Embed Size (px)
Citation preview

For Review OnlyEffect of canola (Brassica napus) cultivar rotation on
Plasmodiophora brassicae pathotype composition
Journal: Canadian Journal of Plant Science
Manuscript ID CJPS-2019-0126.R1
Manuscript Type: Article
Date Submitted by the Author: 14-Aug-2019
Complete List of Authors: Cao, TiesenManolii, Victor; University of Alberta, Agriculture, Food and Nutritional SciencesZhou, Qixing; Crop Diversification Centre North, Alberta Agriculture and Rural DevelopmentHwang, Sheau-Fang; University of Alberta, Dept. of Agricultural, Food and Nutritional Science; Strelkov, Stephen; University of Alberta, Agricultural, Food and Nutrional Science
Keywords: Canola, clubroot, oilseed rape, pathotypes, Plasmodiophora brassicae
Is the invited manuscript for consideration in a Special
Issue?:Not applicable (regular submission)
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
1
Effect of canola (Brassica napus) cultivar rotation on Plasmodiophora
brassicae pathotype composition
Tiesen Cao, Victor P. Manolii, Qixing Zhou, Sheau-Fang Hwang and Stephen E. Strelkov
T. Cao, V.P. Manolii, S.F. Hwang, and S.E. Strelkov. Department of Agricultural, Food and
Nutritional Science, 410 Agriculture/Forestry Centre, University of Alberta, Edmonton AB T6G
2P5, Canada.
Q. Zhou. Crop Diversification Centre North, Alberta Agriculture and Forestry, Edmonton AB
T5Y 6H3, Canada.
Corresponding author: Stephen Strelkov (email: [email protected]).
Page 1 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
2
Abstract: In Canada, clubroot (Plasmodiophora brassicae) disease is managed mainly by
planting clubroot resistant (CR) canola (Brassica napus). New pathotypes of P. brassicae have
emerged recently, however, which are virulent on most CR canola cultivars. To understand the
impact of cultivar rotation on pathotype abundance, greenhouse experiments were conducted in
which different canola cultivar rotations were grown in a soil mix containing equal amounts of
pathotypes 5X and 3, which are virulent and avirulent, respectively, on CR canola. The rotation
treatments included: T1, the same susceptible cultivar planted over four cycles; T2, the same CR
cultivar planted over four cycles; and T3, different CR cultivars planted in each cycle. Clubroot
severity increased from cycles one to four in all treatments, with the exception of one CR
cultivar in T3 that may carry a different source of resistance. Pathogen populations were
recovered with a susceptible bait crop and pathotyped on the differentials of Williams plus a CR
host (B. napus ‘Mendel’). The percentage of galls classified as pathotype 5X in T1 declined from
50% to 6.7% over the course of the experiment, while galls classified as pathotype 5X increased
from 50% to 66.7% in both T2 and T3. Quantitative PCR analysis of the soil with pathotype 5X-
specific primers generally confirmed an increase in 5X DNA. The results suggest that continuous
planting of CR canola favours a rapid proliferation of virulent pathotypes of P. brassicae, as
indicated by the increases in pathotype 5X observed in this study.
Key words: canola, clubroot, oilseed rape, pathotypes, Plasmodiophora brassicae.
Page 2 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
3
Introduction
Plasmodiophora brassicae Woronin is a soil-borne pathogen that causes clubroot on
cruciferous oilseed and vegetable crops in Canada and other countries. Infection by P. brassicae
often results in the formation of clubs or galls on the roots of susceptible hosts, which interrupt
nutrient and water uptake by the plant and cause stunted growth and yellowing when the disease
is severe. Clubroot infection may significantly decrease yield, grain mass and oil content (Pageau
et al. 2006). On a global scale, yield losses associated with clubroot infestations are estimated to
be as high as 10-15% (Dixon 2006, 2009a).
Clubroot has long been recognized in Canada, where it has been a problem mainly on
cruciferous vegetables in Ontario, Quebec, British Columbia and the Atlantic Provinces (Howard
et al. 2010). In Alberta and Manitoba, clubroot has been reported sporadically in home and
market gardens since the 1920s (Howard et al. 2010). However, the disease was not described on
canola (oilseed rape; Brassica napus L.) in Alberta until 2003, when 12 clubroot infested fields
were detected near Edmonton (Tewari et al. 2005). Annual disease surveys since that time have
revealed > 3,000 clubroot infested fields by 2018 (Strelkov et al. 2019). Given its soil-borne
nature, the rapid spread of P. brassicae across canola fields has been attributed to the movement
of infested soil on farm machinery (Cao et al. 2009), some movement on wind-borne dust
(Rennie et al. 2015) and possibly dissemination on the surface of seeds or tubers (Rennie et al.
2011). The widespread occurrence of clubroot in Alberta, along with increasing reports of its
incidence on canola in Saskatchewan, Manitoba, Ontario and North Dakota (Cao et al. 2009;
Dokken-Bouchard et al. 2012; Chittem et al. 2014; Al-Daoud et al. 2017), suggest that it poses a
significant threat to the production of this crop in North America.
Page 3 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
4
Clubroot management is challenging given the fact that P. brassicae is able to survive as
resting spores in infested soil for extended periods of time (Karling 1968; Wallenhammar 1996).
Several practices, including liming the soil to increase pH (Murakami et al. 2002), soil fungicide
applications (Donald et al. 2001, 2004; Hwang et al. 2014), growing bait crops to reduce soil
spore load (Murakami et al. 2000, 2001), the manipulation of seeding date to minimize infection
(Gossen et al. 2009, 2012; Hwang et al. 2012), boron application (Deora et al. 2011), crop
rotation (Peng et al. 2015), and the sanitization of field equipment/machinery to limit field-to-
field disease spread (Howard et al. 2010) have been suggested to minimize the adverse impact of
clubroot. Since P. brassicae inoculum can build up quickly in the soil, the deployment of
clubroot resistant (CR) cultivars has been the most efficient and widely accepted measure for
canola disease management in western Canada (Gossen et al. 2014; Peng et al. 2014; Rahman et
al. 2014). The first CR canola cultivar was released in 2009, and as of fall 2018 there were 28
varieties with clubroot resistance available on the Canadian market. Continuous cropping of
resistant cultivars can, however, exert selection pressure on P. brassicae populations, resulting in
shifts in pathogen virulence (Seaman et al. 1963; LeBoldus et al. 2012). In 2013, only four years
after the introduction of the clubroot resistance trait, a new pathotype of P. brassicae capable of
overcoming resistance in most canola cultivars was detected in central Alberta (Strelkov et al.
2016). This pathotype, classified as pathotype 5 on the differentials of Williams (1966) or as
pathotype X on the Canadian Clubroot Differential Set (Strelkov et al. 2016; Strelkov et al.
2018), is known widely as pathotype ‘5X’. It differs from older pathotypes, including pathotype
3, which was the most prevalent before the introduction of CR canola, by its ability to cause
severe clubroot on most ‘resistant’ canola (Strelkov et al. 2016). Since 2013, multiple other new
pathotypes capable of overcoming resistance have been identified (Strelkov et al. 2018).
Page 4 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
5
In the current study, the effects of three different cultivars on the abundance of two P.
brassicae pathotypes (5X and 3) were examined over multiple cycles of rotation. Upon the
completion of a fourth cycle of rotation, P. brassicae resting spores in the soil were recovered by
growing a bait crop and the resulting galls were pathotyped on the differentials of Williams
(1966) plus the CR oilseed rape cultivar ‘Mendel’. Soil samples also were collected at the end of
each rotation cycle and the amount of pathotype 5X DNA was quantified with a 5X-specific
primer set and probe. The major objective of this study was to understand how canola cultivar
rotation treatments affect P. brassicae pathotype composition.
Materials and Methods
Inoculation methods and treatments
A black chernozemic soil, collected from a field at the Edmonton Research Station,
University of Alberta, with no history of clubroot (i.e., no history of cultivation of canola or
cruciferous vegetables), was used in this study. Testing with the P. brassicae-specific primers
TC1F/TC1R (Cao et al. 2007) confirmed the absence of pathogen DNA. The soil was passed
through a 10-mm metal sieve and air-dried in the laboratory at room temperature for about 1
month. The dried soil then was passed through a 2-mm sieve, and 6 kg of the sieved soil was
placed in each of nine polyethylene tubs (52 cm × 37 cm × 21 cm).
Two P. brassicae field isolates representing pathotypes 3 and 5X (Strelkov et al. 2016, 2018)
served as inoculum. The field isolates were maintained as resting spores in frozen (-20°C) root
galls on the universally susceptible Chinese cabbage (Brassica rapa var. pekinensis (L.) Lour.)
‘Granaat’. A total of 350 g of fresh galls of each pathotype were homogenized in 2 L sterile
distilled water (sdH2O) in a large blender for 3 min at high speed. The resulting spore suspension
Page 5 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
6
was adjusted to 2.5 L with sdH2O, mixed evenly, quantified with a haemocytometer and poured
into each of the nine polyethylene tubs. A total of 5 L of spore suspension was added per tub,
representing 2.5 L of each of pathotypes 3 and 5X. The soil in each tub was mixed manually with
the spore suspension for 1 min with a small shovel to generate an even, muddy mixture. The tubs
containing the homogenized soil inoculated with the spore suspension were placed on a bench in
the greenhouse and allowed to air dry for 2 weeks. The average infestation level after drying was
equivalent to 2 × 107 resting spores g-1 soil of each pathotype. After the soil had dried, any
clumps were homogenized in a mortar with a ceramic pestle and again passed through a 2-mm
sieve. The sieved soil in each tub was mixed evenly with two volumes of Sungro Professional
Growing Mix (Sun Gro Horticulture Canada Ltd., Seba Beach, Canada). The soil mixes were
transferred to nine polyethylene tubs with holes on the bottom, which were ready for subsequent
seeding.
Plant materials included the CR canola cultivars ‘45H29’, ‘6056CR’, ‘1960’, ‘9558C’, the
clubroot susceptible cultivar ‘‘45H26’, and the aforementioned Chinese cabbage ‘Granaat’.
About 160 seeds of a specific canola cultivar were planted in four rows in each of the nine
polyethylene tubs, which were placed in large polyethylene trays (115 cm × 198 cm × 9.5 cm).
The seeded tubs were watered carefully until full saturation of the soil was achieved. Three
weeks after seeding, the seedlings in each tub were thinned to 60 - 70 plants. The cultivar
rotation treatments included: T1, continuous cropping of the same susceptible canola cultivar
(‘45H26’); T2, continuous cropping of the same resistant canola cultivar (‘45H29’); and T3,
alternating resistant canola cultivars in the rotation. Each of the three rotation treatments was
replicated three times and one polyethylene tub was regarded as one experimental unit. All the
canola cultivars included in each rotation are listed in Table 1.
Page 6 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
7
Disease evaluation
The plants were uprooted gently with a spatula 6-7 weeks after seeding, and the roots were
washed in water. Clubroot severity on the roots of each plant was rated on a 0 to 3 scale
(Kuginuki et al. 1999), where: 0 = healthy (no galling), 1 = small galls on less than one-third of
the roots, 2 = small to medium galls on one-third to two-thirds of the roots, and 3 = medium to
large galls on more than two-thirds of the roots. An index of disease (ID) was calculated based
on the disease severity rating data using the formula of Horiuchi and Hori (1980) as modified by
Strelkov et al. (2006):
%1003
)3210((%)
Nnnnn
ID
where n is the number of plants in a class; N is the total number of plants; and 0, 1, 2, and 3 are
the symptom severity classes.
Quantification of P. brassicae pathotype 5X
About 120 g of soil was collected prior to the start of the first rotation (Dec. 19 2016) and at
the end of each rotation cycle (Feb. 21 2017, Apr. 13 2017, Jun. 09 2017, and Aug. 02 2017).
The soil samples were air-dried at room temperature in brown paper bags for 7 - 10 days,
homogenized in a mortar with a pestle, and passed through a 2-mm sieve to remove perlite
granules from the potting mix. Genomic DNA was extracted from 250 mg of the sieved soil
using a PowerSoil® DNA Isolation Kit (Qiagen Inc., Toronto) as per the manufacturer’s
instructions.
The quantitative polymerase chain reaction (qPCR) primers P5XF3, P5XR3 and minor groove
binding (MGB) probe P5XP3 (Zhou et al. 2018) were used to measure the amount of pathotype
Page 7 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
8
5X DNA in each soil sample. The qPCR assay was conducted in a Step One Plus Real-Time
PCR System (AB Applied Biosystems) in a 15-µL total reaction volume consisting of 0.4 µM
each forward or reverse primer, 0.2 µM TaqMan MGB probe, 1× TaqMan Environmental Master
Mix 2.0 (Thermo Fisher Scientific, Waltham, MA), and 5 µL sample DNA. The amplification
cycle was modified slightly and consisted of an initial step at 95oC for 10 min, followed by 40
cycles of 95°C for 15 s, and 60°C for 1 min. A total of 45 independent DNA samples were
included in the qPCR assay. For each qPCR run, a set of known quantities of P. brassicae
pathotype 5X DNA (1 × 108 to 1 × 101 fg) was included in triplicate on the 96-well plate to
generate a standard curve for each plate. The correlation equation generated from the known
amount of DNA (i.e., log10(DNA)) and threshold values (Ct) were used to calculate the amount
of pathotype 5X DNA present in each unknown sample on the same 96-well plate. The DNA
calculated according to each standard curve was normalized on a per gram soil basis, which was
subsequently used for mean comparisons of the treatments.
Pathotyping of galls recovered from the soil mixture
Resting spores of P. brassicae were recovered from the soil mix following the final cycle of
canola by growing the universally susceptible Chinese cabbage ‘Granaat’ as a bait crop. The
roots of the bait plants were collected after 6-weeks as described above, and single galls were
homogenized in ca. 5 ml sdH2O with a pestle in a mortar and filtered through 4 layers of
cheesecloth. The spore concentration was measured with a haemocytometer and adjusted to ca.
1 × 107 resting spores mL-1 with sdH2O. The recovered isolates were pathotyped on the
differentials of Williams (1966), which include the cabbages (B. oleracea var. capitata L.)
‘Badger Shipper’ and ‘Jersey Queen’ and the rutabagas (B. napus var. napobrassica (L.) Rchb.)
‘Laurentian’ and ‘Wilhemsburger’. The CR oilseed rape ‘Mendel’ also was included in the
Page 8 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
9
differentials, to identify isolates that could overcome resistance. Twelve seedlings of each host
were inoculated with inoculum from each gall following Strelkov et al. (2006), and inoculations
were replicated four times. The inoculated seedlings were grown in 6 cm × 6 cm × 6 cm plastic
pots (Kord Products Inc., Brampton, ON) filled with Sungro Professional Growing Mix at a rate
of one seedling per pot, and were maintained in a greenhouse at 20oC ± 2oC with a 16 h
photoperiod at a light intensity of 300 µmol m-2s-1. The growing mix was kept saturated with
water for the first week after inoculation, and then was watered and fertilized as required.
Clubroot development was evaluated as described earlier. Differential hosts were considered
resistant if the mean ID of the four replicates was < 50% and the 95% confidence interval did not
overlap 50% (LeBoldus et al. 2012). A total of 90 single root galls were evaluated for pathotype
classification, representing 10 single root galls from each of the nine combinations of rotation
treatment and replication.
Data analysis
Data were analyzed for statistical significance using the Analysis of Variance Procedure of
SAS (Statistical Analysis System; SAS Institute, Cary, NC), and Tukey’s honest significance test
was used for mean comparisons. The SAS LOGISTIC Procedure was used for analyzing
categorical data such as disease ratings. When comparing IDs, mean IDs (± 1 SE) are presented
for simplicity with statistical significance levels generated from the contrast of the SAS
LOGISTIC Procedure using the ordinal rating data.
Results
Effect of canola cultivar rotation on clubroot severity
Page 9 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
10
At the end of the first cycle of each rotation, the highest ID (91.5%) was observed in T1 on
the canola ‘45H26’, followed by IDs of 78.0% and 69.1% on ‘45H29’ in T2 and T3, respectively
(Table 2). All three IDs were significantly different from each other. At the end of cycle 2, the
IDs were 93.4% on ‘45H26’ in T1, 92.2% on ‘45H29’ in T2, and 91.3% on ‘6056CR’ in T3. At
the end of cycle 3, the IDs were 97.6% on ‘45H26’ (T1), 97.9% on ‘45H29’ (T2) and 98.2% on
‘1960’ (T3). The IDs at the end of cycles 2 and 3 showed no significant differences between
treatments (Table 2). At the end of cycle 4, IDs of 99.7% and 98.4% were observed on ‘45H26’
and ‘45H29’ in T1 and T2, respectively, values that were significantly greater than the ID of
14.1% observed on the cultivar ‘9558C’ in T3 (Table 2). Following each of the canola rotations,
the Chinese cabbage ‘Granaat’ was grown as a bait crop in all of the treatments. The highest ID
developed on the ‘Granaat’ plants in T1 (99.8%), while a significantly lower ID (98.7%)
developed on ‘Granaat’ in T3; in T2, the bait plants developed an ID of 99.1%, which was not
significantly different from the other two treatments (Table 2).
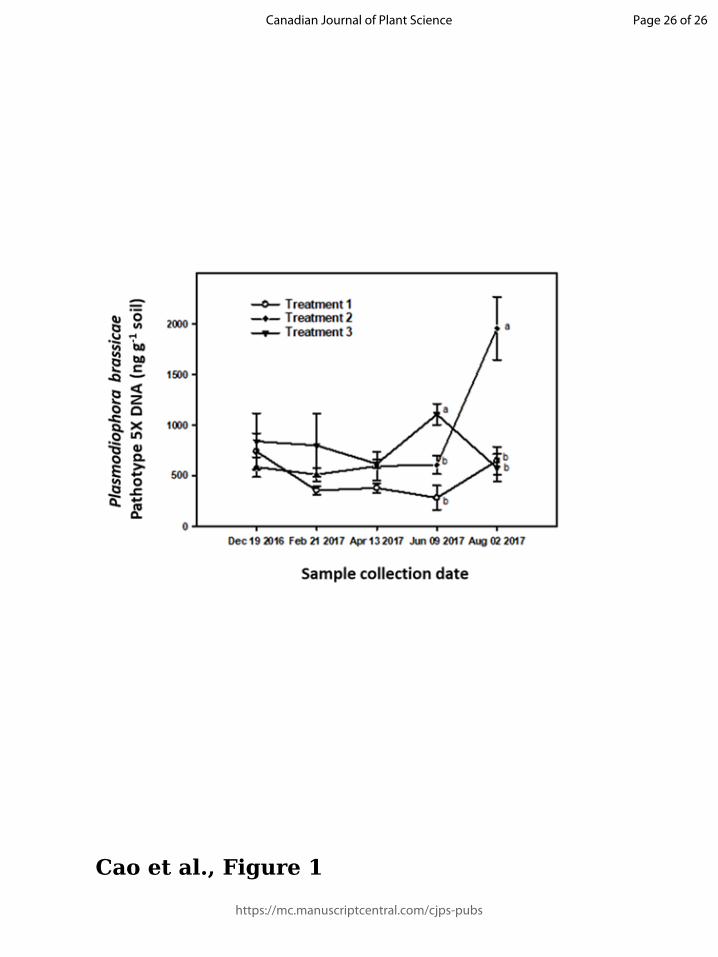
Effect of canola cultivar rotation on amount of pathotype 5X DNA
The amount of pathotype 5X DNA quantified by qPCR analysis was regarded as a proxy for
the size of the 5X population in the soil mix. In T1, this amount (on a per gram air-dried soil
basis, mean ± 1 SE) fluctuated from 7.4 × 102 ng ± 1.8 × 102 ng at the beginning of cycle 1 to 6.5
× 102 ng ± 1.4 × 102 ng by the end of cycle 4 (Fig. 1), with no statitistically significant
differences at any time. In T2, the amount of pathotype 5X DNA also was fairly constant from
the beginning of cycle 1 (5.9 × 102 ng ± 9.7 × 101 ng) to the end of cycle 3 (6.1 × 102 ng ± 8.9 ×
101 ng), but increased significantly to 2.0 × 103 ± 3.1 × 102 ng by the end of cycle 4 (Fig. 1). In
the case of T3, the amount of pathotype 5X DNA ranged from 5.8 × 102 ng ± 1.4 × 102 ng to 8.4
× 102 ng ± 2.8 × 102 ng over most of the cycles, but peaked at 1.1 × 103 ng ± 1.1 × 102 ng at the
Page 10 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
11
end of cycle 3 (Fig. 1). Across treatments, 5X DNA levels were highest for T3 at the end of cycle
3 and for T2 at the end of cycle 4 (i.e., at the end of the canola rotational sequence).
Effect of canola cultivar rotation on pathotypes recovered
Pathotype 5X was recovered at a high frequency (66.7%) from the root galls of bait plants
grown at the end of the rotations in T2 and T3 (Table 3). In contrast, the percentage of galls
classified as 5X was significantly lower (6.7%) in T1. Conversely, while pathotype 3 was most
commonly recovered (63.3%) in T1, it was rare in galls recovered from T2 and not found at all in
galls from T3. In addition to the two pathotypes originally (5X and 3) used to inoculate the soil
mix, several other pathotypes were recovered at lower frequencies in the three treatments. These
included pathotypes 5 (6.7%) and 8 (23.3%) in T1, pathotypes 5 (6.7%), 6 (10%) and 8 (10%) in
T2, and pathotypes 5 (16.7%), 6 (6.7%) and 8 (10%) in T3 (Table 3).
Discussion
Given the widespread and intensive cultivation of CR canola to manage clubroot (Peng et al.
2014), there has been significant selection pressure on P. brassicae populations to overcome host
resistance (Strelkov et al. 2016, 2018). Only four years passed between the release of the first CR
canola cultivar in Alberta and the detection of a new pathotype able to overcome this resistance
(Strelkov et al. 2016). This pathotype, 5X, is now just one of many novel pathotypes reported in
the province that can attack CR canola (Strelkov et al. 2018). The emergence of new pathotypes
represents one of the biggest challenges to clubroot management, since the planting of CR canola
is the most effective and widely used tool to control this disease (Peng et al. 2014; Strelkov and
Hwang 2014). Uncertainties with respect to pathotype shifts or pathogen population dynamics
Page 11 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
12
can complicate the sustainable production of canola in P. brassicae-infested fields. The current
report provides some insights into the nature of pathotype shifts.
An earlier study under greenhouse conditions found that field and single-spore isolates of P.
brassicae can adapt quickly to Brassica host genotypes, resulting in significant increases in
clubroot severity after only a few cycles of repeated exposure to the same hosts (LeBoldus et al.
2012). That study, however, did not compare the abundance of specific pathotypes in mixtures
exposed to different host rotations. Indeed, while it has been postulated that virulent components
of a P. brassicae population are selected preferentially in response to a resistance source
(Howard et al. 2010; Tanaka and Ito 2013), the present study shows how rapidly, after just a few
cycles, one pathotype can become predominant over another. In the absence of selection
pressure (i.e., T1, continuous susceptible canola), pathotype 3 was recovered most commonly
from the ‘Granaat’ bait plants at the end of the rotation. In contrast, when one (T2) or more (T3)
CR canola cultivars were rotated, pathotype 5X became predominant, while pathotype 3 was rare
(T2) or absent (T3).
In addition to the two ‘initial’ pathotypes 3 and 5X inoculated at the start of each rotation,
various other pathotypes (5, 6 and 8) were recovered at low frequencies at the end of the
experiment. Pathotype designations were based on the system of Williams (1966), with the
addition of the CR oilseed rape ‘Mendel’ to detect isolates able to overcome resistance. As such,
the full spectrum of pathotypes that can be distinguished with the recently developed CCD Set
(Strelkov et al. 2018) could not be identified. Nevertheless, aside from those classified as
pathotype 5X, none of the field isolates recovered at the end of the experiment were virulent on
‘Mendel’; this suggests that most if not all could not break clubroot resistance, which was
expected given that pathotypes other than 5X were recovered mainly from T1 (continuous
Page 12 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
13
susceptible canola). The identification of additional pathotypes is interesting and may reflect the
fact that field (vs. single-spore) isolates were used as the starting inoculum. Field isolates often
consist of pathotype mixtures, as has been shown in a number of earlier studies (see for example
Somé et al. 1996; Xue et al. 2008), and it is likely that some of the other pathotypes identified in
addition to 3 and 5X were present as minor components in the original inoculum. Moreover, the
possibility of additional diversity resulting from reproduction by the pathogen over the course of
the experiment cannot be ruled out, since karyogamy and meiosis occur in the P. brassicae life-
cycle (Tommerup and Ingram 1971; Buczacki 1983).
In general, the amount of pathotype 5X DNA in the treatments, as determined by qPCR
analysis, was consistent with the pathotyping results. The level of 5X DNA in T1 (continuous
susceptible canola) was similar throughout the trial, presumably because proliferation of
pathotype 5X was not favored in this treatment and no pathotype shift was observed. In contrast,
the amount of quantifiable 5X DNA increased significantly in T2 (repeated cultivation of
‘45H29’), which was consistent with the predominance of this pathotype among the field isolates
recovered at the end of the experiment. The only apparent discrepancy between the pathotyping
and qPCR results was observed in T3 at the end of the fourth cycle of the rotation, when the
amount of 5X DNA measured by qPCR was lower than expected. The CR canola ‘9558’, which
appeared to be fairly resistant to pathotype 5X, was grown in the fourth cycle of T3. It is
possible, therefore, that the inclusion of a resistant host resulted in a reduction in the quantity of
5X DNA in the soil mix when it was sampled immediately afterwards. This hypothesis is
supported by the observation that the amount of 5X DNA in T3 was highest (relative to T1 and
T2) at the end of the third cycle, when the canola ‘1960’ had been grown and developed severe
clubroot. Since the primers and probe used in the qPCR analysis were specific for pathotype 5X
Page 13 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
14
(‘pathotype 5-like strains’; Zhou et al. 2018), this assay could not be used to monitor the amount
of P. brassicae DNA corresponding to other pathotypes, including pathotype 3. Nonetheless, a
recent study indicated fairly rapid changes in soil inoculum loads under field conditions after the
cropping of a CR canola variety (Ernst et al. 2019).
Clubroot severity increased across all treatments over the course of the rotations; these
increases were most pronounced in T2 and T3, where the resistance-breaking pathotype 5X
initially represented half of the pathogen population but became more common after repeated
exposure to one (T2) or multiple CR canola cultivars (T3). Although the basis for resistance in
most cultivars is not in the public domain, a recent study suggested that the majority of CR
canola cultivars in Canada derived their resistance from the European Clubroot Differential
(ECD) 04 (Brassica rapa subsp. rapifera (L.) Metzg.) or the winter oilseed rape ‘Mendel’
(Fredua-Agyeman et al. 2018). It appears that resistance in ‘9558’, however, may be distinct,
given the low ID that developed on that genotype relative to all others. A generally lower
clubroot severity on ‘9558’ has been reported previously (Strelkov et al. 2018). Without definite
knowledge of the genetics of resistance in specific cultivars, however, it is difficult to draw
conclusions regarding the potential for rotation of resistance sources as a clubroot management
strategy.
It is worth noting that the ID which developed on ‘45H29’ in cycle 1 of T2 (78%), while in
the same general range as the ID on ‘45H29’ in cycle 1 of T3 (69%), was nonetheless
significantly higher at P < 0.05. In theory at least, the two values should have been nearly
identical, since it was the same host in the initial cycle for both of these treatments. It is
important to recall, however, that this experiment was conducted using field isolates of P.
brassicae. As discussed earlier, it is well established that field isolates may consist of pathotype
Page 14 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
15
mixtures (Somé et al. 1996; Xue et al. 2008), and indeed several additional pathotypes were
recovered in this experiment beyond the two (3 and 5X) that were inoculated originally.
Therefore, some limited fluctuation in ID would be expected and likely reflected the presence of
these minor components of the pathogen population.
Canola cultivar rotation had a significant impact on P. brassicae pathotype composition in the
current study. While farming practices and environmental conditions may mitigate this impact
under field conditions, it is clear that continuous cropping of CR canola can result in a rapid shift
towards a more virulent pathotype. This may help to explain the rapid emergence of new
pathotypes of P. brassicae that has been observed in Canada, and underscores the need for an
integrated approach to the management of clubroot of canola.
Acknowledgements
The authors acknowledge financial support from Agriculture and Agri-Food Canada and the
Canola Council of Canada via the Canadian Agricultural Partnership (CAP), as well as from
Alberta Canola, SaskCanola and the Manitoba Canola Growers Association. In-kind
contributions from Alberta Agriculture and Forestry and the University of Alberta are also
acknowledged. The authors also wish to thank H. Askarian, A. Botero, N. Fox, R. Fredua-
Agyeman, L. Galindo-González, C.P.A. Jayasinghege, E. Perez-Lara, X. Ma, M.M. Rahaman, R.
I. Strelkov, B. Wei, and Q. Zhou for technical assistance. Any mention of a commercial brand or
name does not constitute an endorsement by any of the funders.
References
Page 15 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
16
Al-Daoud, F., Moran, M., Gossen, B.D., and McDonald, M.R. 2017. First report of clubroot
(Plasmodiophora brassicae) on canola in Ontario. Can. J. Plant Pathol. 40: 96-99.
Buczacki, S.T. 1983. Plasmodiophora: an inter-relationship between biological and practical
problems. Pages 161-191 in S.T. Buczacki, ed. Zoosporic plant pathogens. Academic
Press, London, UK.
Cao, T., Manolii, V.P., Hwang, S.F., Howard, R.J., and Strelkov, S.E. 2009. Virulence and
spread of Plasmodiophora brassicae [clubroot] in Alberta, Canada. Can. J. Plant Pathol.
31: 321–329.
Cao, T., Tewari, J., and Strelkov, S.E. 2007. Molecular detection of Plasmodiophora brassicae
Woronin, causal agent of clubroot of crucifers, in plant and soil. Plant Dis. 91: 80–87.
Chittem, K., Mansouripour, S.M., and del Río Mendoza, L.E. 2014. First report of clubroot on
canola caused by Plasmodiophora brassicae in North Dakota. Plant Dis. 98: 1438.
Deora, A., Gossen, B.D., Walley, F., McDonald, M.R. 2011. Boron reduces development of
clubroot in canola. Can. J. Plant Pathol. 33: 475-484.
Dixon, G.R. 2006. The biology of Plasmodiophora brassicae Wor. – A review of recent
advances. Acta Hortic. 706: 271–282.
Dixon, G.R. 2009a. The occurrence and economic impact of Plasmodiophora brassicae and
clubroot disease. J. Plant Growth Regul. 28: 194–202.
Dixon, G.R. 2009b. Plasmodiophora brassicae in its environment. J. Plant Growth Regul. 28:
212-228.
Donald, E.C., Porter, I.J., and Lancaster, R.A. 2001. Band incorporation of fluazinam (shirlan)
into soil to control clubroot of vegetable Brassica crops. Aust. J. Exp. Agric. 41: 1223-
1226.
Page 16 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
17
Donald, E.C., Lawrence, J.M., and Porter, I.J. 2004. Influence of particle size and application
method on the efficacy of calcium cyanamide for control of clubroot of vegetable
brassicas. Crop Prot. 23: 297-303.
Dokken-Bouchard, F.L., Anderson, K., Bassendowski, K.A., Bouchard, A., Brown, B., Cranston,
R., Cowell, L.E., Cruise, D., Gugel, R.K., Hicks, L., Ippolito, J., Jurke, C., Kirkham, C.L.,
Kruger, G., Miller, S.G., Moats, E., Morrall, R.A.A., Peng, G., Phelps, S.M., Platford,
R.G., Schemenauer, I., Senko, S., Stonehouse, K., Strelkov, S., Urbaniak, S., and
Vakulabharanam, V. 2012. Survey of canola diseases in Saskatchewan, 2011. Can Plant
Dis. Surv. 92: 125–129.
Ernst, T.W., Kher, S., Stanton, D., Rennie, D.C., Hwang, S.F., and Strelkov, S.E. 2019.
Plasmodiophora brassicae resting spore dynamics in clubroot resistant canola (Brassica
napus) cropping systems. Plant Pathol. 68: 299-408.
Fredua-Agyeman, R., Hwang., S.F., Strelkov, S.E., Zhou, Q., and Feindel, D. 2018. Potential
loss of clubroot resistance genes from donor parent Brassica rapa subsp. rapifera (ECD
04) during doubled haploid production. Plant Pathol. 67: 892-901.
Gossen, B.D., Adhikari, K.K.C., and McDonald, M.R. 2012. Effect of seeding date on
development of clubroot in vegetable Brassica crops. Can. J. Plant Pathol. 34: 516–523.
Gossen, B.D., Deora, A., Peng, G., Hwang, S.F., and McDonald, M.R. 2014. Effect of
environmental parameters on clubroot development and the risk of pathogen spread. Can.
J. Plant Pathol. 36 (Suppl. 1): 37-48.
Gossen, B.D., McDonald, M.R., Hwang, S.F., and Kalpana, K.C. 2009. Manipulating seeding
date to minimize clubroot (Plasmodiophora brassicae) damage in canola and Brassicas.
Phytopathology. 99: S45 [Abstr.].
Page 17 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
18
Howard, R.J., Strelkov, S.E., and Harding, M.W. 2010. Clubroot of cruciferous crops –
perspectives on an old disease. Can. J. Plant Pathol. 32: 43–57.
Hwang, S.F., Ahmed, H.U., Zhou, Q., Strelkov, S.E., Gossen, B.D., Peng, G., and Turnbull, G.D.
2014. Efficacy of Vapam fumigant against clubroot (Plasmodiophora brassicae) of
canola. Plant Pathol. 63: 1374-1383.
Hwang, S.F., Cao, T., Xiao, Q., Ahmed, H.U., Manolii, V.P., Turnbull, G.D., Gossen, B.D., Peng,
G., and Strelkov, S.E. 2012. Effects of fungicide, seeding date and seedling age on
severity, seedling emergence and yield of canola. Can. J. Plant Sci. 92: 1175-1186.
Jones, D.R., Ingram, D.S., and Dixon, G.R. 1982a. Factors affecting tests for differential
pathogenicity in populations of Plasmodiophora brassicae. Plant Pathol. 31: 229–238.
Jones, D.R., Ingram, D.S., and Dixon, G.R. 1982b. Characterization of isolates derived from
single resting spores of Plasmodiophora brassicae and studies of their interaction. Plant
Pathol. 31: 239–246.
Karling, J.S., 1968. The Plasmodiophorales. Hafner Publishing Company, Inc., New York.
Kuginuki, Y., Hiroaki, Y., and Hirai, M. 1999. Variation in virulence of Plasmodiophora
brassicae in Japan tested with clubroot-resistant cultivars of Chinese cabbage (Brassica
rapa L. spp. pekinensis). Eur. J. Plant Pathol. 105: 327–332.
LeBoldus, J.M., Manolii, V.P., Turkington, T.K., and Strelkov, S.E. 2012. Adaptation to
Brassica host genotypes by a single-spore isolate and population of Plasmodiophora
brassicae (clubroot). Plant Dis. 96: 833-838.
Murakami, H., Tsushima, S., Akimoto, T., Murakami, K., Goto, I., and Shishido, Y. 2000.
Effects of growing leafy daikon (Raphanus sativus) on populations of Plasmodiophora
brassicae (clubroot). Plant Pathol. 49: 584-589.
Page 18 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
19
Murakami, H., Tsushima, S., Akimoto, T., and Shishido, Y. 2001. Reduction of spore density of
Plasmodiophora brassicae in soil by decoy plants. J. Gen. Plant Pathol. 67: 85–88.
Murakami, H., Tsushima, S., Kuroyanagi, Y., and Shishido, Y. 2002. Reduction of resting spore
density of Plasmodiophora brassicae and clubroot disease severity by liming. Soil Sci.
Plant Nutr. 48: 685-691.
Pageau, D., Lajeunesse, J., and Lafond, J. 2006. Impact de l’hernie des crucifères
[Plasmodiophora brassicae] sur la productivité et la qualité du canola. Can. J. Plant
Pathol. 28: 137-143.
Peng, G., Lahlali, R., Hwang, S.F., Pageau, D., Hynes, R.K., McDonald, M.R., Gossen, B.D.,
and Strelkov, S.E. 2014. Crop rotation, cultivar resistance, and fungicides/biofungicides
managing clubroot (Plasmodiophora brassicae) on canola. Can. J. Plant Pathol. 36
(Suppl. 1): 99–112.
Peng, G., Pageau, D., Strelkov, S.E., Gossen, B.D., Hwang, S.F., and Lahlali, R. 2015. A > 2-
year crop rotation reduces resting spores of Plasmodiophora brassicae in soil and the
impact of clubroot on canola. Eur. J. Agron. 70: 78-84.
Rahman, H., Peng, G., Yu, F., Falk, K.C., Kulkarni, M., and Selvaraj, G. 2014. Genetics and
breeding for clubroot resistance in Canadian spring canola (Brassica napus L.). Can. J.
Plant Pathol. 36 (Suppl. 1): 122–134.
Rennie, D.C., Holtz, M.D., Turkington, T.K., LeBoldus, J.M., Hwang, S.F., Howard, R.J., and
Strelkov, S.E. 2015. Movement of Plasmodiophora brassicae resting spores in
windblown dust. Can. J. Plant Pathol. 37: 188-196.
Page 19 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
20
Rennie, D.C., Manolii, V.P., Cao, T., Hwang, S.F., Howard, R.J., and Strelkov, S.E. 2011. Direct
evidence of surface infestation of seeds and tubers by Plasmodiophora brassicae and
quantification of spore loads. Plant Pathol. 60: 811-819.
Seaman, W.L., Walker, J.C., and Larson, R.H. 1963. A new race of Plasmodiophora brassicae
affecting Badger Shipper cabbage. Phytopathol. 53: 1426–1429.
Somé, A., Manzanares, M., Laurens, F., Baron, F., Thomas, G., and Rouxel, F. 1996. Variation
for virulence on Brassica napus L. amongst Plasmodiophora brassicae collections from
France and derived single-spore isolates. Plant Pathol. 45: 432-439.
Strelkov, S.E., Hwang, S.F., Manolii, V.P., Cao, T., and Feindel, D. 2016. Emergence of new
virulence phenotypes of Plasmodiophora brassicae on canola (Brassica napus) in
Alberta, Canada. Eur. J. Plant Pathol. 145: 517–529.
Strelkov, S.E., Hwang, S.F., Manolii, V.P., Cao, T., Fredua-Agyeman, R., Harding, M.W., Peng,
G., Gossen, G.D., Mcdonald, M.R., and Feindel, D. 2018. Virulence and pathotype
classification of Plasmodiophora brassicae populations collected from clubroot resistant
canola (Brassica napus) in Canada. Can. J. Plant Pathol. 40: 284-298.
Strelkov, S.E., Manolii, V.P., Lageyre, J., Hwang, S.F., Harding, M.W., and Daniels, G.C. 2019.
The occurrence and spread of clubroot on canola in Alberta in 2018. Can. Plant Dis. Surv.
99: 160-163.
Strelkov, S.E., Tewari, J.P., and Smith-Degenhardt, E. 2006. Characterization of
Plasmodiophora brassicae populations from Alberta, Canada. Can. J. Plant Pathol. 28:
467–474.
Tanaka, S., and Ito, S. 2013. Pathogenic and genetic diversity in Plasmodiophora brassicae
(clubroot) from Japan. J. Gen. Plant Pathol. 79: 297-306.
Page 20 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
21
Tewari, J.P., Strelkov, S.E., Orchard, D., Hartman, M., Lange, R.M., and Turkington, T.K. 2005.
Identification of clubroot disease of crucifers on canola (Brassica napus) in Alberta. Can.
J. Plant Pathol. 27: 143–144.
Tommerup, I.C., and Ingram, D.S. 1971. The life-cycle of Plasmodiophora brassicae Woron. in
Brassica tissue cultures and in intact roots. New Phytol. 70: 327-332.
Wallenhammar, A.C. 1996. Prevalence of Plasmodiophora brassicae in a spring oilseed rape
growing area in central Sweden and factors influencing soil infestation levels. Plant
Pathol. 45: 710–719.
Williams, P.H. 1966. A system for the determination of races of Plasmodiophora brassicae that
infect cabbage and rutabaga. Phytopathol. 56: 624–626.
Xue, S., Cao, T., Howard, R.J., Hwang, S.F., and Strelkov, S.E. 2008. Isolation and varation in
virulence of single-spore isolates of Plasmodiophora brassicae from Canada. Plant Dis.
92: 456-462.
Zhou, Q., Hwang, S.F., Strelkov, S.E., Fredua-Agyeman, R., and Manolii, V.P. 2018. A
molecular marker for the specific detection of new pathotype 5-like strains of
Plasmodiophora brassicae in canola. Plant Pathol. 67: 1582-1588.
Page 21 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science
For Review Only
22
Table 1. Cultivar rotation sequences grown in a soil mix inoculated with Plasmodiophora brassicae pathotypes 3 and 5X (1:1 ratio).Treatmenta Cycle 1 Cycle 2 Cycle 3 Cycle 4 Cycle 5Treatment 1 45H26 45H26 45H26 45H26 GranaatTreatment 2 45H29 45H29 45H29 45H29 GranaatTreatment 3 45H29 6056CR 1960 9558C Granaat
a Treatment 1, continuous cropping of Brassica napus ‘45H26’ (clubroot susceptible); Treatment 2, continuous cropping of B. napus ‘45H29’ (clubroot resistant); Treatment 3, alternating different clubroot resistant B. napus cultivars (‘45H29’ – ‘6056CR’ – ‘1960’ – ‘9558C’). The universally susceptible Chinese cabbage (B. rapa var. pekinensis) ‘Granaat’ was grown in cycle 5 of all treatments to recover P. brassicae from the soil mix.
Page 22 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
23
Table 2. Clubroot index of disease (%, mean ± 1 standard error) at the end of each rotation cycle.Treatmenta Cycle 1 Cycle 2 Cycle 3 Cycle 4 Cycle 5Treatment 1 91.5 ± 1.2 a 93.4 ± 2.5 a 97.6 ± 1.3 a 99.7 ± 0.3 a 99.8 ± 0.2 aTreatment 2 78.0 ± 3.3 b 92.2 ± 0.7 a 97.9 ± 0.8 a 98.4 ± 0.9 a 99.1 ± 0.9 abTreatment 3 69.1 ± 1.5 c 91.3 ± 2.3 a 98.2 ± 0.8 a 14.1 ± 1.2 b 98.7 ± 0.6 b
Note: Means followed by the same letter within each column are not significantly different at P < 0.05 according to the contrast with the SAS LOGISTIC Procedure.
a Treatment 1, continuous cropping of Brassica napus ‘45H26’ (clubroot susceptible); Treatment 2, continuous cropping of B. napus ‘45H29’ (clubroot resistant); Treatment 3, alternating different clubroot resistant B. napus cultivars (‘45H29’ – ‘6056CR’ – ‘1960’ – ‘9558C’). The universally susceptible Chinese cabbage (B. rapa var. pekinensis) ‘Granaat’ was grown cycle 5 of all treatments to recover P. brassicae from the soil mix.
Page 23 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
24
Table 3. Plasmodiophora brassicae pathotypes recovered from a soil mix after various crop rotations.
Pathotypes Recoveredc
TreatmentaNumber of single galls testedb 3 5 5X 6 8
Treatment 1 30 19 (63.3%) a 2 (6.7%) b 2 (6.7%) b 0 (0%) b 7 (23.3%) a
Treatment 2 30 2 (6.7%) b 2 (6.7%) b 20 (66.7%) a 3 (10%) a 3 (10%) a
Treatment 3 30 0 (0%) c 5 (16.7%) a 20 (66.7%) a 2 (6.7%) a 3 (10%) a
Note: Percentages followed by the same letter within a column are not significantly different at 5% significance level according to a t-test.
a Treatment 1, continuous cropping of Brassica napus ‘45H26’ (clubroot susceptible); Treatment 2, continuous cropping of B. napus ‘45H29’ (clubroot resistant); Treatment 3, alternating different clubroot resistant B. napus cultivars (‘45H29’ – ‘6056CR’ – ‘1960’ – ‘9558C’). The universally susceptible Chinese cabbage (B. rapa var. pekinensis) ‘Granaat’ was grown in cycle 5 of all treatments to recover P. brassicae from the soil mix.
b Resting spores extracted from a single gall were regarded as one field isolate. Thirty galls (field isolates) were tested from each treatment.
c Numbers indicate the total field isolates classified into each pathotype, with the percentages in parentheses indicating the percentage out of the total of 30; pathotype classifications were based on the reaction of the differentials of Williams (1966) plus the clubroot resistant B. napus ‘Mendel’; only those field isolates classified as pathotype 5 on Williams system (1966) and able to overcome resistance in ‘Mendel’ were denoted as pathotype 5X. None of the other pathotypes recovered were virulent on ‘Mendel’.
Page 24 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science

For Review Only
25
Figure Legend
Fig. 1. Plasmodiophora brassicae DNA concentration in a soil mix as measured by quantitative PCR
analysis with a pathotype 5X-specific primer and probe set. Means ± 1 standard error labelled with the
same letters are not significantly different at P < 0.05 according to Tukey’s test. Means of DNA
concentrations extracted from the soil samples collected on Dec 19 2016, Feb 21 2017, and Apr 13 2017
are not significantly different based on Tukey’s test, and thus are not labelled. Treatment 1, continuous
cropping of Brassica napus ‘45H26’ (clubroot susceptible); Treatment 2, continuous cropping of B. napus
‘45H29’ (clubroot resistant); Treatment 3, alternating different clubroot resistant B. napus cultivars
(‘45H29’ – ‘6056CR’ – ‘1960’ – ‘9558C’). The start of the rotations was on Dec 19, 2016 (first sampling
date), with additional sampling on Feb. 21 2017, Apr. 13 2017, Jun 09, 2017 and Aug. 02 2017,
corresponding to the end of cycles 1, 2, 3 and 4 of the rotations, respectively.
Page 25 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science
For Review Only
Cao�et�al.,�Figure�1
Page 26 of 26
https://mc.manuscriptcentral.com/cjps-pubs
Canadian Journal of Plant Science





























