Embed Size (px)
Citation preview

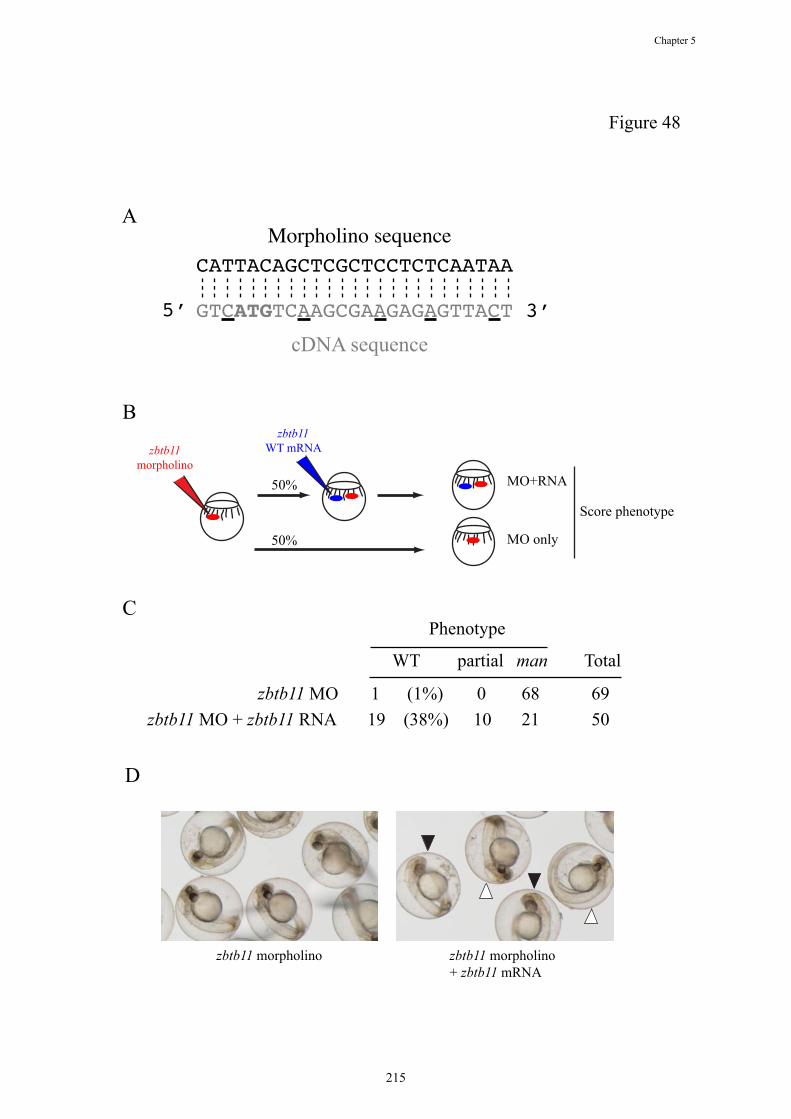
Figure 48. Specificity control for zbtb11 morpholino
A Using site directed mutagenesis, 5 silent mismatches (underlined nucleotides)
were introduced into the zbtb11 WT construct DNA sequence (grey), which
would be predicted to lead to a marked reduction in the binding of the ATG
antisense morpholino (black). The ATG is shown in bold text.
B The specificity of the zbtb11 morpholino was tested by sequentially injecting it,
followed by the zbtb11 WT RNA, into 50% of embryos from the same clutch.
50% of embryos were injected with the morpholino alone. Gross morphology
phenotype of man was then scored.
C Injection of zbtb11 WT RNA into zbtb11 morphant embryos was able to
completely rescue the man phenotype in 38% of embryos, with a further 20%
partially rescued.
D The rescue of the zbtb11 morphant phenotype by zbtb11 RNA was complete in
some embryos (closed arrowheads), which did not display the typical
appearance of a small, darkened head, small eye and hydrocephalus (open
arrowheads).
MO = morpholino.
214
Chapter 5
50%
Score phenotype
zbtb11morpholino
zbtb11WT mRNA
zbtb11 morpholino zbtb11 morpholino+ zbtb11 mRNA
Phenotype
WT man Totalpartial
50%
1 (1%) 0 68 6919 (38%) 10 21 50
A
B
C
CATTACAGCTCGCTCCTCTCAATAA
GTCATGTCAAGCGAAGAGAGTTACT
Morpholino sequence
cDNA sequence
D
MO+RNA
MO only
zbtb11 MOzbtb11 MO + zbtb11 RNA
5’ 3’
Figure 48
215
Chapter 5

zbtb11 morpholino knockdown embryos (Figure 48A-B). Injection of this
mismatched WT zbtb11 RNA into morpholino-injected embryos rescued the
morpholino phenotype completely in 38% of embryos and partially in 20% of
embryos, consistent with the morphant phenotype being due to zbtb11
knockdown only (Figure 48C-D).
5.2.4 Heterologous expression of human ZBTB11
Human ZBTB11 was cloned and synthetic RNA encoding a WT human ZBTB11
protein was transiently over-expressed in man embryos. This rescued the man
phenotype in a similar manner to the zebrafish WT RNA (Table 10),
demonstrating that ZBTB11 can substitute for zbtb11 in this assay. This suggests
that zebrafish and human Zbtb11 have at least some overlap in their biological
roles.
216
Chapter 5

Table 10. Heterologous rescue of man phenotype with human ZBTB11 RNA
Phenotype RNA injected
WT Mutant Other Total
Nil 162 (67%) 60 (25%) 19 (8%) 241
Zebrafish zbtb11 WT 120 (91%) 5 (4%) 7 (5%) 132
Human ZBTB11 WT 113 (65%)2 15 (8%) 45 (26%) 173
2 3/24 randomly selected embryos with WT phenotype genotyped as
homozygous mutant
217
Chapter 5

5.2.5 Structure-function analysis of zbtb11 utilising marsanne rescue
as an in vivo bioassay
Rescue of the man phenotype by over-expression of WT zbtb11 RNA provided
the basis for an assay for testing the function of any modified Zbtb11 protein of
interest. A modified zbtb11 cDNA construct could be created by site directed
mutagenesis or standard cloning techniques, introducing single or multiple amino
acid substitutions, truncations etc. Micro-injection of RNA transcribed from
these modified zbtb11 constructs in man allowed an assessment of their function
relative to the activity of RNA encoding WT Zbtb11, using scoring of the man
phenotype as a readout of biological function. Normally functioning Zbtb11
proteins would rescue the man phenotype equivalently, non-functional proteins
would not. This in vivo bioassay allowed a preliminary structure-function
analysis of the Zbtb11 protein to be undertaken. In particular, the unexpected
finding of Tang et al. that ZBTB11 protein function was normal in the absence of
the C terminal zinc fingers could be tested in an in vivo setting.
To maximise the likelihood that failure to rescue represented a non-functional
mutant protein rather than a technical failure, all modified zbtb11 cDNA
constructs were linearised and RNA in vitro transcribed with a contemporaneous
zbtb11 WT cDNA control. All RNAs were examined by denaturing agarose gel
electrophoresis prior to injection to ensure a band of the expected size and of
optimal quality was produced (Figure 49). All injection RNAs were traced with
rhodamine dextran to ensure delivery, and each modified zbtb11 RNA was
injected in at least 2 independent experiments. On each injection day zbtb11 WT
RNA was injected into embryos from the same clutch as those receiving the
modified RNA. These precautions increased confidence that an intact modified
zbtb11 RNA was injected successfully, but it was not possible to be absolutely
confident that the modified zbtb11 RNA was expressed.
218
Chapter 5

5.2.5.1 Analysis of truncated and mutated Zbtb11 proteins
Using site-directed mutagenesis to introduce premature stop codons into the
Zbtb11 protein at different positions, various truncation constructs were created
(Figure 50). These were tested in the man rescue assay for activity using rescue
of the man gross morphology phenotype as a read-out.
A striking result was that an RNA encoding the Δ657-1146 Zbtb11 construct,
lacking the C terminal zinc fingers, still had normal Zbtb11 function in the man
rescue bioassay (Figure 50). This is an important result because it provides
independent confirmation of the result of (Tang et al. 1999), which showed
conserved repressor function in a CAT reporter assay in a deletion mutant of
human Zbtb11 lacking the zinc fingers. This was an unexpected result given that
other BTB-ZF family members are thought to bind specific DNA sequences via
their zinc fingers, furthermore there is a high degree of sequence conservation
between zebrafish and human Zbtb11 zinc fingers, suggesting a highly conserved
function such as sequence specific DNA binding. In contrast, mRNA encoding
the Δ172-1146 mutant containing only the N terminal part of the protein and
lacking the BTB and zinc finger domains had no bioactivity in this assay (Figure
50).
5.2.6 What is the biological function of the Zbtb11 N-terminus?
5.2.6.1 A putative conserved HHCC zinc integrase motif is present
within the N terminus of Zbtb11
Initial analysis of the Zbtb11 protein using standard domain/motif prediction
tools (InterPro, Pfam, PROSITE) failed to assign any predicted functional
domains or motifs to the 199 amino acids lying N terminal to the BTB domain.
ZBTB11 was the only BTB-ZF family member with a long N terminal amino
acid domain. As that the 116 cysteine residue mutated in man lay within this
219
Chapter 5

Figure 49. Examples of the steps in the preparation of RNAs for
microinjection
Each step in the preparation of RNA for injection was performed in parallel using a
known active zbtb11 construct (WT zbtb11 plasmid) to provide a contemporaneous
active RNA that cold be injected in parallel to the mutant construct to be tested.
A The positive control was linearised contemporaneously with each mutant Zbtb11
construct to be tested in the rescue assay.
B Mutant RNA was synthesised contemporaneously with the positive control
(zbtb11 WT RNA).
220
Chapter 5

A Bzb
tb11
WT
pCS2
+ lin
ear
ZBTB
11 W
T pC
S2+
linea
r
zbtb
11 W
T R
NA
ZBTB
11 W
TR
NA
Figure 49
221
Chapter 5

Figure 50. Test of ability of Zbtb11 truncation mutants to rescue man
mRNAs encoding various Zbtb11 truncation mutants were compared to full-length
Zbtb11 in their ability to rescue the man phenotype at 48 hpf when over-expressed. The
657-1146 mutant was still able to rescue man, however the 172-1146 mutant was
not.
mRNA injections were into man heterozygous in-crosses where 25% mutant embryos
would be expected.
222
Chapter 5

zbtb
11 W
T13
/556
(2
%)
Y Y5/
209
(2%
)
N51
/185
(2
7%)
BTB
Uni
njec
ted
192/
823
(23%
)N
/A
C2H
2 zin
c fin
gers
RN
A in
ject
edR
escu
e?N
o. o
f mut
ant
embr
yos (
%)
Pred
icte
d pr
otei
n
zbtb
11 ∆
657-
1146
1146
zbtb
11 ∆
172-
1146
659
199
Figure 50
223
Chapter 5

highly conserved section of the protein, a more detailed search for potential
functional motifs within this section of the protein was undertaken. BLAST
searching and more detailed analysis of results from motif prediction algorithms
suggested homology of a region of human ZBTB11 N terminus to a HHCC motif
contained within retroviral/retrotransposon DNA integrases (Figure 51). The 116
cysteine residue mutated in man corresponded to the first C in this HHCC motif.
Retroviral/retrotransposon integrases (IN) contain 3 domains, a N terminal
HHCC motif, a catalytic core and a C terminal domain. Within integrases, the N
terminal HHCC motif is essential for IN function and has been shown to bind
and co-ordinate zinc, but does not possess specific DNA binding properties
(Yang et al. 2001). It appears to have a role in promoting IN dimerisation (Yang
et al. 2001). The N terminal HHCC motif in Zbtb11 may be important for
facilitating homo- or hetero-dimerisation of the protein in conjunction with the
BTB domain.
5.2.6.2 Sequential mutagenesis of individual critical residues within the
putative HHCC motif
In order to assess the importance of the residues in this motif in the zebrafish
Zbtb11 protein, site-directed mutagenesis was undertaken to mutate each residue
in Zbtb11 corresponding to the HHCC motif. The 3 mutant Zbtb11 proteins
created were H79A, H86A and C119S; C116S is the man mutant protein. The
H79A, H86A and C119S constructs were inactive in the man rescue bioassay
(Figure 52).
224
Chapter 5

5.2.7 Mammalian Zbtb11 biology
5.2.7.1 Results of literature and database analysis of mammalian
Zbtb11 expression
Published data on mammalian Zbtb11 expression suggests it is expressed in most
adult tissues. In humans, northern blotting demonstrated expression in 8/8 adult
tissues of a 6.2 Kb mRNA (Tang et al. 1999), which was expressed at similar
levels in 7/8 tissues, more weakly in liver (Figure 53A). Microarray data with
Affymetrix probe 204847_at designed from the 5.1 Kb ZBTB11 mRNA
(accession number NM_014415) demonstrated highest expression in some brain
tissues, CD34+ bone marrow cells, T and B lymphocytes, NK cells, CD33+
myeloid cells and CD14+ monocytes (Su et al. 2002) (Figure 53B) but showed
median level expression in many tissues. Low expression was seen in heart, liver,
lung and kidney tissue. Other data supports its constitutive expression throughout
human myelopoiesis from CD34+ stage to mature neutrophil (Ferrari et al. 2007)
and in NK cells (Dybkaer et al. 2007).
During the later stages of the project, a rabbit anti-human polyclonal
ZBTB11antibody became available (HPA015328; Sigma). The expression
pattern of ZBTB11 was characterized in the Human Protein Atlas (Ponten et al.
2008). Highest expression was seen in cells in the colon, duodenum,
hippocampus, cerebellum and adrenal gland, however moderate to strong nuclear
expression was seen in cells in lymphoid tissues such as lymph node, tonsil and
spleen. The expression in lymph nodes was strongest in larger cells within the
germinal follicles and weaker in cells outside the follicles. Expression within
different haematopoietic cell types in the bone marrow was not able to be
determined due to poor specimen quality.
In the adult mouse, microarray data with Affymetrix probes 1454826_at and
gnf1m33018_at designed from a 5.0 Kb Zbtb11 mRNA (accession numbers
225
Chapter 5

Figure 51. Homology between the Zbtb11 N terminus and the zinc integrase
domain
Amino acid alignment of a section of the zinc integrase domain of S. cerevisiae, with
the N terminus of human and zebrafish Zbtb11. The critical HHCC residues are shown
(grey shading) along with other residues demonstrated to be functionally important in
integrase function (boxed).
Alignment was constructed using Clustal W. “*” = identical residue, “:” = conserved,
“.” = semi-conserved.
226
Chapter 5

HLGP-GGTHHTRHQTWHYLSKTYWWRGILKQVKDYIKQCSKCQ
HDHTLFGGHFGVTVTLAKISPIYYWPKLQHSIIQYIRTCVQCQ
HITS-GGEHLNQQQTWEIISQKYWWRGVLKQVKDCIKECIHCQ
H H C C
N C
* . * * * :* *:* : :.: : *: * :**
HH-CC D,D35-E GPY/F
Zinc bindingDimerisation Catalytic domain DNA binding
S.cerevisiae Ty3
H.sapiens ZBTB11
D.rerio Zbtb11
Ty3/Gypsy integrase
Figure 51
227
Chapter 5

Figure 52. Functional analysis of residues within the putative HHCC motif
in Zbtb11
The man rescue assay was used to assess the bioactivity of constructs created by site-
directed mutagenesis of the 4 residues in the HHCC motif (grey shading). The C116S,
C119S and H86A proteins were biologically inactive. Only preliminary data on the
H79A protein was available which suggested it was inactive. The changed residues are
shown in red.
mRNA injections were into man heterozygous in-crosses where 25% mutant embryos
would be expected.
228
Chapter 5

C11
6S
1
WT
6
H86
A
3
C11
9S
2
7986
116
119
HITSGGEHLNQQQTWEIISQKYWWRGVLKQVKDCIKE
LNQQQTWEIISQKYWWRGVLKQVKDCIKECIHIHC
Uni
njec
ted
6
HITSGGEHLNQQQTWEIISQKYWWRGVLKQVKDCIKE
LNQQQTWEIISQKYWWRGVLKQVKDCIKESIHIHC
HITSGGEALNQQQTWEIISQKYWWRGVLKQVKDCIKE
LNQQQTWEIISQKYWWRGVLKQVKDCIKECIHIHC
HITSGGEHLNQQQTWEIISQKYWWRGVLKQVKDCIKE
LNQQQTWEIISQKYWWRGVLKQVKDCIKECIHIHS
zbtb
11 R
NA
Pred
icte
d A
A S
eque
nce
No.
expe
rimen
tsN
o. m
utan
ts (%
)x
+/- s
d
H79
AAITSGGEHLNQQQTWEIISQKYWWRGVLKQVKDCIKE
LNQQQTWEIISQKYWWRGVLKQVKDCIKECIHIHC
2
3.2
+/- 2
.0
11.8
18.9
+/-
4.7
15.0
+/-
1.2
16.2
+/-
14.8
19.5
+/-
5.1
Figure 52
229
Chapter 5

Figure 53. Published expression data on mammalian expression of Zbtb11
A Ubiquitous expression of ZBTB11 was demonstrated by northern blot on
various human tissues.
B Differential ZBTB11 expression was seen across a range of human normal
tissues and malignant cell lines. High expression was seen in CD14+ monocytes,
NK cells, lymphocytes, CD34+ bone marrow cells and in the chronic myeloid
leukaemia cell line K562.
C Expression of Zbtb11 in mouse showed a similar pattern with high expression in
brain tissues, lymphocytes and NK cells.
D Embryonic expression of Zbtb11 in mouse was demonstrated by WISH in the
brain at E10.5.
Data are from (Tang, Westling et al. 1999) (A), (Su, Cooke et al. 2002) (B, C) and
(Ficker, Powles et al. 2004; Gray, Fu et al. 2004) (D).
WISH= Whole mount in situ hybridisation.
230
Chapter 5

Mouse
Organism
A B
C D
Figure 53
231
Chapter 5

XM_001480244 and XM_001481121) showed high expression in T, B, NK cells
and some brain tissues, but also median expression in many other tissues (Su et
al. 2002) (Figure 53C). Embryonic mouse Zbtbt11 expression visualised using
WISH showed expression in the central nervous system at E10.5 and expression
in inner ear structures at E13.5 (Ficker et al. 2004; Gray et al. 2004) (Figure
53D).
5.2.7.2 Expression of Zbtb11 in haematopoietic diseases
Microarray data with probe 204847_at (Su et al. 2002) demonstrated high
expression of ZBTB11 in the K562 chronic myeloid leukaemia cell line, a cell
line derived from a human erythroleukaemia (Lozzio et al. 1975). Sequencing of
the full length ZBTB11 WT cDNA that was amplified by PCR from RNA
derived from K562 cells did not demonstrate any mutations. Several other cell
lines derived from T-lymphoblastic leukaemia (MOLT-4), Burkitt’s lymphoma
(Daudi/Raji) and promyelocytic leukaemia (HL-60) had near-median ZBTB11
expression. Further investigation of the mechanism of ZBTB11 dysregulation in
the K562 cell line would be interesting.
There is some data suggesting ZBTB11 may be dysregulated in B cell lymphoma.
A genome wide association study demonstrated genomic amplification of a
region on human chromosome 3q12 (102.08-103.16 Mb), containing ZBTB11, in
a subtype of DLBCL, activated B-cell-like (ABC-type DLBCL) (Lenz et al.
2008). The authors put forward NFKBIZ as a possible candidate gene but did not
provide further data to support this hypothesis. Direct evaluation of the role of
ZBTB11 amplification in ABC-type DLBCL would be interesting. Data in the
human protein atlas on the ZBTB11 antibody included analysis of its expression
across a panel of tumour specimens. Of interest, specific nuclear expression was
seen in 12/12 lymphoma specimens, with higher expression in higher grade
(DLBCL) than the lower grade lymphomas (Ponten et al. 2008).
232
Chapter 5

5.3 Chapter 5- Discussion
5.3.1 Expression of zbtb11
Zebrafish zbtb11 is expressed ubiquitously in both the embryo and adult. Despite
this ubiquitous expression pattern, the loss of zbtb11 function during embryonic
development had specificity. Within the haematopoietic compartment, there was
marked loss of myeloid cells, but relatively little effect on erythroid cells.
Several congenital neutropenia/bone marrow failure syndromes are due to
mutations in ubiquitously expressed proteins (SDBS, RPS19, HAX1, G6PC3)
involved in ribosome biogenesis, mitochondrial membrane stability and glucose
homeostasis (Klein et al. 2007; Ganapathi et al. 2008) (Boztug et al. 2009).
Despite the ubiquitous expression of these proteins, their loss-of-function disease
phenotypes are most marked within the haematopoietic compartment. It is
possible that the particular proliferative (Li et al. 2003) or apoptotic (Cheung et
al. 2007; Klein et al. 2007; Terzian et al. 2007; Yamaguchi et al. 2007; Ganapathi
et al. 2008; Steimer et al. 2009) profile of some haematopoietic cells renders
them more sensitive than other cell types to the effect of loss of function of some
ubiquitously expressed proteins. Another possibility is that zbtb11 has different
downstream targets in different differentiating cell types. One downstream target
may be more highly expressed in myeloid cells than erythroid cells, leading to
myeloid-specific dysregulation of the pathway. If this target were critical for
myeloid cell development, this could lead to myeloid cell loss without a
pronounced effect on erythroid differentiation. The ubiquitously expressed
transcription factor Sp1 regulates several myeloid-specific targets such as CD11b
and CD14 in this way (Tenen et al. 1997; Hauses et al. 1998).
233
Chapter 5

5.3.2 Conservation of Zbtb11 function across species
Orthologous genes are derived from a common ancestor and share common
function. There are 3 lines of evidence that contribute to recognition of
orthology: (1) sequence homology, (2) shared genomic context (synteny), (3)
shared function. Zebrafish and human Zbtb11 fulfil 2 of these 3 criteria.
Sequence homology is equivalent to other haematopoietic transcription factors
orthologous between human and zebrafish (Liao et al. 1998; Lieschke et al. 2002;
Wei et al. 2008). Syntenic analysis did not demonstrate a shared genomic
context. However zebrafish and humans have had 450 million years to
independently reorganise their genomes and only approximately 80% of putative
orthologue pairs between zebrafish and humans share synteny, compared to 90%
between mouse and humans (Woods et al. 2000). Thus, the lack of synteny does
not disprove orthology. The heterologous rescue with human ZBTB11
demonstrated functional substitution. Together these data suggested that
zebrafish zbtb11 is likely to be the orthologue of human ZBTB11.
The mammalian expression data indicate there is scope for conservation of a cell
autonomous role for Zbtb11 within the haematopoietic compartment, nervous
system and gut at least, although this has not yet been proven.
5.3.3 Structure-function analysis of zbtb11 sub-domains
The rescue of man by constructs encoding a truncated Zbtb11 protein lacking the
C-terminal zinc fingers supports the previous in vitro work on human ZBTB11
showing that the C terminal zinc fingers are not critical for function, at least in
the assay systems used in this work and that of (Tang et al. 1999). Many BTB-ZF
proteins form both homo- and heterodimers, their BTB domains recruiting other
234
Chapter 5

proteins, often co-repressors to assemble large multi-protein complexes
responsible for the transcriptional activity of the complex at its site of DNA
binding (Costoya 2007). If the function of Zbtb11 in embryonic development
were mediated through such a heterodimeric complex, where the dimerisation
partner imparts the DNA binding specificity, the role of Zbtb11 in such a
complex could be limited to formation of the complex itself and/or recruitment of
other proteins. DNA binding via its zinc fingers could be important in other
temporal or spatial settings. Alternatively, DNA binding specificity may lie
within the N terminal section of the protein. As (Tang et al. 1999) were able to
demonstrate that full-length ZBTB11 does bind DNA in an EMSA assay, but that
a ZBTB11 protein consisting of the first 482 amino acids (the N terminal half of
the protein, lacking the zinc fingers) could still bind a promoter (MTIIA) and
cause repression in a CAT assay, this also raises the possibility that DNA binding
function lies outside the zinc fingers. As DNA binding by the BTB domain itself
has not been shown in any BTB-ZF proteins, it is probable that this DNA binding
specificity lies outside the BTB domain elsewhere in the first half of the protein.
The role of the 116 cysteine residue within a putative HHCC motif within the N
terminus of Zbtb11 was explored using sequential mutagenesis of several other
residues within the putative HHCC motif. Although incomplete, these data
suggest a critical role for these residues and this domain in Zbtb11 function. The
mechanism of this is uncertain, but given its close proximity to the BTB domain,
it is possible that the HHCC motif interacts with the BTB domain. As Zbtb11 is
the only BTB-ZF member with a long N terminal extension, and as it is not
homologous to any protein of known structure (at least, as recognised by BLAST
searching using the 199 amino acids), there is no basis on which to predict the
structure of the N terminus. A solved crystal structure of dimeric Zbtb11 would
provide the best data on which to base further analysis of the function of the N
terminus and the residue mutated in man.
235
Chapter 5

Alternatively the HHCC motif could play a role in DNA binding, given that the
DNA binding specificity is known to lie outside the zinc fingers, and is unlikely
to be within the BTB domain itself. Measurement of the DNA binding
specificity, or repression activity in the MTIIA CAT assay of the HHCC mutant
proteins would be informative.
One caveat on these studies is, particularly where no function was observed, that
it would be optimal to directly demonstrate that the mRNA injected resulted in
protein expression. Western blotting with a Zbtb11 antibody would be the most
direct way of demonstrating expression but an antibody against the zebrafish
protein was not available. A commercially available polyclonal rabbit anti-
human antibody only became available during the last phase of this project. This
polyclonal antibody was raised against a 123 amino acid peptide corresponding
to the linker region of the human protein, a region poorly conserved between
species. As such it would be improbable that it would cross-react with zebrafish
Zbtb11.
Where no antibody is available protein production could be verified by epitope
tagging. While this may enable translation to be proved, it remains possible that,
when a loss of activity is observed, the tag itself had impaired function.
Similarly, a GFP tag would enable visualisation of expression of the protein by
fluorescence, but with the same caveat about effect on protein function.
In the assays presented, the validity of the negative results is supported by 1)
sequence verification of the template plasmids, 2) confirm the intactness of the
RNA by gel electrophoresis, 3) repeating the assay with several independently
synthesised RNAs, 4) always injecting a positive control RNA on the same day.
The rescue assay could be sensitised to detect residual function in proteins
encoded by hypomorphic alleles by using zbtb11 morphant embryos or a verified
null allele as the assay vehicle. In this setting any activity of the mutant proteins,
236
Chapter 5

compared to control embryos, would be evidence of protein expression. This
could be done in morphant embryos, although it would require that the mutations
to be tested be introduced on a background of the WT zbtb11 cDNA construct
containing the mismatched sequence to the morpholino.
The mutagenesis analysis on the HHCC motif carries the same technical caveats
as previously discussed with regard to demonstration of expression of the
apparently inactive mutant proteins. In addition, as mutations at all 4 residues of
the HHCC motif were inactive, design of an instructive mutation within this
motif as a negative control was being addressed during the final stages of the
project.
5.3.4 Zbtb11 function studies
5.3.4.1 Comparison between mutant and morphants
Once zbtb11 was identified as the mutant gene in man, descriptive studies of the
effects of loss-of-function could be undertaken in zbtb11 morphant embryos.
Morphants had several advantages: They provided a more homogeneous
population of embryos lacking zbtb11 function than in the one-quarter of
embryos seen in man heterozygous in-crosses and were less likely to be
susceptible to subtle influence of temperature variation. It was also easier to
assess early time-points prior to the onset of a visible phenotype, without
genotyping individual embryos or making an inference based on Mendelian
inheritance and numbers of embryos in a given clutch.
However the analysis of morphant embryos, particularly when making
comparisons to man, has several caveats. Firstly man may be a “weak”
hypomorphic allele and the effect of MO knockdown might be stronger. The
237
Chapter 5

temperature sensitivity data presented in chapter 3 demonstrated that with a 7°C
lowering in temperature, the mpx deficiency could be partially rescued. Such
temperature sensitivity is typical of hypomorphic alleles, which are due to
protein instability rather than gross protein missfolding. However, man may be
temperature sensitive for reasons unrelated to the Zbtb11 mutation itself. Other
aspects of the man phenotype could also be susceptible to variations in
temperature.
Secondly, and alternatively, morphants are always potential hypomorphs, but in
different ways to man. For example, man is a zygotic mutant, an ATG
morpholino, as opposed to a splice-site morpholino or man, knocks down
maternal transcripts in addition to zygotic transcripts. This is only an issue if
quantitative comparison is required, or loss of the maternal transcripts results in a
phenotype that is epistatic to later phenotypes.
Thirdly, the degree of knock-down achieved by a morpholino approach
diminishes over time. Hence morphant embryos were not studied at timepoints >
48 hours. The lack an antibody meant that ongoing Zbtb11 knockdown could not
be confirmed.
As the zbtb11 exon1/intron 1 splice site morpholino was biologically inactive
and only the ATG morpholino produced a phenotype, it was not possible to
quantitate the degree of zbtb11 knockdown by an RT-PCR approach. The lack of
an antibody to zebrafish Zbtb11 precluded analysis of protein levels in the
knockdown embryos. However, as the phenotype in zbtb11 morphant embryos
was more severe than in man, this suggested that the zbtb11 loss-of-function was
at least equivalent to, if not more severe than, the man allele within the first 48
hpf. As a reference point, an allele known or predicted to be null, rather than one
that is potentially hypomorphic, would provide a stable zygotic mutant
background with no zbtb11 function. Such an allele is being sought through a
TILLING approach.
238
Chapter 5

Antisense morpholino oligonucleotides may have non-specific (off-target)
effects. Some of these may be due to activation of p53 (Robu et al. 2007). The
best specificity control for a morpholino is exact concordance between a
morphant and mutant phenotype. Another good specificity control is to
demonstrate complete rescue of the morpholino phenotype by overexpression of
the RNA it is designed to target, thereby rescuing only the on-target aspects of
the morpholino phenotype. Other specificity controls that are commonly used,
such as a second non-overlapping morpholino, are not necessarily required if
these better tests of specificity are available (Eisen et al. 2008).
The difference between zbtb11 morphants and man mutants was in the
expression of markers of primitive haematopoiesis which were normal in man
mutants, but abnormal in morphants. This could be due to any of the factors
mentioned above. Other technical factors may also have affected the expression
of these genes. The degree of zbtb11 knockdown may not have been uniform
across the morphant embryo population and this may have led to variable effects
on gene expression. The 2 genes most significantly reduced in expression, runx1
and cmyb are initially expressed in the PLM closest to the timepoint selected for
examination. If the zbtb11 morpholino caused a delay in global embryo
development, the effect on gene expression would be most marked on those
genes first expressed close to the timepoint examined. This would have to be an
effect of the zbtb11 morpholino itself, not just embryo injection, as the control
morpholino had no effect on runx1 or cmyb expression. This seems unlikely, as
both the zbtb11 and control morphant embryos were staged by somite number.
This could be more rigorously tested by analysis of a non-haematopoietic gene
e.g. deltaC in zbtb11 morphants at the same timepoint, to demonstrate equivalent
developmental age. The specificity of these gene expression differences in
morphants could also be tested by concurrent injection of the mismatched WT
zbtb11 RNA into a subset of embryos, and evaluating for normal expression
pattern of these genes.
239
Chapter 5

5.3.4.2 Limitations of the haematopoietic analysis
The early embryonic lethality of the man mutation precluded an exhaustive
analysis of the effect of zbtb11 loss on later haematopoiesis. This would be
achieved more completely utilising a conditional knockout allele to overcome the
embryonic lethality of homozygous loss of zbtb11. Although the partial rescue of
the man phenotype was possible at a lower temperature, this extended the period
of embryo survival only by a few days. A conditional mouse allele e.g. Mx-Cre
would enable advantage to be taken of well-established reagents and techniques.
Alternatively tissue specific deletion within the haematopoietic compartment
could be undertaken, using a transgenic line with a haematopoietic specific
promoter such as vav-Cre (Georgiades et al. 2002) to analyse the effects of loss
of Zbtb11 within the haematopoietic compartment in isolation from its effects in
other tissues.
A major current limitation of haematopoietic analysis in zebrafish embryos and
adults is the inability to study specific haematopoietic cell sub-populations
quantitatively using multiple antibody markers. In mouse, this is commonly done
using fluorescent antibodies to multiple proteins by flow cytometry (Yeung et al.
2009). Due to a lack of available zebrafish antibodies, particularly to cell surface
proteins differentially expressed on haematopoietic cells, this is currently not
possible to the same extent as in mouse. In addition functional analyses (e.g. by
bone marrow reconstitution or haematopoietic cell culture assays) are not fully
developed in zebrafish, despite some recent advances (Stachura et al. 2009).
Generation of zebrafish-specific antibodies is a high priority for the zebrafish
community (http://zfin.org/zf_info/news/siteNews.html#zfpoll, accessed
29/03/2009).
240
Chapter 5

The haematopoietic phenotype analysis both in the man mutant and in the zbtb11
morpholino injected embryos was limited by this lack of available antibodies.
The loss of transcripts as assessed by WISH does not necessarily translate to loss
of the cell type expressing a particular transcript. The demonstration of loss of
multiple myelomonocytic transcripts (mpx, lcp, lyz, npsn, csf1r) strongly
suggested loss of myelomonocytic cells as distinct from myelomonocytic
transcription, but did not prove it. In addition, compared to the accurate
quantitative analysis of sub-populations possible by using multiple markers by
flow cytometry, WISH is limited to simultaneous analysis of only a few markers.
Although WISH enables an assessment of the actual numbers of marked cells in
the whole organism, rather than the relative quantitation achieved by flow
cytometry, it is a method of analysing a more general haematopoietic population
(e.g. erythroid cells), rather than a specific sub-population defined by the
simultaneous expression of several markers. Several haematopoietic-specific
transgenic zebrafish lines are now available (gata1:GFP/dsRED (Long et al.
1997; Vogeli et al. 2006), mpx:GFP (Mathias et al. 2006; Renshaw et al. 2006),
lyz:GFP/dsRED (Hall et al. 2007), spi1:GFP (Ward et al. 2003; Hsu et al. 2004)
and others) allowing FACS sorting of cells by single or double colour
fluorescence, but this approach is still limited by the number and cell specificity
of transgenic strains available. Significantly, as man is embryonic lethal, very
small numbers of cells (approximately 40 mpx expressing cells per embryo at 48
hpf) are available for study in mutant embryos at the time-points prior embryonic
lethality. As the embryonic lethality in man is prior to the appearance of
haematopoietic cells in the kidney, the isolation of these cells from embryos is
challenging as no haematopoietic organ can be separated from the rest of the
embryo by dissection. Although FACS sorting by fluorescence of haematopoietic
cells from dissociated, pooled embryos has been successfully performed
(Bertrand et al. 2007), it is technically very difficult to isolate or quantitate such
low numbers of fluorescent cells from a background of large numbers of non-
fluorescent cells forming the rest of the embryo. This has been achieved in the
241
Chapter 5

laboratory for WT embryos (Felix Ellett, unpublished data), but has not yet been
used successfully in myeloid-cell depleted mutants.
5.3.5 Conclusion- Zbtb11 function
While the experiments in this chapter are only an initial investigation of the
function of Zbtb11, they provide evidence that it is a protein with an interesting
structure-function relationship of relevance to early haematopoietic development.
242
Chapter 5

Minerva Access is the Institutional Repository of The University of Melbourne
Author/s:Carradice, Duncan Peter
Title:Genetic basis of congenital myeloid failure syndromes in mutant zebrafish
Date:2010
Citation:Carradice, D. P. (2010). Genetic basis of congenital myeloid failure syndromes in mutantzebrafish. PhD thesis, Walter & Eliza Hall Institute of Medical Research, affiliated with theUniversity of Melbourne, The University of Melbourne.
Publication Status:Unpublished
Persistent Link:http://hdl.handle.net/11343/35545
Terms and Conditions:Terms and Conditions: Copyright in works deposited in Minerva Access is retained by thecopyright owner. The work may not be altered without permission from the copyright owner.Readers may only download, print and save electronic copies of whole works for their ownpersonal non-commercial use. Any use that exceeds these limits requires permission fromthe copyright owner. Attribution is essential when quoting or paraphrasing from these works.





























