Embed Size (px)
Citation preview

168 Biochimica et Biophysica Acta, (1992) ! 68-173 © 1992 Elsevier Science Publishers B.V. All rights reserved 0005-2760/92/$05.00
BBALIP 53969
Fibric acid derivatives: effects on the synthesis of isoprenoid lipids in cultured human lymphocytes
Alison Henry a, Char l e s M. Al l en a and Pe te r W. S t acpoo le a,b
a Department of Biochemistry and Molecular Biology, ~ Department of Medicine, College of Medicine, University of Florida, Gainesville, FL (USA)
(Received 30 March 1992)
Key words; Cholesterol; Dolichol; Ubi~,,uinone; Lymphocyte; Mevaionate
Fibric acid derivatives have been demonstrated to reduce circulating iipoprotein and triacylglycerol concentrations and to inhibit hydroxTmethylglutaryl CoA reductase, a key regulatory enzyme of cholesterol biosynthesis. This study describes the effect of four fibric acid derivatives on the biosynthesis of isoprenoid products from acetate and mevalonate in Molt-4 cells, a human leukemic T-lymphocyte cell line. The isoprenoids analyzed were cholesterol as well as dolichoi and ubiquinone, alternative products of the branched isoprenoid biosynthetic pathway. None of the fibric acid derivatives showed significant effects on the synthesis of cholesterol from acetate or mevalonate and there was little change in the flux of these metabolites into either dolichol and ubiquinone compared to cells grown in drug-free medium, Therefore, in contrast to the reported inhibitory effects of fibric acids on hepatic sterol synthesis in rats and humans and on hydroxymethylglutaryl CoA reductase activity in human nonmalignant lymphocytes, our results show that these drugs do not significantly affect any of the post-rcductase enzymes in the branched metabolic pathways leading from acetate to dolichol, ubiquinone and cholesterol in short term culturing of human malignant lymphocytes,
Introduction
lsoprenoid biosynthesis follows a branched pathway that leads to several metabolites including cholesterol, a sterol used for bile acid and steroid hormone produc- tion, dolichol, a sugar carrier in N-linked glycoprotein biosynthesis, and ubiquinone, a component of the mitochondrial electron transport chain. Regulation of the pathway for isoprenoid biosynthesis is important to control the availability of these metabolites for various cellular processes. Several mechanisms function to reg- ulate cholesterol synthesis, including transcriptional control, enzyme degradation, allosteric regulation and covalent modification of key regulatory enzymes in the metabolic pathway including 3-hydrozy-3.methyl- glutaui (HMG) coonzyn.e A reductase (EC 1.1,1.34), the endoplasmic reticulum enzyme that catalyzes the roductive synthesis of mevalonate [1], Several other enzymes involved in cholesterol metabolism, including squalene oxide cyclase, squalene synthase, farnesyl diphosphate synthase and sterol oxidases, are also un-
Correspondence to: C,M. Allen, Department of Biochemistry and Molecular Biology, J. HiUis Miller Health Center, Box J-245, Univer- sity of Florida, Gainesville, FL 32610, USA.
der similar control [2-6]. Studies with whole cells and cell-free systems indicate the rate-limiting function of HMG CoA reductase imposes a tight control over the flow of carbon delivered to the post-reductase steps of stcrol synthesis [4,7], although the biosynthesis of dolichol and ubiquinonc does not appear to be as tightly regulated as that of cholesterol [8,9]. Drugs designed to reduce cholesterol synthesis by the inhibi- tion of HMG CoA reductase and other enzymes early in the biosynthetic pathway reduce carbon flux for cholesterol biosynthesis, but they may also alter rates of formation of other isoprenoids.
The fibric acid derivatives, gemfibrozil and clofi- brate are known to lower predominantly serum triacyl- glycerol and very low-density lipoprotein levels. Clofi- brate also lowers plasma cholesterol levels in hyperlipi- demic patients without an increase in cholesterol bio- synthesis [10]. Drugs of this class exert their lipid- lowering action in part by inhibiting hepatic triacyl- glycerol synthesis [11]. They may also inhibit HMG CoA reductase activity [12-15] and thereby alter the synthesis of various isoprenoids.
We tested this possibility by determining choles- terol, dolichol and ubiquinone formation in cultured human lymphocytes, using radiolabeled acetate and mevalonate as substrates entering the isoprene path-

way prior to and after HMG CoA reductase. None of the fibric acid derivatives tested exhibited significant effects on the rates of synthesis of cholesterol from acetate or mevalonate and :here was little change in the flux of these metabohtes into dolichoi and ubiquinone.
Materials and Methods
The fibric acid derivatives were provided by Dr. Roger S. Newton, Parke-Davis Pharmaceuticals, Ann Arbor, MI.
Tissue culture. Malignant T-cell lymphocytes (Molt-4) cells were obtained from Dr. N. Lawrence Edwards, Department of Medicine, University of Florida. Cells were grown in suspension culture at 37°C in 95% air/5% CO, in 200 ml of culture medium, which consisted of RPMI-1640 (Gibco, Grand Island, NY, USA) supplemented with 10% fetal calf serum (Hy- clone, Logan, UT, USA) and !% antibiotic/antimycotic (Gibco) [16]. Cell numbers were counted with a hemo- cytometcr and cultures exhibiting greater than 90% cell viability, as monitored by Trypan Bluc exclusion, were used. When the cells reached (3-4)" l0 s cells/ml, one quarter of the suspension was diluted with fresh media to maintain ceil growth. The remainder of the cells were used for experimentation.
Metabolic labelling of isoprene lipids. Cells ((2-12)" 10 ~') were incubated in 1 ml RPMI-1640 medium with or without a fibric acid derivative and 10 /~Ci [2- 14C]acetate (53 mCi/mmol, Amersham) or 10 /.LCi [5-3H]mcvalonate (1.26 Ci/mmol, Amersham) as de- scribed below in 25 cm 2 tissue culture bottles gently agitated on a rocking platform at 37°C in an ambicnt atmosphere for the duration of the incubation. As a result of studies to determine the toxicity of the various fibratc derivatives, an incubation protocol was adopted whereby a 4 h incubation period in the presence of 0 /zM, 100 ~M or 250/zM drug preceded the addition of radiolabclled acetate or mevalonate which remained for a further 6 h.
Isolation of isoprene lipids. Following the 6 h incuba- tion the cells were harvested from the incubation medium, washed free of excess radiolabel by centrif- ugation and rcsuspcndcd three times with isotonic saline at room temperature. When [3H]mevalonatc was the labelled precursor, the pellets of cells were resus- pcndcd in 3 mi of 20% KOH and 0.08% pyrogailoi in methanol. In each case 20 ~g each of unlabelled cholesterol, dolichol and ubiquinonc-10 (Sigma) were added as carriers to the saponification mixture. Mild alkaline hydrolysis was achieved by rcfluxing the mix- ture for 1 h at 80°C under nitrogen. The non-saponifia- bit lipids were isolated from the saponification mixture by extraction three times with 3 ml CHCl3:mcthanoi (2:1, v/v). In each case 1 ml methanol was added first,
169
the sample mixed and 2 ml cHC'! 3 added before final vortexing. The pooled CHCI 3 extracts were reduced to dryness under N 2 and the residue redissolved in 2 ml of methanol. The non-saponifiable lipids were purified on a C~8 reverse phase Sep-Pak mini-column (Waters, K " ~ , 4 . . . . l - - . . . . . . . J . . . . . . . . . ,viii,or,.,, MA, I t~A) nr,~viol,~iv equilibrated with methanol. Cholesterol was eluted with 10 ml of methanol and ubiquinone and dolichol with 10 ml of ethanol. The methanol and ethanol eluates were col- lected separately and the solvent removed under vac- uum. The residual material was redissolved in a known volume of 0.1% isopropanol in hexane before HPLC analysis. When [14C]acetate was the precursor, the order of the saponification and lipid extractions were reversed. The iipids were first extracted from the cell pellets with CHCla/methanol; standards were added and saponification was carried out. The nonsaponifi- able lipids were isolated by CHCia/methanol extrac- tion and subjected to Sep-Pak chromatography before final lipid analysis. An assessment of the incorporation of radiolabel and sample recovery were determined by radiochemical analysis of aliquots of the total non- saponifiable material and both methanol and ethanol Se.p-Pak eluates. Preliminary. experiments :!~ilizing a known quantity of radiolabel from both [3H]cholesterol and [3H]dolichol consistently showed a 80-90% recov- ery of non-saponifiable radioactivity from broken cells by these methods.
HPLC. Analysis of ubiquinone, dolichol and choles- terol was achieved by separate analysis of the methanol and ethanol Sep-Pak eluates on straight phase HPLC by the method of Palmer et al. [17]. A Perkin-EImer Series 4 HPLC apparatus (Perkin-EImer, Norwalk, CT, USA) was used with an LC-75 Perkin-Elmer variable wavelength detector and a 10/~m cyanopropyl (250 x 4.9 ram) column (Brownlee Labs, Santa Clara, CA, USA). A mobile phase of 0.1% isopropanoi in hexai~e was pumped at a constant rate of 1 ml/min. The ultraviolet absorbance of the column eluate was moni- tored at 210 nm. This system gave base line separation of ubiquinone (elution volume 7.5 ml), dolichol (9.7 ml) and cholesterol (13.7 ml). When analyzing for the radiolabelled products, the column eluates were col- lected in 500/~l fractions, the solvent removed under a stream of nitrogen and the residue assayed for "~H or ~4C in 4 ml toluene-based scintillation fluid. Recovery of radioactivit~ from the HPLC column was generally > 90%. The incorporation of [s4C]acetate or [3H]me- val-onate into each product is expresse~ as dpm/10 ~' cells. The data is reported as the means + S.D. of the sample. The number (n) of tests for each sample ranged from 2-4 depending on the fibrate tested.
Radiolabei in the nonsaponifiable fractions which was not found in the dolichol, ubiquinone or choles- terol fractions may be accounted for by procedural losses or due to the formation of products which were
I70
not identified and quantitated. For example squalene, which was a component of the methanol (cholesterol) fraction following Sep-Pak chromatography, eluted among several minor radiolabelled products before cholesterol on HPLC, but it was not identified or quantitatcd.
Results
Incorporation of radiolabelled acetate or meval- onate into isoprenoid products varied among cell types. Freshly isolated human mononuclear leukocytes [18,19] and non-malignant B-iymphoblast~ (IM-9) [18] gave poor incorporation of [~H]mevalonate into non- saponifiable lipids including cholesterol, ubiquinone and dolichol. In contrast, malignant T-iymphocytes (Molt-4 cells) demonstrated much better incorporation of radiolabel [18]. The ability of the malignant cells to incorporate mevalonate into non-saponifiable lipids paralleled the specific activity of the cellular HMG Co,& reductase [20]. Therefore, Molt-4 cells were cho- sen to examine the effect of four fibric acid derivatives on the ability of human leukocytes to incorporate [=~C]acetate and [SH]mevalonate into cholesterol, dolichoi and ubiquinone.
Two conditions were established before beginning these experiments: (1) that the biosynthesis of tholes-
TABLE I
The effect of 25-hydroxychok'stero/ on the labelling of ubiquinone dolichol and cholesterol with / 3H/me~'akmh, acid in Molt-4 leukemic lymphocytes
Addition" Incorporation of [~H]mevalonic acid into specific lipids (dpm/I() ~' cells)
ubiquinone dolichol cholesterol
25-Hydroxy- cholesterol 2542 2336 12979
Control 5570 11370 4174 i
" The cells were incubated with or without 25-hydroxycholesterol (2 #g/ml) f o r 2 h before the addition of 10 btCi of [3H]mevalonate (37, t) Ci/mnml) and then incubation continued for 6 h.
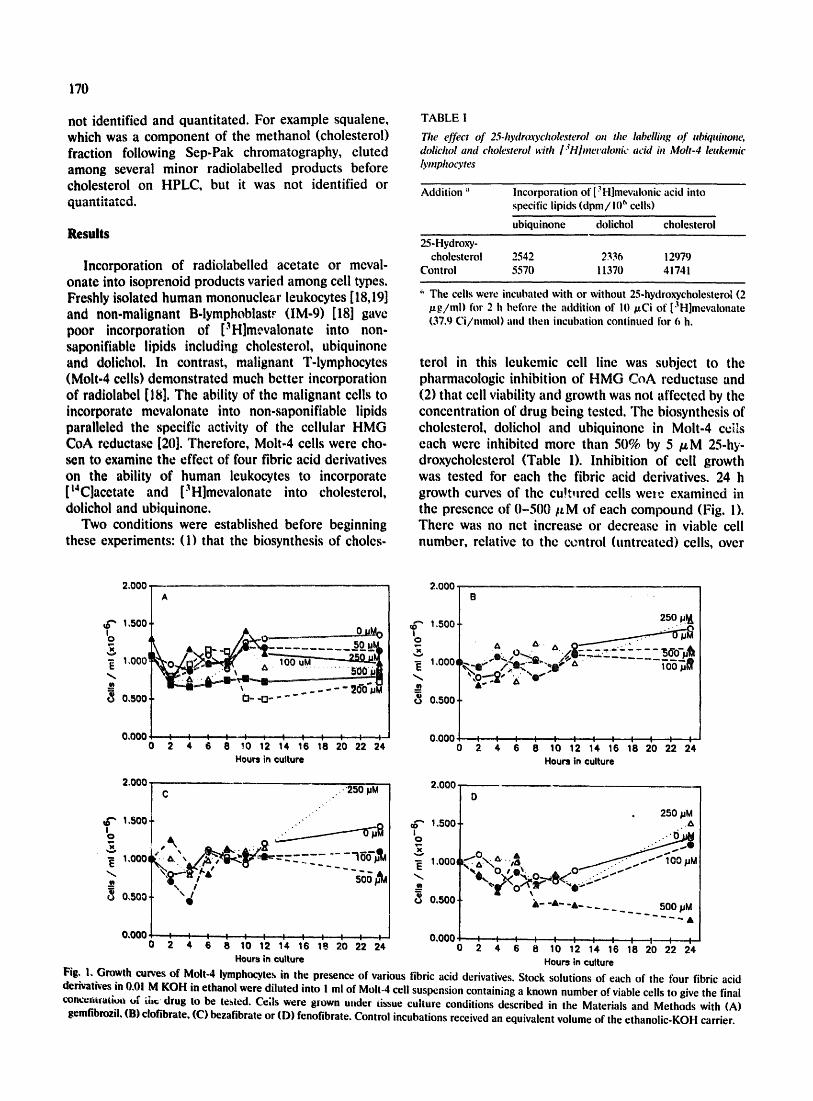
terol in this leukemic cell line was subject to the pharmacologic inhibition of HMG CoA reductase and (2) that cell viability and growth was not affected by the concentration of drug being tested. The biosynthesis of cholesterol, dolichol and ubiquinone in Molt-4 ceils each were inhibited more than 50% by 5 /~M 25-hy- droxycholesterol (Table 1). Inhibition of cell growth was tested for each the fibric acid derivatives. 24 h growth curves of the cu!tured cells we=e examined in the presence of 0 -500 / tM of each compound (Fig. 1). There was no net increase or decrease in viable cell number, relative to the centrol (untreated) cells, over
2,000,
1.500, I o
I,OOO
0,500,
. . . . . . . . . _5o~
0,000 -', : : , ,', =4 , : : o 2 , 6 o ;0 (= I'4 1'6 lh 20 22 24
Hours in culture
2,000.
1,500, I o
E 1,000!
IR
o,5oo,
0,000 : 0 2
B ' '
250 ~1~
• & &,. & ~ . . . ' _
,.. ~..O,.~z_ , w ~ . "7. . . . . . . . . . . ~ ' e / ; =~" ""O" " tb3~f ",o--:2-'., ". '"
', : " : ' : : I I I I ¢ 6 8 10 12 14 16 18 20 22 24
Hours in culture
2 000 ~ . • J C .,'250 pM
i~o 1,500 l ! , . , , ~ ~ ' ~ " " ' " ' "
' g ' ? . K . . . . . . . .
0 ,5oo~ "~
0000 ~ , . . . . . . . . . . ! 0 2 4 6 8 10 12 14 16 18 20 22 24
Hours in culture
2,000
1.500 I o
1.oo0~
m
o.5oo,
0,000 ~
0
D
• 250 ~M ,&
, . . 6 ' , . , . .
%-0 ,~ , "
A - -A - -A . . . . . 500 pM
2 4 6 8 10 12 14 16 18 20 22 24 Hours in culture
Fig. 1. Growth curves of Molt-4 lymphocyte,,, in the presence of various fibric acid derivatives. Stock solutions of each of the four fibric acid dedvatb-es in 0,01 M KOH in ethanol were diluted into ! ml of Molt-4 cell suspension containing a known number of viable cells to give the final conc~m~afiou uF ;,h,~ drug to be tested. Ce;Is were grown under tissue culture conditions described in the Materials and Methods with (A)
gemfibrozil, (B) clofibrate, (C) bezafibrate or (D) fenofibrate. Control incubations received an equivalent volume of the ethanolic-KOH carrier.
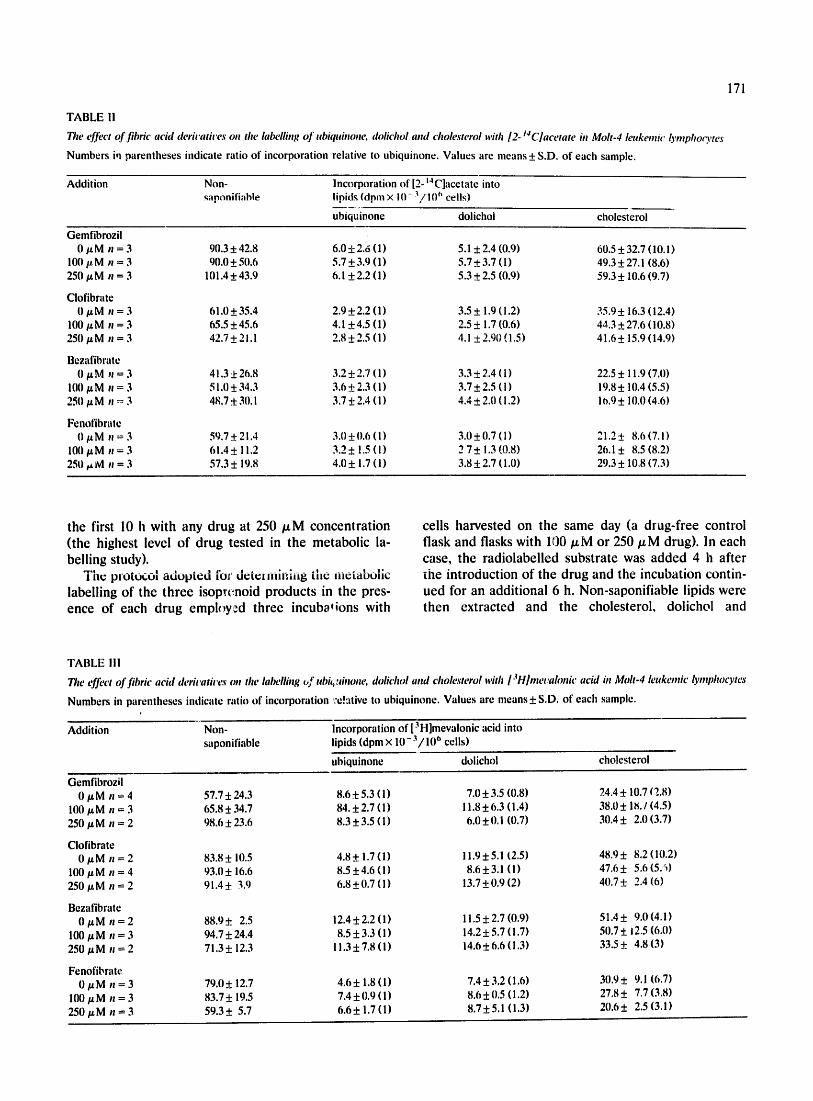
TABLE 11
The effect of fibric acid dericatices on the labelling of ubiquinone, dolichol and cholesterol with/2- t4f.Jacetate in Molt-4 leukemw lymphocytes
Numbers in parentheses indicate ratio of incorporation relative to ubiquinone. Values are means + S.D. of each sample.
171
Addition Non- .~ap~mifiahle
Incorporation of [2-t4C]acetate into lipids {dpm x 10- 3/10~ cells)
ubiquinone dolichol cholesterol
Gemfibrozil 0 ttM n = 3 90.3+42.8 6.0+2.6 (11 5.1 ±2.4 (0.9) 60.5+32.7 (10.1)
100 #M n = 3 90.0+50.6 5.7-+3.9 (11 5.7-+3.7 (1) 49.3±27.1 (8.6) 250 gM n = 3 101.4±43.9 6.1 -+2.2 (11 5.3+2.5 (0.9) 59.3_+ 10.6 (9.7)
CIofibrate 0 gM n = 3 61.0±35.4 2.9+2.2 (11 3.5+ 1.9 (1.2) 35.9+ 16.3 (12.41
100/.tM n = 3 65.5+45.6 4.1 +4.5 (i) 2.5± 1.7 (0.6) 44.3_+27.6 (10.8) 250/z M n = 3 42.7 ± 2 !. I 2.8 ± 2.5 ( I ) 4.1 + 2.90 ( 1.5 ) 41.6 5-15.9 (14.9)
Bezafihrate 0 #M ,: = 3 41.3+26.8 3.25:2.7 (I) 3.3:1:2.4 (1) 22.5± 11.9 (7.11)
I00/zM n = 3 51.0± 34.3 3.6± 2.3 (I) 3.7± 2.5 (I) 19,8± 10.4 (5.5) 8 2511 # M n =- 3 4 , 7 ± 311. I 3,7 ± 2.4 ( 1 ) 4.4 ± 2.0 ( i. 2 ) i 0, 9 + ! 0.0 ( 4.61
Fenofibratc 11 ~tM n = 3 59.7+21.4 3,0±0.6 (I) 3,0+0.7(11 21.2± 8,6(7.11
10(I #M n = 3 61.4+ !1.2 3.2+ t.5 (1) 2 7+ 1.3 (0.8) 26.1 + 8.5 (8.2) 250 ~M n = 3 57.3± 19.8 4.0± 1.7 (i) 3.8±2.7 (1.0) 29.3+ 10.8 (7.3)
the first 10 h with any drug at 250 #M concentration (the highest level of drug tested in the metabolic la- belling study)•
T h e p r o t o c o l a u o p t e u . . . . . . . . . for' ue tUl l l l i i l i l ig ~' . . . . . . ' - : . . . . . . tllC lll~t~l.OOllld' " '"
labelling of the three isoprcnoid products in the pres- ence of each drug empl~:yed three incubations with
cells harvested on the same day (a drug-free control flask and flasks with 100 tzM or 250 #M drug). In each case, the radiolabelled substrate was added 4 h after the introduction of the drug and the incubation contin- ued for an additional 6 h. Non-saponifiable iipids were then extracted and the cholesterol, dolichol and
TABLE Iil
The effect of fibric acid deriratires on the labelling ,,f ubic, inone, dolichol and cholesterol with ['*H/mecalonic acid in Molt-4 leukemic lymphocytes
Numbers in parentheses indicate ratio of incorporation ,re!at(re to ubiquinone. Values are means ± S.D. of each sample.
Addition Non- saponifiable
Incorporation of ['aH]mevalonic acid into lipids (dpm x 10-3/10o cells)
ubiquinone dolichol cholesterol
Gemfibrozil 0/,t M n = 4 57.7 ± 24.3 8.6 + 5.3 (1) 7.0 + 3.5 (0.8) 24.4 + 10.7 t2.8)
100 #M n = 3 65.8+34.7 84.±2.7 (1) 11.8±6.3 (!.4) 38.0± I8./(4.5) 250 ~ M n = 2 98.6 ± 23.6 8.3 + 3.5 ( l ) 6.0 ± 0. I (0.7) 30.4 ± 2.0 (3.7)
CIofibrate 0 #M n = 2 83.8± 10.5 4.8+5:1.7 (1) 11.9±5.1 t2.5) 48.9+ 8.2 (10.2)
100 #M n = 4 93.0+ 16.6 8 .5+4.6(I) 8.6±3.1 (I) 47.6± 5.6 (5/~) 250 #M n = 2 91.45- 3.9 6.8+0.7 (1) 13.7_+0.9 (2) 40.7± 2.4 (6)
Bezafibrate 0 #M n = 2 88.9+ 2.5 12.4±2.2(11 11.5±2.7 (0.9) 51.4+ 9.0(4.11
100 p,M n = 3 94.7:t:24.4 8.5±3.3 (1) 14.2+5.7 (1.71 50.7.+ i2.5 (6.0) 250/zM n = 2 71.3± 12.3 11.3±7.8(I) 14.6+6.6(1.3) 33.5+ 4.8(3)
Fenofibrate 0 #M n = 3 79.0_+ 12.7 4.6± 1.8 (1) 7.4±3.2 (1.61 30.9-+ 9.1 (6.7)
100 #M n = 3 83.7+ 19.5 7.4+(I.9 ( i) 8.6:t:0.5 (1.21 27.8:1:7.7 (3.8) 250 g,M n = 3 59.3+ 5.7 6.6+ 1.7 (1) 8.7+5.1 (1.3) 2(I.6± 2.5 (3.1)

172
ubiquinone were separated and analyzed for radioac- tivity. Although the absolute degree of incorporation varied among experiments, the ratios of incorporation of radiolabel into ubiquinone, dolichol and cholesterol were relatively constant from one day to another. Con- current incubations and analyses of a control and two different concentrations of the same drug with the same cell cultures reduced experimental variation.
The effect of the fibric acid derivatives on the con- version of [2-14C]acetate and [3H]mevalonate to cholesterol, dolichol and ubiquinone are illustrated, respectively, in Tables II and Ill. None of the drugs showed significant effects on the flux of [2-~4C]acetate or [aH]mevalonate into ubiquinone, dolichol and cholesterol at either l t~ /~M or 250 ~M concentra- tions when compared to control values.
Discussion
The present study was initiated to determine if the fibrates affect the metabolic flux of carbon through branched pathways of isoprenoid biosynthesis leading to cholesterol, ubiquinone and dolichol. Since much of the previous work designed to address this question utilized non-mammalian tissue and employed feeding studies which may have measured indirect effects of these drugs or their metabolites, our aim was to study the direct effects of these drugs on a cultured human cell line. Cholesterol synthesis in mononuclear leuko- cytes has been shown to be under similar control mechanisms in both rats and humans [21-23]. There- fore, we studied the effect of four fibric acid deriva- tives oi~ isoprene metabolism using acetate and meval- onic acid as metabolic precursors in lympho~tes. The use of mevalonate in particular permitted the localiza- tion of any effects to events beyond the HMG CoA reductase-catalyzed step,
A malignant lymphocyte cell line was studied be- cause HMG CoA reductase activity and cholesterol synthesis are high in such cells [18,20]. In non-malig- nant cells the rate of sterol synthesis is under negative feedback control by the cellular cholesterol concentra- tion. Although this mechanism of regulation appears to be defective or absent in malignant cells [24-26], HMG CoA reductase activity in leukemic guinea pig iympho- cytes [27] and cholesterol synthesis in cultured human leukemic T-iymphoojtes (as shown here) are both sup- pressed by the classical sterol biosynthesis inhibitor, 25-hydro~cholesterol.
The fibric acid derivatives used in this investigation had no significant effect on rates of cholesterol, dolichol or ubiquinone synthesis from [5-3H]mevalonate or [2- 14C]acetate. Therefore, we conclude that the fibric acid derivatives did not significantly affect any of the en- zl~nes in the branched metabolic pathways leading from acetate to these three isoprenoids in malignant
lymphocytes. These results are consistent with and supported by the earlier observations of McNamara et al. [22] who showed that mononuclear cells from pa- tients receiving clofibrate did no'~: show differences in the rates of cholesterol synthesis from [2-t4C]acetate compared to control cells.
Our results contrast with other observations drawn from both in vivo and in vitro studies with different experiraental designs. HMG CoA reductase activity was inhibited in mononuclear cells taken from Type Ila and Type lib hyperlipoproteinemic patients treated with fenofibrate [13]. Furthermore, in vivo hepatic sterol synthesis from [1-~4C]octanoate was decreased up to 75% following the feeding of seven daily doses of ¢lofibrate or bezafibrate to chow-fed rats [14]. Con- versely, gemfibrozil increased sterol synthesis up to 4-fold. None of the fibrates had ~ significant effect on [2-~4C]mevalonate incorporation into sterols. In con- trast, cholesterol synthesis from [3H]mevalenic acid was stimulated more than 6-fold in rat liver peroxi- somes and 1.5-foid in microsomes obtained from ani- mals fed a diet contab dng clofibrate [28,29]. Appelkvist also reported that, whereas dolichol synthesis from [3H]mevalonic acid was inhibited 50% in rat liver pe~r- oxisomes isolated from animals fed a diet containing clofibrate, it was stimulated over 3-fold in rat liver microsomes [30].
Blasi et al. [15], studied the in vitro effect of bezafi- brate on the synthesis of nunsaponifiable lipids and cholesterol in mononuclear lcukocytcs from normal subjects and patients with heterozygous familial hyper- cholesterolemia and observed effects consistent with the inhibitory in rive results of Maxwell et al. [14]. At bezafibrate concentrations higher than 28/zM, choles- terol biosynthesis from acetate or HMG CoA was inhibited 40-50%. The drug had no effect on choles- terol synthesis from mevalonate at these concentra- tions. Thus, although some consistency exists among reports of the effect of fibrates on sterol biosynthesis and HMG CoA reductase activity, there are also con- flicting results among different laboratories. The stimu- latory or inhibitory effects seen by others contrast with er r results which showed no effect of the fibrates in cultured human lymphocytes.
Several reasons could account for these differences. One explanation lies in the time of exposure to the drug. Our results were obtained after a short term (4 h) exposure of human cells to the fibrates, whereas the other studies involved drug treatment for 20 h to several days. For example, Blasi et al. [15] incubated mononuclear leukocytes with the drug for 20 h before incubation with radiolabelled precursor. Longer incu- bations would permit drug metabolism to a more phar- macologically active compound. However, there is no evidence at this time that the fibrates exhibit their inhibitory effects through biotransformation.

Acknowledgements
Support for this work was furnished by Parke-Davis Pharmaceuticals, the American Cancer Society (ACS 88073A), the National Institutes of Health (DK 40439 and RR 00082-29). We thank Ms. Carolyn Miller and Mrs. Marissa Otal for excellent technical assistance.
References
1 Goldstein, J.L. and Brown, M.S. (19901 Nature 343, 425-430. 2 Edwards, P.A., Popjak, G., Fogelman, A.M. and Edmond, J.
(19771 J. Biol. Chem. 252, 1057-1063. 3 Reynolds, G.A, Goldstein, J.L. and Brown, M.S. (1985) J. Biol.
Chem. 260, 10369-10377. 4 RodweU, V.W., Nordstrom, J. and Mitschelen, J.J. (1976) Adv.
Lipid Res. 14, 1-74. 5 Ashby, M.N. and Edwards, P.A. (19891 J. Biol. Chem. 264,
635 -640, Havel, C., Hanbury, E., Scallen, T.J. and Watson, J.A. (19791 J. Biol. Chem. 254, 9573-0582.
7 Faust, J. and Krieger, M. (1987) J. Biol. Chem. 262, 1996-2004. 8 Keller, R.K. (19861 J. Biol. Chem. 261, 12(153-12059. 9 Nambudiri, A.M.D., Ranganathan, S. and Rudney, H. (1980) J.
Biol. Chem. 255, 5894-5899. 10 Grundy, S.M., Ahrens, Jr., E.H., Salen, G., Schreibman, P.H. and
Vestel, P.J. (19721 J. Lipid Res. 13, 531-551. !1 Rodney, G., Uhlendorf, P. and Maxwell, R.E. (19761 Proc. Roy.
Soc. Med. 69 (Suppl. 2), 6-10. 12 Newton, R.S. and Krause, B.R. (1986) In Pharmacological Con-
173
trol of Hyperlipidemia (Fears, R., ed.), pp. 171-186, J.R. Prous Science Publishers.
13 Schneider, A., Stange, E.F., Ditschuneit, H.tt. and Ditschuneit. H. (1985) Atherosclerosis 56, 257-262.
14 Maxwell, R.E., Nawrocki, J.W. and Uhlendorf, P.D. (19831 Atherosclerosis 48, 195-203.
15 Blasi, F., Sommariva, D., Cosentine, R., Cavaiani, B. and Fasoli, A. (19891 Pharm. Res. 21,247-254.
16 Shive, W. and Matthews, K.S. (19881 Annu. Rev. Nutr. 5, 81-97. 17 Palmer, D.N., Anderson, M.A. and Jolly, R.D. (19841 Anal.
Biochem. 140, 315-319. 18 Henry, A., Stacpoole, P.W. and Allen, C.M. (1991) Biochem. J.
278, 741-747. 19 Burns, C.P., Welshman, I.R., Scallen, T.J. and Spector, A.A.
(19821. Biochim. Biophys. Acta 713, 519-528. 20 Harwood, H.J., Jr., Alvarez, I.M., Noyes, W.D. and Stacpoole,
P.W. (19911 J. Lipid Res. 32, 1237-1252. 21 Young, N.L. and Rodw~.JI, V.W. (1977) J. Lipid Res. 18, 572-581. 22 McNamara, D.J., Davidson, N.O. and Fernandez, S. (19801 J.
Lipid Res. 21, 65-71. 23 Fogelman, A.M., Seager, M.J., ttokom, M. and Edwards, P.A.
(19791 J. Lipid Res. 20, 379-388. 24 Gregg, R.G., Davidson, M. and Wilce, P.A. (19861 Int. J. Biocllem.
18, 389-393. 25 Siperstein, M.D., Gyde, A.M. and Morris, H.P. (1971) Proc. Natl.
Acad. Sci. USA 68, 315-317. 26 Azrolan, N.I. and Coleman, P.S. (1989) Biochem. J. 258, 421-425. 27 Tabacik, C., Alain, S. and Sainte-Marie, J. (1987) Biochim. Bio-
phys. Acta 921,405-410. 28 Appeikvist, E-L. and Dallner, G. (1987) In Pharmacology (Rand,
M.J. and Raper, C., eds.), pp. 857-860, Elsevier, Amsterdam. 29 Applekvist, E-L. (1987) Biosci. Rep. 7, 853-858. 30 Appelkvist, E-L. (1987) Acta Chem. Scand. B 41, 73-75.

![SingleNucleotidePolymorphismsinSelected ...benzo[a]pyrene-7,8-9,10-diol epoxide (BPDE) in cultured primary lymphocytes from 172 cancer-free subjects. We found that We found that among](https://img.pdfslide.us/doc/110x75/608d368227679f70f265b55b/singlenucleotidepolymorphismsinselected-benzoapyrene-78-910-diol-epoxide.jpg)










![Role of heme Oxygenase-1 in low dose Radioadaptive response · 2016. 3. 11. · human lymphocytes were pre-cultured with [3H]thymidine, which acted as a source of low-level chronic](https://img.pdfslide.us/doc/110x75/60e49a5ea41e4060bc41625e/role-of-heme-oxygenase-1-in-low-dose-radioadaptive-2016-3-11-human-lymphocytes.jpg)
















