Embed Size (px)
Citation preview

J. Cell Sci. 25, 313-322 (1977) 313Printed in Great Britain
EXTRACELLULAR MATRIX SYNTHESIS IN
BLASTULA AND GASTRULA STAGES OF
NORMAL AND HYBRID FROG EMBRYOS
I. TOLUIDINE BLUE AND LANTHANUM STAINING
KURT E.JOHNSONBox 3011, Department of Anatomy, Duke University Medical Center,Durham, North Carolina 27710, U.S.A.
SUMMARY
Four developmental stages of Rana pipiens, R. pipiens $ x R. catesbeiana <J, and R. pipiens$ x JR. esculenta 3 embryos were fixed in standard electron microscopic fixatives with or without1 % lanthanum, nitrate (LN) added. Embryos fixed without LN were embedded and sectionedat 05 fim and stained with toluidine blue. Embryos fixed with LN were embedded andsectioned at 70-80 nm and examined in the electron microscope. Both techniques show adramatic increase in the amount of stainable material in the extracellular matrix during develop-ment. Blastula stage embryos have a small amount of material in the extracellular matrix. Earlygastrulae have more stainable material than blastulae and this increased amount appears to bedue to an accumulation of material in the marginal zone of the embryo, especially near thedorsal lip of the blastopore. Late gastrulae have large amounts of stainable material in theextracellular spaces in almost all parts of the embryo. Interspecific arrested hybrid embryosdo not show the same dramatic accumulation of stainable material in their extracellular matrices.
INTRODUCTION
During gastrulation, morphogenetic cell movements lead to a complete rearrange-ment of the embryo. These cell movements occur in a well defined manner, suggestingthat there are intricate control mechanisms to guide cell movements along certainpaths. At present, we have little information about these control processes, but oneimportant possibility for control is in the dynamic properties of the extracellularmatrix (Manasek, 1975). Several studies indicate that there are changes in the com-position of the extracellular matrix that occur during gastrulation in amphibianembryos. Tarin (1971, 1973) showed that there is a hyaluronidase-sensitive, toluidineblue metachromatic material between invaginating chordamesodermal cells inXenopus laevis. Kosher & Searles (1973) have shown that Rana pipiens embryos areactively engaged in sulphated glycosaminoglycan (GAG) synthesis during earlydevelopment. During the blastula period, the predominant sulphated GAG syn-thesized is heparin and/or heparan sulphate with a minor amount of chondroitin-4-sulphate also being made. Beginning at the early gastrula stage, the rate of synthesisof chondroitin-6-sulphate appears to increase. Invaginating chordamesodermal cellsare particularly active in sulphated GAG synthesis. The present study was initiated
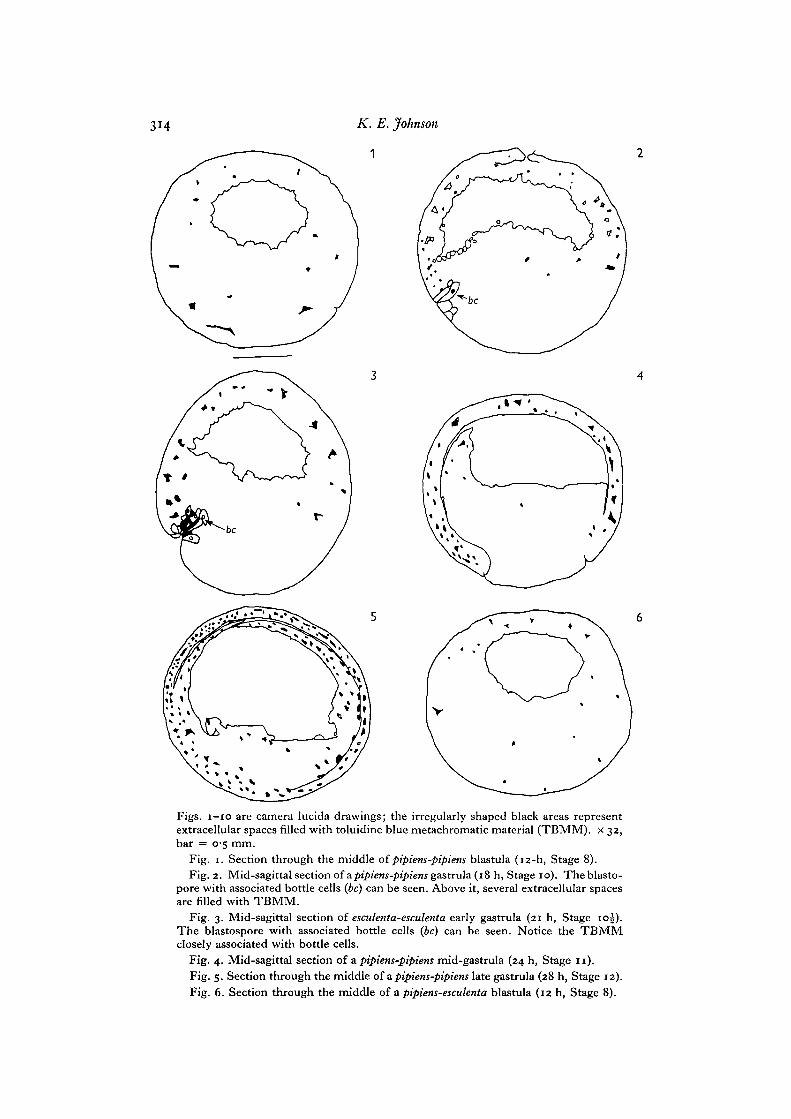
Figs, I - IO are camera lucida drawings; the irregularly shaped black areas representextracellular spaces filled with toluidine blue metachromatic material (TBMM). x 32,bar = 0-5 mm.
Fig. 1. Section through the middle of pipiens-pipiens blastula (12-h, Stage 8).Fig. 2. Mid-sagittal section of a pipiens-pipiens gastrula (18 h, Stage 10). The blasto-
pore with associated bottle cells (be) can be seen. Above it, several extracellular spacesare filled with TBMM.
Fig. 3. Mid-sagittal section of esculenta-esculenta early gastrula (21 h, Stage 10J).The blastospore with associated bottle cells (be) can be seen. Notice the TBMMclosely associated with bottle cells.
Fig. 4. Mid-sagittal section of a pipiens-pipiens mid-gastrula (24 h, Stage 11).Fig. 5. Section through the middle of a pipiens-pipiens late gastrula (28 h, Stage 12).Fig. 6. Section through the middle of a pipiens-esculenta blastula (12 h, Stage 8).
Frog gastrula extracellular matrix, I 315
to compare changes in the composition of the extracellular matrix in developing Ranapipiens embryos and interspecific arrested hybrid embryos. The results of this studysuggest that changes in the composition of the extracellular matrix are involved inmorphogenesis in some way.
MATERIALS AND METHODS
Ratiapipiens, R. catesbeiana, andi?. esculenta were obtained from Nasco (Fort Atkinson, Wis.).Females were ovulated as described previously (Johnson, 1970). /?. esculenta ovulates in response
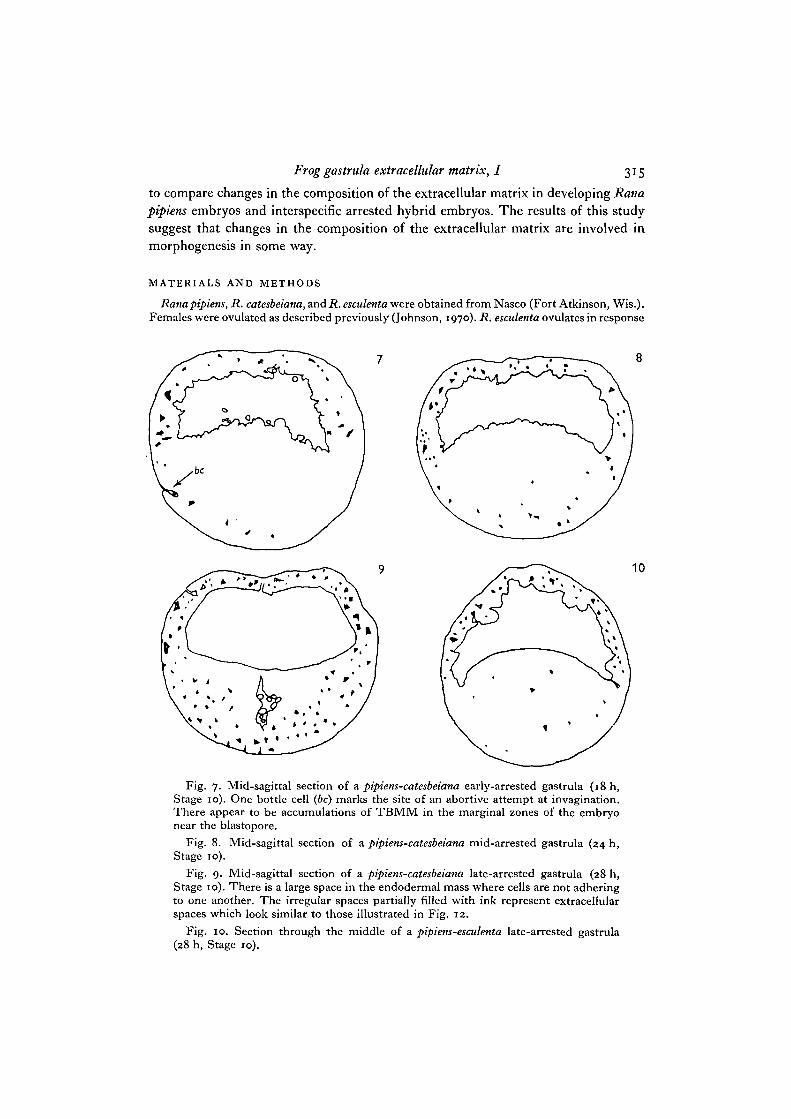
Fig. 7. Mid-sagittal section of a pipiens-catesbeiana early-arrested gastrula (18 h,Stage 10). One bottle cell (6c) marks the site of an abortive attempt at invagination.There appear to be accumulations of TBMM in the marginal zones of the embryonear the blastopore.
Fig. 8. Mid-sagittal section of a pipiens-catesbeiana mid-arrested gastrula (24 h,Stage 10).
Fig. 9. Mid-sagittal section of a pipiens-catesbeiana late-arrested gastrula (28 h,Stage 10). There is a large space in the endodermal mass where cells are not adheringto one another. The irregular spaces partially filled with ink represent extracellularspaces which look similar to those illustrated in Fig. 12.
Fig. 10. Section through the middle of a pipiens-esculenta late-arrested gastrula(28 h, Stage 10).

316 K. E. Johnson
to either R. esculenta or to R. pipiens pituitary-injections during January and February. Jellycoats were removed from embryos by a 15 min treatment with 0-7 % mercaptoacetic acid in50% modified Steinberg's solution (Johnson, 1970) with the pH adjusted to 8-6 with 5 NNaOH.After this treatment, embryos were rinsed 10 times with 10% Steinberg's solution and rearedin this solution at 2i-23°C. Embryos were staged according to Shumway (1940).
Whole dejellied embryos with vitelline membranes intact were fixed essentially as describedearlier (Johnson, 1970) except that in some instances 1 % lanthanum nitrate [La(NO3)3.6H2O,Fisher] (LN) was added to the glutaraldehyde and osmium tetroxide and fixation times werelonger because large fragments or whole embryos were fixed. Thus, glutaraldehyde fixation was4 h, osmium tetroxide was 16 h, and uranyl acetate was 14 h. Embryos not treated with LNwere fixed whole. Embryos treated with LN had vitelline membranes removed during glutar-aldehyde fixation and were cut into large fragments with watchmaker's forceps. After fixationfragments were dehydrated in ethanol, embedded in Epon 812, and sectioned at either 0-5 /imand stained with 1 % toluidine blue in 1% K2B4O,.4H2O or 70-80 nm and stained with leadcitrate. To illustrate the regional distribution of toluidine blue metachromasia in embryos,mid-sagittal sections were traced with the aid of a camera lucida and then extracellular spaceswith extensive metachromasia were marked on the tracing.
In what follows, embryos are named by giving the species of the female followed by thespecies of the male, e.g. the arrested hybrid formed by fertilizing R. pipiens eggs with R.catesbeiana sperm is called pipiens-catesbeiana.
RESULTS
Accumulation of toluidine blue metachromatic material in the extracellular matrix duringdevelopment
From the blastula (Stage 8) to the late gastrula (Stage 12) inpipiens-pipiens embryos,there is an obvious increase in the total amount of toluidine blue metachromaticmaterial (TBMM) in the extracellular matrix (Figs. 1, 2, 4, 5, and Table 1). There isalso a similar but less dramatic trend in esculenta-esculenta embryos. In blastulae andearly gastrulae, only scattered extracellular spaces contain TBMM. By the mid-gastrula stage, many extracellular spaces are either partially or completely filled byTBMM. In the blastula and early gastrula arrest stages of pipiens-catesbeiana andpipiens-esculenta most spaces have no TBMM (Figs. 6, 7). The amount of TBMM inmid- and late-gastrula attest pipiens-esculenta embryos is less than that seen in pipiens-pipiens or pipiens-catesbeiana (compare Figs. 5, 9, 10).
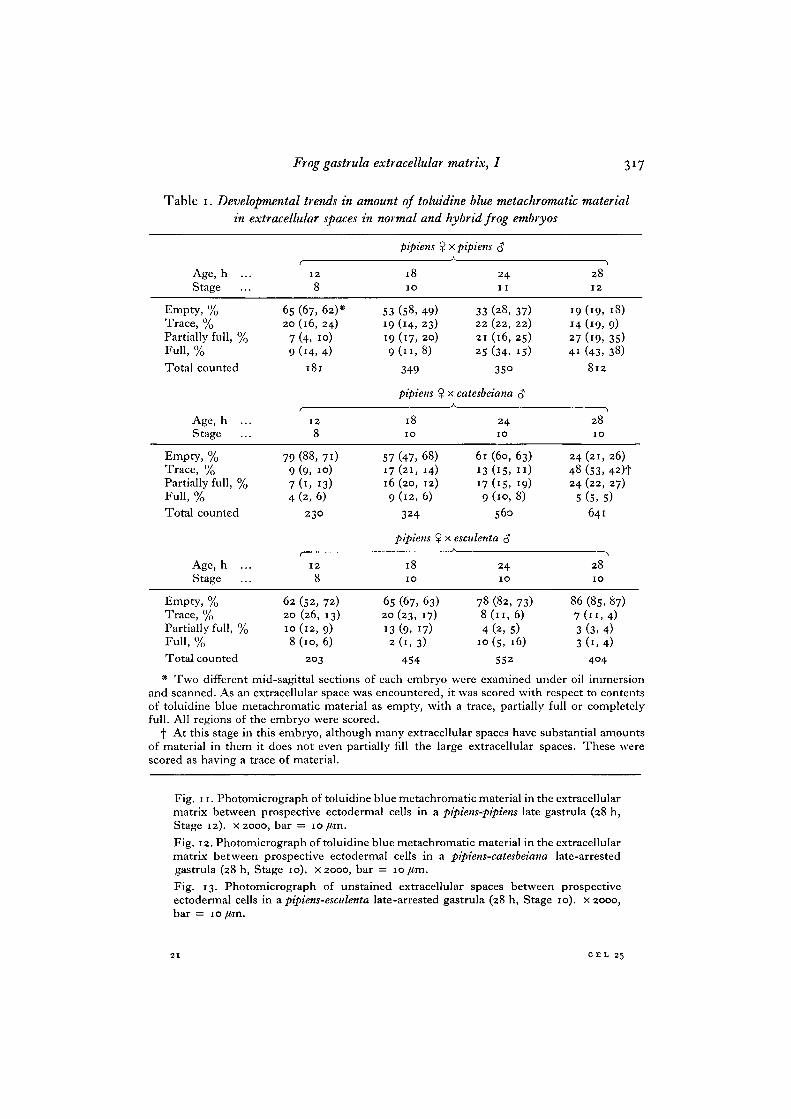
A measurement of the percentage of the total number of extracellular spacescontaining TBMM was made. Sections of embryos were scanned and spaces betweencells in these sections were scored for relative content of TBMM (Table 1).
In many locations in pipiens-catesbeiana embryos there are large extracellular spacesand these contain some TBMM which is not enough to fill them (Fig. 12). Such spaceswere scored as containing a trace of TBMM. This task was undertaken because thetotal number of extracellular spaces and the total volume of the extracellular spacesincreases during development as the number of cells increases. The results of thisquantitation show that there is an increase in all type of embryos in the number ofextracellular spaces which contain TBMM. Normal embryos also show a dramaticincrease in the percentage of spaces with large amounts of TBMM while arrestedhybrid embryos show either a slight increase (pipiens-catesbeiana) or a slight decrease(pipiens-esculenta) in the percentage of spaces with large amounts of TBMM.

K. E. Johnson
i^ ^m^^ ^ ^ * ^ ^ « • ' .̂ / « ^ *^^r « F ^ A ^
(Facing p. 317)
Frog gastrula extracellular matrix, I 3*7
Table i. Developmental trends in amount of toluidine blue metachromatic materialin extracellular spaces in normal and hybrid frog embryos
Age, h ...Stage
Empty, %Trace, %Partially full, %Full, %
Total counted
Age, h ...Stage
Empty, %Trace, %Partially full, %Full, %
Total counted
Age, h ...Stage
6520
79
79974
128
(67, 62)*(16, 24)(4. 10)(14, 4)
181
128
(88, 71)(9, 10)(1, 13)(2,6)
2 3 0
128
5319199
5717169
pipiens %
1810
(58, 49)(14, 23)(17, 20)
349
pipiens $
1810
(47, 68)(21, 14)(20, 12)(12, 6)
324
pipiens $
1810
I x pipiens <J
24I I
33 (28, 37)22 (22, 22)21 (16, 25)25 (34, 15)
35°
! x catesbeiana 3A
24IO
6l (60, 63)13 (15, I I )17 (15, 19)9 (10, 8)
560
x esculenta c?
2410
14274i
244824
5
2812
(19, 18)(19, 9)(19, 35)(43, 38)
8 1 2
2810
(21, 26)
(53, 42)t(22, 27)
(5,5)641
2810
Empty, %Trace, %Partially full, %Full, %Total counted
62 (52, 72)20 (26, 13)10(12, 9)8 (10, 6)
203
65 (67, 63)20 (23, 17)13 (9, 17)2(1 ,3)
454
78 (82, 73)8(11,6)
4 (2, 5)10 (s, 16)
552
86 (85, 87)7 ( n , 4)3 (3, 4)3 (1, 4)
404
* Two different mid-sagittal sections of each embryo were examined under oil immersionand scanned. As an extracellular space was encountered, it was scored with respect to contentsof toluidine blue metachromatic material as empty, with a trace, partially full or completelyfull. All regions of the embryo were scored.
f At this stage in this embryo, although many extracellular spaces have substantial amountsof material in them it does not even partially fill the large extracellular spaces. These werescored as having a trace of material.
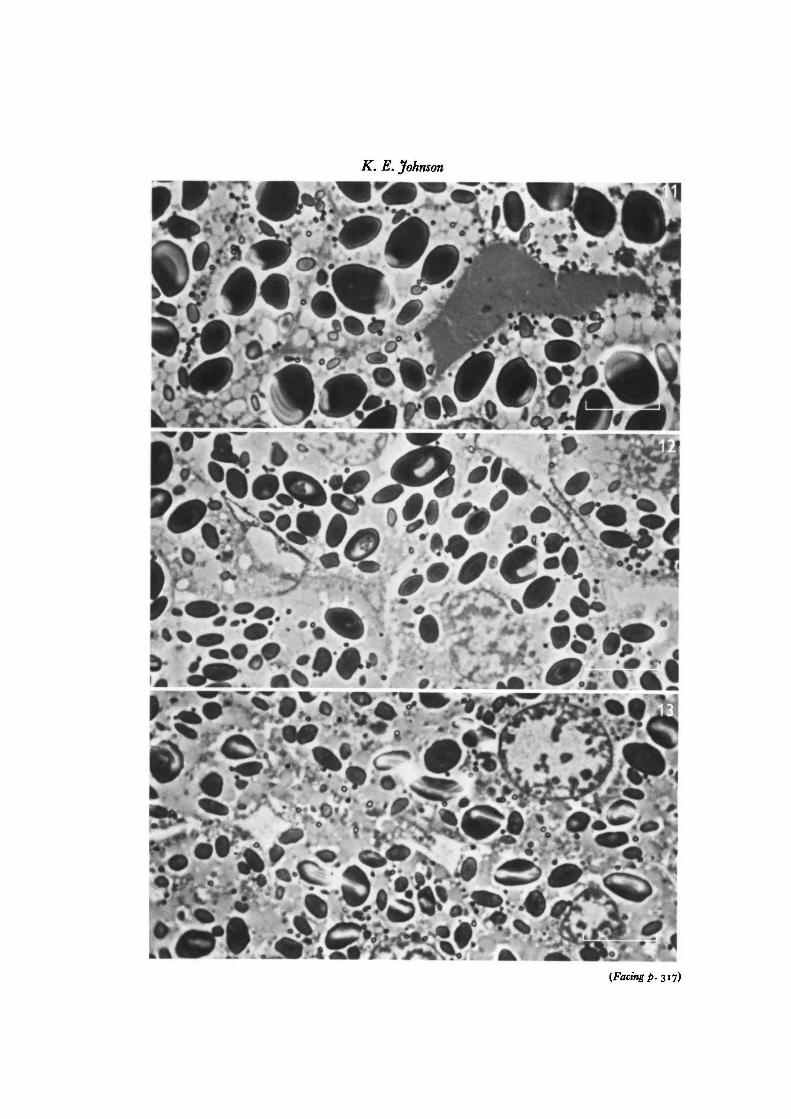
Fig. 11. Photomicrograph of toluidine blue metachromatic material in the extracellularmatrix between prospective ectodermal cells in a pipiens-pipiens late gastrula (28 h,Stage 12). x 2000, bar = io/im.
Fig. 12. Photomicrograph of toluidine blue metachromatic material in the extracellularmatrix between prospective ectodermal cells in a pipiens-catesbeiana late-arrestedgastrula (28 h, Stage 10). x 2000, bar = 10 /tm.
Fig. 13. Photomicrograph of unstained extracellular spaces between prospectiveectodermal cells in a pipiens-esculenta late-arrested gastrula (28 h, Stage 10). x 2000,bar =
C E L 25

318 K.E.Johnson
Regional distribution of toluidine blue metachromatic material in the extracellular matrix
All blastula stage embryos have a scattering of TBMM in extracellular spaces in allregions (Figs, i, 6). There is no obvious concentration of staining material in anyregion of the embryo. In contrast, normal early pipiens-pipiens and esculenta-esculentagastrulae and early arrested pipiens-catesbeiana gastrulae have a clustered group ofextracellular spaces containing TBMM in the marginal zones of the embryo aroundthe dorsal lip of the blastopore (Figs. 2, 3, 7). The bulbous ends of necks of bottlecells are often embedded in a network of TBMM (Fig. 3). Early arrested pipiens-esculenta gastrulae have no accumulation of TBMM in the marginal zone. Bothpipiens-pipiens and pipiens-catesbeiana show definite blastopore formation and anaccumulation of TBMM near the dorsal lip of the blastopore whereaspipiens-esculentashows no blastopore formation and no accumulation of TBMM in the marginal zoneof the embryo.
In the pipiens-pipiens mid-gastrula (Stage 11) there are large extracellular spaceswith TBMM near the animal pole, as well as in the spreading prospective ectoderm,neural plate, and involuting mesoderm. The tip of the advancing archenteron hasnumerous spaces with TBMM, but little occurs between cells in the central endodermalmass (Fig. 4). In the pipiens-catesbeiana mid-arrested gastrula there appears to be anaccumulation of TBMM in all parts of the embryo except the deep parts of theprospective endoderm (Fig. 8) and this accumulation continues in late-arrestedgastrulae (Fig. 9). In many locations, extracellular spaces contain relatively smallamounts of TBMM which appears flocculent and is restricted to peripheral portionsof cells (Figs. 9, 12), rather than filling the extracellular space as is frequently seen inpipiens-pipiens embryos of the same age. In other locations, extracellular spaces arefilled with TBMM. No blastopore formation occurs in the pipiens-esculenta embryoand therefore it is not possible to determine the mid-sagittal plane. In the sectionswhich were made through the maximum diameter of the whole embryo, there is noapparent regional concentration of extracellular spaces with TBMM except that thereare more spaces with TBMM between cells of the roof of the blastocoel than betweencells of the vegetal yolk mass. The most striking observation is that there are manyfewer extracellular spaces with TBMM in pipiens-esculenta embryos after the earlygastrula arrest stage when compared with either pipiens-catesbeiana or pipiens-pipiens.
Accumulation of lanthanum-staining material in the extracellular matrix duringdevelopment
In all pregastrula stages of normal and hybrid embryos there is a small amount oflanthanum-staining material (LSM) in the extracellular matrix. In scattered locations,
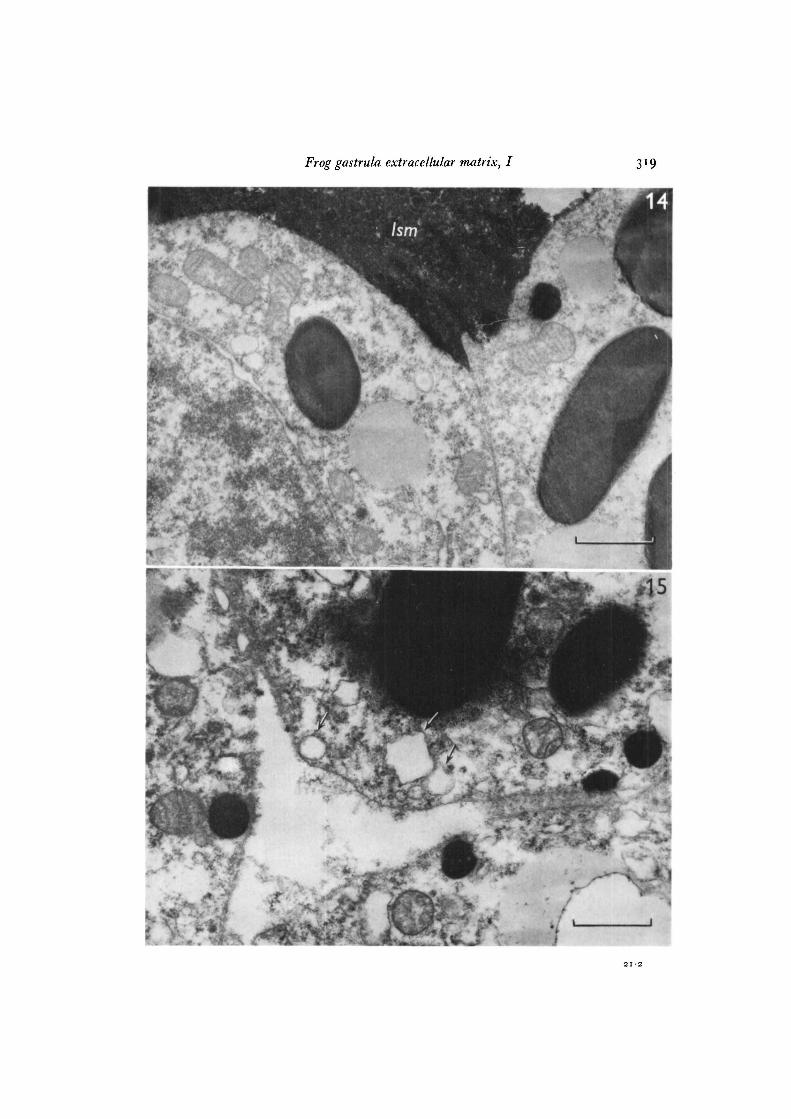
Fig. 14. Electron micrograph of a space between prospective ectodermal cells inpipiens-pipiens late gastrula (28 h, Stage 12). There is a large amount of lanthanum-staining material (Ism) between the cells, x 20000, bar = i-o /tm.
Fig. 15. Electron micrograph of a space between cells in the roof of the blastocoel ina pipiens-catesbeiana late-arrested gastrula (28 h, Stage 10). Notice the sparseness of the/SOT and the membrane-bound vesicles (arrows) in the cell cortex, x 20000, bar= i-o/tm.
Frog gastrula extracellular matrix, I

320 K. E. Johnson
small extracellular spaces have LSM in them but most are empty. In addition, cellsurfaces occasionally have a layer of LSM on them which varies from 35 to 140 nmin thickness. More commonly, however, long stretches of the cell surface are unstained.
There is an obvious increase in the amount of LSM in the extracellular spaces andalong the surfaces of cells in pipiens-pipiens gastrulae. In many locations, extracellularspaces are either partially or completely filled with LSM (Fig. 14). Cell surfaces aresometimes coated with a layer from 30 to 150 nm thick which appears to bridge thegap between cells. The amount of LSM increases throughout gastrulation in pipiens-pipiens. The same results are found in developing Xenopus laevis embryos, (Johnson,
1977)-There is also an increase in the amount of LSM detected in pipiens-catesbeiana
embryos during the 12-h period after arrest at the early gastrula stage, but it is not asdramatic as that seen in pipiens-pipiens. Many extracellular spaces are encounteredwhich contain LSM in amounts which are not sufficient to fill the extracellular space.Instead, the LSM adheres to cell surfaces facing an extracellular space or appears asa sparse flocculent material in the matrix. The appearance of the LSM in the electronmicroscope is quite similar to the appearance of the TBMM in the light microscope(compare Figs. 12 and 15). In some locations, dense accumulations of LSM fillextracellular spaces. The frequency of such full spaces is much less than that seen inpipiens-pipiens. In late pipiens-catesbeiana arrested gastrulae, especially near locationswith small amounts of extracellular LSM, there are many membrane-bound vesiclesin the peripheral cytoplasm (Fig. 15).
Pipiens-esculenta embryos have very little LSM in their extracellular spaces at anydevelopmental stage. In a few locations, LSM can be seen coating cell surfaces or inextracellular spaces. There is no dramatic increase in the total amount of LSM seenin the electron microscope and there is an obvious decrease in the number of spaceswhich contain enough LSM to partially or completely fill them.
DISCUSSION
Two points can be made from these results. First, there is a dramatic increase inthe total amount of extracellular matrix material during gastrulation in pipiens-pipiens.This increase first begins in close proximity to the initial site of invagination. Asgastrulation proceeds there is a further accumulation of matrix material in groups ofcells actively involved in the morphogenetic cell movements of involution, epiboly,and archenteron formation. By the end of gastrulation, large amounts of matrix materialhave accumulated in nearly all locations in the embryo. Second, interspecific hybridembryos that undergo developmental arrest at the onset of gastrulation show defectsin the accumulation of matrix materials. One hybrid [pipiens-catesbeiana) accumulatesrelatively large amounts of matrix material in a regional pattern that is similar to thepattern of accumulation in normal embryos. In this hybrid, however, extracellularspaces usually are not filled with matrix material. Rather, cell surfaces have a coat offlocculent material. Another kind of hybrid {pipiens-esculenta) accumulates very littlematrix material. The results of the present study are in agreement with similar

Frog gastrula extracellular matrix, I 321
histochemical observations on gastrula stage Xenopus embryos (Tarin, 1971, 1973;Johnson, 1977).
Comparative investigations of cell surface adhesiveness in normal and arrestedhybrid embryos have demonstrated that in normal embryos, cell surface adhesivenessbegins to change prior to the onset of gastrulation and continues to change duringgastrulation; these changes fail to occur in arrested hybrid embryos (Johnson, 1970,1972). For example, normal blastulae disaggregate rapidly, normal early gastrulaedisaggregate at an intermediate rate, and normal late gastrulae disaggregate slowlyin Ca2+, Mg2+-free solutions. In contrast, all stages of arrested hybrid embryos dis-aggregate with a high, blastula-like rate. It may be that the accumulating constituentsof the extracellular matrix bind cells together in some fashion during gastrulation andmake the embryos progressively resistant to disaggregation. In addition, the defectivematrix accumulation seen in arrested hybrid embryos may account for the lack ofchanges in cell adhesiveness and morphogenetic defects observed in earlier studies.
What could be the function of new matrix components that appear during gastru-lation ? The appearance of new components, leading to changes in the bulk compositionof the extracellular matrix, may provide a substratum for cell locomotion. There arestrong indications that glycosaminoglycan synthesis is important in the initiation ofcell movements in gastrulating sea-urchin embryos (Immers, 1961; Sugiyama, 1972;Karp & Solursh, 1974) and chick embryos (Manasek, 1975; Solursh, 1976). Ina number of other developmental systems, there appears to be a correlation betweenthe synthesis of glycosaminoglycans and the onset of cell migration (Toole, 1973;Pratt, Goggins, Wilk & King, 1973; Pratt, Larsen & Johnston, 1975). The presentstudy shows that in normal frog embryos, new components are added to the matrixat the time and place of the initiation of cell movements. In addition, it has beenshown that arrested hybrid embryos which are defective in morphogenetic cellmovements are also defective in their capacity to add new components to the extra-cellular matrix. Autoradiographic experiments are in progress to determine the sitesof synthesis and modes of transport of matrix components within cells. In addition,a biochemical analysis of newly synthesized matrix components is in progress.
This work was supported by NIH Grant HD 07082. Dr M. K. Reedy allowed use of equip-ment in his laboratory. Drs S. Roth and S. J. Counce offered helpful advice on this manuscript.The author thanks these people.
REFERENCES
IMMERS, J. (1961). Comparative study of the localization of incorporated 14C-labeled aminoacids and 36SO4 in the sea urchin ovary, egg, and embryo. Expl Cell Res 24, 356-378.
JOHNSON, K. E. (1970). The role of changes in cell contact behaviour in amphibian gastrulation.J. exp. Zool. 175, 391-428.
JOHNSON, K. E. (1972). The extent of cell contact and the relative frequency of small and largegaps between presumptive mesodermal cells in normal gastrulae of Rana pipiens and thearrested gastrulae of the Rana pipiens tyxRana catesbeiana $ hybrid. J. exp. Zool. 179,227-238.
JOHNSON, K. E. (1977). Changes in the cell coat at the onset of gastrulation in Xenopus laevisembryos. J. exp. Zool. 199, 137-142.
KARP, G. C. & SOLURSH, M. (1974). Acid mucopolysaccharide metabolism, the cell surface,and primary mesenchyme cell activity in the sea urchin embryo. Devi Biol. 41, 110-123.

322 K. E. Johnson
KOSHER, R. A. & SEARLES, R. L. (1973). Sulfated mucopolysaccharide synthesis during thedevelopment of Rana pipiens. Devi Biol. 32, 50-68.
MANASEK, F. J. (1975)- The extracellular matrix: a dynamic component of the developingembryo. Curr. Top. dev. Biol. 10, 35-102.
PRATT, R. M., GOGGINS, J. F., WILK, A. L. & KING, C. T. G. (1973). Acid mucopolysaccharidesynthesis in the secondary palate of the developing rat at the time of rotation and fusion.Devi Biol. 32, 230-237.
PRATT, R. M., LARSEN, M. A. & JOHNSTON, M. C. (1975). Migration of cranial neural crest cellsin a cell-free hyaluronate-rich matrix. Devi Biol. 44, 298-305.
SHUMWAY, W. (1940). Stages in the normal development of Rana pipiens. Anat. Rec. 78,139-147-
SOLURSH, M. (1976). Glycosaminoglycan synthesis in the chick gastrula. Devi Biol. 50, 525-530.SUGIYAMA, K. (1972). Occurrence of mucopolysaccharides in the early development of the sea
urchin embryo and its role in gastrulation. Dev., Growth Differ. 14, 63-73.TARIN, D. (1971). Histological features of neural induction in Xenopus laevis. J. Embryol.
exp. Morph. 26, 543-570.TARIN, D. (1973). Histochemical and enzyme digestion studies on neural induction in Xenopus
laevis. Differentiation i, 109-126.TOOLE, B. P. (1973). Hyaluronate and hyaluronidase in morphogenesis and differentiation.
Am. Zool. 13, 1061-1065.(Received 1 November 1976)





























