Embed Size (px)
Citation preview

Biophysical Journal Volume 69 September 1995 1046-1067
Chain Folding and A:T Pairing in Human Telomeric DNA:A Model-Building and Molecular Dynamics Study
Debasisa Mohanty and Manju BansalMolecular Biophysics Unit, Indian Institute of Science, Bangalore 560 012, India
ABSTRACT The various types of chain folding and possible intraloop as well as interloop base pairing in human telomericDNA containing d(TTAG3) repeats have been investigated by model-building, molecular mechanics, and molecular dynamicstechniques. Model-building and molecular mechanics studies indicate that it is possible to build a variety of energeticallyfavorable folded-back structures with the two TTA loops on same side and the 5' end thymines in the two loops forming TATAtetrads involving a number of different intraloop as well as interloop A:T pairing schemes. In these folded-back structures,although both intraloop and interloop Watson-Crick pairing is feasible, no structure is possible with interloop Hoogsteenpairing. MD studies of representative structures indicate that the guanine-tetraplex stem is very rigid and, while the loopregions are relatively much more flexible, most of the hydrogen bonds remain intact throughout the 350-ps in vacuosimulation. The various possible TTA loop structures, although they are energetically similar, have characteristic inter protondistances, which could give rise to unique cross-peaks in two-dimensional nuclear Overhauser effect spectroscopy (NOESY)experiments. These folded-back structures with A:T pairings in the loop region help in rationalizing the data from chemicalprobing and other biochemical studies on human telomeric DNA.
INTRODUCTION
The ends of eukaryotic chromosomes, called telomeres, areknown to play an important role in the stability of chromo-somes and complete replication of chromosomal ends(Blackburn and Szostak, 1984; Henderson and Blackburn,1989; Blackburn, 1991, 1992). The telomeric DNA containsa double-stranded region with tandem repeats of short gua-nine-rich nucleotides at the 3' end, and the guanine-richstrand of the duplex terminates with a short single-strandedoverhang of this repeat sequence. The telomeric repeat ingeneral consists of runs of four contiguous guanines inter-spersed with stretches of two to four thymines (Blackburnand Szostak, 1984; Henderson and Blackburn, 1989; Black-burn, 1991, 1992) or of an adenine (A) and three contiguousguanines (Gs) interspersed with stretches of two to threethymines (Ts) (Moyzis et al., 1988). The exact sequencerepeat varies among species, for example, while sequencemotifs of the type d(T2G4) and d(T4G4) are found in thetelomeres of Tetrahymena and Oxytricha, respectively, inthe human telomeric DNA d(TTAG3) repeats are observed,whereas the telomeric repeat for Arabidopsis is d(T3AG3)(Blackburn and Szostak, 1984; Henderson and Blackburn,1989; Blackburn, 1991, 1992; Moyzis et al., 1988). Thepresence of guanine stretches gives these sequence motifsthe ability to adopt novel four-stranded structures (Sen andGilbert, 1988, 1990; Sundquist and Klug, 1989; Hendersonet al., 1987; Williamson et al., 1989) because of the forma-
Receivedfor publication 27 February 1995 and infinalforn 25 May 1995.Address reprint requests to Dr. Manju Bansal, Molecular Biophysics Unit,Indian Institute of Science, Bangalore 560 012, India. Tel.: 91-080-3092534; Fax: 91-080-3341683; E-mail: [email protected]. Dr. Mo-hanty's current address: Department of Physical Chemistry, The HebrewUniversity of Jerusalem, Jerusalem 91904, Israel.X 1995 by the Biophysical Society0006-3495/95/09/1046/22 $2.00
tion of Hoogsteen bonded guanine tetrads (Sasisekharan etal., 1975; Pinnavaia et al., 1975; Guschlbauer et al., 1990).Apart from telomeric DNA, the G-tetraplex structuresadopted by guanine-rich oligonucleotides have in generalreceived wide attention, because a number of proteins havebeen identified that bind specifically to these structures(Walsh and Gualberto, 1992; Pearson et al., 1993). Forma-tion of G-tetraplex structure has also been postulated to becrucial for dimerization of HIV RNA (Sundquist andHeaphy, 1993). Guanine-rich oligonucleotides are alsoknown to be inhibitors for fibrinogen action in thrombin(Wang et al., 1993; Macaya et al., 1993; Schultze et al.,1994, Padmanabhan et al., 1993) and HIV viral mediatedcell fusion (Wyatt et al., 1994).The conformational features of these G-tetraplex struc-
tures have been investigated by both theoretical (Panyutin etal., 1990; Balagurumoorthy et al., 1992; Mohanty and Ban-sal, 1993, 1994; Ross and Hardin, 1994) and experimental(Arnott et al., 1974; Zimmermann et al., 1975; Jin et al.,1990; Balagurumoorthy et al., 1992; Guo et al., 1992a;Scaria et al., 1992; Wang and Patel, 1992; Aboul-ela et al.,1992; Kang et al., 1992; Smith and Feigon, 1992, 1993)methods. A recent review (Williamson, 1994) gives a de-tailed account of the various studies of G-tetraplex struc-tures that have been reported. These studies indicate thatsequences with isolated stretches of guanines adopt tetramo-lecular structures by association of four guanine strands in aparallel orientation, and all guanines remain in anti confor-mation (Wang and Patel, 1992; Cheong and Moore, 1992;Aboul-ela et al., 1994; Gupta et al., 1993). This observationhas been further supported by the recently reported high-resolution crystal structure of d(TG4T) (Laughlan et al.,1994). On the other hand, sequences with stretches of gua-nines interspersed by thymines adopt folded-back structureswith a G-tetraplex stem and thymines forming loops con-
1 046

Chain Folding and A:T Pairing in Human Telomeric DNA
necting the adjacent or diagonal strands of the G tetraplex(Kang et al., 1992; Smith and Feigon, 1992, 1993). Depend-ing on the number of guanine stretches present in a strand,such folded-back structures could arise from dimerization oftwo hairpins (Kang et al., 1992; Smith and Feigon, 1992,1993) or intramolecular folding of a single strand (Wang etal., 1993; Macaya et al., 1993; Schultze et al.; 1994, Pad-manabhan et al., 1993; Wang and Patel, 1993), and suchstructures necessarily have adjacent or diagonal strands inan antiparallel orientation. The guanines in these antiparal-lel G-tetraplex structures generally adopt an alternating syn-anti conformation along the strand. The x-ray crystallo-graphic structure for d(G4T4G4) (Kang et al., 1992) has ahairpin dimer arrangement with T4 loops over adjacentstrands of a G tetraplex occurring on opposite ends of thetetraplex stem. However, the structure of the same sequencein solution when studied by nuclear magnetic resonance(NMR) has been found to have the T4 loops connecting thediagonal strands (Smith and Feigon, 1992, 1993). In addi-tion, a number of hairpin dimer structures with two thymineloops on the same side and interloop T:T pairing have alsobeen proposed from theoretical studies (Balagurumoorthy etal., 1992; Bansal and Mohanty, 1993; Mohanty and Bansal,1994) for the sequence motifs d(G4TnG4), with n = 2-4.Thus the various folded-back and hairpin dimer structuresfor telomeric DNA differ not only in the folding pattern butalso in the conformation and orientation of thymines inthe loop region. The possibility of occurrence of thyminepairs in the loop region of telomeric DNA had been firstproposed from model-building studies (Balagurumoorthyet al., 1992; Bansal and Mohanty, 1993; Mohanty andBansal, 1994). In the molecular dynamics studies (Rossand Hardin, 1994) of the hairpin dimer structure ofd(G4T2G4) two interloop T:T pairs were observed in thefinal structure, even though the simulation was carried outwith several starting structures without T:T pairs. On theother hand, no T:T pair was found in the NMR and crystalstructures for the telomeric DNA. However, subsequently aGreek-key type of structure with two T2 loops on the sameside and a single T:T pair between the two T2 loops wasobserved in the NMR (Macaya et al., 1993; Schultze et al.,1994) and x-ray crystallographic (Padmanabhan et al.,1993) studies of d(G2T2G2TGTG2T2G2), even though thefolding patterns in crystal and solution were topologicallydifferent. Similarly, a recent NMR study (Wang et al., 1994)of the oligonucleotide d(G4UTUTG4TUTUG4UUTTG4)has indicated that this sequence folds into a Greek-key motifwith T:T and U:U pairs being present in the loop region,whereas earlier NMR studies of a very similar sequenced(T4G4)4 (Smith and Feigon, 1992, 1993) had indicated thatit folds into an Indian-key motif with no T:T pairs. Thevariety of studies of telomeric DNA reported indicate thatthe structure is quite polymorphic, and the major source ofthe conformational variability is the conformation and ori-entation of the loop nucleotides, whereas the guanine stem
In view of the greater conformational variability of theloop regions in telomeric DNA, it is interesting to investi-gate the possible structures adopted by the human telomericsequence d(TTAG3)m or its variant d(T3AG3)m found inArabidopsis, because the presence of adenine in these se-quences gives rise to the possibility of the occurrence ofA:G or A:T pairs in the structure. However, relatively fewstudies (Balagurumoorthy and Brahmachari, 1994; Guo etal., 1992b) of telomeric DNA of humans or Arabidopsishave been reported in contrast to the large number ofexperimental studies of sequences of the type d(TnG4)m.The single NMR study (Wang and Patel, 1993) ofd(AG3(TTAG3)3) indicates that the molecule folds into anIndian-key motif with a G3 stem and TTA loops, but no basepairing is observed in the loops. On the other hand, chem-ical probing studies (Balagurumoorthy and Brahmachari,1994) of d(G3TTAG3) and d(TTAG3) repeats of differentlengths indicate that N7 of adenine is inaccessible, thussuggesting the possibility of the participation of adenine inhydrogen bonding interactions. Similarly, the higher stabil-ity of d(G4TTAG4) compared with d(G4T3G4) has beenexplained on the basis of the occurrence of A:T pairs in theloop regions (Guo et al., 1992b). Therefore, in the presenttheoretical study, model-building, molecular mechanics,and molecular dynamics techniques have been used to elu-cidate various possible structures for d(T.AG3) repeats.The specific questions addressed in this study are the
following: 1. Will these sequences fold, with a Tn loop andAG3 stem consisting of two G tetrads and two AGAGtetrads or with a TnA loop and G3 stem? 2. Is it possible tohave A:T pairs in the loop regions of folded-back structureswith G3 stems and TnA loops? If so, what are the varioustypes of A:T pairs compatible with different types of chainfolding? 3. Are the hydrogen bonds in the loop region reallystable?
MODEL BUILDINGA sequence of the type d(TnAG3)4 or d(G3(TnAG3)3) can adopt a mono-molecular structure by Greek-key- or Indian-key-type folding (Frank-Kamenetskii, 1992), and these structures can have a G-tetraplex stem withthree guanine tetrads (Fig. 1 a and b) and loops containing adenine andthymine residues; altermatively, adenine could occur in the stem, resultingin structures with the tetraplex stem consisting of two G tetrads and twoAGAG tetrads (Fig. 1 c and d) and only thymines constituting the loops.The structures with AGAG tetrads in the stem and Tn loops would besimilar to the structures of d(TnG4)4 (Mohanty and Bansal, 1994), as theAGAG tetrad is isostructural with the guanine tetrad. Structures with G3stems and T,A loops are expected to be different from their thyminecounterparts from the point of view of stacking and hydrogen bonding inthe loop region, because the first thymine and the adenine could inprinciple form intraloop as well as interloop A:T pairing involving Watson-Crick and Hoogsteen types of hydrogen bonding. Moreover, sequences ofthe type d(G3TnAG3) are expected to adopt only hairpin dimer structureswith G3 stems and TnA loops. Inasmuch as all possible intramolecularfolded-back structures with G3 stems and TnA loops can in principle beobtained from a combination of hairpins, model building was carried outfor different possible hairpin dimer structures for d(G3TnAG3) with G3stems and TnA loops.
The sequences of the type d(G3TnAG3) can form three types of hairpindimer with the two T,A loops over adjacent strands and occurring on the
1 047Mohanty and Bansal
has relatively less conformational variability.

Volume 69 September 1995
a b
c d
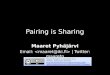
FIGURE 1 Schematic diagrams of the monomolecular structures arisingfrom intramolecular folding of the sequence d(TnAG3)4, n = 2, 3. (a)Greek-key-type folding with G3 in the stem and TnA in the loop. (b)Indian-key-type folding with G3 in the stem and TnA in the loop. (c)Greek-key-type folding with AG3 in the stem and T2 or T3 in the loop. (d)Indian-key-type folding with AG3 in the stem and T3 in the loop. This typeof structure is not possible for d(T2AG3)4, as a minimum of three nucle-otides are required for building a loop across the diagonal strands in a G
tetraplex. The guanines and adenine in the tetraplex stem have beenrepresented as rectangular planes, with the longer edge indicating the sideof guanine that contains Ni and N2 or the side of adenine that contains Niand C2 and the shorter edge indicating the side containing N7 of guanineor adenine. The alternate guanines along a strand, which have their a facesvisible from the top, have been shaded to show alternating syn-antiarrangement of guanines along the strand resulting from base flipover(Mohanty and Bansal, 1993).
same side of the tetraplex stem (Fig. 2 a) or on opposite ends (Fig. 2 b), or
with the loops over diagonal strands and at opposite ends of the G tetraplex(Fig. 2 c). Even though the guanines in the antiparallel G-tetraplex stem ofthese hairpin dimers can have all-syn-all-anti or alternating syn-anti con-
formation (Mohanty and Bansal, 1993, 1994; Balagurumoorthy et al.,1992), only the ones with an alternating syn-anti arrangement of guaninesalong the strand have been observed in x-ray and NMR experiments. Suchstructures have favorable intra-strand as well as interstrand interaction,unlike the all-syn-all-anti structures, in which the all-syn strand is ener-
getically unfavorable (Mohanty and Bansal, 1993). It is important to notethat, once we assume an alternating syn-anti arrangement of guanines inthe stem region, for the hairpin dimers with three guanines in the stem thestructures with loops at opposite ends (Fig. 2 b and c) have to be asym-metric, with the TnA loop in one hairpin joining a guanine in syn confor-mation to a guanine in anti conformation while the other joins a guanine inanti conformation to a guanine in syn conformation (Fig. 2 b and c).However, hairpin dimers with two loops on same side of the G tetraplexcan be symmetric (Fig. 2 a) as well as asymmetric. Thus the hairpin dimerswith G3 stems are different from the hairpin dimers with G4 stems, whereall three types of dimer can be symmetric because of the even number ofguanines in the stem (Mohanty and Bansal, 1994; Smith and Feigon, 1993).In the symmetric hairpin dimers with two loops on the same side thethymines and adenines can form a TATA tetrad (Chernyi et al., 1990),which is nearly isostructural to the G tetrad and can accommodate maxi-mum number of hydrogen bonds between the adenines and thymines thatform the tetrad. Such symmetric dimer structures with TATA tetrads can beobtained by dimerization of hairpins with the loop across the large groove(i.e., the groove in which donor atoms N2 and Ni of the guanine in the anticonformation are hydrogen bonded to acceptors N7 and 06 of the guaninein the syn conformation (Fig. 3 a)) or from hairpins with the loop across thesmall groove (i.e., the groove in which donor atoms N2 and Ni of theguanine in the syn conformation are hydrogen bonded to acceptors N7 and06 of the guanine in the anti conformation (Fig. 3 a)). Thus the objectiveof the model-building study was to build hairpins with appropriate orien-tation of adenine and thymines in the loop so that, in the symmetric dimerstructure obtained by applying a twofold rotation about the helical axis ofthe stem, both intraloop and interloop hydrogen bonds can be formed.
The various types of TATA tetrad (Chernyi et al., 1990) that areexpected from symmetry considerations are the following: Two Watson-Crick (Fig. 3 b) (or Hoogsteen (Fig. 3 c)) AT pairs are related by a twofoldaxis so that the A and the T of one Watson-Crick (or Hoogsteen) pair formone hydrogen bond each with the T and the A of the other Watson-Crick(or Hoogsteen) pair. There are several ways in which this type of basepairing can be accommodated in the hairpin dimer.
WC1 or HGI
An A:T Watson-Crick (Fig. 4 a) (or a Hoogsteen (Fig. 4 c))pair is in the large groove and either of the followingconditions exists: a) The loop is also across the large grooveof the tetraplex. Thus in the dimer one loop connects arms1 and 2 and the other loop connects arms 3 and 4 in Fig. 4a and c). These types of structure will be referred to asWCla or HGla. b) The loop is across the small groove ofthe tetraplex. Thus in the dimer one loop connects arms 1and 4 and the other loop connects arms 2 and 3 in Fig. 4 aand c. These types of structure will be referred as WClb orHGlb.
WC2 or HG2
An A:T Watson-Crick (Fig. 4 b) (or a Hoogsteen (Fig. 4 d))pair is in the small groove and either of the followingconditions exists: a) The loop is also across the smallgroove, i.e., one loop connects arms 1 and 4 and the sym-metry-related loop connects arms 2 and 3 in Fig. 4 b and d.These types of structure will be referred as WC2a or HG2a.b) The loop is across the large groove, i.e., one loop con-nects arms 1 and 2 and the symmetry-related loop connectsarms 3 and 4 in Fig. 4 b and d. These types of structure willbe referred as WC2b or HG2b.
1 048 Biophysical Journal
Chain Folding and A:T Pairing in Human Telomeric DNA
a b c
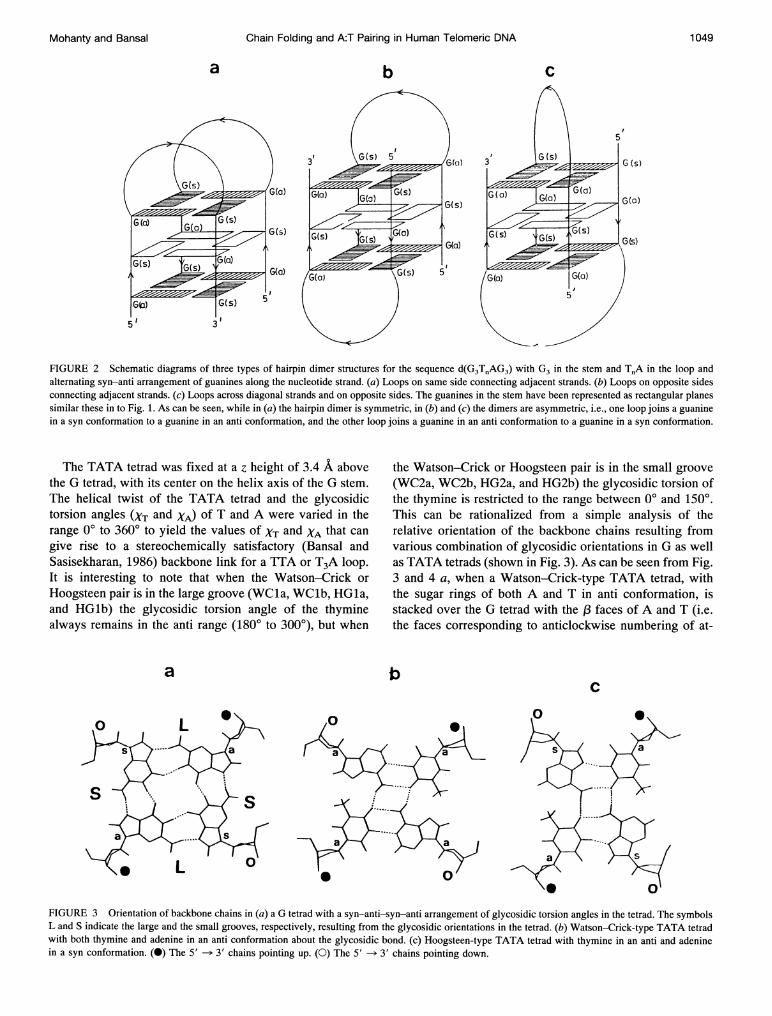
FIGURE 2 Schematic diagrams of three types of hairpin dimer structures for the sequence d(G3T.AG3) with G3 in the stem and T.A in the loop andalternating syn-anti arrangement of guanines along the nucleotide strand. (a) Loops on same side connecting adjacent strands. (b) Loops on opposite sidesconnecting adjacent strands. (c) Loops across diagonal strands and on opposite sides. The guanines in the stem have been represented as rectangular planessimilar these in to Fig. 1. As can be seen, while in (a) the hairpin dimer is symmetric, in (b) and (c) the dimers are asymmetric, i.e., one loop joins a guaninein a syn conformation to a guanine in an anti conformation, and the other loop joins a guanine in an anti conformation to a guanine in a syn conformation.
The TATA tetrad was fixed at a z height of 3.4 A abovethe G tetrad, with its center on the helix axis of the G stem.The helical twist of the TATA tetrad and the glycosidictorsion angles (XT and XA) of T and A were varied in therange 00 to 360° to yield the values of XT and XA that cangive rise to a stereochemically satisfactory (Bansal andSasisekharan, 1986) backbone link for a TTA or T3A loop.It is interesting to note that when the Watson-Crick orHoogsteen pair is in the large groove (WCla, WClb, HGla,and HGlb) the glycosidic torsion angle of the thyminealways remains in the anti range (1800 to 3000), but when
a
the Watson-Crick or Hoogsteen pair is in the small groove(WC2a, WC2b, HG2a, and HG2b) the glycosidic torsion ofthe thymine is restricted to the range between 0° and 1500.This can be rationalized from a simple analysis of therelative orientation of the backbone chains resulting fromvarious combination of glycosidic orientations in G as wellas TATA tetrads (shown in Fig. 3). As can be seen from Fig.3 and 4 a, when a Watson-Crick-type TATA tetrad, withthe sugar rings of both A and T in anti conformation, isstacked over the G tetrad with the ,3 faces of A and T (i.e.the faces corresponding to anticlockwise numbering of at-
bc
A a - a' S a X1:
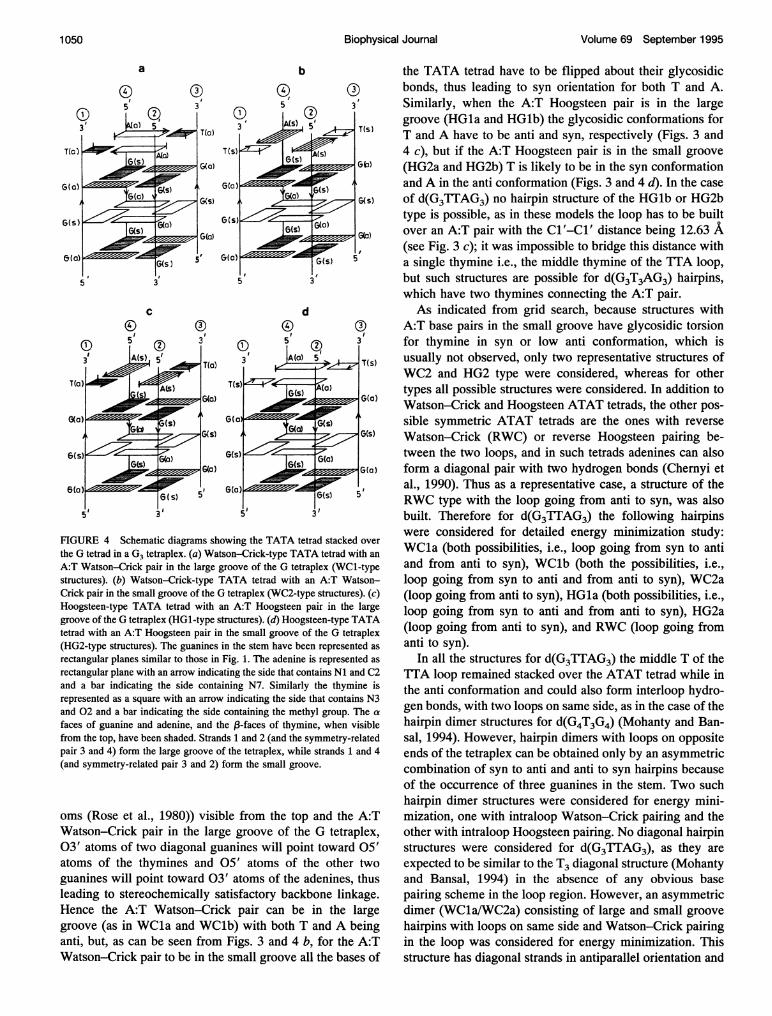
FIGURE 3 Orientation of backbone chains in (a) a G tetrad with a syn-anti-syn-anti arrangement of glycosidic torsion angles in the tetrad. The symbolsL and S indicate the large and the small grooves, respectively, resulting from the glycosidic orientations in the tetrad. (b) Watson-Crick-type TATA tetradwith both thymine and adenine in an anti conformation about the glycosidic bond. (c) Hoogsteen-type TATA tetrad with thymine in an anti and adeninein a syn conformation. (O) The 5' --> 3' chains pointing up. (O) The 5' -->3' chains pointing down.
Mohanty and Bansal 1 049
Volume 69 September 1995
a
(,
T(c)
G(a)
G(s)
6(c)
0
D 5 Q,¢(a) 5
G(a) a
lG(s) G(a)
Gs )
5 3
b
3
c
T(I)
G(a)
G(s)
d
FIGURE 4 Schematic diagrams showing the TATA tetrad stacked over
the G tetrad in a G3 tetraplex. (a) Watson-Crick-type TATA tetrad with an
A:T Watson-Crick pair in the large groove of the G tetraplex (WC1-typestructures). (b) Watson-Crick-type TATA tetrad with an A:T Watson-Crick pair in the small groove of the G tetraplex (WC2-type structures). (c)Hoogsteen-type TATA tetrad with an A:T Hoogsteen pair in the largegroove of the G tetraplex (HG1-type structures). (d) Hoogsteen-type TATAtetrad with an A:T Hoogsteen pair in the small groove of the G tetraplex(HG2-type structures). The guanines in the stem have been represented as
rectangular planes similar to those in Fig. 1. The adenine is represented as
rectangular plane with an arrow indicating the side that contains Ni and C2and a bar indicating the side containing N7. Similarly the thymine isrepresented as a square with an arrow indicating the side that contains N3and 02 and a bar indicating the side containing the methyl group. The cafaces of guanine and adenine, and the 13-faces of thymine, when visiblefrom the top, have been shaded. Strands 1 and 2 (and the symmetry-relatedpair 3 and 4) form the large groove of the tetraplex, while strands 1 and 4(and symmetry-related pair 3 and 2) form the small groove.
oms (Rose et al., 1980)) visible from the top and the A:TWatson-Crick pair in the large groove of the G tetraplex,03' atoms of two diagonal guanines will point toward 05'atoms of the thymines and 05' atoms of the other twoguanines will point toward 03' atoms of the adenines, thusleading to stereochemically satisfactory backbone linkage.Hence the A:T Watson-Crick pair can be in the largegroove (as in WCla and WClb) with both T and A beinganti, but, as can be seen from Figs. 3 and 4 b, for the A:TWatson-Crick pair to be in the small groove all the bases of
the TATA tetrad have to be flipped about their glycosidicbonds, thus leading to syn orientation for both T and A.Similarly, when the A:T Hoogsteen pair is in the largegroove (HGla and HGlb) the glycosidic conformations forT and A have to be anti and syn, respectively (Figs. 3 and4 c), but if the A:T Hoogsteen pair is in the small groove(HG2a and HG2b) T is likely to be in the syn conformationand A in the anti conformation (Figs. 3 and 4 d). In the caseof d(G3TTAG3) no hairpin structure of the HGlb or HG2btype is possible, as in these models the loop has to be builtover an A:T pair with the Cl'-Cl' distance being 12.63 A(see Fig. 3 c); it was impossible to bridge this distance witha single thymine i.e., the middle thymine of the TTA loop,but such structures are possible for d(G3T3AG3) hairpins,which have two thymines connecting the A:T pair.As indicated from grid search, because structures with
A:T base pairs in the small groove have glycosidic torsionfor thymine in syn or low anti conformation, which isusually not observed, only two representative structures ofWC2 and HG2 type were considered, whereas for othertypes all possible structures were considered. In addition toWatson-Crick and Hoogsteen ATAT tetrads, the other pos-sible symmetric ATAT tetrads are the ones with reverseWatson-Crick (RWC) or reverse Hoogsteen pairing be-tween the two loops, and in such tetrads adenines can alsoform a diagonal pair with two hydrogen bonds (Chernyi etal., 1990). Thus as a representative case, a structure of theRWC type with the loop going from anti to syn, was alsobuilt. Therefore for d(G3TTAG3) the following hairpinswere considered for detailed energy minimization study:WCla (both possibilities, i.e., loop going from syn to antiand from anti to syn), WClb (both the possibilities, i.e.,loop going from syn to anti and from anti to syn), WC2a(loop going from anti to syn), HGla (both possibilities, i.e.,loop going from syn to anti and from anti to syn), HG2a(loop going from anti to syn), and RWC (loop going fromanti to syn).
In all the structures for d(G3TTAG3) the middle T of theTTA loop remained stacked over the ATAT tetrad while inthe anti conformation and could also form interloop hydro-gen bonds, with two loops on same side, as in the case of thehairpin dimer structures for d(G4T3G4) (Mohanty and Ban-sal, 1994). However, hairpin dimers with loops on oppositeends of the tetraplex can be obtained only by an asymmetriccombination of syn to anti and anti to syn hairpins becauseof the occurrence of three guanines in the stem. Two suchhairpin dimer structures were considered for energy mini-mization, one with intraloop Watson-Crick pairing and theother with intraloop Hoogsteen pairing. No diagonal hairpinstructures were considered for d(G3TTAG3), as they areexpected to be similar to the T3 diagonal structure (Mohantyand Bansal, 1994) in the absence of any obvious basepairing scheme in the loop region. However, an asymmetricdimer (WC1a/WC2a) consisting of large and small groovehairpins with loops on same side and Watson-Crick pairingin the loop was considered for energy minimization. Thisstructure has diagonal strands in antiparallel orientation and
Biophysical Journal1 050
G(a)
5

Chain Folding and A:T Pairing in Human Telomeric DNA
is a representative case of the hairpin dimers involved inIndian-key type of chain folding (Frank-Kamenetskii,1992). The As and Ts in the tetrad in this structure werearranged as TTAA, unlike the TATA tetrads in other struc-tures.The hairpin dimer structures for d(G3T3AG3) with loops
on same side are essentially similar to the structures ford(G3TTAG3) except for the fact that T and A in the loop arebridged by two thymines instead of one thymine as in thecase of d(G3TTAG3). These two thymines can stack overthe TATA tetrad and can form interloop T:T pairing, as inthe case of d(G4T2G4) (Mohanty and Bansal, 1994; Balagu-rumoorthy et al., 1992). Only two models were built for T3Aloop structures, as these structures had certain features dif-ferent from TTA loop structures. One was of the type WClaand in this case both T and A had an anti conformation,unlike the TTA loop structure in which A was in the synconformation. The other was of the type HGlb, which wasnot possible for d(G3TTAG3).
In addition to the hairpin dimer structures, four strands ofd(G3TTAG3) can also adopt a parallel tetraplex structure(Arnott et al.; 1974, Wang and Patel, 1992; Laughlan et al.,1994) with six G tetrads, two T tetrads, and one A tetrad oran antiparallel tetraplex structure with six G tetrads, twoTATA tetrads, and one T tetrad. Therefore a parallel tetra-plex structure was built for d(G4TTAG4) starting from Ar-nott's fiber model (Arnott et al., 1974) and keeping thethymines and adenines in the stem in exactly the sameconformation as for the guanines. The antiparallel tetraplexstructure was built starting from an antiparallel G-tetraplexstructure (Mohanty and Bansal, 1993) of nine nucleotidelengths and replacing the middle three guanines by TIAwith appropriate glycosidic torsion angles so that thyminesand adenines are in the anti conformation.
Intramolecular folded-back structures were also built ford(TTAG3)4, with hairpins with TTA or TT loops and the 5'TlTA or TT kept in a single-stranded helical conformation.
ENERGY MINIMIZATION AND MOLECULARDYNAMICS
The various parallel tetraplex, hairpin dimer, and intramo-lecular folded-back structures obtained from model buildingwere energy minimized to an rms gradient of 0.09 kcal/moleA by use of an AMBER (1987) all-atom force field (Weineret al., 1986). Solvent effects were mimicked implicitly byuse of a distance-dependent dielectric function Eij = Rij(Weiner et al., 1984, 1986; Bansal and Pattabiraman, 1989),and no cutoff was used for nonbonded pairs. In addition tothe calculations carried out with normal AMBER electro-static charges, the structures were also minimized with areduced charge (by an amount 0.7 of electronic charge, sothat total charge on the molecule (became -0.3 per phos-phate group) on anionic phosphate oxygens, to mimic theeffects of counter ions. Such an approach was used earlier inthe simulation ofDNA duplex (Tidor et al., 1983; Ferentz et
al., 1993) and triplex structures (Cheng and Pettitt, 1992);and, as observed in our earlier molecular mechanics study ofG tetraplexes (Mohanty and Bansal, 1993, 1994), it leads togroove widths in better agreement with the crystal structure.In the minimizations no constraints were used on any of thehydrogen bonds or any other parameters.
Molecular dynamics (MD) calculations at constant tem-perature were carried out with AMBER 4.0 for two repre-sentative intramolecularly folded-back structures with se-quence d(G3(TTAG3)3). The MD studies were carried outwith a reduced charge on phosphate oxygens and a distantdependent dielectric function Ei, = Rij similarly to the min-imization studies, but two K+ ions were placed at the twocation binding sites between the three G tetrads. A cutoff of100 A was used to calculate the nonbonded interactions, andnonbonded pairs were updated every 20 steps. The struc-tures were slowly heated in six steps from 0 to 300 K in thefirst 12 ps of the MD at a rate of 50 K for every 2 ps of MDrun. Then the system was coupled to a heat bath at 300 Kand MD was continued for a further period of 338 ps,resulting in a total simulation time of 350 ps. SHAKE, alogarithm, was used to constrain all bonds involving hydro-gens, and a time step of 1 fs was used in the MD integra-tions. The structures were saved after every 100 steps ofMD run, i.e., every 0.1 ps.
RESULTS AND DISCUSSION
Does adenine occur in the loop or in the stem?
A comparison of energy values in energy-minimized fold-ed-back structures reveals that when adenine remains in thestem the structure has better van der Waal's interaction,owing to more favorable stacking of two AGAG tetrads oneither side of the two GGGG tetrads. As can be seen fromFig. 5 the AGAG tetrad has six hydrogen bonds occurringbetween the 06 and Ni atoms of guanine and the N6 andN7 of adenine. In addition N7 of guanine and Ni of adenine
FIGURE 5 Hydrogen bonding pattern in the AGAG tetrad in the energy-minimized structure of d(TTAG3)4 with adenine in the stems and T2 loopsconnecting adjacent strands.
Mohanty and Bansal 1051
Volume 69 September 1995
are within 3 A in the G:A pairs, which have only onehydrogen bond; hence at low pH, if Ni of adenine isprotonated, the AGAG tetrad could be stabilized by eighthydrogen bonds. The better van der Waals stacking in thesestructures is compensated for by less favorable electrostaticinteraction compared with those with TTA loops and threeG tetrads. In fact TTA loop structures have slightly bettertotal energy than structures in which adenine remains in thestem. The structure with adenine in the stem and AGAGtetrads also lacks the quartet of oxygens required for for-mation of the cation binding site (Sundquist and Klug,1989) in the tetraplex stem. Thus in presence of cationsthese structures are likely to be less stable than TTA loopstructures with three G tetrads. The NMR study ofd(AG3(TTAG3)3) indicates that the sequence folds into anIndian-key motif with a G3 stem and a TfA loop. Chemicalprobing studies (Balagurumoorthy and Brahmachari, 1994)on d(TTAG3)4 and corresponding control sequenced(G3TTAG3) also indicate that d(TTAG3)4 adopts a struc-ture with adenine in the loop region. Therefore only thosestructures with G3 stems and ITA loops are discussed indetail.
Energy component analysis for various hairpindimer structures
Energy-minimization studies indicate that the various hair-pin dimer structures described in the section on modelbuilding are energetically almost equally favorable (Table1). Among the models with Watson-Crick and Hoogsteenbase pairs, WCla (a-s) with the loop going from anti to synis the most favorable (-773 kcal/mol), HGla (a-s) is theleast favorable (-734 kcal/mol), and all other models haveintermediate energy values. The small difference in energyarises mostly from bond, angle, and dihedral distortionenergy components, whereas electrostatic and nonbondedenergy components are very similar for all the Watson-Crick and Hoogsteen types of structure. However, the RWCmodel, i.e., with interloop reverse Watson-Crick pairingand diagonal A:A pairing, has almost 10-15 kcal/mol betterelectrostatic energy than other models, mostly because ofloop stem interaction, but this structure has a larger distor-tion energy component; thus there is no overall gain in totalenergy. The WC1a/WC2a structure, which is an asymmetricdimer of two different Watson-Crick hairpins, also hasenergy values similar to those of the symmetric dimers,even though the arrangement of bases in the top tetrad isdifferent. The models with TTA loops on opposite ends ofthe G3 tetraplex have energy values similar to those withloops on same side. In fact the Watson-Crick type ofstructure WC-opp, which consists of two WCla types ofhairpin, one syn to anti and the other anti to syn, has a totalenergy value that is close to the average of the energy ofWCla (a-s) and WCla (s-a), as seen from Table 1. This isalso true for Hoogsteen-type structure HG-opp, with loopson opposite ends of the G3 tetraplex. If we analyze the
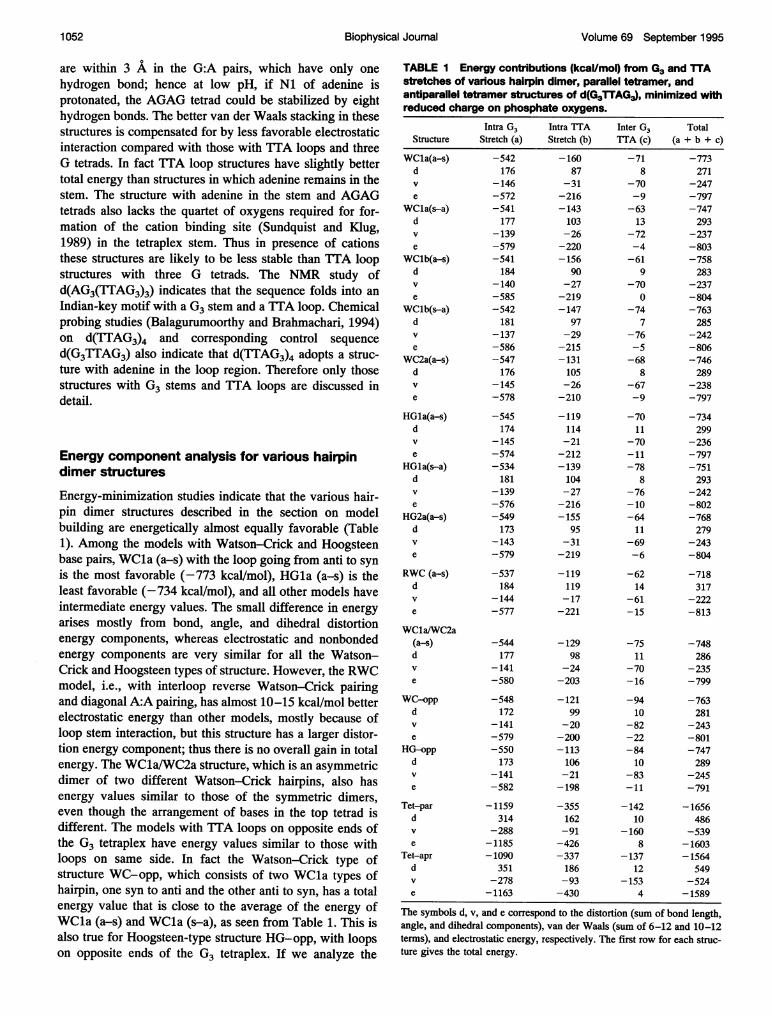
TABLE I Energy contributions (koal/mol) from G3 and TTAstretches of various hairpin dimer, parallel tetramer, andantiparallel tetramer structures of d(G3TTAG3), minimized withreduced charge on phosphate oxygens.
Structure
WCla(a-s)dv
e
WCla(s-a)dv
e
WClb(a-s)dv
e
WClb(s-a)dv
e
WC2a(a-s)dv
e
HGla(a-s)dv
e
HGla(s-a)dv
e
HG2a(a-s)dv
e
RWC (a-s)dv
e
WCla/WC2a(a-s)dv
e
WC-oppdv
e
HG-oppdv
e
Tet-pardv
e
Tet-aprdv
e
Intra G3Stretch (a)
-542176
-146-572-541177
-139-579-541184
-140-585-542181
-137-586-547176
-145-578
-545174
-145-574-534181
-139-576-549173
-143-579
-537184
-144-577
-544177
-141-580
-548172
-141-579-550173
-141-582
-1159314
-288-1185-1090
351-278-1163
Intra TTAStretch (b)-160
87-31-216-143103-26-220-156
90-27-219-147
97-29-215-131105-26-210
-119114-21-212-139104-27-216-155
95-31-219
-119119-17-221
-12998
-24-203
-12199
-20-200-113106-21-198
-355162-91-426-337186-93-430
Inter G3TTA (c)
-718
-70-9
-6313
-72-4-61
9-70
0
-747
-76-5-68
8-67-9
-7011
-70-11-78
8-76-10-6411
-69-6
-6214
-61-15
-7511
-70-16
-9410
-82-22-8410
-83-11
-14210
-1608
-13712
-1534
Total(a + b + c)
-773271
-247-797-747293
-237-803-758283
-237-804-763285
-242-806-746289
-238-797
-734299
-236-797-751293
-242-802-768279
-243-804
-718317
-222-813
-748286
-235-799
-763281
-243-801-747289
-245-791
-1656486
-539-1603-1564
549-524-1589
The symbols d, v, and e correspond to the distortion (sum of bond length,angle, and dihedral components), van der Waals (sum of 6-12 and 10-12terms), and electrostatic energy, respectively. The first row for each struc-ture gives the total energy.
Biophysical Journal1 052

Chain Folding and A:T Pairing in Human Telomeric DNA
energy components in detail (Table 1), we can see that theG3 stem has very similar energy in all the models (near-540 kcal/mol), whereas the energy of the loop varieswithin a range of 40 kcal/mol because of different distortionenergy values for different loops. The loop-stem interactionis more or less similar for various models and varies withina range of 10 kcal/mol. Compared with structures with loopson the same side, the structures with loops on opposite endshave similar stem energy, less favorable intraloop energy,but better loop-stem interaction energy, a trend similar tothat observed for hairpin dimer structures with T2, T3, andT4 loops (Mohanty and Bansal, 1994). The structures whenminimized by normal charge on phosphate oxygens showenergy component trends very similar to those for reducedcharge, and therefore corresponding energy tables have notbeen shown.
Parallel tetraplex versus hairpin dimer structures
The parallel tetraplex structure (Tet-par) for d(G3TTAG3),with all bases in the anti conformation is more favorablethan the antiparallel tetraplex structure (Tet-apr) by almost90 kcal/mol (Table 1) when minimized with reduced charge.In fact an almost 60-kcal/mol difference arises from unfa-vorable distortion energy in the antiparallel structure,whereas the differences arising from van der Waals andelectrostatic components are relatively smaller. This featurehas also been observed for an all-G tetraplex (Mohanty andBansal, 1993). The TTA stretch in the anti structure hasunfavorable distortion energy compared with the TTAstretch in the parallel structure, probably because the start-ing antiparallel structure had an alternating backbone con-formation and only X was changed to the anti conformation.If we compare the conformational energy (reduced charge)of the tetraplexes with that of the hairpin dimer WC1a(a-s)(after scaling it by a factor of 2 to account for the differencein the number of strands), then parallel tetraplex is morefavorable, whereas the antiparallel tetraplex is almost equalin energy. However, for structures minimized with a normalcharge on phosphate oxygens the hairpin dimers are morefavorable because of large phosphate-phosphate repulsionin tetramolecular structures. Thus tetramolecular structuresare likely to be observed only if phosphate charges arescreened by counter ions at high salt concentrations. Ourearlier molecular mechanics calculations of d(G4TnG4) hadindicated a similar result. In fact it was shown recently fromRaman spectroscopic studies that, although d(T4G4)4 adoptsa folded-back structure at low salt concentrations, it adoptsa parallel tetraplex structure at high salt (Miura and Thomas,1994).
It is important to note that, in addition being stabilized byan ambient counter-ion atmosphere, the effect of which wasmodeled by reduction of the charge on phosphate groups,the G tetraplexes are also known to be stabilized by bindingof K+ ions to the central cavity between two adjacent G
earlier molecular mechanics study of d(G4T4G4) had indi-cated that the structural features of G tetraplexes minimizedwith K+ ions in the cavities were almost identical to thoseminimized without K+ ions. Hence in the present study thevarious types of parallel and folded-back structure havebeen energy minimized, without K+ ions being put betweenadjacent G tetrads, and the minimization was carried outwithout the imposition of any constraints. In spite of that,the K+ binding site is preserved in all the models. Ourearlier energy-minimization study of d(G4T4G4), which in-cluded K+ ions in the cavity, also indicated that the bindingof K+ ions in the cavities of the G-tetraplex stem contrib-utes a significant stabilization energy to the structure, inagreement with the experimental observations. However,the energy contribution from K+ binding to the G4 stem isnearly identical for all types of tetraplex, i.e., parallel,Greek-key-type folded-back, and Indian-key-type folded-back structures (Mohanty and Bansal, 1994). This can beattributed to the fact that in all three types of G tetraplexesthe K+-O distance remains the same, regardless of thetwist between the G tetrads or the types of stacking arrange-ment i.e., normal or inverted stacking. Hence binding of K+to the central cavities is unlikely to stabilize preferentiallyany particular type of G tetraplex compared with the others,but can stabilize G tetraplexes of all types, as reportedalready for antiparallel and parallel structures (Kang et al.,1992; Gupta et al., 1993).
Analysis of structural features in theenergy-minimized structures
Stacking and hydrogen bonding in hairpin dimer structures
Stacking and hydrogen bonding in various energy-mini-mized hairpin dimer structures are shown in Fig. 6 a-h. It isinteresting to note that the various hydrogen bonds in theloop region are retained in the energy-minimized structures,even though no constraints were applied to the hydrogenbonds during minimization. Table 2 gives a list of varioushydrogen bonds in the loop region, the H-Y distances, andX-H-Y angle values. In models WCla, WC2a, HGla, andHG2a, because the ]TA loop can have perfect AT basepairs with Watson-Crick or Hoogsteen pairing, these struc-tures are effectively hairpins with a single thymine in theloop. It is important to note that a similar single base loopis not possible over a GG pair (Balagurumoorthy et al.,1992) if the guanines are constrained to have conformationsidentical to those of guanines in the tetraplex stem. Instructures with Watson-Crick and Hoogsteen type hydrogenbonded TATA tetrads (Fig. 6 a-e) the methyl hydrogens ofthymines remain close (-3.0 A) to N7 or Ni of adenine,thus giving rise to the additional possibility of C-H-Nhydrogen bond formation, similar to C-H-O hydrogenbonds (Taylor and Kennard, 1982; Bruskov et al., 1989;Zhurkin et al., 1994). In all the structures shown in Fig. 6a-f the N7 of adenine is either involved in hydrogen bond-ing or is close to a methyl group hydrogen, thus making it
Mohanty and Bansal 1 053
tetrads (Kang et al., 1992; Gupta et al., 1993). However, our

Volume 69 September 1995
b
C 9
d
FIGURE 6 Hydrogen bonding pattern in a TATA or TTAA tetrad andstacking of this tetrad (thick lines) over the guanine tetrads in varioushairpin dimer models of d(G3TTAG3) with two loops on same side. Theresidues are numbered Gl-G2-G3-T4-T5-A6-G7-G8-G9 in one hairpin andG1O-Gll-G12-T13-T14-A15-G16-G17-G18 in the other hairpin of thedimer. (a) WCla(a-s), (b) WClb(a-s), (c) WC2a(a-s), (d) HGla(a-s), (e)HG2a(a-s), (f) RWC(a-s) and (g) an asymmetric hairpin dimer ofWCla(a-s) and WC2a(a-s), with the tetraplex stem in an Indian-keyarrangement (i.e., diagonal strands are antiparallel). In a-g, only backboneatoms have been shown for the middle thymines T(5) and T(14). (h)Representative example of the stacking of the TT pair (thick lines) formedby T(5) and T(14) over the TATA tetrad (thin lines) in the structureWCla(a-s).
less accessible to chemical probes; whereas in the TTAAtetrad shown in Fig. 6 g the N7 of both the adenines aremore accessible. Chemical probing experiments can there-fore distinguish among various types of models discussedhere, and the results of such studies (Balagurumoorthy andBrahmachari, 1994) in fact suggest the possibility of A:Tpairing in the loop region.
In TTA loop structures with loops on same side theTATA as well as the G tetrads have very little buckle andpropeller-type distortion, and the top TT pair is also stackedover the TATA tetrad (shown in Fig. 6 h).The various models with TTA loops, although they are
similar in terms of energy (Table 1), have distinct structuralfeatures that can give rise to unique cross-peaks in nuclearOverhauser effect spectroscopy (NOESY) experiments. Ta-ble 3 gives, for five representative models, a list of hydro-gen pairs that are separated by less than 4 A in some models,although the separation is much larger in other models. Ascan be seen from Table 3 and the stacking diagrams (Fig. 6),H8 ofA6 will come close (3.3 A) to H8 of G3 only in modelHG2a(a-s) (Fig. 6 e); in models (Fig. 6 a-d) that have A:Tpairs in the large grooves or Watson-Crick A:T pairs in thesmall grooves this distance is larger than 8 A. The modelWC2a(a-s) (Fig. 6 c) has H8 of G3 at a distance of 3.3 Afrom H2 of A6; the same proton pair is separated by morethan 7 A in all other models (Fig. 6 a, b, d, e), which haveA:T pairs in the large grooves or Hoogsteen-type A:T pairsin the small grooves. Similarly, NOE will be observedbetween HN2A/HN2B protons of G3 and H2 of A6 only inmodel WC1a(a-s) (Fig. 6 a), which has a WC A:T pair inthe large groove. Table 3 also indicates that NOEs shouldgenerally be observed between H3 of T4 and either H2 orH8 of A6, corresponding to Watson-Crick or HoogsteenA:T pairs, respectively. Thus these NOEs could be used asdiagnostic features for these structures.
Backbone and glycosidic torsion angles
In all the hairpin dimer structures for d(G3TTAG3) theguanines that are one nucleotide away from the loop allhave torsion angles identical to those in the antiparallel Gtetraplex without the loops; i.e., a, ,B, 'y, 5, E, and r forguanine in the syn conformation are in the range g-, g-,g, t(2E), t, and g, and for guanines in the anti confor-mation they are in the g-, t, g+, t(1E), t, and g- regions.Even in the guanines, immediately flanking the loop atthe 5' and the 3' ends, only a few torsion angles havechanged from their values in the tetraplex stem. Thebackbone torsion angles for the nucleotides in the looptake up a wide range of values in various models, but theglycosidic torsion is the one that is directly correlated tothe chain folding and the type of base pairing in the loop,as discussed in the Model Building section. The glyco-sidic torsion angles (Table 4) for thymines and adeninesin various models remain close to their values in thestarting structures. In models WCla(a-s) and WCla-(s-a), which have loops across the large grooves and
Biophysical Journal1054
f
Chain Folding and A:T Pairing in Human Telomeric DNA
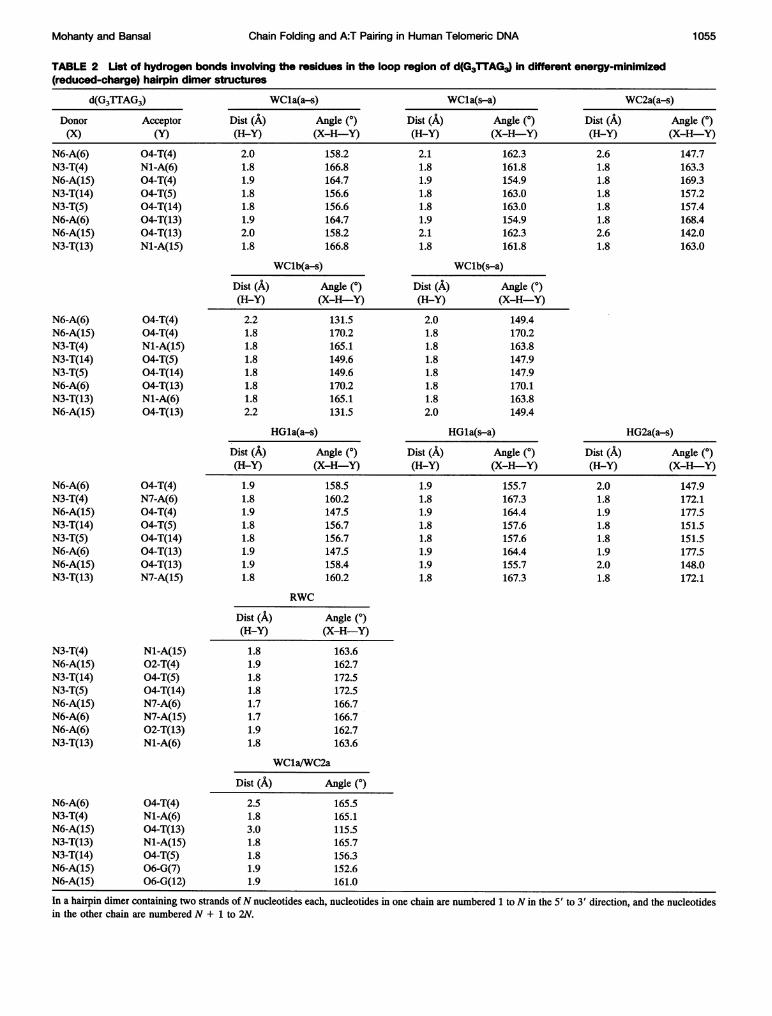
TABLE 2 List of hydrogen bonds involving- the residues in the loop region of d(G3TTAG3) in different energy-minimized(reduced-charge) hairpin dimer structures
d(G3TTAG3) WCla(a-s) WCIa(s-a) WC2a(a-s)Donor Acceptor Dist (A) Angle (0) Dist (A) Angle (0) Dist (A) Angle (0)(X) (Y) (H-Y) (X-H-Y) (H-Y) (X-H-Y) (H-Y) (X-H-Y)
N6-A(6) 04-T(4) 2.0 158.2 2.1 162.3 2.6 147.7N3-T(4)N6-A(15)N3-T(14)N3-T(5)N6-A(6)N6-A(15)N3-T(13)
N1-A(6)04-T(4)04-T(5)04-T(14)04-T(13)04-T(13)Nl-A(15)
1.81.91.81.81.92.01.8
166.8164.7156.6156.6164.7158.2166.8
1.81.91.81.81.92.11.8
161.8154.9163.0163.0154.9162.3161.8
1.81.81.81.81.82.61.8
163.3169.3157.2157.4168.4142.0163.0
WClb(a-s)
Dist (A) Angle (0)(H-Y) (X-H-Y)2.21.81.81.81.81.81.82.2
131.5170.2165.1149.6149.6170.2165.1131.5
WClb(s-a)
Dist (A) Angle (0)(H-Y) (X-H-Y)2.01.81.81.81.81.81.82.0
149.4170.2163.8147.9147.9170.1163.8149.4
HGla(a-s)
Dist (A) Angle (0)(H-Y) (X-H-Y)1.91.81.91.81.81.91.91.8
158.5160.2147.5156.7156.7147.5158.4160.2
HGla(s-a)
Dist (A) Angle (0)(H-Y) (X-H-Y)1.9 155.71.8 167.31.9 164.41.8 157.61.8 157.61.9 164.41.9 155.71.8 167.3
HG2a(a-s)
Dist (A) Angle (0)(H-Y) (X-H-Y)2.0 147.91.8 172.11.9 177.51.8 151.51.8 151.51.9 177.52.0 148.01.8 172.1
RWC
Dist (A) Angle (0)(H-Y) (X-H-Y)1.81.91.81.81.71.71.91.8
163.6162.7172.5172.5166.7166.7162.7163.6
WCla/WC2a
Dist (A) Angle (0)2.51.83.01.81.81.91.9
165.5165.1115.5165.7156.3152.6161.0
In a hairpin dimer containing two strands ofN nucleotides each, nucleotides in one chain are numbered 1 to N in the 5' to 3' direction, and the nucleotidesin the other chain are numbered N + 1 to 2N.
N6-A(6)N6-A(15)N3-T(4)N3-T(14)N3-T(5)N6-A(6)N3-T(13)N6-A(15)
04-T(4)04-T(4)Nl-A(15)04-T(5)04-T(14)04-T(13)N1-A(6)04-T(13)
N6-A(6)N3-T(4)N6-A(15)N3-T(14)N3-T(5)N6-A(6)N6-A(15)N3-T(13)
04-T(4)N7-A(6)04-T(4)04-T(5)04-T(14)04-T(13)04-T(13)N7-A(15)
N3-T(4)N6-A(15)N3-T(14)N3-T(5)N6-A(15)N6-A(6)N6-A(6)N3-T(13)
Nl-A(15)02-T(4)04-T(5)04-T(14)N7-A(6)N7-A(15)02-T(13)N1-A(6)
N6-A(6)N3-T(4)N6-A(15)N3-T(13)N3-T(14)N6-A(15)N6-A(15)
04-T(4)N1-A(6)04-T(13)Nl-A(15)04-T(5)06-G(7)06-G(12)
Mohanty and Bansal 1 055
~~~~~~~~~~Volume69 September 1995
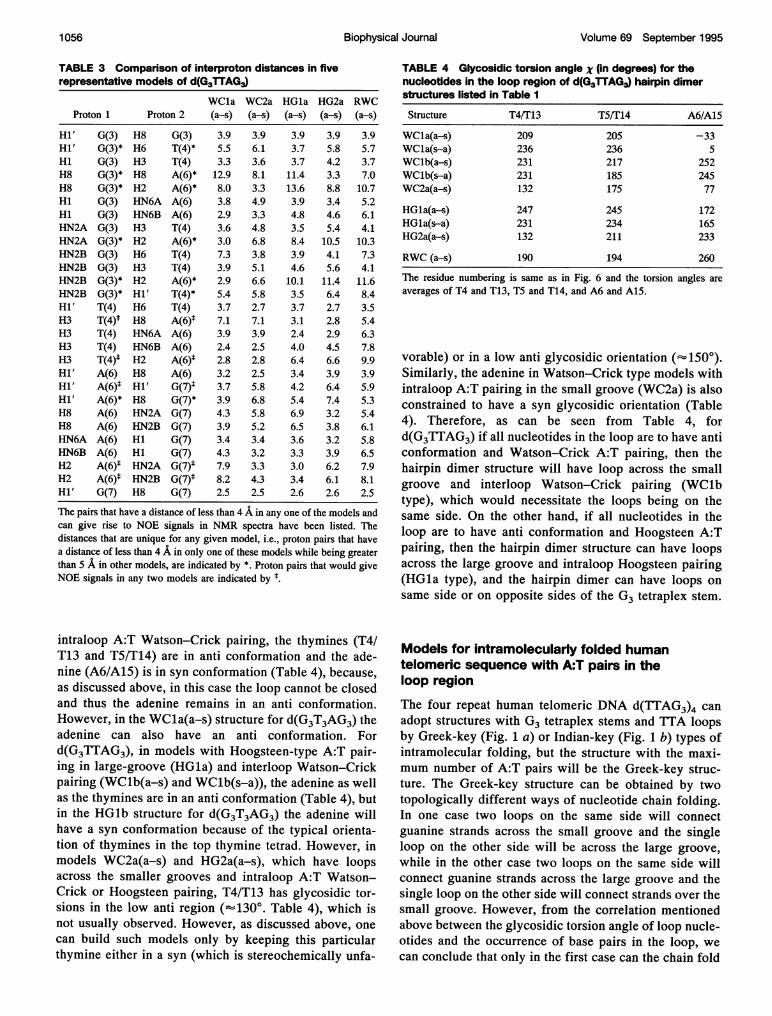
TABLE 3 Comparison of interproton distances in fiverepresentative models of d(G3TTAG3)
WCla WC2a HGla HG2a RWCProton 1 Proton 2 (a-s) (a-s) (a-s) (a-s) (a-s)
Hi' G(3) H8 G(3) 3.9 3.9 3.9 3.9 3.9Hi' G(3)* H6 T(4)* 5.5 6.1 3.7 5.8 5.7Hi G(3) H3 T(4) 3.3 3.6 3.7 4.2 3.7H8 G(3)* H8 A(6)* 12.9 8.1 11.4. 3.3 7.0H8 G(3)* H2 A(6)* 8.0 3.3 13.6 8.8 10.7Hi G(3) HN6A A(6) 3.8 4.9 3.9 3.4 5.2Hi G(3) HN6B A(6) 2.9 3.3 4.8 4.6 6.iHN2A G(3) H3 T(4) 3.6 4.8 3.5 5.4 4.1HN2A G(3)* H2 A(6)* 3.0 6.8 8.4 10.5 10.3HN2B G(3) H6 T(4) 7.3 3.8 3.9 4.1 7.3HN2B G(3) H3 T(4) 3.9 5.1 4.6 5.6 4.1HN2B G(3)* H2 A(6)* 2.9 6.6 10.1 11.4 11.6HN2B G(3)* Hi' T(4)* 5.4 5.8 3.5 6.4 8.4Hi' T(4) H6 T(4) 3.7 2.7 3.7 2.7 3.5H3 T(4)t H8 A(6)1 7.1 7.1 3.1 2.8 5.4H3 T(4) HN6A A(6) 3.9 3.9 2.4 2.9 6.3H3 T(4) HN6B A(6) 2.4 2.5 4.0 4.5 7.8H3 T(4)* H2 A(6)* 2.8 2.8 6.4 6.6 9.9Hi' A(6) H8 A(6) 3.2 2.5 3.4 3.9 3.9Hi' A(6)* Hi' G(7)t 3.7 5.8 4.2 6.4 5.9Hi' A(6)* H8 G(7)* 3.9 6.8 5.4 7.4 5.3H8 A(6) HN2A G(7) 4.3 5.8 6.9 3.2 5.4H8 A(6) HN2B G(7) 3.9 5.2 6.5 3.8 6.1HN6A A(6) HI G(7) 3.4 3.4 3.6 3.2 5.8HN6B A(6) Hi G(7) 4.3 3.2 3.3 3.9 6.5H2 A(6)1 HN2A G(7)* 7.9 3.3 3.0 6.2 7.9H2 A(6)t HN2B G(7)* 8.2 4.3 3.4 6.1 8.1Hi' G(7) H8 G(7) 2.5 2.5 2.6 2.6 2.5
The pairs that have a distance of less than 4 A in any one of the models andcan give rise to NOE signals in NMR spectra have been listed. Thedistances that are unique for any given model, i.e., proton pairs that havea distance of less than 4 A in only one of these models while being greaterthan 5 A in other models, are indicated by *. Proton pairs that would giveNOE signals in any two models are indicated by ~
intraloop A:T Watson-Crick pairing, the thymines (T41T13 and T5/T14) are in anti conformation and the ade-nine (A6/A15) is in syn conformation (Table 4), because,as discussed above, in this case the ioop cannot be closedand thus the adenine remains in an anti conformation.However, in the WCla(a-s) structure for d(G3T3AG3) theadenine can also have an anti conformation. Ford(G3TTAG3), in models with Hoogsteen-type A:T pair-ing in large-groove (HGla) and interloop Watson-Crickpairing (WClb(a-s) and WClb(s-a)), the adenine as wellas the thymines are in an anti conformation (Table 4), butin the HGlb structure for d(G3T3AG3) the adenine willhave a syn conformation because of the typical orienta-tion of thymines in the top thymine tetrad. However, inmodels WC2a(a-s) and HG2a(a-s), which have loopsacross the smaller grooves and intraloop A:T Watson-Crick or Hoogsteen pairing, T4/T13 has glycosidic tor-sions in the low anti region (-13' Table 4), which isnot usually observed. However, as discussed above, onecan build such models only by keeping this particular
TABLE 4 Glycosidic torsion angle X (in degrees) for the
nucleotides in the loop region of d(G3TTAG3) hairpin dimer
structures listed in Table I
Structure T4IT13 TSF[ 14 A6/A15
WCla(a-s) 209 205 -33
WCla(s-a) 236 236 5
WClb(a-s) 231 217 252
WClb(s-a) 231 185 245
WC2a(a-s) 132 175 77
HGla(a-s) 247 245 172
HlGla(s-a) 231 234 165
HG2a(a-s) 132 211 233
RWC (a-s) 190 194 260
The residue numbering is same as in Fig. 6 and the torsion angles are
averages of T4 and T13, TS and T14, and A6 and A15.
vorable) or in a low anti glycosidic orientation (- 1500).
Similarly, the adenine in Watson-Crick type models with
intraloop A:T pairing in the small groove (WC2a) is also
constrained to have a syn glycosidic orientation (Table
4). Therefore, as can be seen from Table 4, for
d(G3TTAG3) if all nucleotides in the loop are to have anti
conformation and Watson-Crick A:T pairing, then the
hairpin dimer structure will have loop across the small
groove and interloop Watson-Crick pairing (WClbtype), which would necessitate the loops being on the
same side. On the other hand, if all nucleotides in the
loop are to have anti conformation and Hoogsteen A:T
pairing, then the hairpin dimer structure can have loopsacross the large groove and intraloop Hoogsteen pairing
(HGla type), and the hairpin dimer can have loops on
same side or on opposite sides of the G3 tetraplex stem.
Models for intramolecularly folded human
telomeric sequence with A:T pairs in the
loop region
The four repeat human telomeric DNA d(TTAG3)4 can
adopt structures with G3 tetraplex stems and TTA loops
by Greek-key (Fig. 1 a) or Indian-key (Fig. 1 b) types of
intramolecular folding, but the structure with the maxi-
mum number of A:T pairs will be the Greek-key struc-
ture. The Greek-key structure can be obtained by two
topologically different ways of nucleotide chain folding.In one case two loops on the same side will connect
guanine strands across the small groove and the single
loop on the other side will be across the large groove,
while in the other case two loops on the same side will
connect guanine strands across the large groove and the
single loop on the other side will connect strands over the
small groove. However, from the correlation mentioned
above between the glycosidic torsion angle of loop nudle-otides and the occurrence of base pairs in the loop, we
thymine either in a syn (which is stereochemically unfa -ca ocueta nyi h isaecntecanflcan conclude that only in the first case can the chain fold
1056 Biophysical Journal
Chain Folding and A:T Pairing in Human Telomeric DNA
FBI
o<a1cni
rC')
010 500 1000 1500 2000 2500 3000
Time (x 0.1 ps)3500
FB2
500 1000 1500 2000 2500 3000Time (x 0.1 ps)
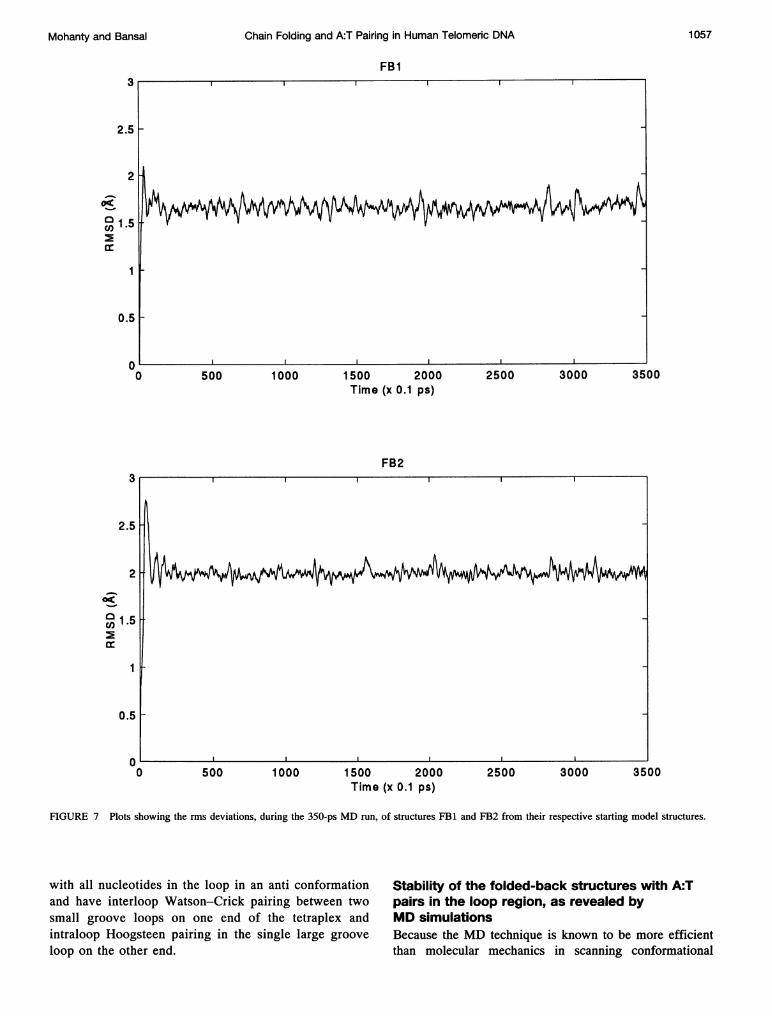
FIGURE 7 Plots showing the rms deviations, during the 350-ps MD run, of structures FB1 and FB2 from their respective starting model structures.
with all nucleotides in the loop in an anti conformationand have interloop Watson-Crick pairing between twosmall groove loops on one end of the tetraplex andintraloop Hoogsteen pairing in the single large groove
loop on the other end.
Stability of the folded-back structures with A:Tpairs in the loop region, as revealed byMD simulationsBecause the MD technique is known to be more efficientthan molecular mechanics in scanning conformational
3
2.5
2
04
v3 1.5
0.5
0-0 3500
Mohanty and Bansal 1057

Volume 69 September 1995
aHN2 GUA_0 --- N7 GUA_1
o4z3a
1
0 1000 2000 3000
1L0 1000 2000 3000
Time (x 0.1 ps)
bHN2 GUA_9 --- N7 GUA_1
4
;3
12 1
0 1000 2000 3000
HN1 GUA_9 --- 06 GUA_14
1-1
0 1000 2000 3000
HN1 GUA_1 --- 06 GUA-214
oil2 1mi_h*w
0 1000 2000 3000
HNI GUA_21 --- 06 GUA_134
010 1000 2000 3000
1000 2000 3000Time (x 0.1 ps)
HN1 GUA_0 --- 06 GUA_14
9 2
112
0 1000 2000 3000
HN2 GUA_1 --- N7 GUA_21 HN1 GUA_1 --- 06 GUA-214 ~~~~~~~~~~~4
11
0 1000 2000 3000 0 1000 2000 3000
HN2 GUA_21 --- N7GUA13 HN1 GUA-21 --- 06 GUA_0134 4
AZ - Z2 2
0 1000 2000 3000 0 1000 2000 3000
HN2GUA 13 N7GUA 9 HNI1GUA 13 ---O0BQUA..9
1~~~~~~~~~~~~~~~~ 1I0 1000 2000 3000 0 1000 2000 3000
Time (x 0.1 ps) Time (x 0.1 ps)
FIGURE 8 Plots of hydrogen-to-acceptor atom distance versus time forall the eight hydrogen bonds in the G tetrad, consisting of guanines Gl, G9,G13, and G21 in the intramolecular folded-back structures: (a) FB1, (b)FB2.
space, we carried out MD simulations for two representativefolded-back structures for d(G3(1TAG3)3) to find outwhether these structures really represent energetically stableminima in the conformational space. Both of the structuresconsidered had Greek-key-type folding, because thesestructures have the maximum number of A:T pairs, but thefolding topology and base parings in the loop region weredifferent in the two structures. In one, the two loops on thesame side were over the small groove joining a guanine inan anti conformation to a guanine in a syn conformation andwith interloop A:T Watson-Crick pairs, and the single loopon the other end joining a guanine in a syn conformation toa guanine in an anti had intraloop Hoogsteen A:T pairs.Thus the folded-back structure had two hairpin loops of theWClb(a-s) type on one end (as shown in Fig. 6 b) and asingle hairpin of the HGla(s-a) type on the other end. Thisstructure will be called FB1. The other folded-back structure(FB2) had hairpin loops of the HG1a(s-a) type on one endand a single hairpin of the type WC2a(a-s) on the other end.As mentioned in the methods the MD calculations were
carried out with distance-dependent dielectric function andreduced charge on the phosphate groups, as for the molec-ular mechanics studies. However, because the structureswere expected to undergo relatively larger structuralchanges during the MD simulations than those observedduring energy minimization, there was a possibility of theK+ binding site's being distorted in the absence of explicitK+ ions. Therefore in the MD calculations two K+ ionswere included between the three G tetrads of the folded-back structures FBl and FB2. A 350- ps MD simulation wascarried out for each of the folded-back structures, at aconstant temperature of 300 K (by coupling to a heat bath).For all the structures saved along the trajectory, rms devi-ations from the starting structure was calculated, and theplot of rms deviation versus time is shown in Fig. 7. As canbe seen from that figure, after the first few picoseconds ofthe MD run, while the rms deviation from the startingstructure stabilizes in the range between 1.6 and 1.8 A forstructure FB1, the rms deviation for FB2 stabilizes at avalue close to 2.0 A. This stable value of the rms deviationindicates that the structures are well equilibrated. The rmsdeviations are reasonably small (<2.0 A) for all the 595atoms, including hydrogens. Thus the MD simulationclearly shows that the structures obtained from our modelbuilding and energy minimization are energetically quitestable.The stability of the folded-back structures is further evi-
dent from an analysis of the various hydrogen bonds in thestem as well as the loop regions. In the stem region of thefolded-back structures almost all the hydrogen bonds re-main intact throughout the MD run. Fig. 8 a and b shows thetrajectories for the all eight hydrogen bonds in the first Gtetrads of FB1 and FB2 as representative examples. It can beseen from Fig. 8 that the hydrogen-to-acceptor distancefluctuates around 1.9 A for all hydrogen bonds. In the caseof FB1 only the hydrogen bond HN2 G1-N7 G21 breaks(i.e. the HN2-N7 distance increases to 3.5 A) during the
1 058 Biophysical Journal
Chain Folding and A:T Pairing in Human Telomeric DNA
HN6 ADE_18--- 04 THY_4
10
0
Cu-0
D
1000 2000 3000
HN6ADE_6---04THY_16
2 fim a -k ~ M j L A~
10 1000 2000 3000
HN6 ADE_6---04 THY_44
'3
I
HN3 THY_4 --- N1 ADE_184.
3.-
1*0 1000 2000 3000
HN3THY_16---NIADE_64
o<,., 30~
2._
a
0
0 Li- s*a...Li.1002i00 300L
0 1000 2000 3000
HN6ADE_18--- 04 THY_164
0<,-,30~U._
coCU2z 1fIijrT Mif trf-T1 w !F1nrr wr- 1r pi
0_ 1000 2000 3000
HN6 ADE_12--- 04 THY_10
0 1000 2000Time (x 0.1 ps)
3000
1'0 1000 2000 3000
HN3 THY_10 --- N7 ADE_124
0, 3-2 ,L10 1000 2000
Time (x 0.1 ps)3000
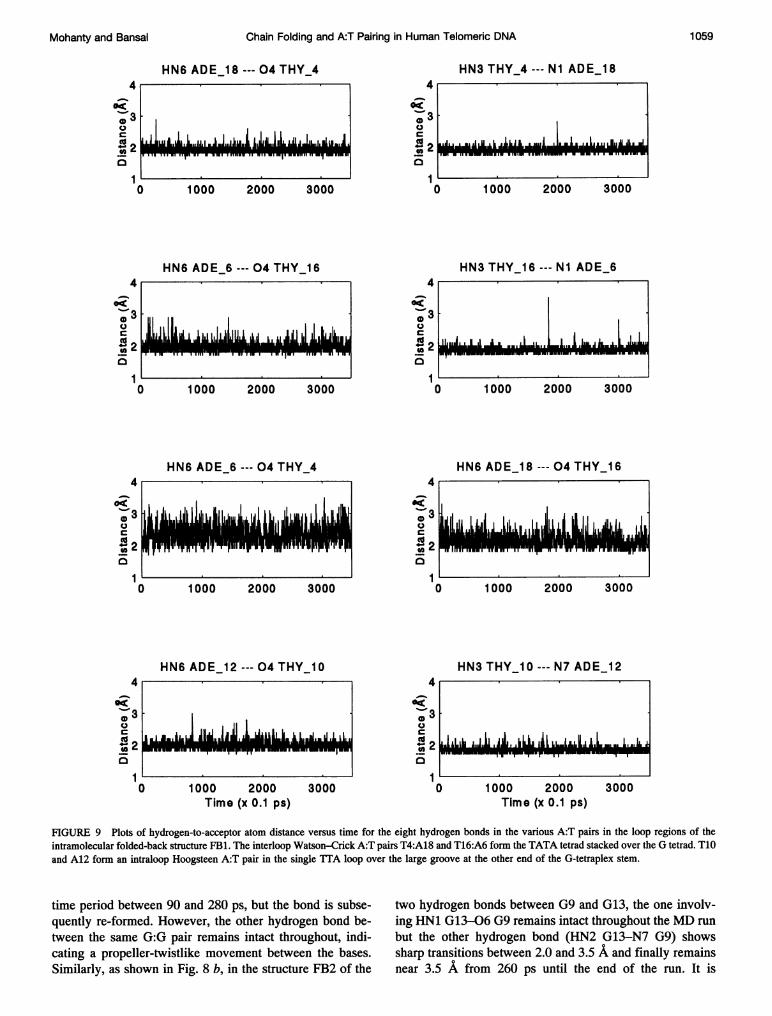
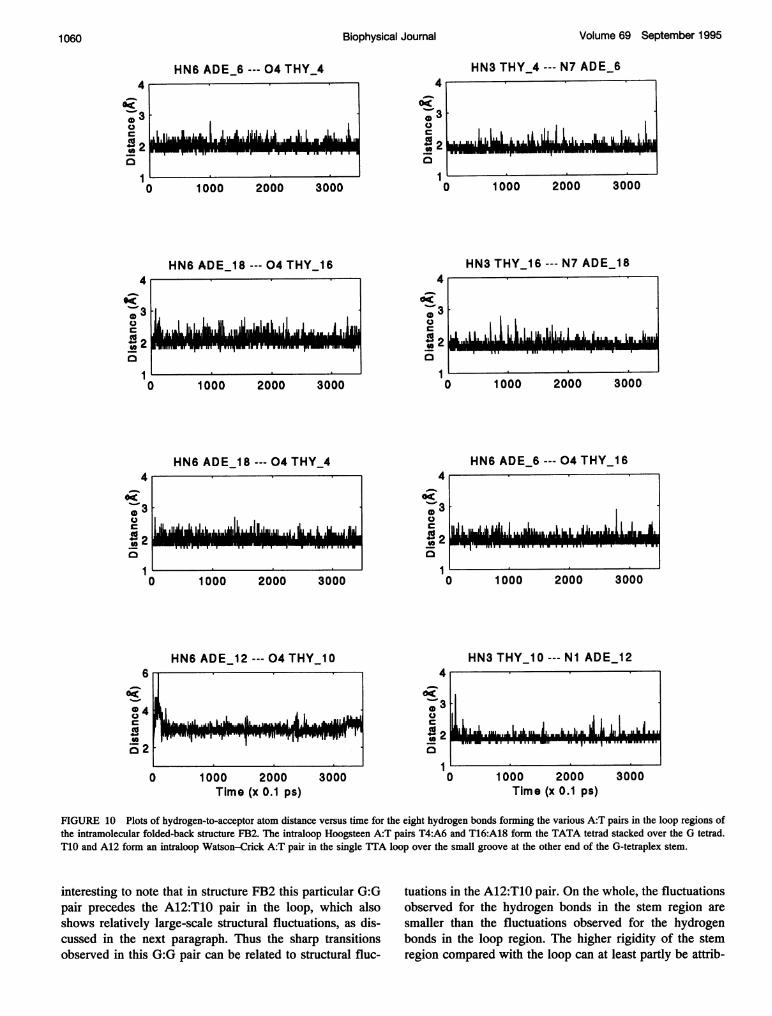
FIGURE 9 Plots of hydrogen-to-acceptor atom distance versus time for the eight hydrogen bonds in the various A:T pairs in the loop regions of theintramolecular folded-back structure FB1. The interloop Watson-Crick A:T pairs T4:A18 and T16:A6 form the TATA tetrad stacked over the G tetrad. T10and A12 form an intraloop Hoogsteen A:T pair in the single TTA loop over the large groove at the other end of the G-tetraplex stem.
time period between 90 and 280 ps, but the bond is subse-quently re-formed. However, the other hydrogen bond be-tween the same G:G pair remains intact throughout, indi-cating a propeller-twistlike movement between the bases.Similarly, as shown in Fig. 8 b, in the structure FB2 of the
two hydrogen bonds between G9 and G13, the one involv-ing HN1 G13-06 G9 remains intact throughout the MD run
but the other hydrogen bond (HN2 G13-N7 G9) showssharp transitions between 2.0 and 3.5 A and finally remainsnear 3.5 A from 260 ps until the end of the run. It is
4
0U_LD a.* 2 1J
._0 v III 1-ii-i Frro g I||Ivnw WI I M ,"II igii I1 l NMw
4
,3U
..1IL .1 ilaIIliJiLu .1 L. ,.lII LIiJmLilMuLw
._
0
ci5-
4
.3UCD 3
0._
i
:
l
1 059Mohanty and Bansal
I
a I III A til L II I I I this i t jig loll I ji I -III I mtla- I
-i II& J6 . L,IF-r7-
Volume 69 September 1995
HN6 ADE_6--- 04 THY_4
0 1000 2000 3000
HN6 ADE_18--- 04 THY_16
0302co _.I II I u1.-4L L" aI 2.
4-
.b30
0
C
HN3 THY_4---N7 ADE_6
. . L.LJ, f L1_. ILI 1L.A
1000 2000 3000
HN3 THY_16---N7 ADE_18A rat
&C-3
2
1 I._
1000 2000 3000
HN6ADE_18---04THY_4
MAL.AWI166 IMh&pME.qii- -Li& L.my miliii 1.1...I I
1000 2000 3000
HN6 ADE_6--- 04 THY_164
07
.,3
cz-21to
0 1000 2000 3000
HN6 ADE_12--- 04 THY_10
0 1000 2000Time (x 0.1 ps)
3000
1000 2000 3000
HN3THY_10---N1ADE_12
1000 2000Time (x 0.1 ps)
FIGURE 10 Plots of hydrogen-to-acceptor atom distance versus time for the eight hydrogen bonds forming the various A:T pairs in the loop regions ofthe intramolecular folded-back structure FB2. The intraloop Hoogsteen A:T pairs T4:A6 and T16:A18 form the TATA tetrad stacked over the G tetrad.T1O and A12 form an intraloop Watson-Crick A:T pair in the single TTA loop over the small groove at the other end of the G-tetraplex stem.
interesting to note that in structure FB2 this particular G:Gpair precedes the A12:T1O pair in the loop, which alsoshows relatively large-scale structural fluctuations, as dis-cussed in the next paragraph. Thus the sharp transitionsobserved in this G:G pair can be related to structural fluc-
tuations in the A12:T10 pair. On the whole, the fluctuationsobserved for the hydrogen bonds in the stem region are
smaller than the fluctuations observed for the hydrogenbonds in the loop region. The higher rigidity of the stemregion compared with the loop can at least partly be attrib-
A r
so.3
,-,0
0g42
49i
0w-
10
_Li"dab L.LA ItI1LUI .jL.I. I
4
.-307
0
C
.-S20
1cz
6
o<
0 4
07
C
co
IJI ILJ.ii l.I .. ILi.. .1 L6 ., 1I LI-= ==F=fvl
,6AjW6jM=Lm&LA.M.Aldh selAAME toME mammiumn WWI lownwom WMININEWr..
;
Biophysical Journal1060
L
.11.1"66did 2 Ili., iiwil14 la I- t- -0
I It mrmmFOM" TT-
6iI
LL i L L.1 , L .u I ai &-- In- -knii

Chain Folding and A:T Pairing in Human Telomeric DNA
uted to the presence of two K+ ions between the G tetradsof the G3 stem.
Most of the hydrogen bonds in the loop regions of both ofthe folded-back structures also remain intact during theentire duration of the MD simulation, with only three hy-drogen bonds being broken, i.e., the hydrogen-to-acceptordistance becomes more than 2.5 A. Figs. 9 and 10 show theplots for all the hydrogen bonds that occur in the TATAtetrad and also in the isolated A:T pair in the single loop inthe folded-back structures FB1 and FB2. As can be seenfrom Fig. 9, in structure FB1 all the hydrogen bonds in thetwo interloop Watson-Crick pairs (i.e., A18:T4 and A6:T16) and in the intraloop Hoogsteen pair (A12:T10) remainintact and show a mean fluctuation of only 0.2 A about themean hydrogen-to-acceptor distance of 2.0 A, with a fewexcursions to slightly longer distances. However, the intra-loop hydrogen bonds in the TATA tetrad of FB1 (i.e., HN6A6-04 T4 and HN6 A18-04 T16) show larger fluctu-ations, in the range between 1.9 and 3.0 A, indicating thatthese hydrogen bonds break and form during the course ofthe simulation.
In structure FB2 all the intraloop as well as the interloophydrogen bonds in the TATA tetrad remain intact andfluctuate about the mean hydrogen-to-acceptor distance of2.0 A (Fig. 10). On the other hand, the two hydrogen bondsin the intraloop WC A12:T10 pair are initially broken. Ascan be seen from Fig. 10, at the very beginning of thesimulation, while the HN6 A12-04 T10 distance increasesto 6.0 A, the HN3 T10-Ni A12 distance approaches 3.4 A,indicating a complete rupture of this A:T pair. However,after -30 ps of the MD run the hydrogen bond HN3T10-Ni A12 is again formed and remains close to 2.0 Athroughout the rest of the MD run, but the potential hydro-gen bond between HN6 A12-04 T10 is not formed, al-though the distance does reduce to -3.0 A after 30 ps andfluctuates about that value for the rest of the 350-ps MD run.It may be noted that this particular A:T pair is a WC pair inthe small groove, and, as discussed in the model buildingsection, it is essential for the thymine to be in syn or low antiglycosidic orientation to retain both the hydrogen bonds inthis A:T pair. As discussed in the next subsection, thebreakage of one of the hydrogen bonds in the A12:T10 pairis correlated to a transition in the glycosidic orientation ofT10. The hydrogen bonds linking the T:T pairs in both ofthe folded-back structures remain intact throughout the350-ps run.
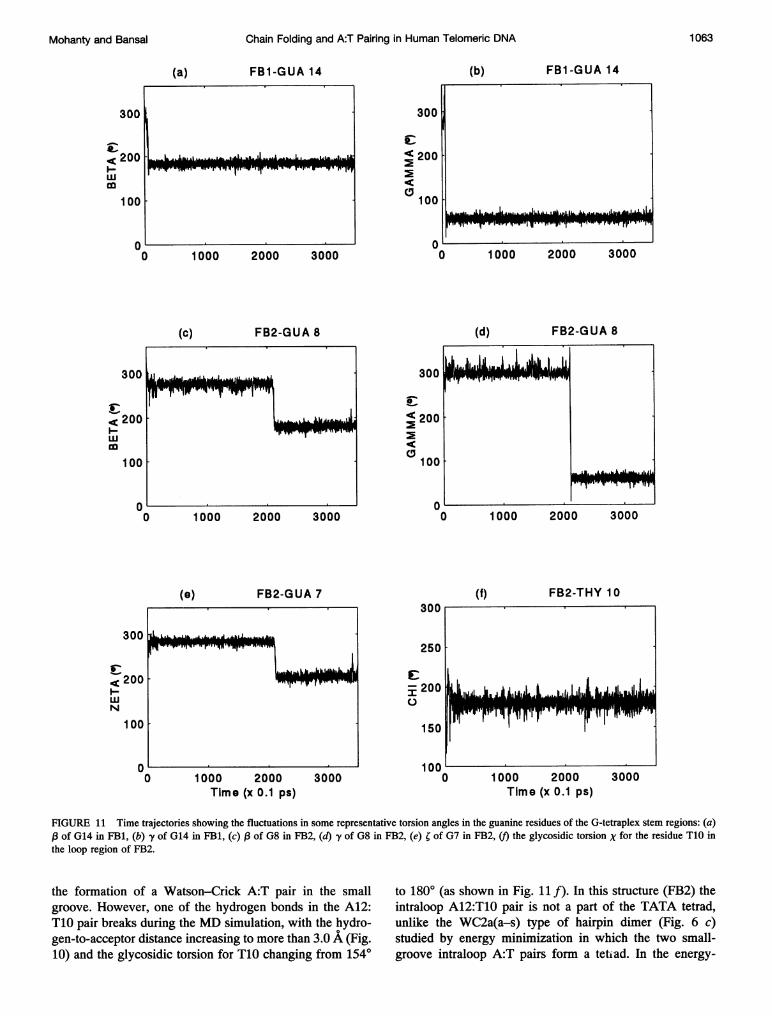
Torsion angle fluctuations in the MD structures
The backbone and glycosidic torsion angles for the twofolded-back structures, averaged over the last 300 ps of theMD simulation, are listed in Table 5, along with the stan-dard deviations and also the corresponding values in thestarting structure. As can be seen from Table 5, many of theguanines in the stem region of FB1 and FB2 do not undergo
small fluctuations about their starting values. The dihedraltransitions in some of the guanines take place in the first fewpicoseconds, and the dihedral angle values subsequentlystabilize. In structure FB1, while the guanines in the anticonformation have their backbone torsion angles close totheir starting values (Table 5), most of the guanines in thesyn conformation have undergone dihedral transitions in-volving a change in (,B, y) from (g-, g-) to (t, g+) withinthe first 10 ps. This dihedral transition for the syn guaninesis shown in Fig. 11 a and b for the nucleotide G14 in themiddle tetrad of FB1, as a representative case. On theotherhand, in structure FB2, a, ,B, and y of some of theguanines in the syn conformation changed from (g-, g-,g-) to (g+, t, t), whereas guanines in the anti conformationshowed a transition in ; from g- to t (Table 5). Only theguanines G7 and G8 of FB2 show sharp dihedral transitionsafter 200 ps of MD run, with a (g-, g-) to (t, g+) transitionin (,B, y) of G8, as seen in Fig. 11 c and d, accompanying a
g to t transition in ; of G7 (Fig. lle). Hence, in thesecases, averages of torsion angles over the period 50-350 ps
have very large values of standard deviations, and the av-
erage torsion angle values do not give a true indication ofthe dihedral angles. In the energy-minimized structures thesix backbone torsion angles for guanines, located one nu-
cleotide away from the loop, were g-, t, g+, t(1E), t, and g-with an anti glycosidic conformation and g-, g-, g t(2E),t, and g- for the guanines with syn glycosidic orientation.As mentioned above, after the MD simulation the six back-bone torsion angles in many of the syn guanines changed toa g-, t, g+, t(1E/2E), t, g- combination, which is known tobe stereochemically more favorable (Bansal and Sasisekha-ran, 1986). However, as can be seen from Table 5, in thestructures obtained from MD a few other unusual combina-tions of torsion angles are found, as also observed for theantiparallel G-tetraplex structures obtained from crystallog-raphy and NMR.
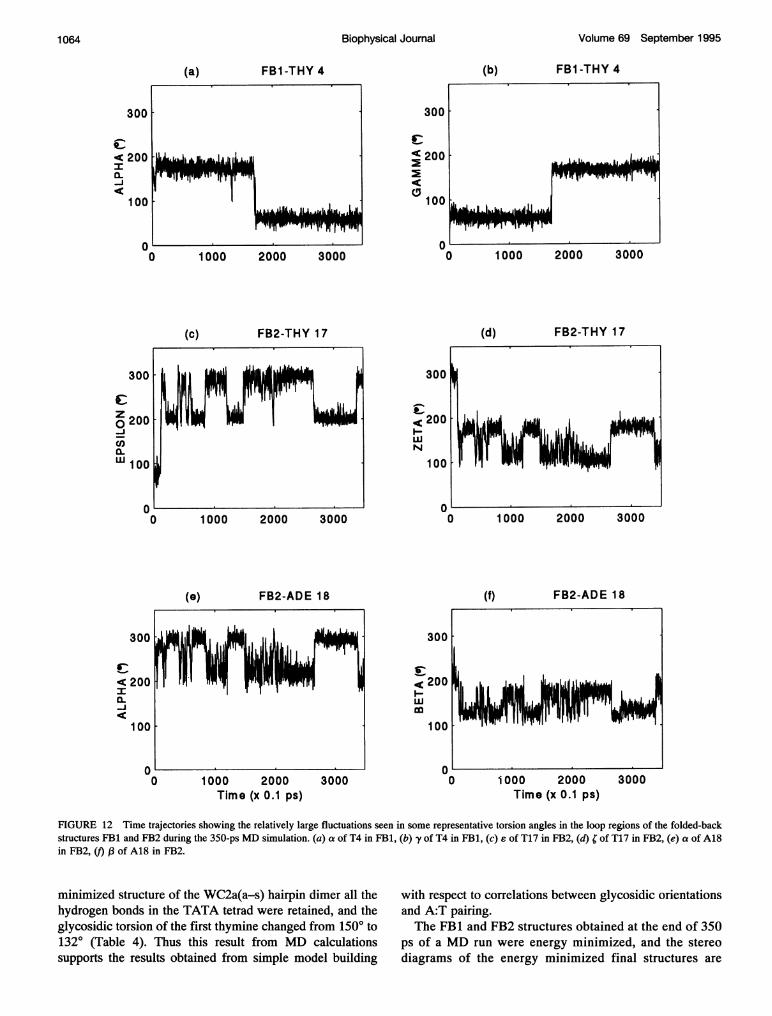
In contrast to the guanines in the stem, the backbonetorsion angles of some loop nucleotides show large fluctu-ations with significant dihedral transitions throughout the350-ps MD run. This is reflected in the larger values ofstandard deviations listed in Table 5. The transitions gen-
erally occur in a pairwise correlated manner for the back-bone torsion angles (a. y) and (£, 0, between the threestaggered conformations. The torsion angle trajectories fora and y in T4 of FB1, e and ; in T17, and a and 13 in A18of FB2 are shown in Fig. 12 as representative examples.Similar transitions are observed for several other loop residuesconstituting the TATA tetrads and TT pairs. It is interesting tonote that the loop nucleotides can undergo such large torsionangle fluctuations while retaining all the intraloop and inter-loop hydrogen bonds, as shown in Fig. 9 and 10.The glycosidic torsion angles of almost all the nucleo-
tides in the two folded-back structures remain very close tothe values obtained from the minimization studies, exceptfor T10 in the structure FB2. As discussed in the ModelBuilding section, this angle was constrained to have a value
any dihedral transition but throughout the simulation show
Mohanty and Bansal 1061
in the syn or low anti range because of t'fle requirements for
Biophysical Journal Volume 69 September 1995
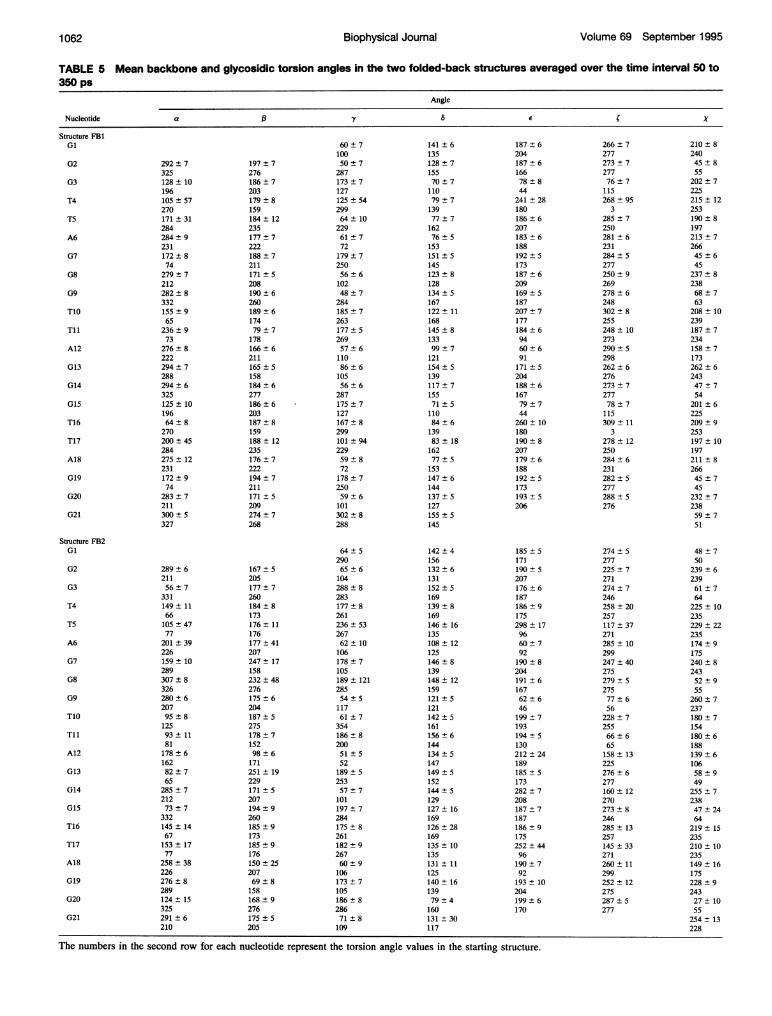
TABLE 5 Mean backbone and glycosidic torsion angles in the two folded-back structures averaged over the time interval 50 to350 ps
Angle
a j y 8 e x
292 7325128 10196105 ± 57270171 31284284 t 9231172 874
279 7212282 ± 8332155 ± 965
236 ± 973
276 ± 8222294 ± 7288294 ± 6325125 ± 1019664 ± 8
270200 ± 45284275 ± 12231172 ± 974
283 ± 7211300 ± 5327
289 621156 ± 7
331149 ± 1166105 4777
201 ± 39226159 ± 10289307 ± 8326280 ±620795 ± 812593 ± 1181178 ± 616282 ± 765
285 ± 721273 ± 7
332145 ± 1467153 ± 1777
258 ± 38226276 ± 8289124 ± 15325291 ± 6210
197 7276186 ± 7203179 ± 8159184± 12235177 7222188 7211171 5208190 ± 6260189 617479 ± 7178166 ± 6211165 5158184 6277186 ± 6203187 ± 8159188 ± 12235176 ± 7222194 ± 7211171 ± 5209274 ± 7268
167 5205177 ± 7260184 ± 8173176 ± 11176177 ± 41207247 ± 17158232 ± 48276175 ± 6204187 ± 5275178 ± 715298 ± 6171251 ± 19229171 ± 5207194 ± 9260185 9173185 ± 9176150 ± 2520769 ± 8158168 ± 9276175 ± 5205
60 ± 710050 ± 7287173 ± 7127125 ± 5429964± 10
22961 ± 772179 t 725056 ± 610248 ± 7
284185 t 7263177 + 526957 ± 611086 t 610556 ± 6287175 ± 7127167 ± 8299101 ± 9422959 + 872178 + 725059 + 6101302 ± 8288
64 + 529065 + 6104288 ± 8283177 + 8261236 ± 5326762 + 10106178 + 7105189 ± 12128554 + 511761 ± 7
354186 ± 820051 + 552189 + 525357 ± 7101197 ± 7284175 t 8261182 ± 926760 t 9106173 ± 7105186 t 828671 ± 8109
141 ± 6135128 ± 715570 t 711079 ± 713977 ± 716276 ± 5153151 ± 5145123 + 8128134 ± 5167122 ± 11168145 + 813399 ± 7121154 ± 5139117 + 715571 ± 511084 + 613983 ± 1816277 + 5153147 + 6144137 ± 5127155 ± 5145
142 ± 4156132 ± 6131152 ± 5169139 ± 8169146 ± 16135108 ± 12125146 ± 8139148 ± 12159121 + 5121142 ± 5161156 + 6144134 ± 5147149 ± 5152144 ± 5129127 + 16169126 ± 28169135 ± 10135131 ± 11125140 ± 1613979 ± 4160131 ± 30117
187 ± 6204187 + 616678 + 844
241 ± 28180186 ± 6207183 ± 6188192 ± 5173187 ± 6209169 ± 5187207 ± 7177184 ± 69460 ± 691171 ± 5204188 ± 616779 + 744260± 10180190 ± 8207179 ± 6188192 + 5173193 + 5206
185 ± 5171190 + 5207176 + 6187186 + 9175298 ± 179660 ± 792190 ± 8204191 ± 616762 4 646199 ± 7193194 ± 5130212 ± 24189185 - 5173282 ± 7208187 4 7187186 ± 91752524 4496190 4 792
193 ± 10204199 ± 6170
266 4 7277273 ± 727776 ± 7115268 ± 95
3285 ± 7250281 ± 6231284 ± 5277250 ± 9269278 ± 6248302 ± 8255248 ± 102732904 5298262 ± 6276273 ± 727778 ± 7115309 4 11
3278 4 12250284 4 6231282 ± 5277288 ± 5276
274 4 5277225 4 7271274 ± 7246258 ± 20257117 ± 37271285 ± 10299247 ± 40275279 ± 527577 4 656228 ± 725566 ± 665158 ± 13225276 ± 6277160 ± 12270273 ± 8246285 ± 13257145 ± 33271260 ± 11299252 ± 12275287 ± 5277
The numbers in the second row for each nucleotide represent the torsion angle values in the starting structure.
1062
Nucleotide
Structure FB1Gl
G2
G3
T4
T5
A6
G7
G8
G9
T10
Tll
A12
G13
G14
G15
T16
T17
A18
G19
G20
G21
Structure FB2Gl
G2
G3
T4
T5
A6
G7
G8
G9
T10
Tll
A12
G13
G14
G15
T16
T17
A18
G19
G20
G21
210 ± 824045 ± 855202 ± 7225215 ± 12253190 ± 8197213 4 726645 ± 645
237 ± 823868 ± 763
208 4 10239187 ± 7234158 4 7173262 ± 624347 4 754
201 ± 6225209 4 9253197 ± 10197211 ± 826645 ± 745
232 4 723859 ± 751
48 4 750
239 ± 623961 ± 764
225 4 10235229 ± 22235174 ± 91752404 824352 4 955
260 ± 7237180 ± 7154180 ± 6188139 ± 610658 ± 949
255 ± 723847 ± 2464
219 4 15235210 ± 10235149 ± 16175228 4 924327 ± 1055254± 13228
Chain Folding and A:T Pairing in Human Telomeric DNA
FB1-GUA 14 (b) FB1-GUA 14
(c)
300
c< 200
100 .
r
1000 2000 3000
FB2-GUA 8
O10 1000 2000 3000
(e)
300
< 200:
100
0
FB2-GUA 7
u0
(d)
1000 2000 3000
FB2-GUA 8
0 1000 2000 3000
(f) FB2-THY 10
0 1000 2000Time (x 0.1 ps)
3000 1000 2000Time (x 0.1 ps)
FIGURE 11 Time trajectories showing the fluctuations in some representative torsion angles in the guanine residues of the G-tetraplex stem regions: (a)l3 of G14 in FB1, (b) -y of G14 in FB1, (c) of G8 in FB2, (d) y of G8 in FB2, (e) ; of G7 in FB2, (f) the glycosidic torsion X for the residue T1O inthe loop region of FB2.
the formation of a Watson-Crick A:T pair in the smallgroove. However, one of the hydrogen bonds in the A12:T10 pair breaks during the MD simulation, with the hydro-gen-to-acceptor distance increasing to more than 3.0 A (Fig.10) and the glycosidic torsion for T10 changing from 1540
to 1800 (as shown in Fig. llf). In this structure (FB2) theintraloop A12:T10 pair is not a part of the TATA tetrad,unlike the WC2a(a-s) type of hairpin dimer (Fig. 6 c)studied by energy minimization in which the two small-groove intraloop A:T pairs form a tetiad. In the energy-
(a)
300
< 200w
100
0D
< 200w
300
£< 200wN
100
0
r-r-r,-TqMw-W"T'T"WTrr"W 'rprl w--,Tw iq
ILIAll ItTwvr Ntrfrgr VwTprr -w-iqw
.I.j
71
1063Mohanty and Bansal
Volume 69 September 1995
(a) FB1-THY 4 (b) FB1-THY 4
I-
)
0 1000 2000 3000
(c)
(e)
FB2-THY 17
FB2-ADE 18
0 1000 2000 3000
(d) FB2-THY 17
1000 2000 3000
(f) FB2-ADE 18
300 300
s-I~~~~~~~~~~~~~~~~~~~~.< 200 <200-J
100 100
0 100 1000 2000 3000 0 1000 2000 3000
Time (x 0.1 ps) Time (x 0.1 ps)
FIGURE 12 Time trajectories showing the relatively large fluctuations seen in some representative torsion angles in the loop regions of the folded-backstructures FB1 and FB2 during the 350-ps MD simulation. (a) a of T4 in FB1, (b) y of T4 in FB1, (c) £ of T17 in FB2, (d) ; of T17 in FB2, (e) a of A18in FB2, () ,(3 of A18 in FB2.
minimized structure of the WC2a(a-s) hairpin dimer all thehydrogen bonds in the TATA tetrad were retained, and theglycosidic torsion of the first thymine changed from 1500 to1320 (Table 4). Thus this result from MD calculationssupports the results obtained from simple model building
with respect to correlations between glycosidic orientationsand A:T pairing.The FB1 and FB2 structures obtained at the end of 350
ps of a MD run were energy minimized, and the stereodiagrams of the energy minimized final structures are
300
< 2001
0-bc
C0
v,)
cL
01Wi1
Biophysical Journal1 064
Chain Folding and A:T Pairing in Human Telomeric DNA
a
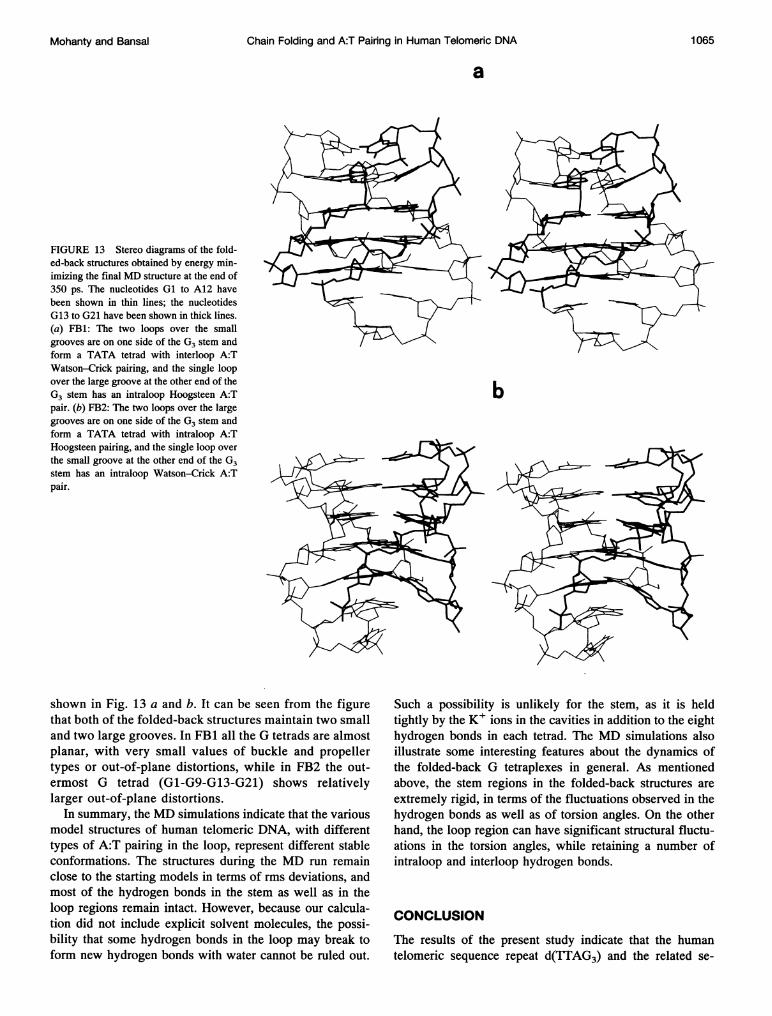
FIGURE 13 Stereo diagrams of the fold-ed-back structures obtained by energy min-imizing the final MD structure at the end of350 ps. The nucleotides Gl to A12 havebeen shown in thin lines; the nucleotidesG13 to G21 have been shown in thick lines.(a) FB1: The two loops over the smallgrooves are on one side of the G3 stem andform a TATA tetrad with interloop A:TWatson-Crick pairing, and the single loopover the large groove at the other end of theG3 stem has an intraloop Hoogsteen A:Tpair. (b) FB2: The two loops over the largegrooves are on one side of the G3 stem andform a TATA tetrad with intraloop A:THoogsteen pairing, and the single loop overthe small groove at the other end of the G3stem has an intraloop Watson-Crick A:Tpair.
b
shown in Fig. 13 a and b. It can be seen from the figurethat both of the folded-back structures maintain two smalland two large grooves. In FB1 all the G tetrads are almostplanar, with very small values of buckle and propellertypes or out-of-plane distortions, while in FB2 the out-ermost G tetrad (G1-G9-G13-G21) shows relativelylarger out-of-plane distortions.
In summary, the MD simulations indicate that the variousmodel structures of human telomeric DNA, with differenttypes of A:T pairing in the loop, represent different stableconformations. The structures during the MD run remainclose to the starting models in terms of rms deviations, andmost of the hydrogen bonds in the stem as well as in theloop regions remain intact. However, because our calcula-tion did not include explicit solvent molecules, the possi-bility that some hydrogen bonds in the loop may break toform new hydrogen bonds with water cannot be ruled out.
Such a possibility is unlikely for the stem, as it is heldtightly by the K+ ions in the cavities in addition to the eighthydrogen bonds in each tetrad. The MD simulations alsoillustrate some interesting features about the dynamics ofthe folded-back G tetraplexes in general. As mentionedabove, the stem regions in the folded-back structures areextremely rigid, in terms of the fluctuations observed in thehydrogen bonds as well as of torsion angles. On the otherhand, the loop region can have significant structural fluctu-ations in the torsion angles, while retaining a number ofintraloop and interloop hydrogen bonds.
CONCLUSION
The results of the present study indicate that the humantelomeric sequence repeat d(lTAG3) and the related se-
Mohanty and Bansal 1065

1066 Biophysical Journal Volume 69 September 1995
quence d(T3AG3) will prefer to adopt folded-back structureswith G3 stems and TTA or T3A loops rather than structureswith AGAG tetrads and T2 or T3 loops. The various G3tetraplex structures, although energetically they are verysimilar, have a wide range of structural variability becausethe presence of adenine along with thymines in the loopgives these sequence motifs the ability to adopt severaldifferent types of intraloop as well as interloop A:T pairs,leading to a number of different TATA tetrads when twoloops are on the same side of the G3-tetraplex stem. Therehave been relatively few studies of structural characteriza-tion of human telomeric DNA so far, and the variouspossible model structures presented here can help in ratio-nalizing the results of chemical probing studies of thesesequence motifs. The calculation of interproton distancesindicates that each of the different possible structures couldgive rise to unique NOE signals in NMR experiments. It isalso observed from model building that, although A:T pairswith Watson-Crick or Hoogsteen hydrogen bondingschemes can be accommodated in an antiparallel G tetra-plex, the thymine can have an anti conformation only whenthe A:T pair occurs in the large groove of the tetraplex,whereas an A:T pair in the small groove of the tetraplexforces the thymine to adopt a syn conformation. Thus bytaking into consideration the known propensity of thyminesto adopt an anti conformation, it might be possible to designguanine-rich oligonucleotides with selective positioning ofA and T, which would favor their folding into a particulartype of structure.The MD simulations on two of the folded-back model
structures indicate that these structures, with various possi-ble A:T pairs in the loop region, represent energeticallystable conformations for human telomeric DNA. The MDsimulations also indicate that the antiparallel G-tetraplexstem in these folded-back structures is very robust, and thenetwork of G:G hydrogen bonds, as well as the presence ofK+ ions between the G tetrads, makes it an extremely rigidand stable structure. Considerably higher flexibility is ex-hibited by the backbone of the loop nucleotides, eventhough all nucleotides in the loop are also involved inhydrogen bonding and most of these are retained during theMD simulations. The fact that folded-back telomeric struc-tures have a very rigid G-tetraplex stem and relativelyflexible loop regions has also been indicated by NMRstudies (Wang and Patel, 1992, Smith and Feigon, 1992,1993).
The authors are grateful to SERC, IISc, for computational facilities. Wealso thank Mr. Ravi Kiran for help in preparation of the manuscript. D.M.was supported by a fellowship from the Department of Biotechnology,India.
REFERENCES
Aboul-ela, F., A. I. H. Murchie, and D. M. Lilly. 1992. NMR study ofparallel-stranded DNA tetraplex formation by d(TG4T). Nature (Lon-don). 360:280-282.
Aboul-ela, F., A. I. H. Murchie, D. G. Norman, and D. M. J. Lilley. 1994.Solution structure of a parallel-stranded tetraplex formed by d(TG4T) inpresence of sodium ions by nuclear magnetic resonance spectroscopy.J. Moi. Bio. 243:458-471.
AMBER, V. 3.0. 1987. Molecular mechanics program from the Universityof California, San Francisco.
Arnott, S., R. Chandrasekharan, and C. M. Martilla. 1974. Structure ofpolyinosinic acid and polyguanylic acid. Biochem. J. 141:537-543.
Balagurumoorthy, P., and S. K. Brahmachari. 1994. Structure and stabilityof human telomeric sequence. J. Biol. Chem. 269:21858-21869.
Balagurumoorthy, P., S. K. Brahmachari, D. Mohanty, M. Bansal, and V.Sasisekharan. 1992. Hairpin and parallel quartet structure for telomericsequences. Nucl. Acids Res. 20:4061-4067.
Bansal, M., and D. Mohanty. 1993. Molecular mechanics studies of telo-meric DNA. J. Biomol. Struct. Dyn. 10:aO10.
Bansal, M., and N. Pattabiraman. 1989. Molecular mechanics studies onpoly(purine).poly(pyrimidine) sequences in DNA: polymorphism andlocal variability. Biopolymers. 28:531-548.
Bansal, M., and V. Sasisekharan. 1986. Molecular model-building ofDNA: constraints and restraints. In Theoretical Chemistry of BiologicalSystems. G. N. Szabo, editor. Elsevier, Amsterdam. 127-288.
Blackburn, E. H. 1991. Structure and function of telomeres. Nature (Lon-don). 350:569-573.
Blackburn, E. H. 1992. Telomerases. Ann. Rev. Biochem. 61:113-129.Blackburn, E. H., and J. W. Szostak. 1984. The molecular structure of
centromeres and telomeres. Ann. Rev. Biochem. 53:163-194.Bruskov, V. I., V. N. Bushuev, M. S. Okon, N. V. Shulyupina, and V. I.
Poltev. 1989. C-H ... 0 hydrogen bonding in solutions of methylatednucleic acid base analogs as revealed by NMR. Biopolymers. 28:589-604.
Cheng, Y. K., and B. M. Pettitt. 1992. Hoogsteen versus reversed-Hoogsteen base pairing: DNA triple helices. J. Am. Chem. Soc. 114:4465-4474.
Cheong, C., and P. B. Moore. 1992. Solution structure of an unusuallystable RNA tetraplex containing G and U-quartet structures. Biochem-istry. 31:8406-8414.
Chernyi, A. A., Yu P. Lysov, I. A. Il'ychova, A. S. Zibrov, A. K.Shchyolkina, 0. F. Borisova, 0. K. Mamaeva, and V. L. Florentiev.1990. Four-stranded DNA helices: conformational analysis of regularpoly(dT).poly(dA).poly(dA).poly(dT) helices with various types of basebinding. J. Biomol. Struct. Dyn. 8:513-527.
Ferentz, A. E., J. Wiorkiewicz-Kuczera, M. Karplus, and G. L. Verdine.1993. Molecular dynamics simulation of disulfide cross-linked DNAdecamers. J. Am. Chem. Soc. 115:7569-7583.
Frank-Kamenetskii, M. 1992. The quadruplex and the vase. Nature (Lon-don). 356:105-105.
Guo, Q., M. Lu, and N. R. Kallenbach. 1992a. Effect of thymine tractlength on the structure and stability of model telomeric sequences.Biochemistry. 32:3596-3603.
Guo, Q., M. Lu, and N. R. Kallenbach. 1992b. Adenine affects the structureand stability of telomeric sequences. J. Biol. Chem. 267:15293-15300.
Gupta, G., A. E. Garcia, Q. Guo, M. Lu, and N. R. Kallenbach. 1993.Structure of a parallel-stranded tetramer of the Oxytricha telomere repeatoligonucleotide d(T4G4). Biochemistry. 32:7098-7103.
Guschlbauer, W., J. F. Chantot, and D. Thiele. 1990. Four-stranded nucleicacid structures 25 years later: from guanosine gels to telomer DNA.J. Biomol. Struct. Dyn. 8:491-511.
Henderson, E., C. C. Hardin, S. K. Walk, I. Tinoco, Jr., and E. H.Blackburn. 1987. Telomeric DNA oligonucleotides form novel intramo-lecular structures containing guanine-guanine base pairs. Cell. 51:899-908.
Henderson, E. R., and E. H. Blackburn. 1989. An overhanging 3' terminusis a conserved feature of telomeres. Mol. Cell. Biol. 9:345-348.
Jin, R., K. J. Breslauer, R. A. Jones, and B. L. Gaffney. 1990. Tetraplexformation of a guanine-containing nonameric DNA fragment. Science.250:543-546.
Kang, C., X. Zhang, R. Ratliff, R. Moyzis, and A. Rich. 1992. Crystalstructure of four-stranded Oxytricha telomeric DNA. Nature (London).356:126-131.

Mohanty and Bansal Chain Folding and A:T Pairing in Human Telomeric DNA 1067
Laughlan, G., A. I. H. Murchie, D. G. Norman, M. H. Moore, P. C. E.Moody, D. M. J. Lilley, and B. Luisi. 1994. The high resolution crystalstructure of a parallel-stranded guanine tetraplex. Science. 265:520-524.
Macaya, R. F., P. Schultze, F. W. Smith, J. A. Roe, and J. Feigon. 1993.Thrombin-binding DNA aptamer forms a unimolecular quadruplexstructure in solution. Proc. Natl. Acad. Sci. USA. 90:3745-3749.
Miura, T., and G. J. Thomas, Jr. 1994. Structural polymorphism of telo-meric DNA: interquadruplex and duplex- quadruplex conversionsprobed by Raman spectroscopy. Biochemistry. 33:7848-7856.
Mohanty, D., and M. Bansal. 1993. Conformational polymorphism inG-tetraplex structures: strand reversal by base flipover or sugar flipover.Nucl. Acids Res. 21:1767-1774.
Mohanty, D., and M. Bansal. 1994. Conformational polymorphism intelomeric structures: loop orientation and interloop pairing ind(G4TJG4). Biopolymers. 34:1187-1211.
Moyzis, R. K., J. M. Buckingham, L. S. Cram, M. Dani, L. L. Deaven, M.D. Jones, J. Meyne, R. L. Ratliff, and J.-R. Wu. 1988. A highlyconserved repetitive DNA sequence, (TTAGGG)., present at the telo-meres of human chromosomes. Proc. Natl. Acad. Sci. USA. 85:6622-6626.
Padmanabhan, K., K. P. Padmanabhan, J. D. Ferrara, J. E. Sadler, and A.Tulinsky. 1993. The structure of a-thrombin inhibited by a l5mer singlestranded DNA aptamer. J. Biol. Chem. 268:17651-17654.
Panyutin, I. G., 0. I. Kovalsky, E. I. Budowsky, R. E. Dickerson, M. E.Rikhirev, and A. A. Lipanov. 1990. G-DNA: A twice folded DNAstructure adopted by single-stranded oligo(dG) and its implications fortelomeres. Proc. Natl. Acad. Sci. USA. 87:867-870.
Pearson, A. M., A. Rich, and M. Krieger. 1993. Polynucleotide binding tomacrophage scavanger receptors depends on the formation of base-quartet-stabilized four stranded helixes. J. Biol. Chem. 268:3546-3554.
Pinnavaia, T. J., H. T. Miles, and E. D. Becker. 1975. Self-assembled5'-guanosine monophosphate. Nuclear magnetic resonance evidence fora regular, ordered structure and slow chemical exchange. J. Am. Chem.Soc. 97:7198-7200.
Rose, I. A., K. R. Hanson, K. D. Wilkinson, and M. J. Wimmer. 1980. Asuggestion for naming faces of ring compounds. Proc. Natl. Acad. Sci.USA. 77:2439-2441.
Ross, W. S., and C. C. Hardin. 1994. Ion-induced stabilization of theG-DNA quadruplex: free energy perturbation studies. J. Am. Chem. Soc.116:6070-6080.
Sasisekharan, V., S. B. Zimmermann, and D. R. Davies. 1975. The struc-ture of helical 5'-guanosine monophosphate. J. Mol. Biol. 92:171-179.
Scaria, P. V., S. J. Shire, and R. H. Shafer. 1992. Quadruplex structure ofd(G3T4G3) stabilized by K' or Na+ is an asymmetric hairpin dimer.Proc. Natl. Acad. Sci. USA. 89:10336-10340.
Schultze, P., R. F. Macaya, and J. Feigon. 1994. Three-dimensional solu-tion structure of the thrombin-binding DNA aptamer d(GGTTGGTGT-GGTTGG). J. Mol. Biol. 235:1532-1547.
Sen, D., and W. Gilbert. 1988. Formation of parallel four-stranded com-plexes by guanine-rich motifs for meiosis. Nature (London). 334:364-366.
Sen, D., and W. Gilbert. 1990. A sodium-potassium switch in the formationof four stranded G4-DNA. Nature (London). 344:410-414.
Smith, F. W., and J. Feigon. 1992. Quadruplex structure of Oxytrichatelomeric DNA oligonucleotides. Nature (London). 356:164-168.
Smith, F. W., and J. Feigon. 1993. Strand orientation in the DNA quadru-plex formed from the Oxytricha telomere repeat oligonucleotided(G4T4G4) in solution. Biochemistry. 32:8682-8692.
Sundquist, W. I., and S. Heaphy. 1993. Evidence for interstrand quadruplexformation in the dimerization of human immunodeficiency virus Igenomic RNA. Proc. Natl. Acad. Sci. USA. 90:3393-3397.
Sundquist, W. I., and A. Klug. 1989. Telomeric DNA dimerises by for-mation of guanine tetrads between hairpin loops. Nature (London).342:825-829.
Taylor, R., and 0. Kennard. 1982. Crystallographic evidence for theexistence of C-HO..., C-H...N and C-H...Cl hydrogen bonds. J. Am.Chem. Soc. 104:5063-5070.
Tidor, B., K. K. Irikura, B. R. Brooks, and M. Karplus. 1983. Dynamics ofDNA oligomers. J. Biomol. Struct. Dyn. 1:231-252.
Walsh, K., and A. Gualberto. 1992. MyoD binds to the guanine tetradnucleic acid structure. J. Biol. Chem. 267:13714-13718.
Wang, K. Y., S. McCurdy, R. G. Shea, S. Swaminathanan, and P. H.Bolton. 1993. A DNA aptamer which binds to and inhibits thrombinexhibits a new structural motif for DNA. Biochemistry. 32:1899-1904.
Wang, K. Y., S. Swaminathan, and P. H. Bolton. 1994. Tertiary structuremotif of Oxytricha telomere DNA. Biochemistry. 33:7517-7527.
Wang, Y., and D. J. Patel. 1992. Guanine residues in d(T2AG3) andd(T2G4) form parallel-stranded potassium cation stabilized G-quadruplexes with anti glycosidic torsion angles in solution. Biochem-istry. 31:8112-8119.
Wang, Y., and D. J. Patel. 1993. Solution structure of human telomericrepeat d[AG3(T2AG3)3] G-tetraplex. Structure. 1:263-282.
Weiner, S. J., P. A. Kollman, D. A. Case, U. C. Singh, C. Ghio, G.Alagona, S. Profeta, Jr., and P. Weiner. 1984. A new force field formolecular mechanical simulation of nucleic acids and proteins. J. Am.Chem. Soc. 106:765-784.
Weiner, S. J., P. A. Kollman, D. T. Nguyen, and D. A. Case. 1986. An allatom force field for simulation of proteins and nucleic acids. J. Comp.Chem. 7:230-252.
Williamson, J. R. 1994. G-quartet structures in telomeric DNA. Annu. Rev.Biophys. Biomol. Struct. 23:703-730.
Williamson, J. R., M. K. Raghuraman, and T. R. Cech. 1989. Monovalentcation-induced structure of telomeric DNA: the G-quartet model. Cell.59:871-880.
Wyatt, J. R., T. A. Vickers, J. L. Robertson, R. W. J. Buckheit, T. Klimkait,E. DeBaets, P. W. Davis, B. Rayner, J. L. Imbach, and D. J. Ecker. 1994.Combinatorially selected guanosine-quartet structure is a potent inhibi-tor of human immuno deficiency virus envelope-mediated cell fusion.Proc. Natl. Acad. Sci. USA. 91:1356-1360.
Zhurkin, V. B., G. Raghunathan, N. B. Ulyanov, R. D. Camerini-Otero,and R. L. Jernigan. 1994. A parallel DNA triplex as a model forhomologous recombination. J. Mol. Biol. 239:181-200.
Zimmermann, S. B., G. H. Cohen, and D. R. Davies. 1975. X-ray fiberdiffraction and model-building study of polyguanylic acid and poly-inosinic acid. J. Mol. Biol. 92:181-192.