Embed Size (px)
Citation preview

Review Article
Extraembryonic Endoderm cells as a model of endodermdevelopment
Asja T. Moerkamp,1 Agnieszka Paca,2 Marie-Jos�e Goumans,1 Tilo Kunath,2
Boudewijn P. T. Kruithof1 and Marianna Kruithof-de Julio1,*1Department of Molecular and Cell Biology, Centre of Biomedical Genetics, Leiden University Medical Center, Leiden,
the Netherlands; and 2Medical Research Council Centre for Regenerative Medicine, University of Edinburgh, Edinburgh,UK
In recent years the multipotent extraembryonic endoderm (XEN) stem cells have been the center of much atten-tion. In vivo, XEN cells contribute to the formation of the extraembryonic endoderm, visceral and parietal endo-derm and later on, the yolk sac. Recent data have shown that the distinction between embryonic andextraembryonic endoderm is not as strict as previously thought due to the integration, and not the displace-ment, of the visceral endoderm into the definitive embryonic endoderm. Therefore, cells from the extraembry-onic endoderm also contribute to definitive endoderm. Many research groups focused on unraveling thepotential and ability of XEN cells to both support differentiation and/or differentiate into endoderm-like tissues asan alternative to embryonic stem (ES) cells. Moreover, the conversion of ES to XEN cells, shown recently with-out genetic manipulations, uncovers significant and novel molecular mechanisms involved in extraembryonicendoderm and definitive endoderm development. XEN cell lines provide a unique model for an early mammalianlineage that complements the established ES and trophoblast stem cell lines. Through the study of essentialgenes and signaling requirements for XEN cells in vitro, insights will be gained about the developmentalprogram of the extraembryonic and embryonic endodermal lineage in vivo. This review will provide an overviewon the current literature focusing on XEN cells as a model for primitive endoderm and possibly definitive endo-derm as well as the potential of using these cells for therapeutic applications.
Key words: definitive endoderm, development, extraembryonic endoderm, stem cells, XEN cells.
Introduction: Extraembryonic endodermdevelopment
The mature mouse blastocyst (4.5 days post-coitum
(dpc)) consists of three distinct cell types: the trophec-
toderm, which gives rise to the trophoblast and extra-
embryonic ectoderm (ExEc), the pluripotent cells of the
epiblast, and the primitive or extraembryonic endo-
derm (ExEn), an epithelial layer of cells on the surfaceof the epiblast. The primitive endoderm gives rise to: (i)
visceral endoderm (VE) that surrounds the epiblast and
the ExEc; and (ii) parietal endoderm (PE) that interacts
with the trophoblast giant cell layer. PE cells migratealong the inner surface of the trophectoderm and
together with trophoblast giant cells form the parietal
yolk sac (Hogan et al. 1980). The PE, as well as the
VE, mediates nutrient-waste exchange for the develop-
ing embryo. Initially the VE overlays only the epiblast,
but as the ExEc increases in size the VE quickly
expands to also cover the ExEc. The VE overlaying the
epiblast becomes molecularly and morphologically dis-tinct from the VE in contact with the ExEc around
5.0 dpc, representing the embryonic VE (emVE) and
extraembryonic VE (exVE), respectively. The cells from
the exVE are columnar and cuboidal, while the emVE
cells are flatter and more epithelial in shape (Takito &
Al-Awqati 2004).
Around 5.5 dpc a group of cells at the distal tip of
the epiblast differentiates into a morphologically distin-guishable subset of emVE, the distal visceral endo-
derm (DVE) (Rivera-Perez et al. 2003; Srinivas et al.
2004). This marks the formation of the first axis of the
*Author to whom all correspondence should be addressed.Email: [email protected] 3 December 2012; revised 19 December 2012;
accepted 19 December 2012.ª 2013 The AuthorsDevelopment, Growth & Differentiation ª 2013 Japanese
Society of Developmental Biologists
Develop. Growth Differ. (2013) 55, 301–308 doi: 10.1111/dgd.12036
The Japanese Society of Developmental Biologists

body, the distal–proximal axis. Within 4–5 h (betweenapproximately 5.75 and 6.0 dpc) the DVE migrates
proximally as a continuous epithelial sheet to the pro-
spective anterior pole of the embryo. The underlying
mechanism of this migration is yet to be fully charac-
terized and both active migration and differences in
the proliferation rate of the anterior versus posterior
epiblast have been suggested (Srinivas et al. 2004;
Migeotte et al. 2010; Stuckey et al. 2011; Trichaset al. 2011). The unilateral movement of the DVE
changes the distal–proximal axis into the anterior–posterior axis of the embryo and the DVE is now
called anterior visceral endoderm (AVE).
The VE and its derivatives, play critical roles in orga-
nization and differentiation of the epiblast. The VE is
the first site of hematopoiesis (Toles et al. 1989; McG-
rath & Palis 2005) and induces through the expressionof Indian hedgehog and vascular endothelial growth
factor the formation of blood islands and endothelial
cells (Dyer et al. 2001; Byrd et al. 2002; Damert et al.
2002). In addition, the proximal VE was shown to be
involved in early primordial germ cell differentiation (de
Sousa Lopes et al. 2004, 2007). Finally, microsurgical
removal of AVE resulted in anterior neural structures
truncations (Thomas & Beddington 1996) and itsderived BMP2 signals have been shown to take part in
heart positioning and foregut invagination (Madabhushi
& Lacy 2011).
Interestingly, the lineage distinction between embry-
onic and ExEn tissue was marked by the assumption
that the VE that surrounds the epiblast was displaced
by the definitive endoderm. Recently it has been
shown that cells from the VE persist within the defini-tive endoderm layer of the embryo and contribute to
the early gut tube. This suggests that the distinction
between extraembryonic and embryonic tissues is not
as strict as believed and the lineage that was previ-
ously considered to be exclusively embryonic has
extraembryonic contributions (Kwon et al. 2008). An
interesting question is whether, within the definitive
endoderm and potentially its derivative tissues, thereare molecular and functional differences between the
ExEn- and epiblast-derived cells that may be studied
by in vitro culturing and manipulating cells representa-
tive of the ExEn.
Derivation, properties and applications ofExEn-derived stem cells
Stem cells can be derived from each of the primary lin-
eages of the mammalian embryo (Fig. 1). ES cells from
the inner cell mass (ICM) or early epiblast, trophoblast
stem (TS) cells from the trophectoderm layer and XEN
stem cells from the primitive endoderm (or ExEn). Most
importantly each one of these stem cell systems arecapable of indefinite self-renewal in culture and once
reintroduced into the mouse embryo will display line-
age restricted contributions in the resulting chimeric
embryos that are consistent with their lineage of origin
(Beddington & Robertson 1989; Tanaka et al. 1998;
Kunath et al. 2005). Interestingly, in vivo, XEN cells
can only repopulate the PE, rarely the VE (Kunath
et al. 2005; Kruithof-de Julio et al. 2011).Initially, parietal endoderm cell (PEC) lines were iso-
lated by Fowler et al. (1990). In vitro studies have
shown that these cells have characteristics of PE, clo-
sely resembling the basement membrane matrix of
Reichert’s membrane. However, their chimera contri-
bution potential was not assessed (Fowler et al. 1990).
PECs morphologically resemble the more recently
isolated XEN cells (Kunath et al. 2005; reviewed inRossant 2007) being round and refractile or stellate or
epithelial-like cells. Several methods of isolation have
been proposed for XEN cell derivation, which lead all
to cells with similar morphological characteristics; how-
ever, their response to growth factors seems to be
influenced by the derivation process. In this context, it
is essential to appreciate that primitive endoderm
already exhibits heterogeneous expression of some ofthe DVE/AVE markers, such as Cer1 and Lefty1 (Tak-
aoka et al. 2006, 2011; Torres-Padilla et al. 2007).
Furthermore, Nodal/Activin signaling can be differen-
tially perceived within the primitive endoderm (Granier
et al. 2011). This would imply that, at the time of sig-
naling pathway maturation, primitive endoderm cells
are sensitive to even minor changes in signaling inten-
sity. Interestingly, primitive endoderm-progenitorsexpressing Oct4 exhibit greater developmental plastic-
ity than Oct4-expressing epiblast progenitors at a simi-
lar stage (Grabarek et al. 2012) which could reflect the
in vitro heterogeneous nature of XEN cells.
Extensive microarray analysis on XEN cells per-
formed by several groups all agree in the expression of
primitive endoderm markers SOX7, GATA4, GATA6
and the VE markers hHEX and DKK1. Furthermore,XEN cells are characterized by the lack of AFP expres-
sion as well as the absence of definitive endoderm
markers (Kunath et al. 2005; Brown et al. 2010b; Kru-
ithof-de Julio et al. 2011). Given their reproducible der-
ivation from the ExEn and their expression profile
similar to this extraembryonic tissue, XEN cells can be
a powerful tool to study inductive effects attributed to
the AVE. Brown et al. (2010a,b) have undertaken anextensive array analysis of three cell lines that are simi-
lar to the heart-inducing AVE: two embryonal carci-
noma-derived (END2 and PYS2) and the XEN cells. By
comparing the gene expression profiles they have
identified a discrete set of genes that could support
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
302 A. T. Moerkamp et al.
myocardial differentiation. In addition, by using XEN
cell-derived conditioned media on embryoid bodies
they were able to enhance cardiogenesis (Brown et al.
2010a). These are interesting observations with poten-
tial therapeutic properties. In order to obtain the
appropriate cell products (in this context, cardiomyo-cytes for cardiac repair), it is of importance to stepwise
direct the fate of the stem cell of interest with the
proper instructive signals in vitro. During development,
the VE interacts with the nascent mesoderm to induce
the cardiac fate. Therefore, XEN cells could be used to
recapitulate this developmental process in vitro and
induce and accelerate differentiation of the susceptible
stem cells into the cardiac lineage.Although derived from primitive endoderm, XEN cells
contribute efficiently in chimeras to PE but not to VE
(Kunath et al. 2005; Kruithof-de Julio et al. 2011). This
lack of contribution to the VE lineage could be caused
by many factors, including the preferential interaction
with the mural trophectoderm (Artus et al. 2012; Kruit-
hof-de Julio and Shen, unpubl. data, 2009), the alter-
ation of a signaling pathway in the establishment ofthe cell line based on the derivation process (XEN cells
derived in the presence of LIF poorly respond to
growth factor stimulation) or the derivation of a “com-
mitted” cell that has lost potency. In support of the lat-
ter, extraembryonic endoderm precursor (XEN-P) stem
cells have been derived from rat blastocyst (Debeb
et al. 2009; Galat et al. 2009). These cells are charac-
terized by a less “endoderm” defined gene expression
profile; they express the ES markers OCT4, REX1, AP,
and SSEA1 and contribute to the PE as well as the VE
lineages in chimeras. The authors suggest they are
precursor endoderm cells as they could represent the
first committed step of the ExEn.
Finally, with respect to the plasticity or commitmentof stem cells, recently Cho et al. (2012) was able to
derive XEN cells from mouse ES cells via the addition
of exogenous retinoic acid and Activin. These XEN
cells are indistinguishable from embryo-derived XEN
cells, including their differentiation capacity (Cho et al.
2012). These data show a high degree of plasticity
within the ICM and provide the possibility of deriving
XEN cell lines from the various mutant ES cell linesavailable, thereby shedding light on the factors
required for XEN cell derivation and the development
of ExEn.
Signaling pathways in XEN cells
Characteristic of XEN cells are the transcription factors
GATA4 and GATA6, specifically expressed in theExEn. GATA6 is present from 3.5 dpc in the ICM in a
salt and pepper distribution with NANOG and then
restricted to the primitive endoderm (Chazaud et al.
2006). Gata6-deficient mice are embryonically lethal
and Gata6-null ES cells fail to specify VE in vivo and
in vitro, suggesting a key role for GATA6 in both VE
and PE differentiation (Morrisey et al. 1998). Forced
and maintained expression of the Gata4 and Gata6 in
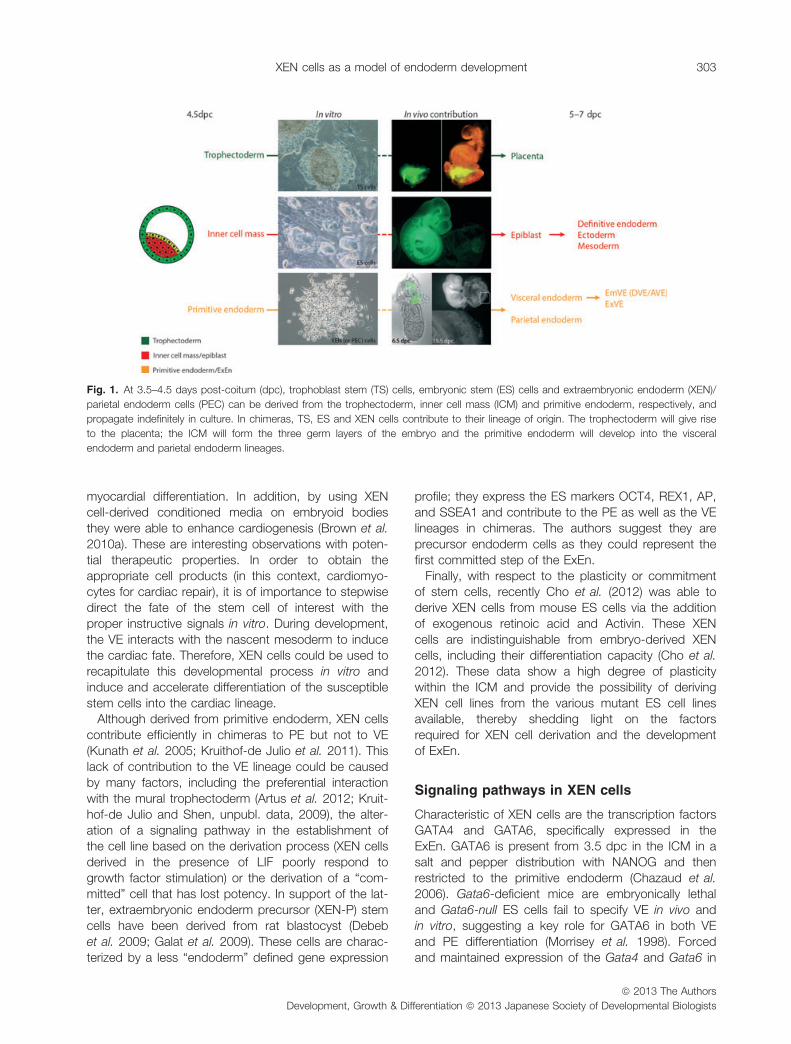
Fig. 1. At 3.5–4.5 days post-coitum (dpc), trophoblast stem (TS) cells, embryonic stem (ES) cells and extraembryonic endoderm (XEN)/
parietal endoderm cells (PEC) can be derived from the trophectoderm, inner cell mass (ICM) and primitive endoderm, respectively, and
propagate indefinitely in culture. In chimeras, TS, ES and XEN cells contribute to their lineage of origin. The trophectoderm will give rise
to the placenta; the ICM will form the three germ layers of the embryo and the primitive endoderm will develop into the visceral
endoderm and parietal endoderm lineages.
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
XEN cells as a model of endoderm development 303

mouse ES cells mimic XEN cell characteristics bothin vivo and in vitro (Fujikura et al. 2002; Shimosato
et al. 2007), implying that GATA factors play a crucial
role in XEN cell specification. Recently, an essential
stemness factor that has been identified in XEN cells is
SALL4. In fact, XEN cells cannot be derived from
Sall4-null mice. Given it lies upstream of GATA4 and
GATA6, SALL4 seems to play a role as an activator of
key lineage-defining genes in the ExEn. This partiallyexplains why loss of Gata4 or Gata6 leads to a VE
defect and not to a broader loss of primitive endoderm
(Lim et al. 2008).
Interestingly, human ES (hES) cells can also mimic
ExEn and definitive endoderm by constitutively
expressing SOX7 or SOX17, respectively (Seguin et al.
2008). In addition, differentiation of these cells by
either BMP4 treatment in SOX7 overexpressing hEScells increased expression of ExEn markers, whereas
Activin A treatment in SOX17 overexpressing hES cells
increased expression of definitive endoderm markers
as expected for hES cells. Both cell lines, however,
maintained expression of both NANOG and OCT4 in
their undifferentiated state. This suggests a similarity of
the SOX7 overexpressing hES to the XEN-P cells
rather than to the mouse XEN cells and implies a rolefor the SOX transcription factors in maintaining a pre-
cursor state in the cells preventing them from differen-
tiation. Sox17 is expressed in mouse-derived XEN
cells and plays a role in their establishment since XEN
cells cannot be derived from Sox17-null embryos.
However, in Sox17 mutant mice no significant effect
on ExEn development was observed, which the
authors attributed to the highly regulative environmentof the mouse embryo and the continuous expression
of Gata6 and Sox7 in Sox17-deficient embryos (Niakan
et al. 2010).
Endogenous non-coding small RNAs (miRNAs) play
a role in the pluripotency regulatory network of ES, TS
and XEN cells (Marson et al. 2008; Xu et al. 2009;
Spruce et al. 2010). Interestingly, Dicer mutants do not
correctly pattern the VE. In addition, depletion ofmiRNAs in XEN cells leads to a loss of multipotency,
which seems to be mediated via the modulation of the
ERK1/2 signaling pathway and is contrary to what
occurs in embryonic-derived cells. This suggests, for
the extraembryonic lineage, a crucial role for miRNAs
in establishment and maintenance of self-renewal
capability (Spruce et al. 2010). Furthermore, extra-
embryonic tissues have been reported to be hypome-thylated when compared to embryonic tissue
(Chapman et al. 1984; Monk et al. 1987; Gardner &
Davies 1992). Similarly, XEN cells have been recently
shown to express low levels of repressive chromatin
modifications, such as H3K27me3 (Rugg-Gunn et al.
2010). Low methylation of ExEn could potentially givethis tissue a significant competence to easily undergo
differentiation or transdifferentiation.
The lack of XEN cell contribution to VE has inter-
ested several groups and lead to the findings that
Nodal and BMP4, both members of the transforming
growth factor (TGF)b superfamily, treatments direct
XEN cells toward a VE phenotype in vivo and in vitro
(Kruithof-de Julio et al. 2011; Artus et al. 2012; Pacaet al. 2012). Nodal binds to type I (ALK4) or type II
receptors in the presence of the co-receptor Cripto or
Cryptic (members of the EGF-CFC family). Its signal is
propagated to the nucleus via phosphorylation of
SMAD2 and is tightly regulated by inhibitors including
Lefty and Cerberus. XEN cells, treated with either
Nodal or Cripto can be differentiated into a VE pheno-
type in vitro, and in chimeras they contribute moreefficiently to VE. This contribution, however, is not VE
exclusive, which could be due to the highly heteroge-
neous population of XEN cells. In XEN cells, Nodal
signals solely via the ALK4 receptor and the EGF-CFC
Cryptic, as both SB431542 (as inhibitor of the ALK4
receptor) treated XEN cells and XEN cells derived from
Cryptic-null mice do not respond to Nodal. Cripto is,
at least partially, Nodal-independent as its functioncannot be inhibited by SB431542. The authors pro-
pose two possible mechanistic scenarios: an alterna-
tive low affinity receptor or a non-canonical pathway
through an unknown signal transducer (Kruithof-de
Julio et al. 2011). Interestingly, similar conclusions
have been independently reached by Clements et al.
(2011). The authors focus on the cross talk between
the Nodal/Activin signaling pathway and the MAPKp38 in XEN cells revealing a novel role for p38 in regu-
lating Nodal thresholds. They observed that the activa-
tion of p38 is not ALK4, 5 or 7 dependent as it cannot
be inhibited by SB431542; however, it can be trig-
gered by Cripto. This suggests that the non-canonical
pathway by which Cripto functions may be p38
dependent (Clements et al. 2011).
As previously mentioned, BMP4 treatment alsodirects XEN cells to a VE phenotype. In this case
through the canonical BMP receptors, as the chemical
inhibitor Dorsomorphin prevented the VE induction
mediated by BMP4. Interestingly, the mainly induced
VE subtype resembles the VE adjacent to ExEc, and
not emVE (Artus et al. 2012; Paca et al. 2012). This
observation fits with the known expression of BMP4 in
the early embryo, which is highest in the ExEc closestto the proximal epiblast (Lawson et al. 1999). BMP4
was also capable of inducing VE differentiation of PE,
showing that this cell type is not terminally differenti-
ated, and retains the ability to form VE (Artus et al.
2012; Paca et al. 2012).
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
304 A. T. Moerkamp et al.

Conclusions
Multipotent stem cells have the potential to develop
into different cell types in the body during early life and
growth. They are distinguished from other cell types bytwo important characteristics. First, they can self-
renew and second, under certain physiologic or exper-
imental conditions, they can be induced to become
tissue- or organ-specific cells with special functions.
In order to fulfill the desire to generate a cell-based
therapy, it is necessary to manipulate the cells, in vitro,
to successfully differentiate them towards the cell-type
of interest. The meticulous application of developmen-tal principals to stem cell culture systems is the basis
of this kind of research. The major limitation is the very
low frequency of differentiated cells identified and the
cellular heterogeneity. Only when large numbers of
highly enriched progenitors are accessible, methods
can be defined for their maturation and their functional
capacity rigorously tested in animal models. XEN cells
might give the missing link in this differentiation pro-cess.
Extraembryonic and embryonic tissues interact tospecify each other’s commitment. By using XEN cells
as an inductive feeder layer for ES cells, epiblast
stem cells (EpiSC) or their multipotent progeny, a par-
ticular developmental or differentiation fate can be
induced (Kruithof-de Julio, Moerkamp and Goumans,
unpubl. data, 2012). In this context, XEN cells may
not only be a model to understand ExEn develop-
ment, but also to elucidate the inductive role of theExEn in embryonic tissue specification, for example,
the formation of heart and blood lineages (Brown
et al. 2010a; Artus et al. 2012). Once established,
their roles in these inductive processes, mutant lines
can be derived to pin point the specific genes
involved. Thereby, XEN cells may be a powerful
in vitro tool (Fig. 2), by either the use of conditioned
medium or co-culture, for providing an overview ofthe factors that support the differentiation during
developmental processes, like cardiogenesis and
hematopoiesis. The identification of the exogenous
differentiation stimuli may be translated into clinical
practice for stem cell-based approaches to efficiently
Fig. 2. Extraembryonic endoderm (XEN) cells, or its derived conditioned medium (CDM) may be a valuable in vitro tool to study develop-
mental processes including the communication between the visceral endoderm (VE) and mesoderm as well as these cells may have
therapeutic properties. Understanding the inductive factors underlying mesoderm specification, like cardiogenesis and hematopoiesis,
may be useful for stem cell-based therapeutic approaches.
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
XEN cells as a model of endoderm development 305

obtain a high number of the differentiated cell type ofinterest. For cell-based therapies, XEN cells are pref-
erably derived from human embryos. However, the
derivation from human embryos was only partly
supplemented by the SOX7 overexpressing hES cells
and remains subject to future studies. Alternatively,
for the identification of developmental stimuli, XEN
cells could be derived from the mouse postimplanta-
tion epiblast. Mouse EpiSC are highly similar to hEScells in their pluripotent state and growth factor
requirements and therefore its derived XEN cells may
resemble human ExEn development more closely.
Cho et al. (2012) was unable to derive XEN cells from
mouse EpiSC by using the same protocol as for the
derivation of XEN cells from mouse ES cells (Cho
et al. 2012). In the case of EpiSC, due to their epige-
netic state, XEN cell derivation may require differentexogenous stimuli. However, there remains the possi-
bility that EpiSC are indeed unable to convert their
developmental fate. Finally, although it has not been
shown to date, XEN cells may be derived from
somatic cells via direct reprogramming, thereby
circumventing the requirement of human embryos.
Although one must keep in mind the highly hetero-
geneous population and the existence of species-spe-cific differences, which may be due to the stage and
derivation protocols, XEN cells can also be a model to
study extraembryonic contribution to definitive endo-
derm and its differentiated derivatives. This raises the
question as to whether XEN cells per se can indeed
contribute to the primitive gut tube or must they be
prior differentiated towards a definitive endodermal
lineage. In later stage chimeras derived from Nodal,Cripto or untreated XEN cells contribution to definitive
endoderm has never been observed (Kruithof-de Julio
and Shen, unpubl. data, 2010), only the contribution
to the visceral yolk sac.
In summary, XEN cells provide a valuable cell culture
model to dissect the extraembryonic endodermal line-
age and its potential contribution to the embryonic
one. Thereby, XEN cells may increase our understand-ing of the molecular mechanism behind endoderm
behavior and function during development. Further-
more, XEN cells may give an insight into the inductive
factors underlying mesoderm specification, like cardio-
genesis and hematopoiesis, which may be reflected
into stem cell-based therapeutic approaches.
References
Artus, J., Douvaras, P., Piliszek, A., Isern, J., Baron, M. H. &Hadjantonakis, A. K. 2012. BMP4 signaling directs primitiveendoderm-derived XEN cells to an extraembryonic visceralendoderm identity. Dev. Biol. 361, 245–262.
Beddington, R. S. & Robertson, E. J. 1989. An assessment ofthe developmental potential of embryonic stem cells in themidgestation mouse embryo. Development 105, 733–737.
Brown, K., Doss, M. X., Legros, S., Artus, J., Hadjantonakis, A.K. & Foley, A. C. 2010a. eXtraembryonic ENdoderm (XEN)stem cells produce factors that activate heart formation.PLoS ONE 5, e13446.
Brown, K., Legros, S., Artus, J., Doss, M. X., Khanin, R., Had-jantonakis, A. K. & Foley, A. 2010b. A comparative analysisof extra-embryonic endoderm cell lines. PLoS ONE 5,e12016.
Byrd, N., Becker, S., Maye, P., Narasimhaiah, R., St-Jacques,B., Zhang, X., McMahon, J., McMahon, A. & Grabel, L.2002. Hedgehog is required for murine yolk sac angiogene-sis. Development 129, 361–372.
Chapman, V., Forrester, L., Sanford, J., Hastie, N. & Rossant, J.1984. Cell lineage-specific undermethylation of mouse repet-itive DNA. Nature 307, 284–286.
Chazaud, C., Yamanaka, Y., Pawson, T. & Rossant, J. 2006.Early lineage segregation between epiblast and primitiveendoderm in mouse blastocysts through the Grb2-MAPKpathway. Dev. Cell 10, 615–624.
Cho, L. T., Wamaitha, S. E., Tsai, I. J., Artus, J., Sherwood, R.I., Pedersen, R. A., Hadjantonakis, A. K. & Niakan, K. K.2012. Conversion from mouse embryonic to extra-embryonicendoderm stem cells reveals distinct differentiation capacitiesof pluripotent stem cell states. Development 139, 2866–2877.
Clements, M., Pernaute, B., Vella, F. & Rodriguez, T. A. 2011.Crosstalk between Nodal/activin and MAPK p38 signaling isessential for anterior-posterior axis specification. Curr. Biol.
21, 1289–1295.Damert, A., Miquerol, L., Gertsenstein, M., Risau, W. & Nagy, A.
2002. Insufficient VEGFA activity in yolk sac endoderm com-promises haematopoietic and endothelial differentiation.Development 129, 1881–1892.
de Sousa Lopes, S. M., Hayashi, K. & Surani, M. A. 2007. Proxi-mal visceral endoderm and extraembryonic ectoderm regu-late the formation of primordial germ cell precursors. BMC
Dev. Biol. 7, 140.de Sousa Lopes, S. M., Roelen, B. A., Monteiro, R. M., Em-
mens, R., Lin, H. Y., Li, E., Lawson, K. A. & Mummery, C.L. 2004. BMP signaling mediated by ALK2 in the visceralendoderm is necessary for the generation of primordial germcells in the mouse embryo. Genes Dev. 18, 1838–1849.
Debeb, B. G., Galat, V., Epple-Farmer, J., Iannaccone, S.,Woodward, W. A., Bader, M., Iannaccone, P. & Binas, B.2009. Isolation of Oct4-expressing extraembryonic endo-derm precursor cell lines. PLoS ONE 4, e7216.
Dyer, M. A., Farrington, S. M., Mohn, D., Munday, J. R. & Baron,M. H. 2001. Indian hedgehog activates hematopoiesis andvasculogenesis and can respecify prospective neurectoder-mal cell fate in the mouse embryo. Development 128, 1717–1730.
Fowler, K. J., Mitrangas, K. & Dziadek, M. 1990. In vitro produc-tion of Reichert’s membrane by mouse embryo-derived pari-etal endoderm cell lines. Exp. Cell Res. 191, 194–203.
Fujikura, J., Yamato, E., Yonemura, S., Hosoda, K., Masui, S.,Nakao, K., Miyazaki Ji, J. & Niwa, H. 2002. Differentiation ofembryonic stem cells is induced by GATA factors. Genes
Dev. 16, 784–789.Galat, V., Binas, B., Iannaccone, S., Postovit, L. M., Debeb, B.
G. & Iannaccone, P. 2009. Developmental potential of ratextraembryonic stem cells. Stem Cells Dev. 18, 1309–1318.
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
306 A. T. Moerkamp et al.

Gardner, R. L. & Davies, T. J. 1992. Environmental factors andthe stability of differentiation in mammalian development. C.R. Acad. Sci. III, Sci. Vie 314, 67–69.
Grabarek, J. B., Zyzynska, K., Saiz, N., Piliszek, A., Frankenberg,S., Nichols, J., Hadjantonakis, A. K. & Plusa, B. 2012. Differ-ential plasticity of epiblast and primitive endoderm precur-sors within the ICM of the early mouse embryo.Development 139, 129–139.
Granier, C., Gurchenkov, V., Perea-Gomez, A., Camus, A., Ott,S., Papanayotou, C., Iranzo, J., Moreau, A., Reid, J., Koent-ges, G., Saberan-Djoneidi, D. & Collignon, J. 2011. Nodalcis-regulatory elements reveal epiblast and primitive endo-derm heterogeneity in the peri-implantation mouse embryo.Dev. Biol. 349, 350–362.
Hogan, B. L., Cooper, A. R. & Kurkinen, M. 1980. Incorporationinto Reichert’s membrane of laminin-like extracellular pro-teins synthesized by parietal endoderm cells of the mouseembryo. Dev. Biol. 80, 289–300.
Kruithof-de Julio, M., Alvarez, M. J., Galli, A., Chu, J., Price, S.M., Califano, A. & Shen, M. M. 2011. Regulation of extra-embryonic endoderm stem cell differentiation by Nodal andCripto signaling. Development 138, 3885–3895.
Kunath, T., Arnaud, D., Uy, G. D., Okamoto, I., Chureau, C.,Yamanaka, Y., Heard, E., Gardner, R. L., Avner, P. & Ros-sant, J. 2005. Imprinted X-inactivation in extra-embryonicendoderm cell lines from mouse blastocysts. Development
132, 1649–1661.Kwon, G. S., Viotti, M. & Hadjantonakis, A. K. 2008. The endo-
derm of the mouse embryo arises by dynamic widespreadintercalation of embryonic and extraembryonic lineages. Dev.Cell 15, 509–520.
Lawson, K. A., Dunn, N. R., Roelen, B. A., Zeinstra, L. M., Davis,A. M., Wright, C. V., Korving, J. P. & Hogan, B. L. 1999.Bmp4 is required for the generation of primordial germ cellsin the mouse embryo. Genes Dev. 13, 424–436.
Lim, C. Y., Tam, W. L., Zhang, J., Ang, H. S., Jia, H., Lipovich,L., Ng, H. H., Wei, C. L., Sung, W. K., Robson, P., Yang, H.& Lim, B. 2008. Sall4 regulates distinct transcription circuit-ries in different blastocyst-derived stem cell lineages. Cell
Stem Cell 3, 543–554.Madabhushi, M. & Lacy, E. 2011. Anterior Visceral Endoderm
Directs Ventral Morphogenesis and Placement of Head andHeart via BMP2 Expression. Dev. Cell 21, 907–919.
Marson, A., Levine, S. S., Cole, M. F., Frampton, G. M., Bram-brink, T., Johnstone, S., Guenther, M. G., Johnston, W. K.,Wernig, M., Newman, J., Calabrese, J. M., Dennis, L. M.,Volkert, T. L., Gupta, S., Love, J., Hannett, N., Sharp, P. A.,Bartel, D. P., Jaenisch, R. & Young, R. A. 2008. ConnectingmicroRNA genes to the core transcriptional regulatory cir-cuitry of embryonic stem cells. Cell 134, 521–533.
McGrath, K. E. & Palis, J. 2005. Hematopoiesis in the yolk sac:more than meets the eye. Exp. Hematol. 33, 1021–1028.
Migeotte, I., Omelchenko, T., Hall, A. & Anderson, K. V. 2010.Rac1-dependent collective cell migration is required forspecification of the anterior-posterior body axis of themouse. PLoS Biol. 8, e1000442.
Monk, M., Boubelik, M. & Lehnert, S. 1987. Temporal and regio-nal changes in DNA methylation in the embryonic, extraem-bryonic and germ cell lineages during mouse embryodevelopment. Development 99, 371–382.
Morrisey, E. E., Tang, Z., Sigrist, K., Lu, M. M., Jiang, F., Ip, H.S. & Parmacek, M. S. 1998. GATA6 regulates HNF4 and isrequired for differentiation of visceral endoderm in the mouseembryo. Genes Dev. 12, 3579–3590.
Niakan, K. K., Ji, H., Maehr, R., Vokes, S. A., Rodolfa, K. T.,Sherwood, R. I., Yamaki, M., Dimos, J. T., Chen, A. E.,Melton, D. A., McMahon, A. P. & Eggan, K. 2010. Sox17promotes differentiation in mouse embryonic stem cells bydirectly regulating extraembryonic gene expression andindirectly antagonizing self-renewal. Genes Dev. 24, 312–326.
Paca, A., Seguin, C. A., Clements, M., Ryczko, M., Rossant, J.,Rodriguez, T. A. & Kunath, T. 2012. BMP signaling inducesvisceral endoderm differentiation of XEN cells and parietalendoderm. Dev. Biol. 361, 90–102.
Rivera-Perez, J. A., Mager, J. & Magnuson, T. 2003. Dynamicmorphogenetic events characterize the mouse visceral endo-derm. Dev. Biol. 261, 470–487.
Rossant, J. 2007. Stem cells and lineage development in themammalian blastocyst. Reprod. Fertil. Dev. 19(Suppl. 1), 111–118.
Rugg-Gunn, P. J., Cox, B. J., Ralston, A. & Rossant, J. 2010.Distinct histone modifications in stem cell lines and tissue lin-eages from the early mouse embryo. Proc. Natl Acad. Sci.USA 107, 10783–10790.
Seguin, C. A., Draper, J. S., Nagy, A. & Rossant, J. 2008. Estab-lishment of endoderm progenitors by SOX transcription fac-tor expression in human embryonic stem cells. Cell Stem
Cell 3, 182–195.Shimosato, D., Shiki, M. & Niwa, H. 2007. Extra-embryonic
endoderm cells derived from ES cells induced by GATA fac-tors acquire the character of XEN cells. BMC Dev. Biol. 7,80.
Spruce, T., Pernaute, B., Di-Gregorio, A., Cobb, B. S., Mer-kenschlager, M., Manzanares, M. & Rodriguez, T. A. 2010.An early developmental role for miRNAs in the maintenanceof extraembryonic stem cells in the mouse embryo. Dev. Cell19, 207–219.
Srinivas, S., Rodriguez, T., Clements, M., Smith, J. C. & Bedd-ington, R. S. 2004. Active cell migration drives the unilateralmovements of the anterior visceral endoderm. Development
131, 1157–1164.Stuckey, D. W., Clements, M., Di-Gregorio, A., Senner, C. E., Le
Tissier, P., Srinivas, S. & Rodriguez, T. A. 2011. Coordina-tion of cell proliferation and anterior-posterior axisestablishment in the mouse embryo. Development 138,1521–1530.
Takaoka, K., Yamamoto, M. & Hamada, H. 2011. Origin and roleof distal visceral endoderm, a group of cells that determinesanterior-posterior polarity of the mouse embryo. Nat. Cell
Biol. 13, 743–752.Takaoka, K., Yamamoto, M., Shiratori, H., Meno, C., Rossant, J.,
Saijoh, Y. & Hamada, H. 2006. The mouse embryo autono-mously acquires anterior-posterior polarity at implantation.Dev. Cell 10, 451–459.
Takito, J. & Al-Awqati, Q. 2004. Conversion of ES cells to colum-nar epithelia by hensin and to squamous epithelia by laminin.J. Cell Biol. 166, 1093–1102.
Tanaka, S., Kunath, T., Hadjantonakis, A. K., Nagy, A. & Ros-sant, J. 1998. Promotion of trophoblast stem cell prolifera-tion by FGF4. Science 282, 2072–2075.
Thomas, P. & Beddington, R. 1996. Anterior primitive endodermmay be responsible for patterning the anterior neural plate inthe mouse embryo. Curr. Biol. 6, 1487–1496.
Toles, J. F., Chui, D. H., Belbeck, L. W., Starr, E. & Barker, J. E.1989. Hemopoietic stem cells in murine embryonic yolk sacand peripheral blood. Proc. Natl Acad. Sci. USA 86, 7456–7459.
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
XEN cells as a model of endoderm development 307

Torres-Padilla, M. E., Richardson, L., Kolasinska, P., Meilhac, S.M., Luetke-Eversloh, M. V. & Zernicka-Goetz, M. 2007. Theanterior visceral endoderm of the mouse embryo is establishedfrom both preimplantation precursor cells and by de novogene expression after implantation. Dev. Biol. 309, 97–112.
Trichas, G., Joyce, B., Crompton, L. A., Wilkins, V., Clements,M., Tada, M., Rodriguez, T. A. & Srinivas, S. 2011. Nodal
dependent differential localisation of dishevelled-2 demar-cates regions of differing cell behaviour in the visceral endo-derm. PLoS Biol. 9, e1001019.
Xu, N., Papagiannakopoulos, T., Pan, G., Thomson, J. A. & Kos-ik, K. S. 2009. MicroRNA-145 regulates OCT4, SOX2, andKLF4 and represses pluripotency in human embryonic stemcells. Cell 137, 647–658.
ª 2013 The Authors
Development, Growth & Differentiation ª 2013 Japanese Society of Developmental Biologists
308 A. T. Moerkamp et al.