sucrose , and a s t e p w i s e gradient o f sucrose was m a d e as descr ibed in Materials and M e t h o d s . A f t e r c e n t r i f u g a t i o n at 6 5 0 0 0 × g fo r 2 h, fract ions w e r e c o l l e c t e d f r o m the b o t t o m o f the tube . T h e act iv i t ies of m a r k e r e n z y m e s , radioact iv i ty of d e x t r a n and pro te in w e r e as sayed and results are e x p r e s s e d as p e r c e n t a g e recover ies o f the tota l act iv i ty . T u b e s Nos 1 - -5 ( P e a k 1) a n d 6 - - 1 0 (Peak 2) w e r e p o o l e d . See t e x t for detai ls .
the activities of marker enzymes and radioactivity of dextran were recovered in Peak 1. After incorporation of only dextran phagolysosomes are known to have a higher density [16 ] . However, no change in the density of phagolysosomes after injection of dextran alone could be observed using the present procedure, because the range of concentrations of sucrose used was insufficient for their separation.
Recovery of 14 C-labeled dextran in phagolysosomes containing Triton WR 1339
After injection of dextran and then Triton WR 1339, about 20% of the radioactivity of dextran applied to the gradient centrifugation was recovered in the ~.ower density fraction (Peak 2, Table I). This suggests that phagolysosomes which have incorporated dextran could still incorporate Triton WR 1339 to form phagolysosomes of lower density. Moreover; when dextran was injected after Triton WR 1339 radioactivity was observed in Peak 2, suggesting that phagolysosomes formed after injection of Triton WR 1339 can still incorporate

220
T A B L E I
D I S T R I B U T I O N O F L Y S O S O M A L E N Z Y M E S A N D D E X T R A N IN P H A G O L Y S O S O M E S F R O M T H E L I V E R O F M I C E I N J E C T E D W I T H T R I T O N W R 1 3 3 9 A N D D E X T R A N
M i c e w e r e i n j e c t e d w i t h d e x t r a n a n d T r i t o n W R 1 3 3 9 f o l l o w i n g t h e d e s i g n o f e x p e r i m e n t s (A , B
a n d C) d e s c r i b e d in M a t e r i a l s a n d M e t h o d s . T h e l a rge g r a n u l a r f r a c t i o n f r o m t h e l ive r w a s f r a c t i o n a t e d b y
s u c r o s e g r a d i e n t c e n t r i f u g a t i o n , as s h o w n in Fig . 1. L i g h t e r f r a c t i o n s ( P e a k 2, F r a c t i o n s N o s 6 10 o f the
g r a d i e n t ) w e r e p o o l e d a n d t h e r a d i o a c t i v i t y o f d e x t r a n , a c t i v i t i e s o f l y s o s o m a l e n z y m e s a n d p r o t e i n w e r e
a s s a y e d . V a l u e s in t h e T a b l e a re a v e r a g e s o f t h o s e in f o u r e x p e r i m e n t s . T h e s t a n d a r d d e v i a t i o n s w e r e less
t h a n 3% o f t h e a v e r a g e v a l u e s . See t e x t f o r d e t a i l s .
A s s a y
P e r c e n t a g e r e c o v e r i e s o f e n z y m e a c t i v i t i e s a n d r a d i o a c t i v i t y
( A ) D e x t r a n T r i t o n * (B) T r i t o n T r i t o n (C) T r i t o n * * t h e n o n l y t h e n o n l y w i t h
T r i t o n ( c o n t r o l ) D e x t r a n ( c o n t r o l ) D e x t r a n
14 C - l a b e l e d
D e x t r a n 2 1 . 1 --- 2 8 . 1 3 1 . 8
A c i d p h o s p h a t a s e 1 1 . 9 1 2 . 2 1 3 . 3 1 9 . 7 1 3 . 9 / 3 - G l u c u r o n i d a s e 3 3 . 3 2 3 . 5 3 6 . 0 4 7 . 9 4 7 . 7 f l - N - A c e t y l - G l u c o s a m i n i d a s e 5 0 . 6 3 8 . 1 4 5 . 5 6 0 . 2 6 1 . 8 A r y l s u l f a t a s e 3 5 . 8 2 3 . 2 3 7 . 9 6 1 . 4 4 9 . 2
P r o t e i n 4 . 8 2 .9 8 .2 9 .1 7 .3
* C o n t r o l e x p e r i m e n t s w e r e d o n e o n t h e s a m e d a y u s i n g l i ve r s f r o m m i c e i n j e c t e d w i t h o n l y T r i t o n
W R 1 3 3 9 as d e s c r i b e d i n M a t e r i a l s a n d M e t h o d s ( G r o u p s A a n d B) . E s s e n t i a l l y n o r a d i o a c t i v i t y o f
d e x t r a n o r e n z y m e a c t i v i t y w a s r e c o v e r e d in t h i s f r a c t i o n a f t e r i n j e c t i o n o f d e x t r a n o n l y .
** T h e c o n t r o l f o r t h i s e x p e r i m e n t is t h e s a m e as f o r G r o u p B.
dextran (Table I). Probably not all the phagolysosomes in Peak 2 contain both dextran and Triton WR 1339. It is worth mentioning that Peak 2 was in the same position in the gradient in all these experiments.
Recovery of activities o f lysosomal marker enzymes in phagolysosomes (Peak 2)
After injection of Tri ton WR 1339 with or wi thout dextran, 10--20% of acid phosphatase activity in the large granular fraction was recovered into Peak 2 in all the experiments (Table I). The light fraction (Peak 2) obtained from the liver of mice after injection of dextran and then Triton WR 1339 contained 11% of the acid phosphatase, 33% of the fi-glucuronidase, 50% of the fl-N-acetylglucosaminidase and 35% of the arylsulfatase. Almost all the rest of these activities were recovered into Peak 1.
Similar results were obtained after injection of Triton WR 1339 with or without dextran. This suggests that the contents of different enzyme activities in phagolysosomes containing Triton WR 1339 are not the same as those in lysosomes in the large granular fraction.
Discussion
In the present work using Triton WR 1339 and dextran, it was shown that lysosomes could participate in the endocytic process twice and phagolysosomes containing both materials were isolated. Phagolysosomes containing Triton WR

221
1339 have been shown to fuse with phagosomes containing trypan blue [7] , ceruloplasmin [8] or invertase [9] , although they could not fuse with phago- somes containing peroxidase [8] or albumin [11]. De Duve [17] suggested that aged phagolysosomes could not participate in endocytosisl Thus it is pos- sible that aged phagolysosomes which participated in endocytosis only once could not fuse with phagosomes. This may be one explanation of the apparent discrepancy in the above literature [7--11] . Thus since phagolysosomes con- taining Triton WR 1339 show lower density and can be separated from other cellular organelles, using the proper conditions, it may be possible to prepare relatively pure phagolysosomes containing desired materials.
Results suggest that the lysosomes in mouse liver may be heterogeneous with respect to their contents of acid hydrolases. Evidence in support of this was that the lighter phagolysosomes (Peak 2, Fig. 1) separated after injection of Triton WR 1339 appeared to be relatively rich in ~-N-acetylglucosaminidase, ~-glucuronidase and arylsulfatase. The possible heterogeneity in the distribution of marker enzymes in lysosomes from rat liver has already been discussed [12] , and confirmed in mouse liver*. However, it must be emphasized that this heterogeneity was observed in lysosomes fractionated after disruption of cells. Therefore, the interpretation that the heterogeneity was due to selective leakage of enzymes from phagolysosomes (Peak 2) during the preparation could not be completely excluded. It seems unlikely that the membranes allow leakage of one enzyme but not another, but no final conclusion on this is possible until the properties of the membranes of phagolysosomes are known. Another explanation of the heterogeneity observed here is that some lysosomal enzymes are selectively induced after injection of Triton WR 1339 or dextran. In support of this possibility it has been shown that different lysosomal en- zymes increase at different rates in macrophages after endocytosis of serum proteins [ 18] .
Liver is known to be composed of different types of cells, such as paren- chymal and Kupffer cells. Thus the reason for the different recoveries of en- zyme activities in phagolysosomes (Peak 2) may be that either (A) one type of cells has heterogeneous lysosomes one of which responded to injected foreign material, or {B} the composit ion of lysosomal enzymes differs in different cell types and only a certain type(s) of cells responded to the injection. Experi- ments using fractionated liver cells may elucidate this point. Recently, Milson and Wynn [19] have shown that lysosomes from a homogeneous cell popula- tion of Chinese hamster ovary fibroblasts are heterogeneous. This supports explanation {A) above.
Acknowledgements
We thank Dr T. Takano (University of Hiroshima) for valuable discussions. This work was aided in part by a grant from the Ministry of Education of Japan.
* L ysosom es of m o u s e l iver w e r e s h o w n t o b e h e t e r o g e n e o u s u s i n g t h e same e x p e r i m e n t a l c o n d i t i o n s as for ra t l iver [ 1 1 ] (Tsung, P.K. and Mizuno, D., unpub l i shed observa t ion) .

222
References
1 G o r d o n , G.B. , Miller, L .R. a n d Bensch , K .G , ( 1 9 6 5 ) J . Ceil Biol. 25 , No. 2, p a r t 2, 41 55 2 Hi r sch , J .G . ( 1 9 6 2 ) J. Exp . Med. 116 , 8 2 7 - - 8 4 6 3 S t raus , W. ( 1 9 6 4 ) J . CeU Biol. 20 , 4 9 7 - - 5 0 7 4 T r o u e t , A. ( 1 9 6 4 ) Arch . Int . Phys io l . B i o c h e m . 72, 6 9 8 - - 7 0 0 5 L e i g h t o n , F. , Poo le , B., B e a u f a y , H,, B a u d h u i n , P., C o f f e y , J .W. , Fowle r , S. a n d de Duve, C. ( 1 9 6 8 ) J.
Ceil Biol. 37 , 4 8 2 - - 5 1 3 6 W a t t i a u x , R. , Wibo, M. a n d B a u d h u i n , P. ( 1 9 6 3 ) Ciba F o u n d a t i o n S y m p o s i u m o n L y s o s o m e s (de
R e u c k , A.V.S . a n d C a m e r o n , M.P., eds) , pp . 1 7 6 - - 2 0 0 , J . a n d A. Church i l l L td , L o n d o n 7 L l o y d , J .B. , Beck , F., Gr i f f i t h , A. a n d P a r r y , L.M. ( 1 9 6 8 ) A S y m p o s i u m o n the I n t e r a c t i o n of Drugs
a n d S u b c e l l u l a r C o m p o n e n t s in A n i m a l Cells ( C a m p b e l l , P .N. , ed . ) , pp . 1 7 1 - - 2 0 0 , J . a n d A. C h u r c h i l l L td , L o n d o n
8 Grego r i ad i s , G. a n d R y m a n , B.E. ( 1 9 7 2 ) B i o c h e m . J. 129 , 123 1 3 3 9 Grego r i ad i s , G., Morel l , A .G. , S tern l ieg , I. a n d S c h e i n b e r g , I .H. ( 1 9 7 0 ) J . Biol. C h e m . 245 ,
5 8 3 3 - - 5 8 3 7 10 J a c q u e s , P.J, ( 1 9 6 8 ) P h . D . Thesis , Univers i ty o f L o u v a i n (c i t ed in ref. 11) 11 Davies, M. ( 1 9 7 3 ) B i o c h e m . J. 136 , 5 7 - - 6 5 12 F u t a i , M., Tsung , P.K. a n d Mizuno , D. ( 1 9 7 2 ) B i o c h i m . B i o p h y s . A c t a 261 , 5 0 8 - - 5 1 6 13 Bray , G.A. ( 1 9 6 0 ) Anal . B i o c h e m . 1, 2 7 9 2 8 5 14 P a t t e r s o n , M.S. a n d Greene , R .C. ( 1 9 6 5 ) Ana l . C h e m . 37 , 8 5 4 - - 8 5 7 15 L o w r y , O .H, , R o s e b r o u g h , N.J . , Fa r r , A.L. a n d R a n d a l l , R .J . ( 1 9 5 1 ) J . Biol. C h e m . 193 , 2 6 5 - - 2 7 5 16 T h i n e s - S e m p o u x , D., J a c q u e s , P.J . a n d Bruns , G.P. ( 1 9 6 6 ) Proc . 3 rd FEBS Meet . , Warsaw, Po l and ,
A b s t r . F 71, P. 2 1 3 17 de Duve , C. ( 1 9 6 9 ) L y s o s o m e s in B io logy a n d P a t h o l o g y (Dingle , J .T. a n d Fell , H.B. , eds) , Vol . I, pp .
3 - - 4 0 , N o r t h - H o l l a n d Publ . Co. , A m s t e r d a m 18 C o h n , Z .A. , F e d o r k o , M.E. a n d Hi rsch , J .G . ( 1 9 6 6 ) J . Exp . Med. 123 , 7 5 7 - - 7 6 6 19 Milson, J .P . a n d W y n n , C.H. ( 1 9 7 3 ) B i o c h e m . J. 132 , 4 9 3 5 0 0