Embed Size (px)
Citation preview

1
Fire Intensity Effects on Coastal Redwood Recruitment
Axel Eaton, Mark Goering, Quinton Mells, Catherine Ohlin, Elliott Perry, and Ariana Rojas Introduction
Fire is a powerful disturbance that has been shaping Earth's ecosystems since land was colonized by terrestrial plants. In the following eons, fire has moderated Earth's atmospheric composition and become a key component in nutrient cycling and ecological succession. The overall effects of fire have heavily influenced the global distribution of vegetation and shaped the ecosystem structures we see today (Bowman et al. 2009). To prepare for the role of fire in a warmer and uncertain future, better understanding of how fire shapes ecosystem is imperative.
California landscapes have been shaped by a long history of persistent wildfire activity. Frequent lightning-ignited fires have burned throughout the state, resulting in many specially adapted plant communities. We have come to understand the importance of fire in these specially adapted ecosystems as fire cycles nutrients and creates space for new growth. Wildfires produce forest canopy gaps, increase understory light, and remove surface litter which exposes high mineral concentration soil (Meyer and Safford 2011). In a study done by Meyer and Safford (2011), canopy gaps exceeding 0.3 - 0.8 ha demonstrated the highest number of saplings. These large gaps allow more light to reach the base of the tree where the sapling is positioned to successfully prosper with newly available forest floor space (Ramage et al. 2010). Fire also returns nutrients back into the soil in the form of ashes. Phosphorus and other key elements, which were previously bound in plant tissues, are released back into the soil. However, since California settlement, fire suppression efforts have drastically altered fire regimes. Fire suppression has resulted in understory litter accumulation, reduced nutrient cycling, and a dense, homogeneous forest structure. Contrary to previous thought, the exclusion of fire has created an ecosystem prone to high-severity, stand replacing fires.
Specific effects on ecosystems can be both positive and negative. Although the presence of wildfires may increase the abundance of nonnative plant species in pine forests, many species rely on the occurrence of fire to regenerate and expand their population (Keeley 2002). For example, fire is the single most important factor in maintaining lodgepole pine (Pinus Contorta) forests (Brown 1975). By burning old growth trees and other plants, fires replenish the soil with carbon and other nutrients for new growth seedlings to use (Webster and Halpern 2010). Moderate to intense surface fires create a bed of soft, ashy, nutrient rich soil where conifer seeds

2
can become buried and are more likely to germinate. More severe fires in mixed conifer forests actually lead to an increase in species diversity and total plant cover (Webster and Halpern 2010). Subsequently, conifers are dependent on recurring fires for their own regeneration. This is especially true for the coastal redwood (Sequoia semperviren).
Redwoods are an iconic, endemic species found only along a narrow 900 km strip of coastline ranging from central California to southern Oregon which represents only 4 to 5 percent of their historical range (Noss 2000). They face stressors such the lack of moisture availability and recent disturbance of changing fire regimes causing a cataclysmic shrink in their range. Despite being affected by these stressors, redwoods can still flourish in severe fires. Thick insulated bark and a high crown have allowed redwoods to outcompete all other associated species in fire-disturbed ecosystems (Lazzeri-Aerts and Russell 2014).
Wildfires have been observed to be a crucial part of the redwood reproductive cycle (Ramage et al. 2010). A sister species, Sierra redwoods (Seqoiadendron giganteum), have developed serotinous cones, which only release their seeds upon the occurrence of fire. The coastal redwood species do not have serotinous cones, and therefore rely heavily on basal sprouting. Sprouting is used as insurance against seedling failure. According to Lazzeri-Aerts (2014) up to 81% of redwoods originate from basal sprouts. Sequoia sempervirens are one of the few conifers with the ability to sprout from their roots—sprouting being initiated from underground burls, known as lignotubers (Ramage et al. 2010). In response to a variety of life-threatening disturbances, redwoods will divert resources into clonal regeneration in an attempt to persist despite potential bole death. This trait has contributed to redwoods’ pliancy to anthropogenic and natural disturbances, such as frequent fire exposure. In the short term, species with the highest amount of post-fire basal sprout growth will become the most prominent. Redwoods outcompeted post-fire sprouting abilities of other broad-leafed trees with similar recruitment strategies (Lazzeri-Aerts and Russell 2014). Many studies have confirmed that wildfires are crucial for redwood recruitment and increase their overall dominance compared to other species with less fire adaptive qualities. (Lazzeri-Aerts and Russell 2014).
In this study, we take a closer look at the understudied response to fire severity by coastal redwoods in the southernmost part of their range. Specifically, we are studying how basal sprouting as a post-fire response correlates to fire severity. Based on the existing literature, we predicted that coastal redwoods would have a positive basal sprout response to fires of higher severity. This study will contribute to more informed management decisions in an effort to most efficiently and effectively conserve redwoods in the face of climate change and shifting fire regimes.

3
Methods Natural History of the Study Site
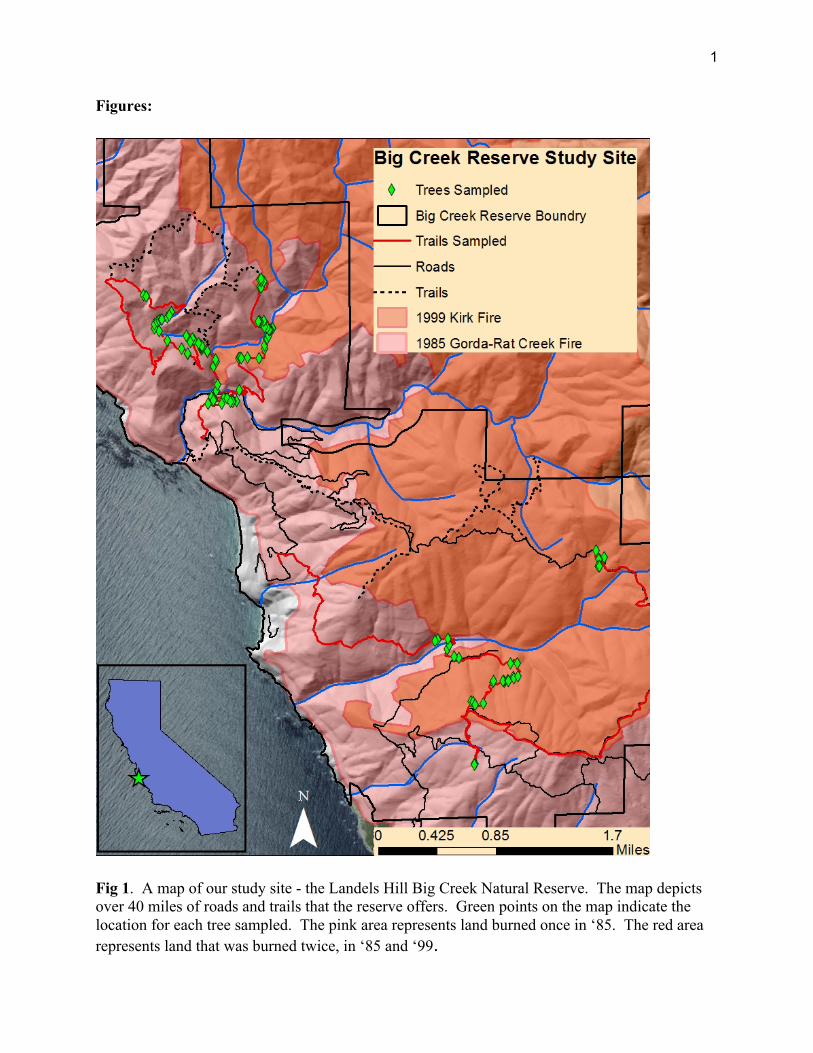
Research was conducted at the Landels-Hill Big Creek Natural Reserve (W 36.071984°, N -121.599245°). Located in Monterey County, California, the reserve is managed by UC Santa Cruz and encompasses approximately 3,200 hectares of land along the Big Sur Coast. It hosts a diverse array of habitats including rugged shore-line, dense redwood forests, coastal chaparral, and rolling grass hills. The communities are supported by annual precipitation ranging from 100 cm at the coast to 210 cm at the higher elevations of up to 1,220 meters. The reserves landscape has also been heavily impacted by fire, completely burning in 1985 during the Rat-Creek Gorda Fire and partially burning in the Kirk Complex in 1999. The many diverse communities are accessible through 40+ miles of roads and trails; Figure 1 depicts those covered in our study and the extent of the fires. Research Design
We visited every coastal redwood tree (Sequoia sempervirens) accessible or visible from the hiking trails to measure burn severity, clonal regeneration, and various site characteristics. Redwoods grow in a ‘cathedral’ formation, where mature trees sprout basal stems from the central trunk that then grow to surround the ‘parent’. We targeted only obvious ‘parent’ trees (meaning the central, largest tree acknowledging they’re all biologically the same individual) Any time it was not immediately obvious which parent tree a group of clonal stems originated from, we did not sample those individuals.
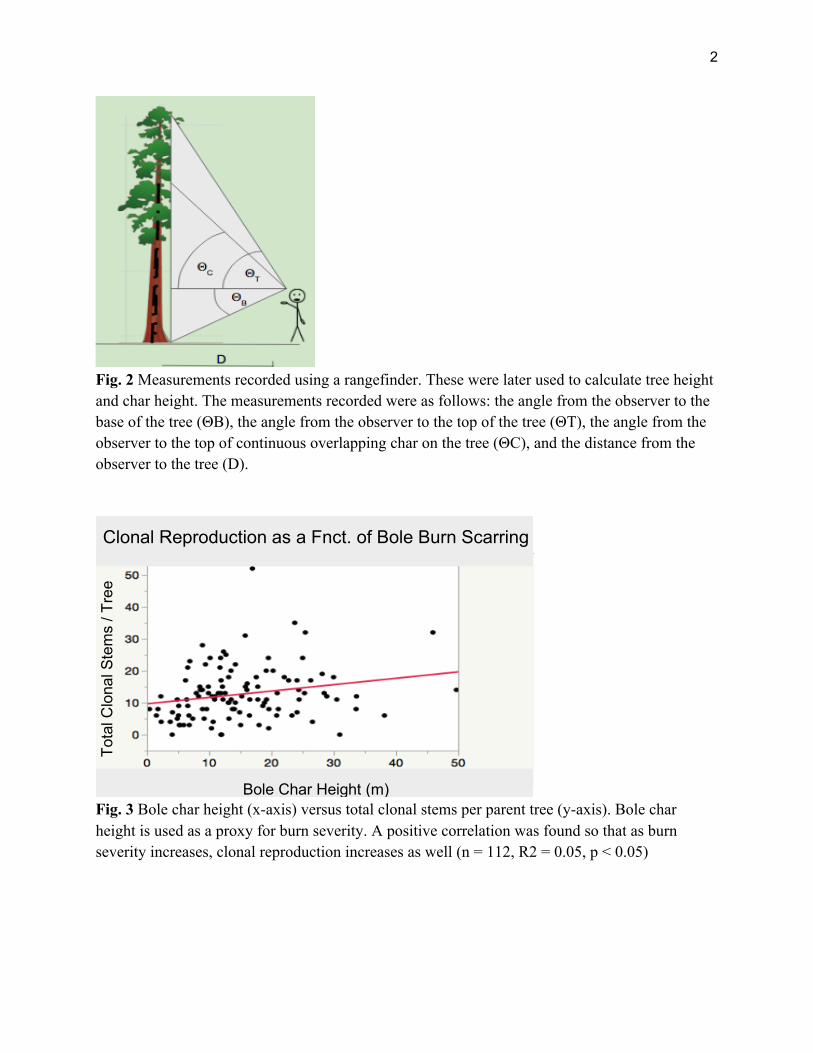
To estimate the size of the parent tree, we used a diameter at breast height (dbh) tape to measure the tree’s diameter. We also calculated tree height using a laser rangefinder to measure the distance to the tree, the angle to the top of the tree, and the angle to the base of the tree (see Figure 2). Because our measurements for dbh were much more precise and accurate than those for height, we chose to focus on dbh as our main proxy for tree size when analyzing our data.
We estimated burn severity according to multiple metrics. We devised an ordinal and a continuous ranking to estimate burn damage for each ‘parent’ tree. The least severe scarring did not compromise the tree’s fire-resistant bark. The next most severe burn is heartwood exposure, where fire was able to burn through the outer bark layer, exposing the tree’s inner, more sensitive wood. The most severe burn scar is a goosepen, which is a cavity formed when a fire burns through the bark and continues to burn towards the center of the tree through a significant portion of the heartwood. Continuous bole char height was also calculated using the same method as tree height.
To quantify regenerative outputs of parent trees, we counted the number of living and dead clonal stems (new growth stemming from parent tree base or root network) around each parent tree, and then classified stems into one of five categories based on size. Our clonal stem

4
categories were as follows: 1- less than 6 feet tall with woody tissue, 2- taller than 6 feet with a dbh of less than 1 inch, 3- a dbh between 1 and 4 inches, 4- a dbh between 4 and 8 inches, and 5- a dbh greater than 8 inches. We also noted presence or absence of a sprout mat, which we defined as a cluster of green, non-woody sprouts less than 3 feet tall. We did not include dead saplings less than 6 feet tall in our counts. We chose to combine live and dead sapling counts when analyzing our data for a more accurate understanding of the parent tree’s total regenerative output. Statistical Analysis
We used statistical software JMP 12 to analyze our data. Each clonal stem size category, and total clonal stem counts for parent trees, were tested individually against the presence of a goosepen, heartwood exposure, or intact bark using one-way ANOVAs. We used linear regressions to investigate the relationship between total clonal stems and tree size—using dbh, tree height, and tree volume as proxies. A linear regression was also fit to total saplings as a function of bole char height. We used a linear regression to test the relationship between tree size and burn severity. Finally, we ran an analysis of covariance test (ANCOVA) on burn severity to account for confounding variables, using our three ordinal rankings as a proxies for burn severity, dbh as a proxy for tree size, and total clonal stem production. Results
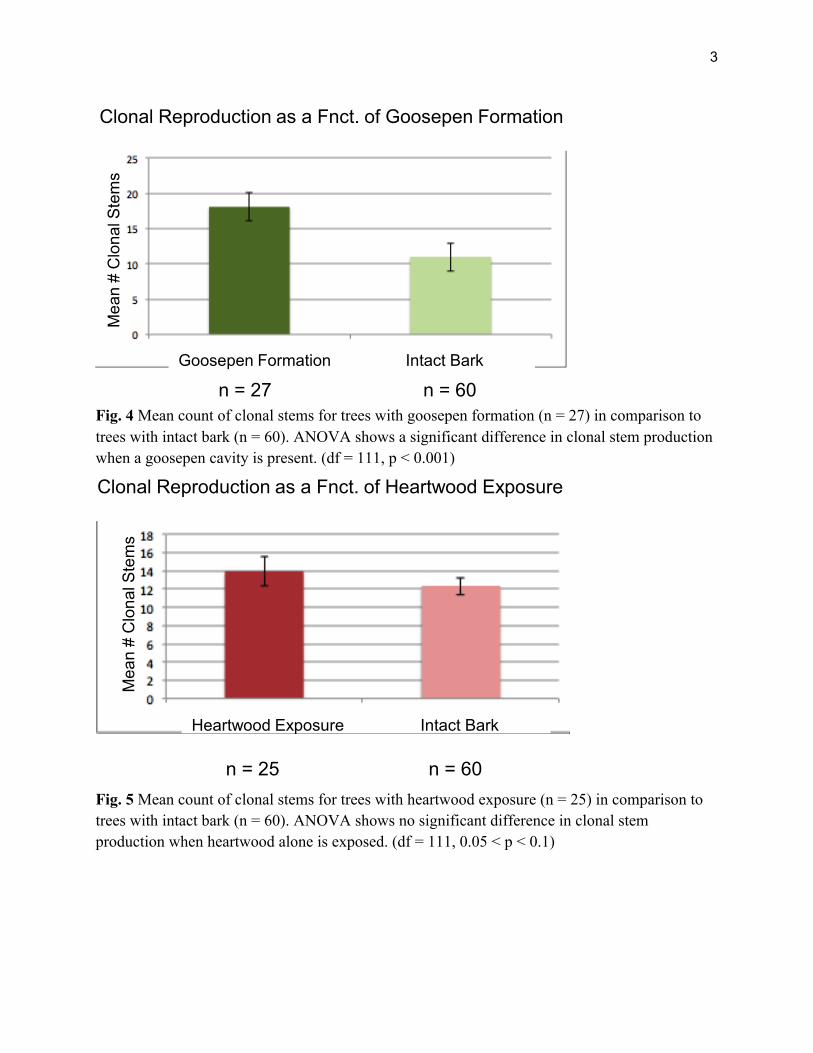
We accessed 112 ‘parent’ redwood trees, with all trees showing some amount of char on their trunks. Of all trees sampled 60 had intact bark, 25 had heartwood exposure, and 27 had goosepen formation. We found positive relationships between regenerative output and burn severity according to two of our three burn severity metrics. Bole char height is positively related to clonal reproduction (n = 112, R2 = 0.05, p < 0.05; Figure 3). With heartwood exposure alone, there is no significant relationship between burn severity and clonal reproduction (df = 111, F = .83, 0.05 p > 0.1; Figure 4). However, when burn severity is intense enough to form a goosepen, clonal reproduction significantly increases (df = 111 F= 16.89, p < 0.0001; Figure 5).
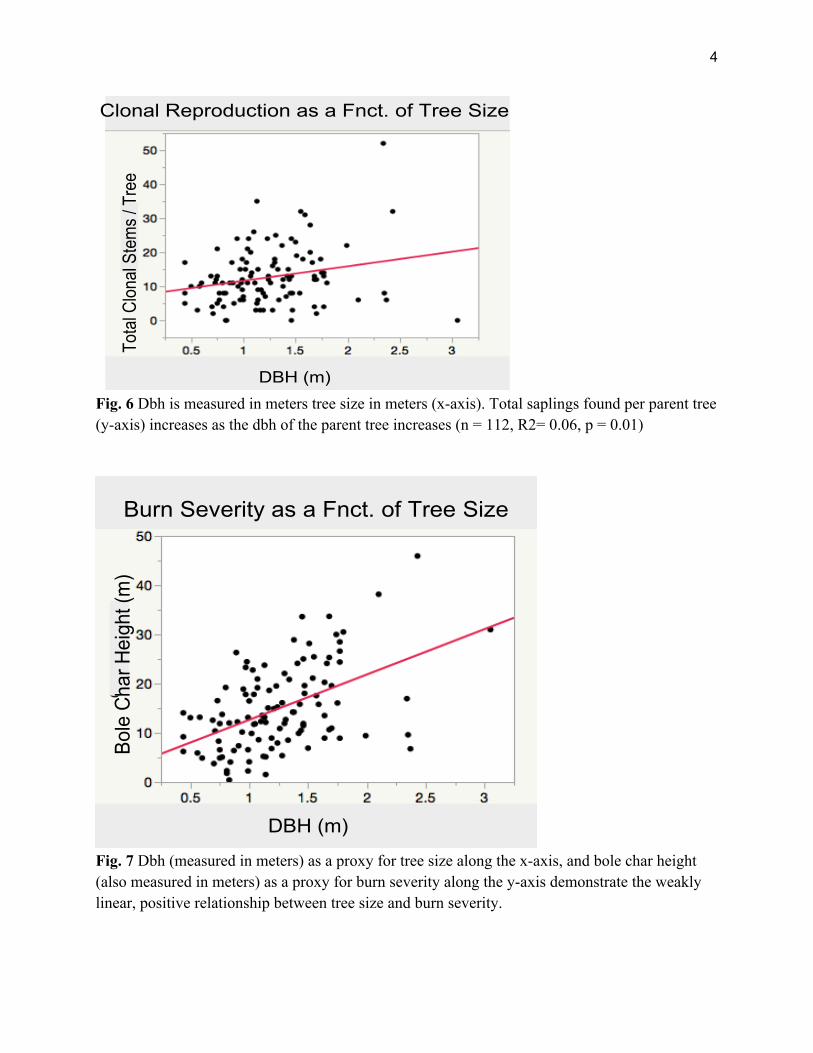
In addition to burn severity, our data shows a weakly linear, positive correlation between
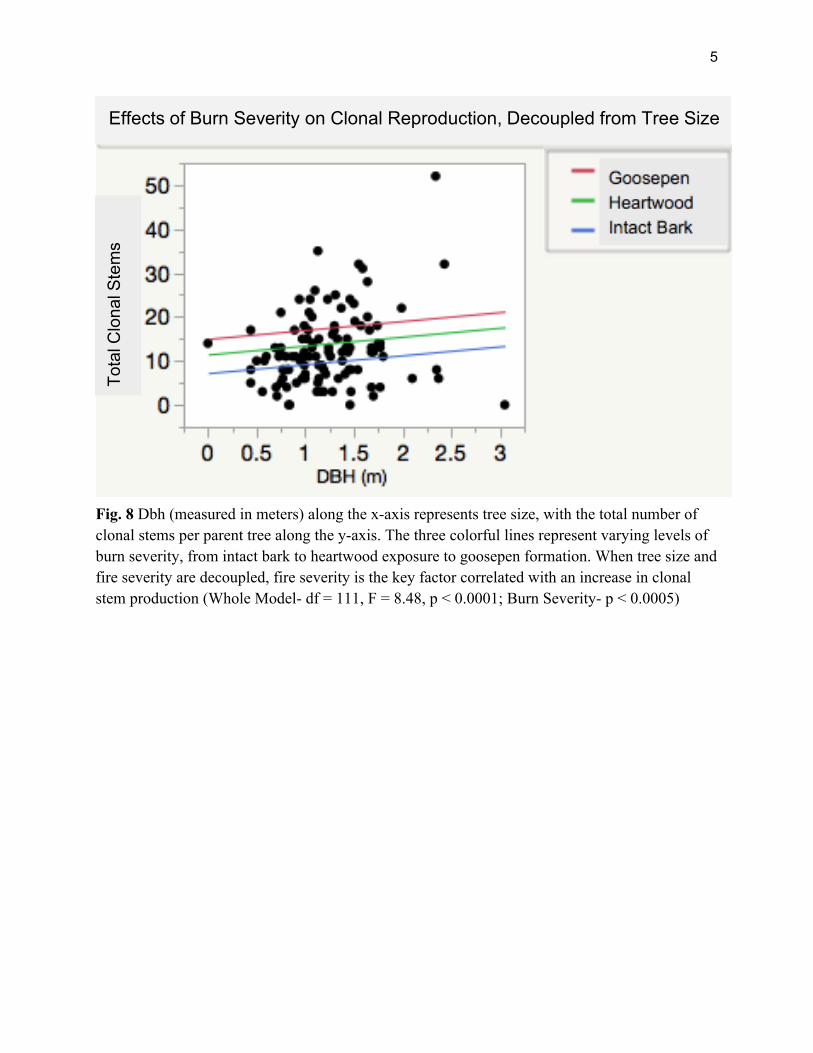
clonal regeneration and tree size measured as DBH. (n = 112, R2= 0.06, p = 0.01; Figure 6). Tree size and burn severity, as measured by dbh and bole char height, also show a positive correlation (n = 112, R2 = 0.25, p < 0.0001; Figure 7). When dbh covariation is taken into account, we find that clonal reproduction mainly increases in correlation with increasing burn severity. (ANCOVA Whole Model- df = 111, F = 8.48, p < 0.0001; Burn Severity- t = 3.18, p < 0.005; dbh - t = 1.27, p > 0.05; Figure 8).

5
Discussion Burn severity showed a strong influence on redwood regeneration in our Big Creek sample. The number of clonal trees surrounding a ‘parent’ tree was positively related to both our ordinal metric of burn severity and our linear metric of burn severity, each based on completely different features from the trees. Trees with higher char marks on their bark produced more clones. And trees with more severe indicators of burn damage produced more clones. Even when the positive influence of tree size on overall clone production is accounted for (as larger trees also showed higher degrees of burn overall), the positive relationship between burn damage and clone production is strong. We have come up with two possible driving factors for this regenerative response. The first being a response to light availability. The formation of a large cavity at the base of a tree occurs as a result of a high intensity burn which also clears out the understory around the parent tree. Because of increases of light availability, clonal stems have the opportunity to flourish. (Lorimer et al. 2009). Thus, the trait that prioritizes post-fire clonal regeneration survive is selected.
Another possible driving factor of the regenerative response we are seeing involves the redirection of nutrients. The vascular cambium, along with the xylem and phloem of a ‘parent’ redwood tree, are severely wounded in the formation of a goosepen. Because the regulation of plant growth is driven by the transport of nutrients and hormones through these systems, a compromise in their ability to do so may direct more nutrients to the clonal stems (Finney 1975). Additional methods could be brought to bear on this investigation to strengthen and build on the inferences we have drawn. Measuring the size of a goosepen in relation to tree size could provide a more accurate representation of burn severity. This data could also shed light on whether increased clonal regeneration is a result of decreased parent tree vascular tissue or a function of light availability. By sampling more redwood groves across the California coast we can become certain that this pattern is not unique to Big Creek Reserve. By more rigorously distinguishing clonally regenerated trees as opposed to saplings generated by seed via DNA sequencing, our data would be more representative of true clonal regeneration. Despite these potential improvements to our study, this does not diminish the efficacy of our study. Implications for Forest Management
Current fire suppression tactics are to put fires out as quickly as possible, which leads to a buildup of ladder fuels in the understory. Ultimately, fuel buildup increases the likelihood of high severity, stand replacing fires that would completely devastate a coastal redwood forest instead of stimulate new growth. Our findings suggest new potential approaches to fire suppression in redwood forests. Such approaches could include the thinning of understory ladder fuels at the base of old growth redwoods. This could be easily accomplished by incentivizing

6
private logging companies to remove ladder fuel while allowing them to take a portion of the usable lumber in the stand as an incentive. Once ladder fuels have been removed fire can now burn through the area with less threat to old growth redwood canopies. When fire is allowed back into the system, the clonal regenerative response of the remaining redwoods would be stimulated, potentially increasing redwood recruitment overall.
Additionally, wildland firefighting tactics could be slightly altered in order to allow for areas with redwoods stands to be burned in the event of a large wildland fire. This could be accomplished by constructing fire retarding control lines in such a way as to allow these areas to burn instead of excluding fires from these old-growth redwood stands. We fully recognize these methods may be increasing fire risk. This recommendation is not intended to compromise safety by any extent by prioritizing redwood regrowth over fire suppression. Our findings could maintain and potentially increase fire safety, while simultaneously benefiting the natural ecology of coastal redwood forests.
Research has not only demonstrated that redwoods can tolerate fire, but that they can directly benefit from fire exposure. A better understanding of how this evocative tree species responds to harsh environmental effects, like fire damage, will enhance our ability to allow populations of coastal redwoods to flourish in an uncertain future climate. The coastal redwoods’ diminished range will potentially face more pressure as climatic shifts in temperature reduce coastal fog, and in turn increase drought stress on redwoods (Johnson and Dawson 2010). Because these climatic factors are predicted to increase the severity and frequency of fires, forestry management in line with our current research can transform the threat of fires into an opportunity to conserve redwood communities. By leveraging the recruitment effects that fires have on individual redwood trees, fires can be a possible natural strategy to conserve a species that is currently at risk.
1
Figures:
Fig 1. A map of our study site - the Landels Hill Big Creek Natural Reserve. The map depicts over 40 miles of roads and trails that the reserve offers. Green points on the map indicate the location for each tree sampled. The pink area represents land burned once in ‘85. The red area represents land that was burned twice, in ‘85 and ‘99.
2
Fig. 2 Measurements recorded using a rangefinder. These were later used to calculate tree height and char height. The measurements recorded were as follows: the angle from the observer to the base of the tree (ΘB), the angle from the observer to the top of the tree (ΘT), the angle from the observer to the top of continuous overlapping char on the tree (ΘC), and the distance from the observer to the tree (D).
Fig. 3 Bole char height (x-axis) versus total clonal stems per parent tree (y-axis). Bole char height is used as a proxy for burn severity. A positive correlation was found so that as burn severity increases, clonal reproduction increases as well (n = 112, R2 = 0.05, p < 0.05)
Bole Char Height (m)
Clonal Reproduction as a Fnct. of Bole Burn Scarring
Tota
l Clo
nal S
tem
s / T
ree
3
Fig. 4 Mean count of clonal stems for trees with goosepen formation (n = 27) in comparison to trees with intact bark (n = 60). ANOVA shows a significant difference in clonal stem production when a goosepen cavity is present. (df = 111, p < 0.001)
Fig. 5 Mean count of clonal stems for trees with heartwood exposure (n = 25) in comparison to trees with intact bark (n = 60). ANOVA shows no significant difference in clonal stem production when heartwood alone is exposed. (df = 111, 0.05 < p < 0.1)
Mea
n #
Clo
nal S
tem
s
n = 27 n = 60Goosepen Formation Intact Bark
Clonal Reproduction as a Fnct. of Goosepen FormationM
ean
# C
lona
l Ste
ms
n = 25 n = 60
Heartwood Exposure Intact Bark
Clonal Reproduction as a Fnct. of Heartwood Exposure
4
Fig. 6 Dbh is measured in meters tree size in meters (x-axis). Total saplings found per parent tree (y-axis) increases as the dbh of the parent tree increases (n = 112, R2= 0.06, p = 0.01)
Fig. 7 Dbh (measured in meters) as a proxy for tree size along the x-axis, and bole char height (also measured in meters) as a proxy for burn severity along the y-axis demonstrate the weakly linear, positive relationship between tree size and burn severity.
DBH (m)
Tota
l Clo
nal S
tem
s/ T
ree
Clonal Reproduction as a Fnct. of Tree Size
DBH (m)
Bole
Cha
r Hei
ght (
m)
Burn Severity as a Fnct. of Tree Size
5
Fig. 8 Dbh (measured in meters) along the x-axis represents tree size, with the total number of clonal stems per parent tree along the y-axis. The three colorful lines represent varying levels of burn severity, from intact bark to heartwood exposure to goosepen formation. When tree size and fire severity are decoupled, fire severity is the key factor correlated with an increase in clonal stem production (Whole Model- df = 111, F = 8.48, p < 0.0001; Burn Severity- p < 0.0005)
Effects of Burn Severity on Clonal Reproduction, Decoupled from Tree SizeTo
tal C
lona
l Ste
ms

1
References Bowman, D. M. J. S., Balch, J. K., Artaxo, P., Bond, W. J., Carlson, J. M., Cochrane, M. A., …
Pyne, S. J. (2009). Fire in the Earth System. Science, 324(5926), 481–484. http://doi.org/10.1126/science.1163886
Brown, J. K., (1975). Fire Cycles and Community Dynamics in Lodgepole Pine Forests.
Management of lodgepole pine ecosystems, 429-456. Finney, M. A. (1993). Effects of thermal wounding, shading and exogenous auxin on some
sprouting responses of coast redwood seedlings. Tree Physiology, 12(3), 301-309. doi:10.1093/treephys/12.3.301
Finney, M. A., & Martin, R. E. (1993). Modeling effects of prescribed fire on young-growth
coast redwood trees. Canadian Journal of Forest Research Can. J. For. Res., 23(6), 1125-1135. doi:10.1139/x93-143
Johnstone, J. A., & Dawson, T. E. (2010). Climatic context and ecological implications of
summer fog decline in the coast redwood region. Proceedings of the National Academy of Sciences, 107(10), 4533-4538. doi:10.1073/pnas.0915062107
Keeley, J. E. (2002). Fire Management of California Shrubland Landscapes. Environmental
Management, 29(3), 395-408. doi:10.1007/s00267-001-0034-y Keeley, J.E. (2001). Fire and invasive species in Mediterranean-climate ecosystems of
California. Pages 81–94 in K.E.M. Galley and T.P. Wilson (eds.). Proceedings of the Invasive Species Workshop: the Role of Fire in the Control and Spread of Invasive Species. Fire Conference 2000: the First National Congress on Fire Ecology, Prevention, and Management. Miscellaneous Publication No. 11, Tall Timbers Research Station, Tallahassee, FL.
Kilgore, B. M. (1973). The ecological role of fire in Sierran conifer forests. Quaternary
Research, 3(3), 496-513. doi:10.1016/0033-5894(73)90010-0 Lazzeri-Aerts, R., & Russell, W. (2014). Survival and Recovery Following Wildfire in the
Southern Range of the Coast Redwood Forest. Fire Ecology, 10(1), 43-55. doi:10.4996/fireecology.1001043

2
Lorimer, C. G., Porter, D. J., Madej, M. A., Stuart, J. D., Veirs, S. D., Norman, S. P., … Libby, W. J. (2009). Presettlement and modern disturbance regimes in coast redwood forests: Implications for the conservation of old-growth stands. Forest Ecology and Management, 258(7), 1038–1054. http://doi.org/10.1016/j.foreco.2009.07.008
Meyer, M. D., & Safford, H. D. (2011). Giant Sequoia Regeneration in Groves Exposed to
Wildfire and Retention Harvest. Fire Ecology, 7(2), 2-16. doi:10.4996/fireecology.0702002
Noss, R.F. 2000. The redwood forest: history, ecology and conservation of the coast redwoods. Covelo, CA: Island Press. Ramage, B. S., O'hara, K. L., & Caldwell, B. T. (2010). The role of fire in the competitive
dynamics of coast redwood forests. Ecosphere, 1(6). doi:10.1890/es10-00134.1 Webster, K.M., & Halpern C.B. (2010). Long-term Vegetation Responses to Reintroduction and
Repeated Use of Fire in Mixed-conifer Forests of the Sierra Nevada. Ecosphere 1(5). doi: 10.1890/ES10-00018.
![The Redwood gazette. (Redwood Falls, Minn.), 1925-06-17, [p ]. · 2019-10-27 · THE REDWOOD GAZETTE, REDWOOD FALLS, MINNESOTA The Redwood Gazette prints wedding an- nouncements or](https://img.pdfslide.us/doc/110x75/5fa04f2ead664330d06ddb4a/the-redwood-gazette-redwood-falls-minn-1925-06-17-p-2019-10-27-the.jpg)



















![The Redwood gazette. (Redwood Falls, Minn.), 1921-10-05, [p ]](https://img.pdfslide.us/doc/110x75/61e75daafb034a33595cb385/the-redwood-gazette-redwood-falls-minn-1921-10-05-p-.jpg)






![The Redwood gazette. (Redwood Falls, Minn.), 1909-05-19, [p ]](https://img.pdfslide.us/doc/110x75/61f3066c4fb1c01f2e62eb08/the-redwood-gazette-redwood-falls-minn-1909-05-19-p-.jpg)

