Embed Size (px)
Citation preview

Photochemisrry and Photobiology Vol. 53, No. 2, pp. 271-274, 1991 Printed in Great Britain
003 1 -8655/9 1 $03.00 + 0.00 Pergamon Press plc
EXCITATION OF MESOPHYLL AND BUNDLE SHEATH CHLOROPLASTS OF Atriplex repanda IN THE ABSENCE
OF LIGHT EDUARDO SILVA'*, GLADYS RODRfGUEZ', ADELAIDE FALJONI* and GIUSEPPE CILENTO*
'Facultad de Quimica, Pontificia Universidad Catdlica de Chile, Casilla 6177, Santiago, Chile and ZDepartamento de Bioquimica, Instituto de Quimica, Universidade de SSo Paulo, C.P. 20780,
Sio Paulo, 01498, Brazil
(Received 16 May 1990; accepted 6 September 1990)
Abstract-Mesophyll and bundle sheath chloroplasts isolated from Afriplex repanda cells promote oxygen consumption by isobutyraldehyde or phenylacetaldehyde. In all cases, a red emission and reduction of tetrazolium blue was observed. Addition of horseradish peroxidase greatly increases the reduction of the dye. In the presence of 3-(3,4-dichlorophenyI)-l,l-dimethylurea, the reduction of the Hill acceptor was fully suppressed. This suppression was abolished when 2,6-dichlorophenolindophenol and ascorbate were added to the systems. These results indicate that, in mesophyll and bundle sheath chloroplasts, chlorophylls can be efficiently excited in the absence of light and an electron flow through the photosystems can be promoted.
peroxidase [ I /H ] p* 0 H2 //
*-C-C +Oz -> C---OH + H C O O H + 3 M , H 0- 0
'H
INTRODUCTION
The great potential of generation and transfer of triplet energy in enzymatic systems for driving pho- toprocesses in the absence of light has recently been stressed in a review of photochemistry and photobi- ology without light (Cilento and Adam, 1988). A remarkable enzyme for generating triplet species is peroxidase and one of the key substrates is phenyl- acetaldehyde (PAA)t. Peroxidase-containing plant extracts were found t o oxidize PAA to benzal- dehyde and formic acid (Kenten, 1953). Since these are the products expected from an intermediate dioxetane, there was the possibility-already confirmed-that benzaldehyde is generated in elec- tronically excited states, mainly in the triplet state. Indeed, cleavage of simple dioxetanes leads to an electronically excited triplet carbonyl compound (Adam, 1977, 1982). Directly and/or indirectly (via induced lipid peroxidation) related to the generation of excited species, when PAA was added to spinach chloroplasts (Nassi and Cilento, 1982) or to the
'To whom correspondence should be addressed. tAbbreviations: 'Ac', triplet acetone; Chl, chlorophyll;
'Chi', excited singlet chlorophyll; DCMU, 3-(3,4- dichloropheny1)-1,l-dimethylurea; DCPIP, 2,6-di- chlorophenolindophenol; EDTA, ethylenediaminetetra- acetic acid; HRP, horseradish peroxidase; IBAL, isobu- tyraldehyde; PAA, phenylacetaldehyde; PS-I and PS- 11, photosystems I and 11, respectively; PUFA, polyun- saturated fatty acid; TB, tetrazolium blue; TJ3H2, reduced tetrazolium blue.
photosynthetic protozoan Euglena gracilis (De Mello et al . , 1988), excitation of chlorophyll was observed, as attested to by red emission and Hill activity. We report here that isobutyraldehyde (1BAL)-a precursor of triplet acetone-and PAA promote the excitation of chlorophylls in dark- adapted mesophyll and bundle sheath chloroplasts of a C4 plant (Atriplex repanda). The present results greatly expand the earlier ones and also provide a wealth of new information.
MATERIALS AND METHODS
Horseradish peroxidase (HRP; type VI), sodium ascorb- ate, 3-(3,4-dichlorophenyI)-l .I-dimethylurea (DCMU), 2, 6-dichlorophenolindophenol (DCPIP), PAA and IBAL were obtained from Sigma Chemical Company (St. Louis, MO). Tetrazolium blue (TB) was from Mann Research Laboratory (New York, NY). PAA was distilled in vacuo under nitrogen and the center cut collected. IBAL was distilled under nitrogen. Ethanolic solutions of IBAL and PAA (1 : 4, vol/vol; EtOH-IBAL, EtOH-PAA) were prepared immediately prior to use.
Mesophyll and bundle sheath chloroplasts from Atriplex repanda were isolated according to Edwards et al. (1979). Macerozyme R-10 and cellulase Onozuka R-10 were pur- chased from Serva (Heidelberg, FRG).
All the following experiments were performed in dark- ness.
The standard reaction mixture was prepared as follows. To 2.9 mL of 0.62 M phosphate buffer, pH 7.4, were added 10 )I.L of 0.1 M EDTA and chloroplasts. The total chlorophyll content was 6.3 F g h L . To determine the Hill activity, 2.23 mM TB was present. The reaction was started by adding 100 pL of 2.2 M IBAL or of 1.8 M PAA in ethanol at 37°C. In experiments with HRP. 10 pL of a
271 PAP 53:2-H
212 EDUARDO SILVA er al.
Table I . Oxygen consumption by isolated mesophyll and bundle sheath chloroplasts in the absence and presence of
isobutyraldehyde (IBAL) or phcnylacetaldehyde (PAA)
Rate of oxygen consumption (pmol OJmg Chl. min)
Additions Mesophyll Bundle sheath
None IBAL PAA
0 0 6.9 6.6 2.7 2.4
solution of the enzyme (20 mg/mL) were added. In all cases, after 15 min incubation the solutions were
centrifuged and the reduction of TB was followed spectro- photometrically in 1 cm path length cells by the increase in absorbance at 580 nm (Ash eral., 1961). This incubation period was significantly longer than that of the emission and was selected this way with the purpose to make certain that the biochemical reactions were concluded. Oxygen consumption was determined with a Yellow Springs Instru- ments Model 5300 oxygen monitor. Chemiluminescence was measured in a Hamamatsu TVC-767 photon counter with and without a Toshiba R-63 filter, which eliminates all the emission at wavelengths shorter than 630 nm. For spectrophotometric studies, a Varian Super Scan 3 rec- ording spectrophotometer was employed. Arnon’s method (1949) was used to determine chlorophyll concentrations.
RESULTS AND DISCUSSION
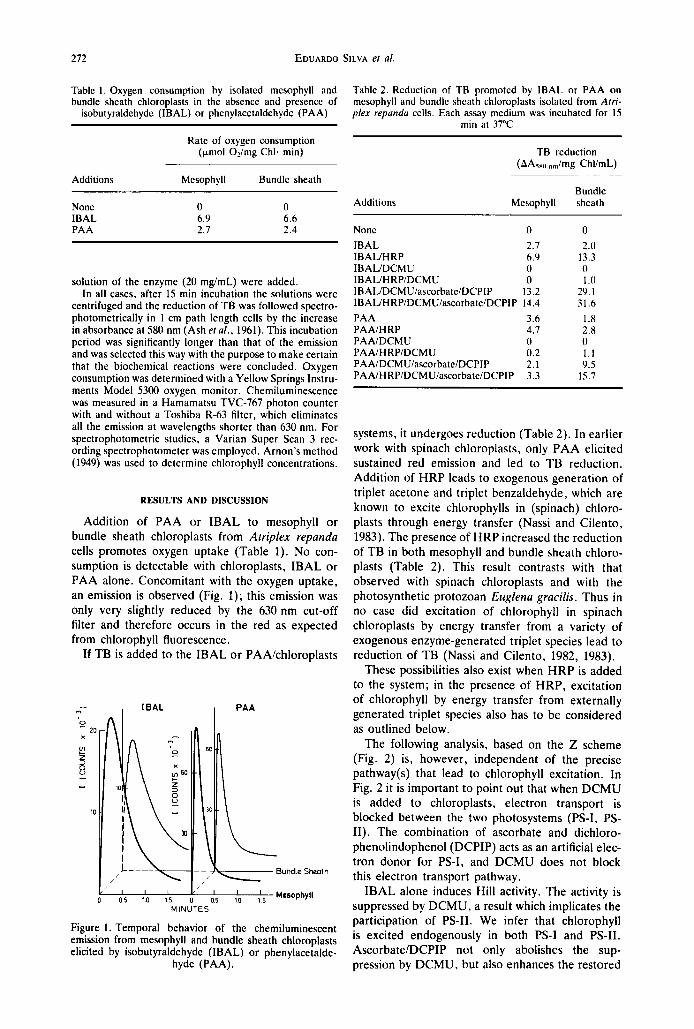
Addition of PAA or IBAL to mesophyll or bundle sheath chloroplasts from Atriplex repanda cells promotes oxygen uptake (Table 1). No con- sumption is detectable with chloroplasts, IBAL or PAA alone. Concomitant with the oxygen uptake, an emission is observed (Fig. 1); this emission was only very slightly reduced by the 630 nm cut-off filter and therefore occurs in the red as expected from chlorophyll fluorescence.
If TB is added to the IBAL or PAAlchloroplasts
Fl , I B A L I P A A I
x
51
r - I
Bundle Sheath / - I -
MINUTES
Figure 1. Temporal behavior of the chemiluminescent emission from mesophyll and bundle sheath chloroplasts elicited by isobutyraldehyde (IBAL) or phenylacetalde-
hyde (PAA).
Tdbk 2. Reduction of TB promoted by IBAL or PAA on mesophyll and bundle sheath chloroplasts isolatcd from Arri- plex repundu cells. Each assay medium was incubated for 15
min at 37°C
Additions
TB reduction (PA,,,, J m g ChlImL)
_ _ -
Bundlc Mesophyll sheath
None 0 0 IBAL 2.7 2.0 IBALIHRP 6.Y 13.3 IBALIDCMU 0 0 IBALIHRPIDCMU 0 1 .o IBALIDCMUIascorbatclDCPIP 13.2 29.1 IBALIHRPIDCMUlascorbateiDCPIP 14.4 31.6 PAA 3.6 1.8 PAAIHRP 4.7 2.R PAAlDCMU 0 0 PAA/HRP/DCMU 0.2 1.1 PAAIDCMUIascorbatelDCPIP 2.1 9.5 PA A/H RPIDCMUIascorbatelDCPI P 3.3 15.7
systems, it undergoes reduction (Table 2). In earlier work with spinach chloroplasts, only PAA elicited sustained red emission and led to TB reduction. Addition of HRP leads to exogenous generation of triplet acetone and triplet benzaldehyde, which are known to excite chlorophylls in (spinach) chloro- plasts through energy transfer (Nassi and Cilento, 1983). The presence of HRP increased the reduction of TB in both mesophyll and bundle sheath chloro- plasts (Table 2). This result contrasts with that observed with spinach chloroplasts and with the photosynthetic protozoan Euglena gracilis. Thus in no case did excitation of chlorophyll in spinach chloroplasts by energy transfer from a variety of exogenous enzyme-generated triplet species lead to reduction of TB (Nassi and Cilento, 1982, 1983).
These possibilities also exist when HRP is added to the system; in the presence of HRP, excitation of chlorophyll by energy transfer from externally generated triplet species also has to be considered as outlined below.
The following analysis, based on the Z scheme (Fig. 2) is, however, independent of the precise pathway(s) that lead to chlorophyll excitation. In Fig. 2 it is important to point out that when DCMU is added to chloroplasts, electron transport is blocked between the two photosystems (PS-I, PS- 11). The combination of ascorbate and dichloro- phenolindophenol (DCPIP) acts as an artificial elec- tron donor for PS-I, and DCMU does not block this electron transport pathway.
IBAL alone induces Hill activity. The activity is suppressed by DCMU, a result which implicates the participation of PS-11. We infer that chlorophyll is excited endogenously in both PS-I and PS-11. Ascorbate/DCPIP not only abolishes the sup- pression by DCMU, but also enhances the restored

Dark excitation of chloroplasts 273
Figure 2. Pathway of enzymically induced electron flow from H,O to tetrazolium blue (TB) through PS-I and PS- 11. 0 and EP correspond to the primary acceptor of PS- I1 and to endogenous peroxidase or peroxidase like activity
respectively.
activity. Therefore, in our system ascorbate/DCPIP is a better supplier of electron to PS-I than the endogenous excited PS-II/H20 system.
With PAA, the results are similar to those observed with IBAL, except that, in the case of mesophyll chloroplasts, ascorbate/DCPIP does not fully restore the activity. This fact suggests that PS- I is not efficiently excited by PAA. It also suggests that PS-I1 is, in this case, a better source of electrons than ascorbate/DCPIP which in turn might imply that PAA efficiently excites PS-I1 in mesophyll chlo- roplasts.
Summarizing, the fact that IBAL and PAA induce Hill activity, concomitant with oxygen uptake and emission of light in both mesophyll and bundle sheath chloroplasts, indicates the presence of an endogenous peroxidase like activity in the Atriplex repanda cells. In mesophyll chloroplasts, IBAL is better than PAA at exciting PS-I, whereas PAA is better at exciting PS-11. In bundle sheath chloroplasts, IBAL is better than PAA at exciting PS-I (compare in Table 2 reduction of TB by IBAL/ DCMU /axorbate / DCPIP and by PAA/ DCMU/ascorbate/DCPIP) .
On the basis of O2 consumed, PAA is more efficient than IBAL in generating TB reducing power; this applies to both mesophyll and bundle sheath chloroplasts.
Mesophyll und bundle sheath chloroplasts; com- parative behavior
Let us first consider the results with IBAL as substrate. Considering the system that contains DCMU/ascorbate/DCPIP and that therefore “meas- ures” excitation in PS-I, we infer that PS-I is more efficiently excited in bundle sheath chloroplasts than in mesophyll chloroplasts. On the other hand, the PS-I1 system may be excited somewhat better in mesophyll than in bundle sheath chloroplasts; thus, in spite of a greater excitation of PS-I in bundle sheath, there is a somewhat larger Hill activity of IBAL with mesophyll chloroplasts.
A greater excitation of PS-I in bundle sheath relative to mesophyll chloroplasts is also seen with PAA as substrate. Excitation of PS-I1 is apparently favored in mesophyll chloroplasts because a greater activity is seen relative to bundle sheath chloroplasts despite a less efficient excitation of PS-I.
Summarizing, with either IBAL or PAA as substrate, PS-I is excited better in bundle sheath chloroplasts, whereas PS-I1 is excited better in mesophyll chloroplasts.
Presence of H R P
It is certainly appropriate to recall that both exter- nally generated triplet acetone and triplet benzal- dehyde transfer energy to thylakoid membrane frac- tions (from spinach chloroplasts) enriched in either PS-I or PS-I1 (Nassi and Cilento, 1985).
In the case of IBAL as substrate, with either mesophyll or bundle sheath chloroplasts, added HRP only very modestly enhances the excitation of PS-I. Indeed the rate of TB reduction is only very modestly enhanced in going from IBAL/DCMU/ ascorbate/DCPIP to IBAL/HRP/DCMU/ascor- bate/DCPIP. Yet HRP markedly enhances Hill activity promoted by IBAL. Therefore exogenously generated triplet acetone must efficiently excite PS- I1 through energy transfer. The enhanced Hill activity requires that the excitation of PS-I is not rate limiting. Accordingly the failure of externally generated triplet acetone to enhance significantly the reduction of TB by IBAL/DCMU/ascorba- te/DCPIP may imply that PS-I is efficiently excited by the endogenous route. The high rates of TB reduction by IBAL in the presence of DCMU/ascor- bate/DCPIP support this view.
When PAA is the substrate, added HRP signifi- cantly enhances the excitation of PS-I, the rate of TB reduction being increased 57% with mesophyll and 66% with bundle sheath chloroplasts. However HRP enhances the Hill activity promoted by PAA alone by 30 (mesophyll) and 55% (bundle sheath) only. In mesophyll chloroplasts either PS-I1 is some- what less efficiently excited than PS-I by the exter- nal donor or PS-I1 is already efficiently excited by the endogenous donor.
CONCLUSION
Following addition of IBAL or PAA to chloro- plasts, there is an electron flow mediated by the photosystems excited by energy transfer from elec- tronically excited species generated in the oxidation of IBAL and PAA catalyzed by endogenous peroxi- dase or peroxidase-like activity. This behavior is similar to that observed in the presence of light. The dramatic suppression of the Hill activity by DCMU indicates that the reducing power comes from the splitting of water. The possibility that dark excitation of chloroplasts may play a role has al-

274 EDUARDO SILVA er al.
ready been mentioned by Salim-Hanna et al. (1987).
Acknowledgements-The authors are deeply indebted to Professor F. H. Quina (SBo Paulo University) for a critical reading of the manuscript. They thank DIUC P. Universi- dad Catolica de Chile, FINEP (Rio de Janeiro), FAPESP (SBo Paulo). CODAC University of Sio Paulo, CNPq (Brasilia), the Volkswagen Foundation (Hanover) and GTZ (Eschborn) for financial support.
REFERENCES
Adam, W. (1977) The chemistry of 1,2-dioxetanes. Adv. Hererocycl. Chem. 21, 437-481.
Adam, W. (1982) Determination of chemiexcitation yields in the thermal generation of electronic excitation from 1.2-dioxetanes. In Chemical and Biological Generation of Excited Srares (Edited by W . Adam and G. Cilento). pp. 115-152. Academic Press, New York.
Arnon, D. I. (1949) Copper enzymes in isolated chloro- plasts. Polyphenoloxidase in Bera vulgaris. Planr Phy- siol. 24, 1-15.
Ash, 0. K., W. S . Zaugg and L. P. Vernon (1961) Photo- reduction of methyl red and tetrazolium blue by spinach chloroplasts and chromatophores of Rhodospirillum rubrum. Acra Chem. Scand. 15, 1629-1638.
Cilento, G. and W. Adam (1988) Photochemistry and
photobiology without light. Phofochem. Photobiol. 48.
De Mello. M. P.. A. L. T. 0. Nascimento, C. Bohne and G. Cilento (1988) Excitation of chloroplasts in Euglena gracilis in the absence of light. Phofochem. Phorobiol. 47. 457-461.
Edwards, G. W., R. McC. Lilley, S . Craig and M. D. Hatch (1979) Isolation of intact and functional chloro- plasts from mesophyll and bundle sheath protoplasts of the C, plant Panicum miliaceum. Planr Physiol. 63.
Kenten, R. H. (1953) The oxidation of phenylacetalde- hyde by plant saps. Biochem. J . 55, 350-360.
Nassi, L. and G. Cilento (1982) Excitation of chloroplasts induced by phenylacetaldehyde. Photochem. Phorobiol.
Nassi. L. and G. Cilento (1983) Red emission from chloro- plasts elicited by enzyme-generated triplet acetone and triplet indole-3-aldehyde. Phorochem. Phorobiol. 37. 233-237.
Nassi, L. and G. Cilento (1985) Energy transfer from enzyme-generated triplet carbonyls to thylakoid mem- brane fractions enriched in Photosystems I and 11. Pho- rochem. Phorobiol. 41, 195-201,
Salim-Hanna, M., A. Campa and G. Cilento (1987) The a-oxidase system of young pea leaves (Pisum sativum) as generator of electronically excited states. Excitation in the dark under natural conditions. Phorochem. Phoro- biol. 45, 849-854.
36 1-368.
821-827.
36, 121-123.












![Estimating Mesophyll Conductance from Measurements of ... · Estimating Mesophyll Conductance from Measurements of C18OO Photosynthetic Discrimination and Carbonic Anhydrase Activity1[OPEN]](https://img.pdfslide.us/doc/110x75/5e218e60b49cd34ffe11f49e/estimating-mesophyll-conductance-from-measurements-of-estimating-mesophyll-conductance.jpg)
















