Embed Size (px)
Citation preview

Neural Pathways for Bilateral VocalControl in Songbirds
J. MARTIN WILD,1* MATTHEW N. WILLIAMS,1AND RODERICK A. SUTHERS2
1Department of Anatomy, School of Medicine and Health Science, University of Auckland,Auckland 92019, New Zealand
2Section of Physiology, Medical Sciences, Indiana University, Bloomington, Indiana 47405
ABSTRACTIpsilateral and contralateral projections of nucleus robustus archistriatalis (RA), a tel-
encephalic vocal premotor nucleus, to respiratory-vocal nuclei in the brainstem were definedin adult male Wasserschlager canaries, grey catbirds, and zebra finches, three songbirdspecies that appear to differ in the degree of lateralized syringeal dominance. In all threespecies, ipsilateral projections of RA to the medulla included the tracheosyringeal part of thehypoglossal nucleus (XIIts), that innervates the syrinx, the bird’s vocal organ, the suprahy-poglossal area (SH), and two respiratory-related nuclei, retroambigualis (RAm) and param-bigualis (PAm; Reinke and Wild [1998] J Comp Neurol 391:147–163). Projections of RA to thecontralateral XIIts, SH and RAm, were substantial in canaries, which use the left side of thesyrinx predominantly during singing; less pronounced in catbirds, which have no lateraldominance for song control; and least pronounced in zebra finches, in which there is aright-sided dominance for song control. There were no obvious differences in the number ofcrossed projections in birds injected in the left or right RA. Local sources of inputs to XIItsand RAm were defined anatomically in zebra finches and canaries. RAm, including neuronsin close proximity to XIIts, was found to project to XIIts and the suprahypoglossal area bilaterallybut predominantly ipsilaterally. RAm also had reciprocal connections with its contralateralhomologue. These results suggest a pattern of connections between premotor and motorrespiratory-vocal nuclei that may be involved in bilateral control of vocal output at medullarylevels, a control that involves a high degree of coordination between vocal and respiratorystructures on both sides of the body. J. Comp. Neurol. 423:413–426, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: nucleus robustus; nucleus retroambigualis; zebra finch; canary; catbird
Song in songbirds, as in humans, is produced by thecoordinated action of vocal, supravocal, and respiratorymuscles (Wild, 1997). Unlike the larynx of humans, how-ever, the sound source of songbirds is a duplicated struc-ture, with the muscles of each syringeal half receivingindependent neural control from the ipsilateral hypoglos-sal nucleus. This configuration allows songbirds to pro-duce song components using the two sides of the syrinx ineither sequential or simultaneous fashion, depending onthe species (Suthers, 1997). An early finding in canary,chaffinch, white-throated sparrow, white-crowned spar-row, and java sparrow was that song production is asym-metric, in the sense that the majority of syllables in thesongs of these species is produced using the left side of thesyrinx, as usually measured by the number of syllableslost or retained following unilateral syringeal denervation(Nottebohm, 1970, 1971, 1972, 1977; Lemon, 1973; Notte-bohm and Nottebohm, 1976; Nottebohm et al., 1976;Seller, 1979; Hartley and Suthers, 1989; Suthers, 1992).
In the zebra finch, however, it was later found that syrin-geal denervation did not affect the number of syllablesretained postoperatively so much as their acoustic struc-ture, this being more affected following denervation of theright than of the left side of the syrinx (Williams et al.,1992; Floody and Arnold, 1997). Furthermore, with graycatbirds and brown thrashers, Suthers (1990) and Gollerand Suthers (1996a) found that neither side of the syrinxwas dominant for syllabic production. In these mimicthrushes, different notes or syllables can be produced ei-ther on the left side or on the right side of the syrinxsimultaneously, thus producing two harmonically unre-
*Correspondence to: Dr. J.M. Wild, Department of Anatomy, School ofMedicine and Health Science, University of Auckland, Private Bag, Auck-land 92019, New Zealand. E-mail: [email protected]
Received 2 December 1999; Revised 15 March 2000; Accepted 23 March2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 423:413–426 (2000)
© 2000 WILEY-LISS, INC.

lated voices, or on different sides in sequential fashion, thebird rapidly switching back and forth from one side to theother, even within the short time constraints of a singlesyllable. Frequently in these birds, and in northern cardi-nals, the left and right sides of the syrinx coordinate toproduce a frequency-modulated syllable, in which lowerfrequency components are produced on the left side of thesyrinx and higher frequency components on the right(Nottebohm, 1977; Allan and Suthers, 1994; Suthers andGoller, 1996; reviewed by Suthers, 1997, and by Suthersand Goller, 1997).
In mimic thrushes and brown-headed cowbirds, the gat-ing of phonation and the determination of which side ofthe syrinx produces sound at any particular moment isaccomplished by the dorsal syringeal muscles, the mo-toneurons of which are concentrated caudally in XIIts(Vicario and Nottebohm, 1988; Suthers et al., 1994; Ruanand Suthers, 1996; Goller and Suthers, 1996b; Goller andLarsen, 1997). The control of fundamental frequency, onthe other hand, appears to be by the action of ventralsyringeal muscles, the motoneurons of which are concen-trated rostrally in XIIts (Vicario and Nottebohm, 1988;Ruan and Suthers, 1996). The ventral syringeal muscles,like the expiratory muscles that provide the pressurehead, are active on both sides, irrespective of which side ofthe syrinx is being used to produce the sound (Goller andSuthers, 1996a,b). This seems to imply that fundamentalfrequency is controlled by the same motor program beingsent to both sides of rostral XIIts and that phonation usingone side of the syrinx is controlled by different motorprograms being sent to the two sides of caudal XIIts.During two-voiced syllables, however, in which air is flow-ing through both sides of the syrinx, either the same motorprogram or different motor programs could be sent to thetwo sides of caudal XIIts.
However, whether in any particular species there is or isnot a lateral dominance for song production, the questionarises of how the bird coordinates the left and right sidesof the syrinx and respiratory apparatus to produce thespecies-typical song pattern, which includes syllables pro-duced on both the left and the right sides. This question isall the more pertinent in the context of the ipsilateralinnervation of each syringeal half by the hypoglossal mo-tor nucleus (XIIts) and nerve (ts) in songbirds (Nottebohmand Nottebohm 1976; Nottebohm, 1977; Vicario and Not-tebohm, 1988), the absence of any known morphologicaldifferences between the song control nuclei on the left andright sides of the brain rostral to the hypoglossal nucleusitself (DeVoogd and Nottebohm, 1981; Nottebohm et al.,1982; DeVoogd et al., 1991), and the absence of any cal-losal connections between song control nuclei on oppositesides of the brain. However, several neural pathways haverecently been described that may function to transfer in-formation derived from the vocal control circuitry in onecerebral hemisphere, or on one side of the brainstem, tothat in the other, in a feedback manner (Vates et al., 1997;Striedter and Vu, 1997; Reinke and Wild, 1998). One ofthese, which originates in the rostroventrolateral medullaand projects bilaterally upon the thalamic nucleus uvae-formis (Uva; Okuhata and Nottebohm, 1992; Striedterand Vu, 1997; Reinke and Wild, 1998), may be particularlyconcerned with the coordination of the outputs of the leftand right premotor vocal control nuclei in the two hemi-spheres, so that the temporal sequencing of syllables fol-lows the learned, species-typical pattern (Williams andVicario, 1993; Coleman et al., 1999; see also Vu et al.,1994, 1996). It has not yet been established, however,whether this pathway actually feeds back informationthat is derived directly from descending motor commands.
Abbreviations
A archistriatumAi archistriatum intermediumCb cerebellumCbd tractus spinocerebellaris dorsaliscc canalis centralisCI commissura infimaCu nucleus cuneatusDIP nucleus dorsalis intermedius posterior thalamiDLL nucleus dorsolateralis anterior thalami, pars lateralisDM dorsomedial nucleus of the intercollicular complexDMP nucleus dorsomedialis posterior thalamiFA tractus frontoarchistriatalisFLM fasciculus longitudinalis medialisFPL fasciculus prosencephali lateralisGCt substantia grisea centralisHIP tractus habenulointerpeduncularisHL nucleus habenularis lateralisHV hyperstriatum ventraleICo nucleus intercollicularisIOS nucleus infraolivaris superiorIpc nucleis isthmi, pars parvocellularisLa nucleus laminarisLAD lamina archistriatalis dorsalisMLd nucleus mesencephalicus lateralis, pars dorsalisMV nucleus motorius nervi trigeminiN neostriatumnBOR nucleus of the basal optic rootNC neostriatum caudaleNIII nervus oculomotoriusNVIII nervus octavusOI nucleus olivaris inferior
OM tractus occipitomesencephalicusOS nucleus olivaris superiorPA paleostriatum augmentatumPAm nucleus parambigualis (Reinke and Wild, 1998)PM nucleus pontis medialisPP paleostriatum primitivumPrV nucleus sensorius principalis nervi trigeminiRA nucleus robustus archistriatalisRAm nucleus retroambigualis (Wild, 1993a)Rt nucleus rotundusRu nucleus ruberRVL ventrolateral nucleus of the rostral medualla (Wild,
1993b)SCE stratum cellulare externumSGC stratum griseum centraleSGF stratum griseum et fibrosum superficialeSH suprahypoglossal area (Wild, 1993b)SHL nucleus subhabenularis lateralisSpM nucleus spiriformis medialisSSp nucleus supraspinalisTn nucleus taeniaeTPc nucleus tegmenti pedunculopontinus, pars compactaTTD nucleus et tractus descendens nervi trigeminiUva nucleus uvaeformisv ventriculusVc nucleus descendens nervi trigemini, pars caudalisVI nucleus nervi abducentisVIId nucleus nervi facialis, pars dorsalisVIIv nucleus nervi facialis, pars ventralisX nucleus motorius dorsalis nervi vagiXIIts nucleus nervi hypoglossi, pars tracheosyringealis
414 J.M. WILD ET AL.

An alternative to contralateral feedback control of vocaloutput is contralateral feedforward control, by which mo-tor commands from RA are routed to brainstem nuclei onthe opposite side. In zebra finches and green finches, RAhas a minor contralateral projection directly upon XIItsand RAm, but it also has more numerous indirect projec-tions, by way of a largely ipsilateral projection upon thepontine nucleus infraolivaris superior (IOS) and the ven-trolateral nucleus of the rostral medulla (RVL), whichthen project upon the vocal motoneurons and respiratorypremotor neurons bilaterally (Wild, 1993a,b, 1994; Wild etal., 1997; Reinke and Wild, 1997, 1998).
In the present study, we have again focussed our atten-tion on the descending projections of RA, with a view todetermining whether there was an obvious difference inthe presence or extent of contralateral projections to XIItsand RAm between species that appear to vary in thedegree of lateralized syringeal dominance. Thus, a com-parison was made between the descending projections ofRA in male zebra finches (Taeniopygia guttata; Wild,1993b; Vicario, 1993), which are said to have a right-sideddominance for song production (Price, 1977; Williams etal., 1992); male Wasserschlager canaries (Serinus cana-ria), which are strongly left-side-dominant (Nottebohmand Nottebohm, 1976), and whose RA projections to thebrainstem, originally thought to be entirely ipsilateral,have not been reexamined in detail since the originallesion/silver degeneration study of Nottebohm et al.(1976); and grey catbirds (Dumatella carolinensis), a spe-cies of mimic thrush having no lateralized syringeal dom-inance (Goller and Suthers, 1996a,b) and whose RA pro-jections have not previously been charted. In addition,because RAm projects bilaterally to spinal motoneuronsthat innervate expiratory muscles (Wild, 1993a), and be-cause it has also been suggested to project upon XIIts,where it may impose an expiratory-related rhythm (Vi-cario, 1993; Wild, 1993a), a further objective of the presentstudy was to determine whether RAm projects upon XIItscontralaterally as well as ipsilaterally and whether it hasreciprocal projections with its contralateral homologue.Such bilateral and reciprocal projections of RAm could beinvolved in ensuring that syringeal and expiratory muscleactivities are temporally coordinated on both sides of thebody during phonation. These local projections were ex-amined in zebra finches and canaries.
MATERIALS AND METHODS
Surgical procedures were approved by and performedaccording to the guidelines of the Animal Ethics Commit-tee of the University of Auckland. Four male zebra finches(Taeniopygia guttata), four male Wasserschlager canaries(Serinus canaria), and four male grey catbirds (Dumatellacarolinensis) were each anesthetized either with isoflu-rane gas or with an intramuscular injection of ketamine(50 mg/kg) and xylazine (20 mg/kg) and the heads fixed ina stereotaxic apparatus with the beak tilted downward at45° to the horizontal. Nucleus RA was identified eitherusing the stereotaxic atlas of Stokes et al. (1974) and/orelectrophysiologically by recording multiunit activity withtungsten microelectrodes, the dorsal border of RA beingreadily identified by a marked increase in neuronal activ-ity and by the fact that, under ketamine/xylazine anesthe-sia, RA neurons have a characteristic irregular and“bursty” pattern of discharge. Injections of biotinylated
dextran amine [BDA; 10,000 mw; Molecular Probes, Eu-gene, OR; 10% in 0.1 M phosphate-buffered saline (PBS;pH 7.4); Veenman et al., 1992] were then made in RA viaglass micropipettes (outside diameter 30 mm) attached toa picospritzer (General Valve, Fairfield, NJ). BDA was thetracer of choice because of its proved ability to label the RApathway with clarity and completeness (Wild, 1993b). Twobirds of each species were injected on the left and two onthe right. The injections were deliberately large (30–40nl) in order to maximize the anterograde labeling of de-scending axons.
In two other male canaries, and in 10 more male zebrafinches, unilateral iontophoretic injections of BDA (4 mApositive current, 7 seconds on, 7 seconds off, for 10–20minutes) were made into either the left or the right XIItsor RAm using glass micropipettes, following identificationof the nuclei by a combination of stereotaxis and electro-physiological recording of expiratory-related unit activity(Wild, 1993a), using tungsten microelectrodes (FrederickHaer and Company, 3–5 MV). Again, BDA was the tracerof choice, because of the high reliability with which rela-tively small, iontophoretic injections could be made in thisconfined region of the caudal medulla and because of thedetail with which individual fibers could be traced to andfrom the injection site.
The birds were allowed to survive 4 or 5 days before theywere anesthetized with an intramuscular injection of ket-amine (50 mg/kg) and xylazine (20 mg/kg) and perfusedtranscardially with 50 ml of saline followed by 100 ml 4%paraformaldehyde in 0.1 M phosphate buffer, pH7.4. Thecalvaria were removed and the head placed in fixative for3–4 hours before blocking the brain in the transverseplane, extracting it from the skull, and postfixing it over-night. Following equilibration in PBS plus 30% sucrose for24 hours, the brains were sectioned on a feezing mic-rotome at 40 mm in the transverse plane, and the sectionswere collected in three series. These were treated with50% methanol and 1% H2O2 to enhance the penetration ofsecondary reagents and to quench endogenous peroxidaseactivity and then were incubated at room temperature for1.0–1.5 hours in streptavidin-peroxidase conjugate (Mo-lecular Probes) at 1:1,000 dilution in PBS, pH 7.4, con-taining 0.4% Triton X-100. Following washing in PBS, thesections were reacted with 0.025% diaminobenzidine inPBS and H2O2. Sections were mounted on subbed slides,and at least one series was counterstained with Giemsafor the identification of nuclear groups.
RESULTS
Ipsilateral projections resulting fromarchistriatal injections
As anticipated, none of the injections in RA was con-fined to the nucleus, there being spread of tracer to thearchistriatum surrounding RA and up the pipette trackinto the overlying caudal neostriatum (Fig. 1D). In nocase, however, was there a part of RA that appeared to befree of reaction product. The result of the extranuclearspread of deposited tracer was that, in addition to thediencephalic and mesencephalic terminal fields that areknown to originate from RA (e.g., certain dorsal thalamicnuclei and the dorsomedial nucleus (DM) of the intercol-licular complex (Nottebohm et al., 1976; Gurney, 1981;Wild, 1993b; Vates et al., 1997), there were terminal fields
415BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS
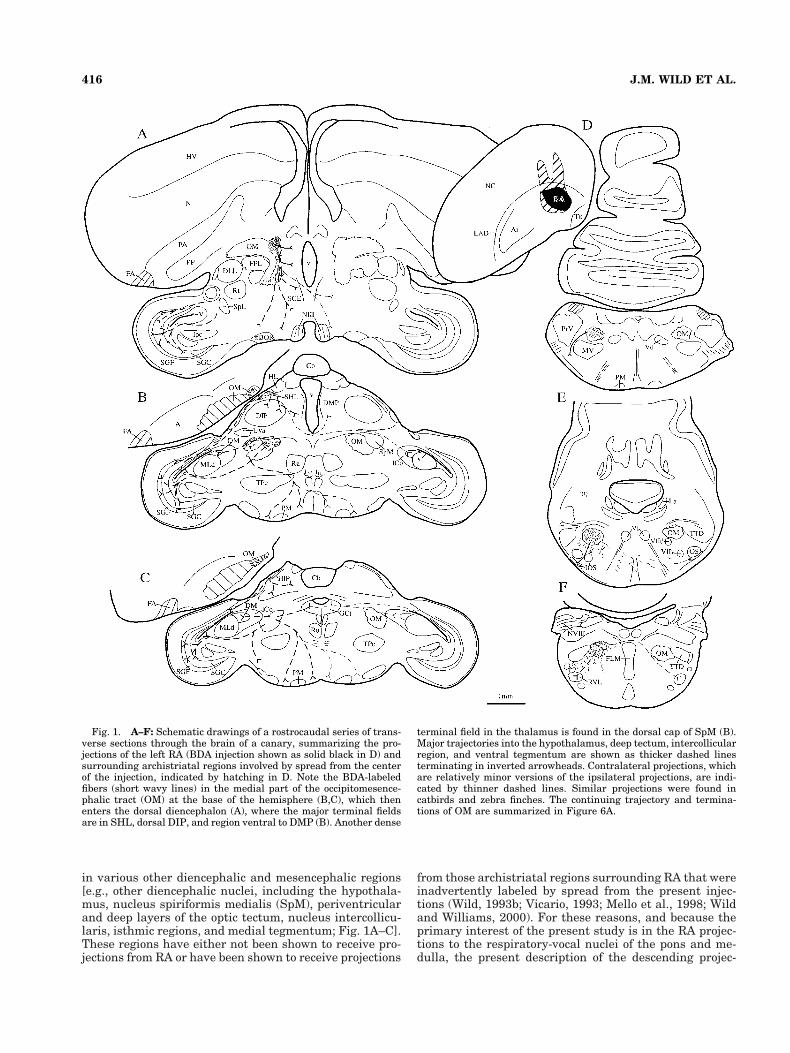
in various other diencephalic and mesencephalic regions[e.g., other diencephalic nuclei, including the hypothala-mus, nucleus spiriformis medialis (SpM), periventricularand deep layers of the optic tectum, nucleus intercollicu-laris, isthmic regions, and medial tegmentum; Fig. 1A–C].These regions have either not been shown to receive pro-jections from RA or have been shown to receive projections
from those archistriatal regions surrounding RA that wereinadvertently labeled by spread from the present injec-tions (Wild, 1993b; Vicario, 1993; Mello et al., 1998; Wildand Williams, 2000). For these reasons, and because theprimary interest of the present study is in the RA projec-tions to the respiratory-vocal nuclei of the pons and me-dulla, the present description of the descending projec-
Fig. 1. A–F: Schematic drawings of a rostrocaudal series of trans-verse sections through the brain of a canary, summarizing the pro-jections of the left RA (BDA injection shown as solid black in D) andsurrounding archistriatal regions involved by spread from the centerof the injection, indicated by hatching in D. Note the BDA-labeledfibers (short wavy lines) in the medial part of the occipitomesence-phalic tract (OM) at the base of the hemisphere (B,C), which thenenters the dorsal diencephalon (A), where the major terminal fieldsare in SHL, dorsal DIP, and region ventral to DMP (B). Another dense
terminal field in the thalamus is found in the dorsal cap of SpM (B).Major trajectories into the hypothalamus, deep tectum, intercollicularregion, and ventral tegmentum are shown as thicker dashed linesterminating in inverted arrowheads. Contralateral projections, whichare relatively minor versions of the ipsilateral projections, are indi-cated by thinner dashed lines. Similar projections were found incatbirds and zebra finches. The continuing trajectory and termina-tions of OM are summarized in Figure 6A.
416 J.M. WILD ET AL.

tions is largely limited to these levels, where, for the mostpart, the trajectory follows that previously described spe-cifically for the RA pathway in zebra finches and greenfinches, excepting contralateral terminations (Vicario,1993; Wild, 1993b).
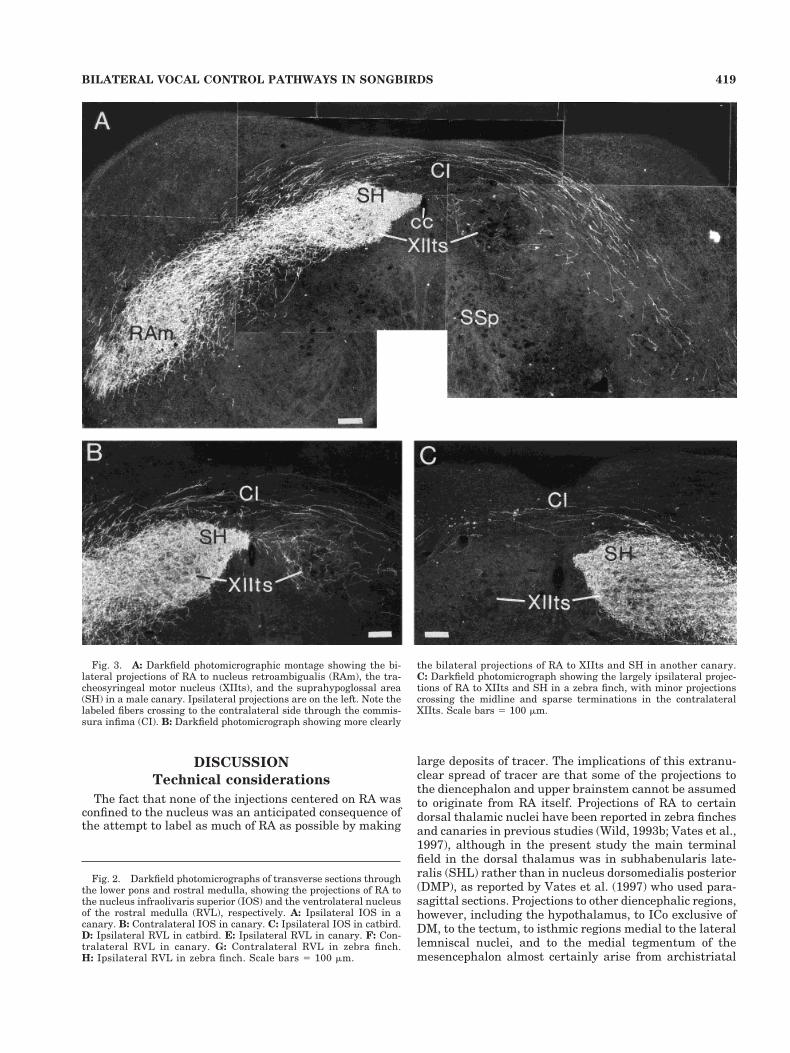
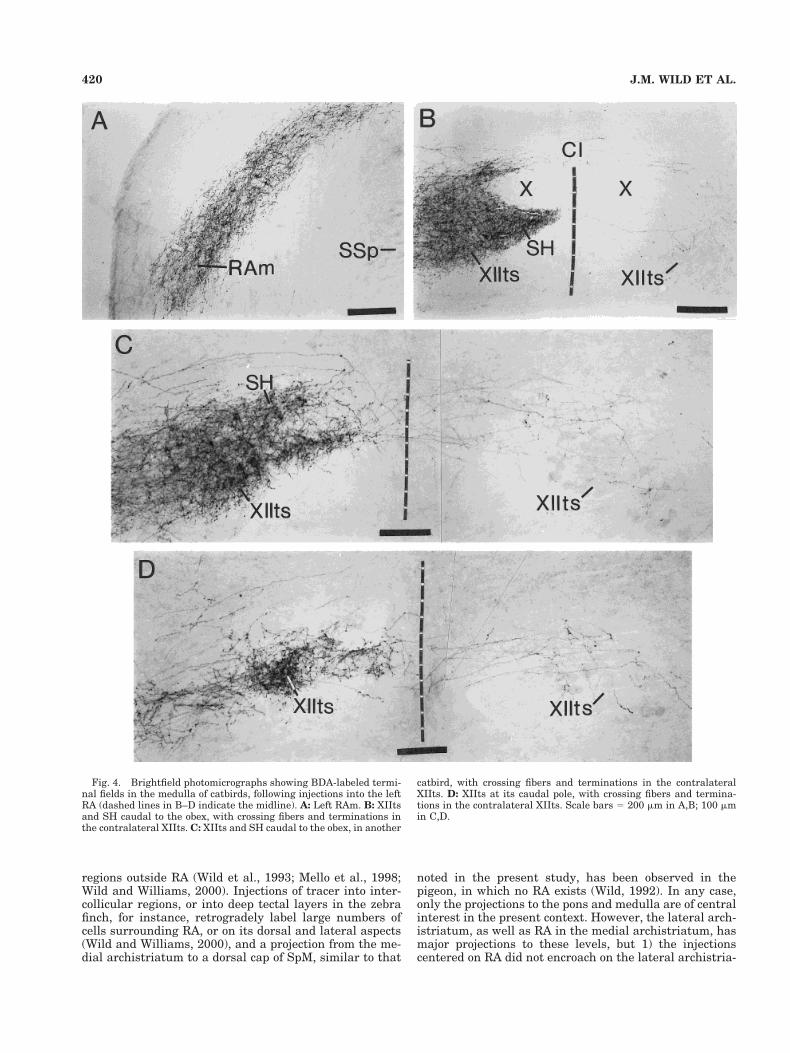
In each case in each of the three species, labeled fiberscoursed rostrally from the injection site, collected in themedial part of the occipitomesencephalic tract (OM) at thebase of the hemisphere (Fig. 1A–C), and passed throughthe dorsal diencephalon and into the brainstem. En route,terminal fields were primarily in the nucleus subhabenu-laris lateralis (SHL), dorsal cap of nucleus spiriformismedialis (SpM), deep and periventricular layers of theoptic tectum, and DM of the intercollicular complex (Fig.1B,C). In the pons, fibers left OM to pass ventrally aroundand through the subnuclei of the trigeminal motor com-plex (Fig. 1D) and terminated in the nucleus infraolivarissuperior (IOS; Figs. 1E, 2A,C) and, more caudally, in theventrolateral nucleus of the rostral medulla (RVL; Figs.1F, 2D,E,H). In all three species at middle and caudallevels of the medulla, fibers left OM medially to terminatewithin XIIts and the suprahypoglossal area (SH), a rela-tively cell-poor region that is most evident dorsal andmedial to XIIts (Figs. 3A–C, 4B–D). In catbirds, the ter-minal field in and around XIIts caudal to the obex formeda characteristic pattern, in which the terminations in SHand the fibers leaving OM to cross to the opposite sidewere closely applied to the lateral aspect of the caudal poleof the dorsal motor nucleus of the vagus (NX), grasping itin pincer-like fashion (Fig. 4B). Fibers also left OM ven-trolaterally to terminate throughout the arc that extendsfrom XIIts and the suprahypoglossal area to the ventro-lateral periphery of the medulla (Figs. 3A, 4A). This arcincludes the inspiratory premotor nucleus parambigualis(PAm) at periobex levels and the expiratory premotornucleus retroambigualis more caudally, as far as the spi-nomedullary junction (Wild, 1993a; Reinke and Wild,1998).
Contralateral projections resulting fromarchistriatal injections
In the diencephalon and mesencephalon, contralateralprojections were a minor version of the ipsilateral projec-tions, although projections to the contralateral tectumwere limited to periventricular layers (Fig. 1A–C). In thepons and medulla of canaries, and to a lesser extent inzebra finches, but not in catbirds, a few fibers were seen toextend into the contralateral pontine tegmentum, wherethey passed ventrally and caudally to terminate sparselyin IOS and RVL (Figs. 1E,F, 2B,F,G). More caudally in themedulla of canaries, a substantial contingent of fibers leftthe ipsilateral OM medially and crossed to the oppositeside, primarily through the commissura infima (Fig.3A,B). Some of these fibers extended into and terminatedthroughout the contralateral RAm and PAm, particularlythe former (Fig. 3A). Others terminated within the con-tralateral XIIts, and particularly within SH (Fig. 3A,B).This pattern of crossed projections was also evident incatbirds (Fig. 4B–D), but to a lesser extent than in canar-ies, and was least evident in zebra finches (Fig. 3C), inwhich only a few fibers and terminations were observed inthe contralateral XIIts and RAm, confirming previous ob-servations (Wild, 1993b).
Local sources of projections to XIItsand RAm
In canaries and zebra finches, the iontophoretic injec-tions of BDA that were centered on XIIts always includedparts of the relatively cell-poor region that surrounds XI-Its, including the suprahypoglossal area. These injectionsgave rise to a characteristic pattern of anterograde andretrograde labeling within the medulla. Invariably theinjections resulted in the labeling of fibers and termina-tions, but not cell bodies, in the contralateral XIIts (Fig.5A,F–H). In addition, when the injection was centered onXIIts caudal to the obex (see, e.g., Fig. 5F), there weremany labeled cell bodies, fibers, and terminations scat-tered throughout RAm bilaterally, but predominantly ip-silaterally (Fig. 5A–D). If, however, the injection was cen-tered on XIIts rostral to the obex (see, e.g., Fig. 5E), thelabeled cell bodies, fibers, and terminations tended to clus-ter in the ipsilateral nucleus parambigualis (Fig. 5E),which extends from the rostral pole of RAm caudal to theobex, to a position rostral to the obex (Reinke and Wild,1998).
Some of the nonmotoneurons that were labeled bilater-ally in the medulla as a result of injections centered onXIIts and its surroundings were situated in close proxim-ity to XIIts. These cells and their processes could moreeasily be identified on the contralateral side, where theywere not obscured by the fringes of the injection itself (Fig.5H). Occasionally, such cells were located between XIItsand the suprajacent dorsal motor nucleus of the vagus,i.e., within SH, but more frequently they were locatedlaterally adjacent to XIIts, from which their labeled pro-cesses could be followed into the adjacent XIIts itself (Fig.5H). Some of these processes were clearly dendrites, butothers could have been axons. Retrogradely labeled cellslocated at a greater distance from XIIts, e.g., those in moreventrolateral parts of RAm on both sides of the brain, senttheir axons across the midline through the central me-dulla and/or through the commissura infima to ramifywithin the opposite RAm and XIIts.
This pattern of bilateral projections of retrogradely la-beled neurons located in more or less close proximity toXIIts is consistent with a process of somatopetal and so-matofugal transport and was substantiated by the injec-tions in RAm. These gave rise to dense projections to theipsilateral XIIts and less dense, but nonetheless substan-tial, projections to the contralateral XIIts and RAm (Fig.6B–D). A summary diagram that is consistent with thelocal medullary circuitry delineated in the present studyand with the descending projections of RA is shown inFigure 6A.
The XIIts injections also retrogradely labeled cells inPAm, RVL, IOS, and dorsomedial nucleus of the inter-collicular complex (DM), predominantly ipsilaterally,and also supplied projections to these nuclei, the specificorigin of which has not been determined but which mayinclude cells in the region surrounding XIIts that wereincluded in the injection. The XIIts injections also ret-rogradely labeled substantial numbers of cells in all butthe dorsal part of the ipsilateral RA. Only in canarieswere a very few labeled cells observed in the contralat-eral RA.
417BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS
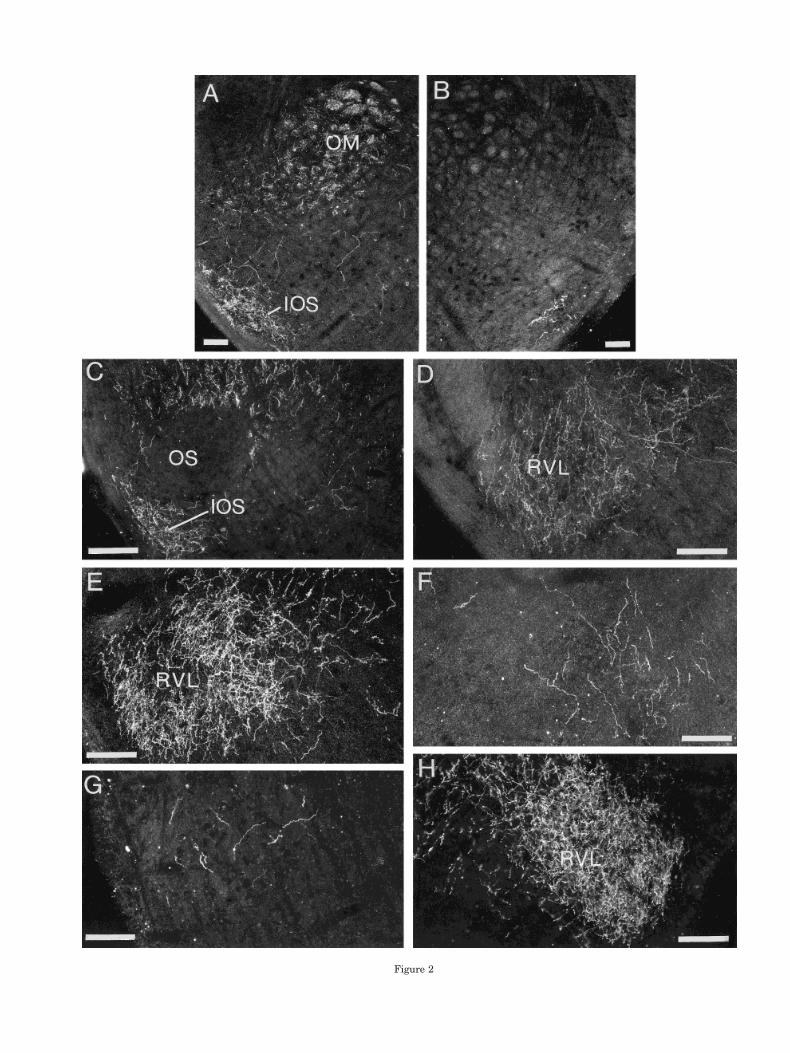
Figure 2
DISCUSSION
Technical considerations
The fact that none of the injections centered on RA wasconfined to the nucleus was an anticipated consequence ofthe attempt to label as much of RA as possible by making
large deposits of tracer. The implications of this extranu-clear spread of tracer are that some of the projections tothe diencephalon and upper brainstem cannot be assumedto originate from RA itself. Projections of RA to certaindorsal thalamic nuclei have been reported in zebra finchesand canaries in previous studies (Wild, 1993b; Vates et al.,1997), although in the present study the main terminalfield in the dorsal thalamus was in subhabenularis late-ralis (SHL) rather than in nucleus dorsomedialis posterior(DMP), as reported by Vates et al. (1997) who used para-sagittal sections. Projections to other diencephalic regions,however, including the hypothalamus, to ICo exclusive ofDM, to the tectum, to isthmic regions medial to the laterallemniscal nuclei, and to the medial tegmentum of themesencephalon almost certainly arise from archistriatal
Fig. 2. Darkfield photomicrographs of transverse sections throughthe lower pons and rostral medulla, showing the projections of RA tothe nucleus infraolivaris superior (IOS) and the ventrolateral nucleusof the rostral medulla (RVL), respectively. A: Ipsilateral IOS in acanary. B: Contralateral IOS in canary. C: Ipsilateral IOS in catbird.D: Ipsilateral RVL in catbird. E: Ipsilateral RVL in canary. F: Con-tralateral RVL in canary. G: Contralateral RVL in zebra finch.H: Ipsilateral RVL in zebra finch. Scale bars 5 100 mm.
Fig. 3. A: Darkfield photomicrographic montage showing the bi-lateral projections of RA to nucleus retroambigualis (RAm), the tra-cheosyringeal motor nucleus (XIIts), and the suprahypoglossal area(SH) in a male canary. Ipsilateral projections are on the left. Note thelabeled fibers crossing to the contralateral side through the commis-sura infima (CI). B: Darkfield photomicrograph showing more clearly
the bilateral projections of RA to XIIts and SH in another canary.C: Darkfield photomicrograph showing the largely ipsilateral projec-tions of RA to XIIts and SH in a zebra finch, with minor projectionscrossing the midline and sparse terminations in the contralateralXIIts. Scale bars 5 100 mm.
419BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS
regions outside RA (Wild et al., 1993; Mello et al., 1998;Wild and Williams, 2000). Injections of tracer into inter-collicular regions, or into deep tectal layers in the zebrafinch, for instance, retrogradely label large numbers ofcells surrounding RA, or on its dorsal and lateral aspects(Wild and Williams, 2000), and a projection from the me-dial archistriatum to a dorsal cap of SpM, similar to that
noted in the present study, has been observed in thepigeon, in which no RA exists (Wild, 1992). In any case,only the projections to the pons and medulla are of centralinterest in the present context. However, the lateral arch-istriatum, as well as RA in the medial archistriatum, hasmajor projections to these levels, but 1) the injectionscentered on RA did not encroach on the lateral archistria-
Fig. 4. Brightfield photomicrographs showing BDA-labeled termi-nal fields in the medulla of catbirds, following injections into the leftRA (dashed lines in B–D indicate the midline). A: Left RAm. B: XIItsand SH caudal to the obex, with crossing fibers and terminations inthe contralateral XIIts. C: XIIts and SH caudal to the obex, in another
catbird, with crossing fibers and terminations in the contralateralXIIts. D: XIIts at its caudal pole, with crossing fibers and termina-tions in the contralateral XIIts. Scale bars 5 200 mm in A,B; 100 mmin C,D.
420 J.M. WILD ET AL.

tum and 2) the targets of projections arising in the lateralarchisriatum are for the most part different from those ofRA and do not include the respiratory-vocal nuclei (Wildand Farabaugh, 1996). On the ipsilateral side of the me-dulla, RVL, XIIts, SH, RAm, and PAm were almost thesole targets of the projections labeled by the injectionscentered on RA, and, insofar as no extratelencephalic pro-jections are known to arise from the neostriatum, to whichtracer spread from the injections in RA, it is consideredthat the pontine and medullary projections described herearose from RA and not from surrounding archistriatal orneostriatal regions. It is also considered that this is astrue for the contralateral projections as it is for the ipsi-lateral projections, because, in the pons and medulla, con-tralateral projections were entirely confined to therespiratory-vocal nuclei. Furthermore, injections in XIItsretrogradely label cells only within RA and not withinother parts of the archistriatum (Vicario, 1991; Wild,1993a,b; Reinke and Wild, 1998).
Following XIIts injections, only a very few retrogradelylabeled cells were observed in the contralateral RA in thecanary and none in the zebra finch. These findings areconsistent with the relative proportions of ipsi- and con-tralateral RA projections in the two species and the smallsize of the injections, which were delivered iontophoreti-cally and were largely confined to XIIts and the suprahy-poglosssal area. Retrogradely labeled cells have beennoted in the contralateral RA following large pressureinjections of cholera toxin B-chain into OM and RAm ofzebra finches (Wild, 1993a). Despite the paucity of retro-grade confirmation of contralateral RA projections in thepresent study, the evidence of contralateral projectionsresulting from the unilateral RA injections cannot be de-nied. The contralateral projections of RA also cannot beassumed to arise by way of collaterals of somatopetallylabeled neurons in other nuclei, because the retrogradelylabeled cells that were observed following RA injections,such as those in the high vocal center (HVc) and thelateral magnocellular nucleus of the anterior neostriatum(lMAN), known sources of RA afferents, do not projectbeyond the telencephalon (Nottebohm et al., 1982; Bottjeret al., 1989).
Projections of nucleus robustus (RA) to thepons and medulla
After providing a relatively small terminal field to thenucleus infraolivaris superior (IOS) in the pons, and amore substantial one to the ventrolateral nucleus of therostral medulla (RVL), the projections of nucleus robustus(RA) terminate, as summarized in Figure 6, upon XIIts,upon the suprahypoglossal area (SH), and throughout thearc of RAm [and, at more rostral, periobex, levels of themedulla, throughout nucleus parambigualis (PAm); notshown in Fig. 6]. A variable and relatively small propor-tion of RA projections in different species leaves OM,crosses through the commissura infima (CI), and termi-nates in all the same nuclei on the contralateral side. RAmprojects upon XIIts bilaterally, but again predominanltlyipsilaterally, and also has reciprocal connections with itscontralateral homologue.
The ipsilateral projections of RA to the pons and me-dulla in the three species examined were essentially sim-ilar and did not appear to differ depending on which sidewas injected. The projections in the zebra finch have beendescribed previously (Gurney, 1981; Wild, 1993b; Vicario,
1993) and were completely confirmed in the present studyusing either left- or right-sided injections. Descending pro-jections of RA to the brainstem in canaries have not beenreported since Nottebohm et al.’s (1976) original descrip-tion, also in canaries of the Wasserschlager strain, basedon silver impregnation of degenerating fibers subsequentto RA lesions. In that study, the RA projections weredescribed as strictly ipsilateral and were predominantly toXIIts, although a relatively minor projection to the ven-trolateral brainstem was also observed. In the presentstudy, a pattern of projections resembling that in thezebra finch, with substantial ipsilateral terminations inIOS and RVL, and major ipsilateral terminations in PAmand RAm, as well as in XIIts and SH, is thus describedhere for the first time in this species. In addition, the RAprojections were shown to have a contralateral compo-nent. The projections of RA in catbirds are also describedfor the first time and are generally very similar to those inthe other two species, although the pattern of termina-tions in and around the catbird XIIts is slightly different,there being a characteristic pincer-like clustering of fibersand terminations lateral to the dorsal motor nucleus of thevagus caudal to the obex (see Fig. 4B).
The contralateral projections of RA to the medulla dodiffer between species, however; those in the zebra finchbeing very sparse, as was noted previously (Wild, 1993b),whereas those in catbirds and canaries are more substan-tial, particulary in canaries. On the face of it, these resultsdo not provide for a simple, consistent correlation with thedegree of lateralized syringeal dominance reported in thethree species. That is, if lateralized syringeal dominance isassumed to be greatest in canaries, least in zebra finches,and not present in catbirds, then a correlated pattern ofcontralateral RA projections might suggest that they aremost evident in canaries, which is true, least evident inzebra finches, which is also true, and not present at all incatbirds, which is not true. However, what is meant by“lateralized syringeal dominance” differs depending onhow dominance is measured. Zebra finches have been saidto be right-side-dominant for song control, but this refersnot so much to the loss of syllables following unilateralsyringeal denervation, as it does in canaries, for instance,as to changes in the acoustic structure of syllables follow-ing section of the right vs. the left tracheosyringeal nerveor lesion of the high vocal center (Nottebohm and Notte-bohm, 1976; Williams et al., 1992; Floody and Arnold,1997). Syllables that are lost postoperatively in zebrafinch song tend to be the high-frequency ones, especiallyfollowing section of the right tracheosyringeal nerve, afinding that could be seen as consistent with the generaltendency in passerine species for high notes to be pro-duced on the right side (Suthers, 1997). It may be, there-fore, that there is very little lateralized syringeal domi-nance for syllabic production in the zebra finch, in thesense in which this term has been applied to canary andsparrow song, and it is tempting to speculate that thepaucity of contralateral RA projections to XIIts and RAmin this species reflects this situation. However, in catbirds,in which both sides of the syrinx frequently contribute tosyllabic production, and in which there is no lateral dom-inance for syllabic production, there were considerablymore RA projections to the contralateral XIIts than in thezebra finch. It must be concluded, therefore, that the rel-ative number of contralateral RA projections in differentspecies does not necessarily reflect the degree of lateral-
421BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS
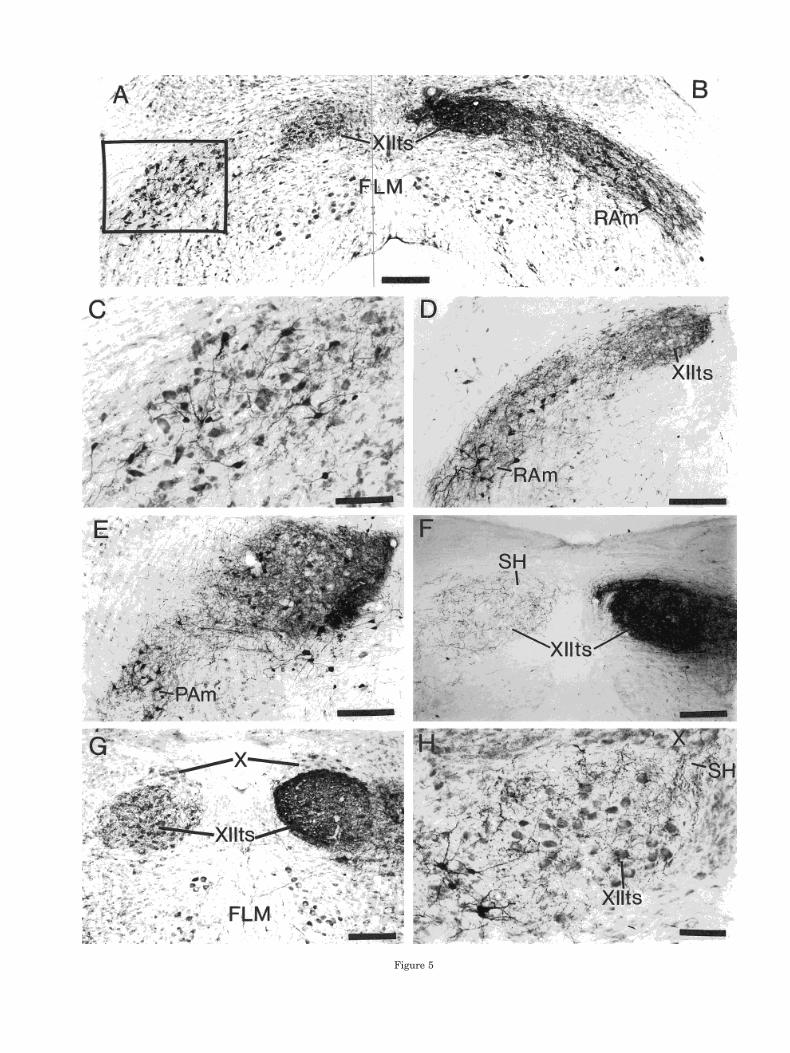
Figure 5

ized syringeal dominace. Furthermore, even in thestrongly lateralized canary, if there is a difference in theextent of contralateral projections originating in the leftvs. the right RA, it was not apparent in the small numberof animals used in the present study. The pronouncedbehavioral left–right syringeal differences in syllabic pro-duction cannot therefore readily be explained on the basisof an obvious anatomical asymmetry in the premotor con-trol pathway.
Although contralateral RA projections may not be re-lated to syringeal dominance in a general way, the possi-bility remains that, within any one species, they are usedfor the coordination of left and right sides during vocalproduction. These contralateral inputs of RA to XIIts andRAm are also substantially reinforced by the bilateralprojections of more rostral brainstem nuclei, such as IOSand RVL, to which RA also projects bilaterally in canariesand zebra finches (present study; Wild, 1993b; Vicario,1993). DM also receives a bilateral input from RA in allthree species, and this nucleus also projects bilaterallyupon XIIts and RAm (Wild, 1993b; Wild et al., 1997).There are thus multiple ways for the RA of one side toinfluence the activity of XIIts and RAm on both sides ofthe medulla, one of which is via a direct projection,whereas others are indirect. A possible function of theindirect projections (e.g., via IOS and RVL) might be tointroduce temporal delays into the descending motor com-mands that eventually reach XIIts and RAm, delays thatcould be important in the temporal patterning of thespecies-typical song.
Local projections of nucleusretroambigualis (RAm)
In contrast to the marked differences in the contralat-eral projections of RA to the medulla in canaries and zebrafinches, no such differences were observed in the projec-tions of RAm in these species. In both there was a strongprojection of RAm to the ipsilateral XIIts and a compara-tively weak, but nonetheless substantial, projection to the
contralateral XIIts and the opposite RAm, although,again, left–right asymmetries in these projections werenot obvious to the eye. As described here, RAm includesneurons throughout the whole arc that extends from theborders of XIIts to the ventrolateral periphery of the me-dulla. This arc is largely coextensive, on the one hand,with the terminal field of nucleus robustus in the caudalmedulla (present study; Wild, 1993b) and, on the other,with the origins of bulbospinal neurons that may projectupon expiratory motoneurons in the lower thoracic andupper lumbosacral spinal cord (Reinke and Wild, 1998).RAm may even include some cells that lie dorsal to XIIts,i.e., within the suprahypoglossal area (SH).
Thus, the bilateral projections of RAm to XIIts mayexplain the findings of Vicario and Nottebohm (1988), whofound that applying microstimulation unilaterally to XIItsin zebra finches resulted in a short-latency response in theipsilateral syrinx and a longer latency response that in-cluded the contralateral syrinx. The contralateral re-sponses were usually evoked from regions dorsal to thosefrom which ipsilateral responses were evoked, an obser-vation that may be consistent with the activation of su-prahypoglossal (SH) and RAm neurons that in the presentstudy were found to project to XIIts bilaterally. Similarly,these bilateral projections of RAm to XIIts, and of therelatively minor contralateral projections of nucleus ro-bustus to RAm and XIIts, may also bear on the findings ofPaton and Manogue (1982) in canaries and zebra finchesto the effect that electrical stimulation of the high vocalcenter in the telencephalon could evoke a weak responsein the contralateral tracheosyringeal nerve, in addition toa strong response in the ipsilateral nerve, only when highrates of stimulation were used. The suggestion that thevocal motoneurons themselves could mediate such a con-tralateral response by sending a collateral axon across themidline through the commissura infima (Nottebohm,1980) can now be ruled out on the basis of the presentresults; no motoneuronal cell bodies were retrogradelylabeled in XIIts following injections of tracer into theopposite XIIts. Nottebohm (1980) assumed correctly, how-ever, that an alternative source of bilateral interactionsbetween the two XIIts nuclei might be an interneuron,such as was identified in RAm in the present study.
The present results also provide the anatomical sub-stantiation of the suggestions of Manogue and Paton(1982) regarding the medullary source of “respiratory gat-ing” of vocal output. These authors found that electricalstimulation of premotor vocal control nuclei in the telen-cephalon of budgerigars and zebra finches producedgreater activity in nucleus XIIts or tracheosyringeal nerveduring expiration than during inspiration, but RA excit-ability was not affected by antidromic stimulation of XIIts,suggesting that neither RA nor the high vocal center wasthe source of the respiratory gating. The present resultsstrongly suggest that RAm and SH neurons that mayproject bilaterally to XIIts are the source of the increasedexcitability of XIIts neurons during expiration. It is alsopossible, however, that the projections to XIIts frominspiratory-related neurons in PAm (Reinke and Wild,1998; present results) reduce the excitability of XIIts neu-rons during inspiration. In any case, the local projectionsto XIIts from RAm and PAm seem likely to be involved inrespiratory-vocal coordination, by ensuring that the activ-ity of syringeal motoneurons is timed with that of appro-priate expiratory effort during phonation.
Fig. 5. Brightfield photomicrographs showing, in transverse sec-tions, retrograde and/or anterograde labeling in the medulla of canar-ies and zebra finches, following iontophoretic injections of BDA inXIIts. A,B: Montage of a Nissl-counterstained section caudal to theobex in a canary. The injection of BDA is in the right XIIts, whichproduced dense retrograde labeling in the ipsilateral RAm, substan-tial retrograde labeling in the contralateral RAm (boxed area), andanterograde labeling in the contralateral XIIts (the cells in the leftXIIts are not retrogradely labeled). C: Higher power view of the boxedarea in A. D: Zebra finch, uncounterstained section. Retrograde andanterograde lebeling in the left RAm, and anterograde labeling in theleft XIIts, following an injection of BDA in the left XIIts at a morecaudal level. E: Zebra finch, Nissl-counterstained section. An injectionof BDA in the ventromedial part of the left XIIts rostral to the obexand retrogradely labeled cells in the dorsomedial part of the ipsilat-eral nucleus parambigualis (PAm). F: Zebra finch, uncounterstainedsection. An injection in the right XIIts and SH caudal to the obex andanterograde labeling in the contralateral XIIts and SH. G: Zebrafinch, Nissl-counterstained section. An injection in the right XIIts andSH rostral to the obex and anterograde labeling in the contralateralXIIts and SH (the cell bodies in the left XIIts are not retrogradelylabeled). H: Zebra finch, Nissl-counterstained section. Left XIIts andsurrounding regions, including SH, showing retrogradely labeled cellsventrolateral to XIIts, and anterograde labeling in XIIts and SH,following an injection of BDA centered on the right XIIts and SH (seetext). Note the processes (dendrites) of the retrogradely labeled cellsentering XIIts. Scale bars 5 200 mm in A,B,D–G; 100 mm in C,H.
423BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS
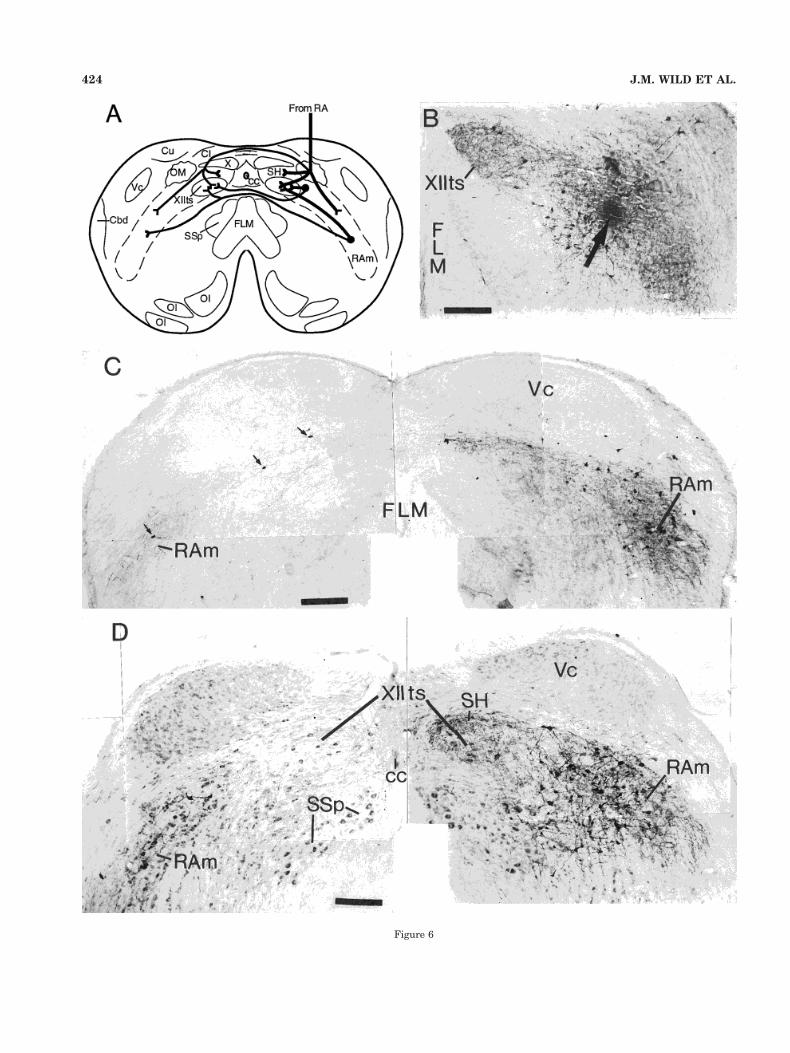
Figure 6
424 J.M. WILD ET AL.

In the case of neither RAm nor PAm, however, is itcurrently known whether the neurons that project to XIItsare the same ones that project upon respiratory motoneu-rons in the cord (Reinke and Wild, 1998), nor do thepresent data allow us to determine with confidencewhether the projections of RAm involve syringeal mo-toneurons innervating groups of both dorsal and ventralsyringeal muscles or are restricted to only one of thesegroups. This will be important to determine in futurestudies (using intracellular labeling techniques) becauseof the possible role of these groups of motoneurons inlateralized phonation and bilateral frequency control, re-spectively (Goller and Suthers, 1996a; b).
In addition to whatever role the RAm projections toXIIts may play in vocalization, they seem likely to sub-serve a more basic function related to syringeal muscletone during respiration, because expiratory-related activ-ity is present in XIIts even during quiet respiration, atleast in zebra finches, budgerigars, and canaries(Manogue and Paton, 1982; Vicario and Nottebohm, 1988;Wild, unpublished observations). These findings are sup-ported by the finding that cutting the motor roots of thetracheosyringeal nerve in zebra finches may result in abird that phonates with every expiration, particularly ifthis is done on the right side and if the bird is stressed(Wild and Gahr, unublished observations). This is pre-sumably because the syringeal membranes are therebyrendered flaccid and hence partially obstruct the airway(Manogue and Paton 1982).
Comparison to mammals
A neural system for the mediation of respiratory-vocalcontrol, similar to that delineated in the brainstem ofbirds, is also present in the brainstem of mammals (Daviset al., 1996a,b; Reinke and Wild, 1997). In particular,RAm and PAm in birds are components of an avian ven-tral respiratory group (VRG), which, like nucleus retroam-biguus (NRA) in the cat, project upon spinal motoneuronsinnervating respiratory muscles (Holstege, 1989; Wild,1993a; Reinke and Wild, 1997, 1998). NRA in the cat wasalso thought to project upon many of the perioral, hypo-glossal, and possibly laryngeal motor neurons and wastherefore considered to be the nexus of a final common
path for vocalization (Holstege, 1989; cf. Thoms and Jur-gens, 1987, for the squirrel monkey). However, whetherthese brainstem projections actually originate from NRAor from neurons in close proximity to NRA is not clear.
A significant descending input to NRA in the cat origi-nates from the periaqueductal grey (PAG; Holstege, 1989;Jurgens, 1994; Gerrits and Holstege, 1996), which inmammals, including humans, is considered to be the ma-jor pattern generator involved in respiratory-vocal control(Davis et al., 1996a,b). A similar projection to RAm andPAm in birds originates, not in the central grey thatsurrounds the aqueduct (i.e., GCt), but in the dorsomedialnucleus of the intercollicular complex (DM), which, be-cause of the massive lateral expansion of the tectum inbirds, may be homologous with parts of the lateral PAG inmammals. In nonsongbirds, such as pigeons, DM is alsolikely to be a major pattern generator for respiratory-vocalmovements involved in calling (Wild et al., 1997), but, innonsongbirds and most mammals, apart from the human,there appears to be little or no projection originating in thetelencephalon that terminates directly on vocal motoneu-rons, and the extent of cortical terminations on respira-tory premotor neurons is unknown (Kuypers, 1958;Muller-Preuss and Jurgens, 1976; Sutton and Jurgens,1988). In contrast, songbirds have a major telencephalicsource of direct projections onto vocal motoneurons andrespiratory premotor neurons, shown here and elsewhereto originate in nucleus robustus (RA; Nottebohm et al.,1976). A major reason for this difference between song-birds and nonsongbirds, and between songbirds and non-human primates and other mammals, appears to be thatsong in songbirds, as with speech in humans, is largely alearned phenomenon and hence requires a dedicated neu-ral system within the telencephalon not only for vocallearning but also for vocal production (Doupe and Kuhl,1999). As we have shown here and elsewhere, vocal pro-duction in birds in general and songbirds in particular ismediated via neural pathways that incorporate bilateralprojections for the efficient coordination of structures onboth sides of the body.
ACKNOWLEDGMENTS
This work was supported by Whitehall Foundation, Inc.grant M94RO7 to J.M.W. and National Institutes ofHealth grant 2 RO1 NS29467-07 to R.A.S.
LITERATURE CITEDAllan SE, Suthers RA. 1984. Lateralization and motor stereotopy of song
production in the brown headed cowbird. J Neurobiol 25:1154–1166.Bottjer SW, Halsema KA, Brown SA, Meisner EA. 1989. Axonal connec-
tions of a forebrain nucleus involved with vocal learning in zebrafinches. J Comp Neurol 279:312–326.
Colemen MJ, Sule PJ, Vu ET. 1999. Recovery of impaired songs followingunilateral but not bilateral lesions of nucleus uvaeformis of adult zebrafinches. Soc Neurosci Abstr 25:1367.
Davis PJ, Zhang SP, Winkworth A, Bandler R. 1996a. Neural control ofvocalization: respiratory and emotional influences. J Voice 10:23–38.
Davis PJ, Zhang SP, Bandler R. 1966b. Midbrain and medullary regulationof vocalization. In: Davis PJ, Fletcher NH, editors, Vocal fold physiol-ogy, controlling complexity and chaos. San Diego: Singular PublishingGroup, Inc. p 121–135.
DeVoogd TJ, Nottebohm F. 1981. Sex differences in dendritic morphologyof a song control nucleus in the canary: a quantitative Golgi study.J Comp Neurol 196:309–316.
DeVoogd TJ, Pyskaty DJ, Nottebohm F. 1991. Lateral asynmmetries and
Fig. 6. A: Summary diagram (transverse section through the cau-dal medulla of a finch) depicting respiratory-vocal pathways consis-tent with the main neural circuitry delineated in the present study.Descending projections from RA terminate predominantly in the ip-silateral XIIts, in SH, and throughout the entire arc that includes therespiratory premotor nucleus Ram as well as PAm more rostrally. Inthe canary, in particular, the RA projections also terminate in thecontralateral XIIts, SH, and RAm. Cell bodies (black dots) adjacent toXIIts, in more ventrolateral parts of RAm (and in PAm), projectbilaterally upon XIIts, and cells in RAm also have reciprocal projec-tions with the contralateral RAm. The extent to which single cellsproject to XIIts and RAm via branched axons is not known. B: Aninjection (at the arrow) in the right RAm of a canary that producedpredominantly anterograde labeling in XIIts at that level. C: Montageshowing the caudal end of the injection depicted in B and anterogradeand retrograde labeling (small arrows) in the contralateral RAm.D: Montage of a Nissl-counterstained section through the caudalmedulla of a zebra finch showing part of an injection in the right RAm,anterograde labeling in XIIts and SH bilaterally and anterogradelabeling and retrogradely labeled cells in the left (contralateral) RAm.Scale bars 5 200 mm.
425BILATERAL VOCAL CONTROL PATHWAYS IN SONGBIRDS

testosterone-induced changes in the gross morphology of the hypoglos-sal nucleus in adult canaries. J Comp Neurol 307:65–76.
Doupe AJ, Kuhl PK. 1999. Birdsong and human speech. Annu Rev Neu-rosci 22:567–631.
Floody OR, Arnold AP. 1997. Song lateralization in the zebra finch. HormBehav 31:25–34.
Gerrits PO, Holstege G. 1996. Pontine and medullary projections to nu-cleus retroambiguus: a wheat germ agglutinin-horseradish peroxidaseand autoradiographic tracing study in the cat. J Comp Neurol 373:173–185.
Goller F, Larsen ON. 1997. A new mechanism of sound generation insongbirds. Proc Natl Acad Sci USA 94:14787–14791.
Goller F, Suthers RA. 1996a. Role of syringeal muscles in gating airflowand sound production in singing brown thrashers. J Neurophysiol75:867–876.
Goller F, Suthers RA. 1996b. Role of syringeal muscles in controlling thephonology of bird song. J Neurophysiol 76:287–300.
Gurney ME. 1981. Hormonal control of cell form and number in the zebrafinch song system. J Neurosci 1:658–673.
Hartley RS, Suthers RA. 1989. Airflow and pressure during canary song:direct evidence for mini-breaths. J Comp Physiol 165:15–26.
Holstege G. 1989. Anatomical study of the final common path for vocaliza-tion in the cat. J Comp Neurol 284:242–252.
Jurgens U. 1994. The role of the periaqueductal grey in vocal behaviour.Behav Brain Res 62:107–117.
Kuypers HGJM 1958. Corticobulbar connections to the pons and lowerbrain stem in man. An anatomical study. Brain 81:364–388.
Lemon RE. 1973. Nervous control of the syrinx in white-throated sparrows(Zonotrichia albicollis). J Zool (Lond) 71:131–140
Manogue KR, Paton JA. 1982. Respiratory gating of activity in the avianvocal control system. Brain Res 247:383–387.
Mello CV, Vates GE, Okuhata S, Nottebohm F. 1998. Descending auditorypathways in the adult male zebra finch (Taeniopygia guttata). J CompNeurol 395:137–160.
Muller-Preuss P, Jurgens U. 1976. Projections from the “cingular” vocal-ization area in the squirrel monkey. Brain Res 103:29–43.
Nottebohm F. 1970. Ontogeny of bird song. Science 167:950–956.Nottebohm F. 1971. Neural lateralization of vocal control in a passerine
bird. I. Song. J Exp Zool 177:229–262.Nottebohm F. 1972. Neural lateralization of vocal control in a passerine
bird. II. Subsong, calls, and a theory of vocal learning. J Exp Zool179:35–50.
Nottebohm F. 1977. Asymmetries in neural control of vocalization in thecanary. In: Harnad S, Doty RW, Goldstein L, Jaynes, J, Krauthamer G.editors. Lateralization in the nervous system. New York: AcademicPress. p 23–44.
Nottebohm F. 1980. Brain pathways for vocal learning in birds: a review ofthe first ten years. Progr Psychobiol Physiol Psychol 9:85–124.
Nottebohm F, Nottebohm ME. 1976. Left hypoglossal dominance in thecontrol of canary and white-crowned sparrow song. J Comp Physiol108A:171–192.
Nottebohm F, Stokes TM, Leonard CM. 1976. Central control of song in thecanary, Serinus canaria. J Comp Neurol 165:457–486.
Nottebohm F, Kelley DB, Paton JA. 1982. Connections of vocal controlnuclei in the canary telencephalon. J Comp Neurol 207:344–357.
Okuhata S, Nottebohm F. 1982. Nucleus uva might be part of a feedbackcircuit for song processing. Soc Neurosci Abstr 18:527.
Paton JA, Manogue KR. 1982. Bilateral interactions within the vocalcontrol pathway of birds: two evolutionary alternatives. J Comp Neurol212:329–335.
Reinke H, Wild JM. 1997. Distribution and connections of inspiratorypremotor neurons in the brainstem of the pigeon (Columba livia).J Comp Neurol 379:347–362.
Reinke H, Wild JM. 1998. Identification and connections of inspiratorypremotor neurons in songbirds and budgerigar. J Comp Neurol 391:147–163.
Ruan J, Suthers RA. 1996. Myotopic representation of syringeal muscles inthe hypoglossal nucleus of the cowbird. Soc Neurosci Abstr 22:1402.
Seller TJ. 1979. Unilateral nervous control of the syrinx in Java sparrows(Padda oryzivora). J Comp Physiol 129:281–288.
Stokes TM, Leonard CM, Nottebohm F. 1974. The telencephalon, dienceph-alon, and mesencephalon of the canary, Serinus canaria, in stereotaxiccoordinates. J Comp Neurol 156:337–374.
Striedter GF, Vu ET. 1997. Bilateral feedback projections to the forebrainin the premotor network for singing in zebra finches. J Neurobiol34:27–40.
Suthers RA. 1990. Contributions to birdsong from the left and right sidesof the intact syrinx. Nature 347:473–477.
Suthers RA. 1992. Lateralization of sound production and motor action onthe left and right sides of the syrinx during bird song. Proceedings ofthe 14th International Congress on Acoustics, Beijing, China, vol 4, pI1–I5.
Suthers RA. 1997. Peripheral control and lateralization of birdsong. J Neu-robiol 33:632–652.
Suthers RA, Goller F. 1996. Respiratory and syringeal dynamics of songproduction in northern cardinals. In: Burrows M, Matheson, T, New-land P, Schuppe H. editors. Nervous systems and behaviour: proceed-ings of the 4th International Congress on Neuroethology. Stuttgart:Georg Thieme Verlag. p 333.
Suthers RA, Goller F. 1997. Motor correlates of vocal diversity in song-birds. In: Nolan V Jr, et al., editors. Current ornithology, vol 14. NewYork: Plenum Press. p 235–288.
Suthers RA, Goller F, Hartley RS. 1994. Motor dynamics of song produc-tion by mimic thrushes. J Neurobiol 25:917–936.
Sutton D, Jurgens, U. 1988. Neural control of vocalization. Comp PrimateBiol Vol 4: Neurosciences. p 625–647.
Thoms G, Jurgens U. 1987. Common input of the cranial motor nucleiinvolved in phonation in the squirrel monkey. Exp Neurol 95:85–99.
Vates GE, Vicario DS, Nottebohm F. 1997. Reafferent thalamo-“cortical”loops in the song system of oscine songbirds. J Comp Neurol 380:275–290.
Veenman CL, Reiner A, Honig MG. 1992. Biotinylated dextran amine as anantergrade tracer for single- and double-labeling studies. J NeurosciMethods 41:239–254.
Vicario DS. 1991. Organization of the zebra finch song control system: II.Functional organization of outputs from nucleus robustus archistria-talis. J Comp Neurol 309:486–494.
Vicario DS. 1993. A new brain stem pathway for vocal control in the zebrafinch song system. Neuroreport 4:983–986.
Vicario D, Nottebohm F. 1988. Organization of the zebra finch song controlsystem. I. Representation of syringeal muscles in the hypoglossal nu-cleus. J Comp Neurol 271:346–354.
Vu ET, Mazurek ME, Kuo Y-C. 1994. Identification of a forebrain motorprogramming network for the learned song of zebra finches. J Neurosci14:6924–6934.
Vu ET, Schmidt MF, Mazurek ME. 1996. Interhemispheric coordination ofpremotor activity during singing by zebra finches. Soc Neurosci Abstr22:151.
Wild JM. 1992. Direct and indirect “cortico”-rubral and rubro-cerebellarcortical projections in the pigeon (Columba livia). J Comp Neurol326:623–636.
Wild JM. 1993a. The avian nucleus retroambigualis: a nucleus for breath-ing, singing and calling. Brain Res 606:119–124.
Wild JM. 1993b. Descending projections of the songbird nucleus robustusarchistriatalis. J Comp Neurol 338:225–241.
Wild JM. 1994. The auditory-vocal-respiratory axis in birds. Brain BehavEvol 44:192–209.
Wild JM. 1997. Neural pathways for the control of birdsong production.J Neurobiol 33:653–670.
Wild JM, Farabaugh SM. 1996. Organization of afferent and efferentprojections of nucleus basalis prosencephali in a passerine (Taeniopy-gia guttata). J Comp Neurol 365:306–328.
Wild JM, Williams MN. 2000. Rostral wulst in passerine birds. I. Origin,course and terminations of an avian pyramidal tract. J Comp Neurol416:429–450.
Wild JM, Karten HJ, Frost BJ. 1993. Connections of the auditory forebrainin the pigeon (Columba livia). J Comp Neurol 337:32–62.
Wild JM, Li D, Eagleton C. 1997. Projections of the dorsomedial nucleus ofthe intercollicular complex in relation to respiratory-vocal nuclei in thebrainstem of pigeon (Columba livia) and zebra finch (Taeniopygiaguttata). J Comp Neurol 377:392–413.
Williams H, Vicario DS. 1993. Temporal patterning of song production:participation of nucleus uvaeformis of the thalamus. J Neurobiol 24:903–912.
Williams H, Crane LA, Hale TK, Esposito MA, Nottebohm F. 1992. Right-sided dominance for song control in the zebra finch. J Neurobiol 23:1006–1020.
426 J.M. WILD ET AL.





























