Embed Size (px)
Citation preview

http://wwwsoc.nii.ac.jp/jsme2/
Microbes Environ. Vol. 22, No. 1, 59–70, 2007
Enhancement of Nitrogen-fixing Activity of Enterobacteriaceae Strains
Isolated from Sago Palm (Metroxylon sagu) by Microbial Interaction with
Non-nitrogen Fixers
ARCHANA SHRESTHA1*, KOKI TOYOTA1, MASANORI OKAZAKI
1, YUKO SUGA2, MARCELO A. QUEVEDO3,
ALAN B. LORETO3 and ALGERICO A. MARISCAL
3
1 Graduate school of Bio-Applications and System Engineerings, Tokyo University of Agriculture and Technology,
2–24–16, Nakamati, Koganei City, Tokyo 184–8588, Japan2 National Agricultural Research Center for Western Region, Kyoto, Japan3 Philippine Root Crop Research and Training Center (PhilRootcrops), Leyte State University, Baybay, Leyte,
Philippines
(Received September 25, 2006—Accepted January 9, 2007)
This study was done to determine microbial interactions between nitrogen-fixing bacteria (NFB) and different
indigenous bacteria, both of which were isolated from the same samples of sago palm. Co-culture of NFB and
indigenous bacteria, isolated using nutrient agar medium, into a nitrogen-free Rennie medium showed signifi-
cantly higher nitrogen-fixing activity (NFA) than single inoculations of NFB did in almost all combinations. A
reduced oxygen status also enhanced the NFA of NFB. All NFB preferred simple sugars as their substrates for
NFA and showed extremely low levels of NFA in starch, hemicellulose, and pectin-containing media. NFA was
markedly stimulated in the consortium of starch-degrading Bacillus sp. strain B1 and NFB, ranging from 0–1.5
to 150–270 nmol C2H4 culture−1 h−1. The consortium of hemicellulose-degrading Agrobacterium sp. strain HMC1
or Flexibacter sp. strain HMC2 and NFB also showed enhanced NFA, ranging from 0–0.1 to 16–38 nmol. In
contrast, no enhanced NFA was detected in the consortium of pectin-degrading Burkholderia sp. strain BT1 or
Paenibacillus sp. strain P1 and NFB. These results may indicate that beneficial microbial interactions occur in
sago palm to enhance nitrogen-fixing activity through collaborative utilizations of starch, hemicellulose and their
degradation products.
Key words: starch-degradation, pectin-degradation, hemicellulose-degradation, PCR-DGGE, nifH
In agriculture, crop productivity is mainly governed by
the use of inorganic N-fertilizers, which are expensive and
damaging to the environment32). Finding alternative ways of
supplying nitrogen to grasses and cereals has been a major
research challenge in tropical agriculture1,3). From ecologi-
cal and economical perspectives, plant-associated biological
nitrogen fixation is highly desirable in non-leguminous
plants as well as leguminous plants29).
Sago palm (Metroxylon sagu) is one of the oldest tropical
plants exploited by humanity33) and its starch has been used
in Southeast Asia, including Papua New Guinea, Indonesia,
and Malaysia, and in Pacific Oceania16). These plants have
great potential as a source of food for people in the regions
in which they grow5). Sago is still an important crop with a
vital role as a subsistence food in some areas of Papua New
Guinea and Irian Jaya33). In East Malaysia and Indonesia,
industrial utilization has helped in the development of this
crop33). Sago palm grows naturally under semi-cultivated
conditions in vast tracts of lowland15). It has several charac-
teristics that make it a quite remarkable plant: it is well
adapted to swampy, acidic peat soils, where few other crops
survive, and it tolerates flooded or drought conditions43).
In our previous study40), nitrogen-fixing bacteria (NFB)
* Corresponding author. E-mail address: [email protected]; Tel. &
Fax: +81–42–388–7915.

SHRESTHA et al.60
were isolated from the different parts of sago palm and were
identified as Klebsiella pneumoniae, Klebsiella oxytoca,
Pantoea agglomerans, Enterobacter cloacae, and Bacillus
megaterium. Moreover, it was suggested that NFB utilized
exclusively simple carbon sources like glucose and sucrose,
and not complex carbon sources like starch, pectin, and
hemicellulose. Haahtela et al.10) also found that simple car-
bon sources are preferable for nitrogen-fixing activity by
NFB. In addition, Halsall et al.12) reported that straw (hemi-
cellulose is the major component) was not utilized as an
energy source for nitrogen fixation. Simple carbon sources
like glucose and sucrose, however, are not readily available
in natural microenvironments10) while the major carbohy-
drates of plants like cellulose, hemicellulose and pectin24)
are easily and readily available in natural microenviron-
ments for different microbial activities2). Besides, this sago
palm bears a typical characteristic, the storing of starch in
its trunk33).
Typically, numerous bacterial populations with different
characteristics coexist in a natural environment2). Hence,
microbial interactions providing simple carbon sources
from naturally existing polymers might be one way to
enhance nitrogen-fixing activity of NFB. Therefore, the
aims of the present study were to examine the nitrogen-
fixing activity of NFB in the presence of different types of
indigenous bacteria: (1) indigenous dominant bacteria, (2)
starch-degrading bacteria, (3) pectin-degrading bacteria,
and (4) hemicellulose-degrading bacteria.
The application of molecular biological methods has
greatly facilitated the study of natural bacterial communities
and the identification of functionally significant organisms
within them27,31). In this study, we confirmed the existence
of NFB isolated from root and starch of sago palm in natural
samples by comparing banding patterns from denaturing
gradient gel electrophoresis (DGGE) targeted at nifH
between the isolates and community DNA extracted from
root and starch samples.
Materials and Methods
Isolation and identification of bacteria
The sago palm parts (root, starch, midrib and trunk) were
collected during March 2005 from Hilusig (N10°38'80'',
E124°56'72''), Leyte, The Philippines. Roots, starch, trunk
and midrib were washed with tap water to remove all soil
particles and rinsed with sterile distilled water. The washed
roots and midribs were cut into 1 to 2 cm segments with
scissors, and 3 g (fresh basis) of sample was transferred into
a vial. The vials were capped with rubber stoppers, and
acetylene gas was injected into the headspace of the vial at a
final concentration of 10%. Experiments were done as one
replicate, in duplicate, or in triplicate, depending on the
availability of samples. Acetylene-reducing activity (ARA)
was quantified, using nitrogen-free Rennie medium36), 4 to 7
days after the injection of acetylene by injecting 0.5 ml of
the headspace into a gas chromatograph (GC-14B; Shi-
madzu, Kyoto, Japan) equipped with a flame ionization
detector and a column packed with Shincarbon S (3 mm×2
m, 60/80 mesh, Shimadzu GLC Center, Tokyo, Japan).
Nutrient agar medium was used for the isolation of indig-
enous dominant bacteria from sago root, midrib, and trunk.
For the isolation of starch-, pectin- and hemicellulose-
degrading bacteria, an enrichment culture was done using
an inorganic salt medium (1.5 g NaNH4HPO4. 4H2O, 1.0 g
KH2PO4, 0.2 g MgSO4, 1000 ml distilled water) supple-
mented with 2.5 g of starch, pectin, or hemicellulose, and
incubated at 27°C for 48 h without shaking, and then the
corresponding polymer-degrading bacteria were isolated
using the same media supplemented with agar (15 g L−1)
and the same conditions. For the enrichment culture of
hemicellulose- and pectin-degrading bacteria, 1 ml of seri-
ally diluted sample of root (1 g macerated root) was used
along with the same technique for isolating starch-degrad-
ing bacteria from starch samples (1 g macerated starch).
To estimate phylogenic positions of NFB and polymer
degraders, DNA was extracted from the purified bacterial
colonies by using the conventional method30). PCR primers
(27f; 5'-AGAGTTTGATCCTGGCTCAG-3' and (1378r; 5'-
TACAAGGCCCGGGAACG-3') universal to the domain
Bacteria18) were used to amplify the segment of the 16S
rRNA gene corresponding to nucleotide positions 27 to
1378 in terms of Escherichia coli numbering25). Then, direct
sequencing of the PCR products was done by Hitachi Sci-
ence Systems, Ltd. (Tokyo, Japan). Approximately 750-bp
fragments of the 16S rRNA gene were sequenced between
nucleotides 27 to 1378 from the reverse side. The sequences
obtained were examined using the DDBJ homology search
system BLAST (http://www.ddbj.nig.ac.jp/search/blast-
j.html). For phylogenetic analysis, the nucleotide sequences
were aligned using CLUSTAL W (http://www.ddbj.nig.
ac.jp/search/clustalw-j.html).
PCR-DGGE analysis of nifH genes from the pure cul-
tures and the root and starch of sago palm
The roots collected (ca. 2–3 g) were cut into pieces a few
millimeter long with scissors and in the case of starch, 2–3 g
was collected. Both the root and starch samples used for
DNA extraction was the same samples used for assaying

Microbial Consortium Enhance N-fixation 61
ARA activity and the isolation of NFB. DNA was extracted
from a portion (0.1 g) of each sample using a bead beater
(Bead Smash-12, WAKENYAKU, Kyoto, Japan) with 1 ml
of extraction buffer (0.1 M Tris-HCl (pH 8), 40 mM EDTA,
0.2 M NaCl, 20 g L−1 SDS). The supernatant was collected
and to it, 400 µl of 7.5 M NH4OAc was added. This mixture
was kept on ice for 5 minutes and centrifuged for 3 min at
13,200×g. To the supernatant, a 70% of volume of ethanol
was added and the mixture was kept at −80°C for 1 h, cen-
trifuged at 13,200×g for 10 min, washed with 70% ethanol,
and finally centrifuged and suspended in TE buffer.
The first PCR was performed with the forward primer
FGPH19 (5'-TACGGCAARGGTGGNATHG-3') and the re-
verse primer PoIR (5'-ATSGCCATCATYTCRCCGGA-3')6).
The amplification product was 429 bp. Then, the second
PCR was performed with the forward primer PoIF-GC (5'-
TGCGAYCCSAARGCBGACTC-3' with a GC clamp and
the reverse primer AQER (5'-GACGATGTAGATYTCCTG-
3')6). The amplification product was 360 bp including a GC
clamp sequence. The cycling conditions used were: 30 cy-
cles of denaturation at 94°C for 1 min, annealing for 1 min
at 55°C for the first and at 48°C for the second PCR, and
primer extension at 72°C for 2 min, with a final extension at
72°C for 5 min. The 2nd PCR products were then subjected
to DGGE.
The DGGE analysis was done by using a Bio-Rad
DCode, Universal Mutation Detection System (Bio-Rad
Laboratories, Hercules, Ca, USA). About 15–20 µl of PCR
product was loaded onto a 6% (w/v) polyacrylamide gel
with 40 to 70% gradients (urea and formamide). The elec-
trophoresis was run for 14 to 16 h at 60°C and 100 V. The
gel was then stained with a SYBR green I nucleic acid gel
stain (1:10000 dilution; Cambrex Bio Science, Rockland,
ME, USA) for 30 min in Tris-acetate EDTA buffer. DNA
from the isolated diazotrophs in this study, Klebsiella
pneumoniae, K. oxytoca, Pantoea agglomerans, and Enter-
obacter cloacae, was used as positive controls.
NFA with different carbon sources
Various carbohydrates like sucrose, glucose, mannitol,
starch, lactate, pectin, hemicellulose galactose, mannose,
cellobiose and galactouronic acid were used to estimate
suitable carbon sources for NFA. All the reagents were spe-
cial grade and purchased from Wako Chemicals, except
galactouronic acid which was from Sigma Co. Ltd. and cel-
lobiose which was from MP Biomedicals, LLC. In this
experiment, a 30-ml sterilized rubber capped vial, into
which 3 ml of semi-solid nitrogen-free Rennie medium
(0.3% agar) containing different individual carbohydrates at
a rate of 5 g L−1, was dispensed and used. Individual pure
bacterial colonies were first inoculated into 3 ml of sterile
distilled water to remove extra nutrients accompanied by
cells, and then a loopful of the bacterial suspension was
inoculated into each medium containing a single carbohy-
drate source. The vials were capped with rubber stoppers,
and acetylene gas was injected into the headspace of the vial
at a final concentration of 10% and incubated at 28°C for 48
h without shaking. The amount of ethylene in the headspace
was quantified by gas chromatography.
NFA in consortia
NFB were inoculated into semi-solid N-free Rennie
medium along with different types of bacteria, all of which
were isolated from the sago palm samples used for ARA
and isolation. Four experimental sets were designed in
which, 1) indigenous dominant bacteria, 2) starch-degrading
bacteria, 3) hemicellulose-degrading bacteria, and 4) pectin-
degrading bacteria were inoculated. In experiment 1, semi-
solid Rennie medium was used, while in experiments 2, 3,
and 4, starch, hemicellulose or pectin was used as a sole
carbon source, instead of mannitol, lactate, or glucose. In
experiment 1, a total of 50 indigenous bacteria were isolated
(25 from sago root, 10 from sago midrib, and 15 from sago
trunk) on simple nutrient agar medium and then separated
into different groups randomly, named as A, B, C, D and E
for root isolates; F and G for midrib isolates, and H, I and J
for trunk isolates and each group contained 5 strains. Exper-
iments were done in triplicate and the amount of ethylene in
the headspace was quantified after 3 to 4 days by gas chro-
matography.
In experiment 2, N-free starch broth medium was used to
estimate the population density of i) NFB, ii) Bacillus sp.
strain B1, and iii) Bacillus sp. strain B1+NFB and incubated
at 27°C for 48 h. From each broth culture, a set of serial
dilutions was prepared and then colony counts were made
on simple nutrient agar media by using a spread plate tech-
nique after incubation at 27°C for 48 h. In the mixed cul-
ture, the colonies of Bacillus sp. strain B1 and NFB were
separated on the basis of colony morphology.
NFA under reduced oxygen conditions and in the cell-
free filtrates of indigenous bacteria
A 30 ml sterilized rubber capped vial was used in this
experiment into which 3 ml of N-free Rennie medium was
dispensed. Then, individual pure bacterial colonies were
inoculated into 3 ml of sterile distilled water and a loopful
of bacterial suspension was inoculated into the medium.
The vials were capped with rubber stoppers and then about
SHRESTHA et al.62
18 ml (2 times of 9 ml) of air was withdrawn using a syringe
and replaced with the same amount of N2 gas and acetylene
gas was injected into the headspace of the vial at a final
concentration of 10% and incubated at 28°C for 48 h with-
out shaking and ARA was measured.
In order to obtain fresh cultures, all the indigenous domi-
nant bacteria isolated from, root, midrib, and trunk, were
sub-cultured on fresh nutrient agar media and incubated at
27°C for 24 h. Then the bacterial suspensions of each group
were prepared by inoculating the colonies in 5 ml of sterile
distilled water. Then the prepared suspensions of each
group of indigenous dominant bacteria were cultured in
semisolid nutrient broth at 27°C for 48 h and cell-free fil-
trate fluid was obtained by filtering the culture using a 0.2
µm filter. Thus collected filtrates were used to measure the
ARA of NFB.
Statistical analysis
Data were analyzed using a standard analysis of variance
(ANOVA) (Excel Toukei, 2002 for Windows).
Results
Isolation and identification of bacteria
16S rRNA gene sequencing showed that all of the NFB
with high levels of ARA belong to the family Enterobacte-
riaceae with Klebsiella oxytoca, K. pneumoniae, Entero-
bacter cloacae, and Pantoea agglomerans as their closest
relatives (Table 1 and Fig. 1). The nitrogenase activity of
the strains varied from 2.4 to 19 nmol C2H4 culture−1 h−1, the
lowest value being measured in Klebsiella sp. strain R3 and
the maximum, in Klebsiella sp. strain R4 (Table 2). Both of
these species were isolated from sago root (Table 1). All of
the partial sequences of the 16S rRNA gene determined in
NFB have been submitted to the DDBJ database under the
accession number AB270603 to AB270612.
Starch-degrading bacteria, Bacillus sp. strain B1 (acces-
sion number, AB270626), was isolated from sago starch;
pectin-degrading bacteria, Burkholderia sp. strain BT1
(AB271166) and Paenibacillus sp. strain P1 (AB271167),
and hemicellulose-degrading bacteria, Agrobacterium sp.
strain HMC1 (AB270627) and Flexibacter sp. strain HMC2
(AB270628), were isolated from sago root (Table 1).
Table 1. Bacterial strains isolated from sago palm in this study
Bacterial strains
isolated
Isolated
from
Accession number for
16S rRNA gene sequences
determined in this study
16S rRNA gene sequences comparison
Closest relative (accession number) % similarity
Nitrogen fixers
R1 Root AB270612 Pantoea agglomerans (AM184307) 100
R2 Root AB270610 Enterobacter cloacae (AY914097) 98.4
R3 Root AB270608 Klebsiella oxytoca (AY873801) 99.8
R4 Root AB270605 K. pnuemoniae (DQ379506) 100
S1 Starch AB270611 E. cloacae (DQ379506) 98.4
S2 Starch AB270603 K. pnuemoniae (DQ379506) 99.8
S3 Starch AB270607 P. agglomerans (AM184307) 99.8
M Midrib AB270606 P. agglomerans (AM184307) 100
T1 Trunk AB270604 K. pnuemoniae (DQ379506) 99.5
T2 Trunk AB270609 K. oxytoca (AY873801) 99.8
Starch-degraders
B1 Starch AB270626 Bacillus megaterium (DQ872156) 100
Pectin-degraders
BT1 Root AB271166 Burkholderia tropicalis (AY321307) 100
P1 Root AB271167 Paenibacillus sp. (AY728023) 99.4
Hemicellulose-degraders
HMC1 Root AB270627 Agrobacterium tumifaciens (DQ14505) 99.5
HMC2 Root AB270628 Flexibacter sancti (AB078068) 98.8

Microbial Consortium Enhance N-fixation 63
Fig. 1. Phylogenetic tree based on the 16S rRNA gene sequence analysis of sago root, starch, midrib and trunk bacteria. Scale bar indicates 10%
sequence divergence. Strain names are preceded by their GenBank accession numbers and the date of sample collection is written in parenthe-
ses. DNA sequences of some control strains were obtained from the DDBJ database.
Table 2. Nitrogen-fixing activities (acetylene-reducing activity) of parts of sago palm plants collected from Hilsug, Leyte, Philippines in March
2005 and strains isolated from the plant
Plant part of isolationAcetylene-reducing activity
(nmol C2H4 g−1 day−1)
Strains isolated from
the corresponding plant part
Acetylene-reducing activity
(nmol C2H4 culture−1 h−1)
Root 26±14a) Pantoea sp. strain R1 15±9.2
Enterobacter sp. strain R2 5.3±8.1
Klebsiella sp. strain R3 2.5±0.4
Klebsiella sp. strain R4 19±0.1
Starch 210±21 Enterobacter sp. strain S1 11±1.4
Klebsiella sp. strain S2 14±3.1
Pantoea sp. strain S3 15±0.8
Midrib 66±22 Pantoea sp. strain M 12±3.2
Trunk 590±0.8 Klebsiella sp. strain T1 2.4±0.5
Klebsiella sp. strain T2 4.4±0.2
a) Mean values in triplicates.

SHRESTHA et al.64
PCR-DGGE analysis of nifH genes from the pure cul-
tures and the roots and starch
DGGE revealed the diversity of the nitrogen-fixing
microbial populations in sago root and sago starch (Fig. 2).
There were at least 5 prominent bands in the sago root
(Fig. 2, bands a, b, c, d and e) and in the starch sample
(Fig. 2. band g, h, i, j and k). The bands of 4 isolated nitro-
gen fixers were clearly separated (A, B, C and D). Although
the band corresponding to Klebsiella sp. strain R3 was not
observed in the root or starch sample, the bands d, e, and l
were identical to the positions of the remaining 3 reference
strains and thus were excised and sequenced (Fig. 2). The
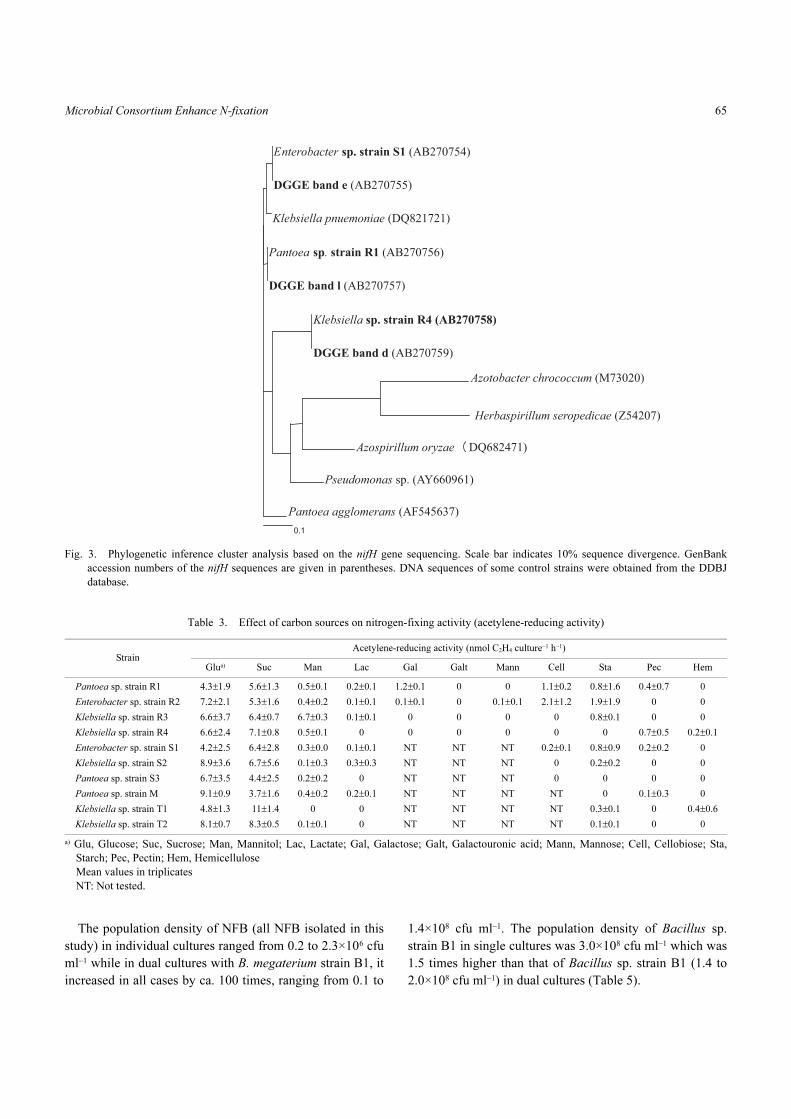
sequences of bands “d”, “e” and “l” were identical to nifH
gene sequences (100% homology) of the reference strains
of bands A, B, and D, respectively (Fig. 3).
NFA under different carbon sources
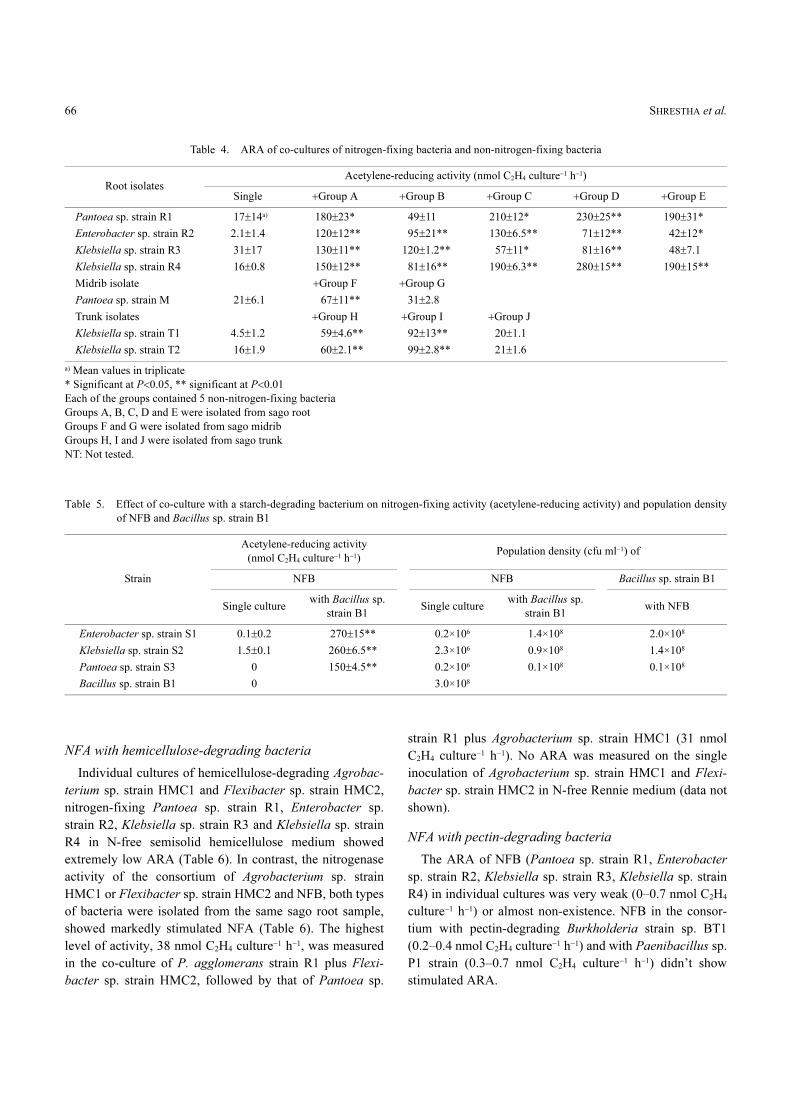
Among all the carbon sources, almost all the NFB pre-
ferred glucose for high NFA and some preferred sucrose
(Table 3). The highest nitrogenase activity was found to be
9.1±0.9 nmol C2H4 culture−1 h−1 (Pantoea sp. strain M) with
glucose and 11±1.4 C2H4 culture−1 h−1 (Klebsiella sp. strain
T1) with sucrose. The use of other carbon sources like man-
nitol, lactate, cellobiose, galactose, mannose, and galactou-
ronic acid resulted in little or no nitrogenase activity and the
same results were obtained with the polymers like starch,
pectin and hemicellulose.
NFA with indigenous dominant bacteria
The consortium of NFB plus indigenous bacteria showed
increased NFA in all the cases: from 2.1–31 (single culture)
to 20–280 (dual culture) nmol C2H4 culture−1 h−1 (Table 4).
However, in Pantoea sp. strain M+group G and Klebsiella
sp. strain T2+group J, the ARA activity was not so
enhanced, as shown by the single culture of NFB. No ARA
activity was measured in any of the groups of indigenous
bacteria (data not shown).
NFA with starch-degrading bacteria
The ARA of individual cultures of Enterobacter sp. strain
S1, Klebsiella sp. strain S2 and Pantoea sp. strain S3 in
semi-solid N-free starch medium was 0.1, 1.5 and 0 nmol
C2H4 culture−1 h−1, respectively. Starch-degrading Bacillus
sp. strain B1 did not show any ARA. The consortium of
Bacillus sp. strain B1 and Enterobacter sp. strain S1, Kleb-
siella sp. strain S2, or Pantoea sp. strain S3 showed signifi-
cantly (P<0.01) enhanced NFA. Bacillus sp. strain B1 plus
Enterobacter sp. strain S1 showed the highest level of nitro-
genase activity, 270 nmol C2H4 culture−1 h−1, while Kleb-
siella sp. strain S2 and Pantoea sp. strain S3 showed 260
and 150 nmol C2H4 culture−1 h−1 respectively (Table 5).
Fig. 2. nifH DGGE banding patterns from sago palm root and starch sample. Lane 1 and 2 are sago root replications; Lane 3 and 4 are sago
starch replications. Marker “M” is positive control (A, Klebsiella sp. strain S2; B, Enterobacer sp. strain R2; C, Klebsiella sp. strain R3; D,
Pantoae sp. strain R1).
Microbial Consortium Enhance N-fixation 65
The population density of NFB (all NFB isolated in this
study) in individual cultures ranged from 0.2 to 2.3×106 cfu
ml−1 while in dual cultures with B. megaterium strain B1, it
increased in all cases by ca. 100 times, ranging from 0.1 to
1.4×108 cfu ml−1. The population density of Bacillus sp.
strain B1 in single cultures was 3.0×108 cfu ml−1 which was
1.5 times higher than that of Bacillus sp. strain B1 (1.4 to
2.0×108 cfu ml−1) in dual cultures (Table 5).
Fig. 3. Phylogenetic inference cluster analysis based on the nifH gene sequencing. Scale bar indicates 10% sequence divergence. GenBank
accession numbers of the nifH sequences are given in parentheses. DNA sequences of some control strains were obtained from the DDBJ
database.
Table 3. Effect of carbon sources on nitrogen-fixing activity (acetylene-reducing activity)
StrainAcetylene-reducing activity (nmol C2H4 culture−1 h−1)
Glua) Suc Man Lac Gal Galt Mann Cell Sta Pec Hem
Pantoea sp. strain R1 4.3±1.9 5.6±1.3 0.5±0.1 0.2±0.1 1.2±0.1 0 0 1.1±0.2 0.8±1.6 0.4±0.7 0
Enterobacter sp. strain R2 7.2±2.1 5.3±1.6 0.4±0.2 0.1±0.1 0.1±0.1 0 0.1±0.1 2.1±1.2 1.9±1.9 0 0
Klebsiella sp. strain R3 6.6±3.7 6.4±0.7 6.7±0.3 0.1±0.1 0 0 0 0 0.8±0.1 0 0
Klebsiella sp. strain R4 6.6±2.4 7.1±0.8 0.5±0.1 0 0 0 0 0 0 0.7±0.5 0.2±0.1
Enterobacter sp. strain S1 4.2±2.5 6.4±2.8 0.3±0.0 0.1±0.1 NT NT NT 0.2±0.1 0.8±0.9 0.2±0.2 0
Klebsiella sp. strain S2 8.9±3.6 6.7±5.6 0.1±0.3 0.3±0.3 NT NT NT 0 0.2±0.2 0 0
Pantoea sp. strain S3 6.7±3.5 4.4±2.5 0.2±0.2 0 NT NT NT 0 0 0 0
Pantoea sp. strain M 9.1±0.9 3.7±1.6 0.4±0.2 0.2±0.1 NT NT NT NT 0 0.1±0.3 0
Klebsiella sp. strain T1 4.8±1.3 11±1.4 0 0 NT NT NT NT 0.3±0.1 0 0.4±0.6
Klebsiella sp. strain T2 8.1±0.7 8.3±0.5 0.1±0.1 0 NT NT NT NT 0.1±0.1 0 0
a) Glu, Glucose; Suc, Sucrose; Man, Mannitol; Lac, Lactate; Gal, Galactose; Galt, Galactouronic acid; Mann, Mannose; Cell, Cellobiose; Sta,
Starch; Pec, Pectin; Hem, Hemicellulose
Mean values in triplicates
NT: Not tested.
SHRESTHA et al.66
NFA with hemicellulose-degrading bacteria
Individual cultures of hemicellulose-degrading Agrobac-
terium sp. strain HMC1 and Flexibacter sp. strain HMC2,
nitrogen-fixing Pantoea sp. strain R1, Enterobacter sp.
strain R2, Klebsiella sp. strain R3 and Klebsiella sp. strain
R4 in N-free semisolid hemicellulose medium showed
extremely low ARA (Table 6). In contrast, the nitrogenase
activity of the consortium of Agrobacterium sp. strain
HMC1 or Flexibacter sp. strain HMC2 and NFB, both types
of bacteria were isolated from the same sago root sample,
showed markedly stimulated NFA (Table 6). The highest
level of activity, 38 nmol C2H4 culture−1 h−1, was measured
in the co-culture of P. agglomerans strain R1 plus Flexi-
bacter sp. strain HMC2, followed by that of Pantoea sp.
strain R1 plus Agrobacterium sp. strain HMC1 (31 nmol
C2H4 culture−1 h−1). No ARA was measured on the single
inoculation of Agrobacterium sp. strain HMC1 and Flexi-
bacter sp. strain HMC2 in N-free Rennie medium (data not
shown).
NFA with pectin-degrading bacteria
The ARA of NFB (Pantoea sp. strain R1, Enterobacter
sp. strain R2, Klebsiella sp. strain R3, Klebsiella sp. strain
R4) in individual cultures was very weak (0–0.7 nmol C2H4
culture−1 h−1) or almost non-existence. NFB in the consor-
tium with pectin-degrading Burkholderia strain sp. BT1
(0.2–0.4 nmol C2H4 culture−1 h−1) and with Paenibacillus sp.
P1 strain (0.3–0.7 nmol C2H4 culture−1 h−1) didn’t show
stimulated ARA.
Table 4. ARA of co-cultures of nitrogen-fixing bacteria and non-nitrogen-fixing bacteria
Root isolatesAcetylene-reducing activity (nmol C2H4 culture−1 h−1)
Single +Group A +Group B +Group C +Group D +Group E
Pantoea sp. strain R1 17±14a) 180±23* 49±11 210±12* 230±25** 190±31*
Enterobacter sp. strain R2 2.1±1.4 120±12** 95±21** 130±6.5** 71±12** 42±12*
Klebsiella sp. strain R3 31±17 130±11** 120±1.2** 57±11* 81±16** 48±7.1
Klebsiella sp. strain R4 16±0.8 150±12** 81±16** 190±6.3** 280±15** 190±15**
Midrib isolate +Group F +Group G
Pantoea sp. strain M 21±6.1 67±11** 31±2.8
Trunk isolates +Group H +Group I +Group J
Klebsiella sp. strain T1 4.5±1.2 59±4.6** 92±13** 20±1.1
Klebsiella sp. strain T2 16±1.9 60±2.1** 99±2.8** 21±1.6
a) Mean values in triplicate
* Significant at P<0.05, ** significant at P<0.01
Each of the groups contained 5 non-nitrogen-fixing bacteria
Groups A, B, C, D and E were isolated from sago root
Groups F and G were isolated from sago midrib
Groups H, I and J were isolated from sago trunk
NT: Not tested.
Table 5. Effect of co-culture with a starch-degrading bacterium on nitrogen-fixing activity (acetylene-reducing activity) and population density
of NFB and Bacillus sp. strain B1
Strain
Acetylene-reducing activity
(nmol C2H4 culture−1 h−1)Population density (cfu ml−1) of
NFB NFB Bacillus sp. strain B1
Single culturewith Bacillus sp.
strain B1Single culture
with Bacillus sp.
strain B1with NFB
Enterobacter sp. strain S1 0.1±0.2 270±15** 0.2×106 1.4×108 2.0×108
Klebsiella sp. strain S2 1.5±0.1 260±6.5** 2.3×106 0.9×108 1.4×108
Pantoea sp. strain S3 0 150±4.5** 0.2×106 0.1×108 0.1×108
Bacillus sp. strain B1 0 3.0×108
Microbial Consortium Enhance N-fixation 67
NFA under reduced oxygen concentrations and in cell
filtrates of indigenous bacteria
Under normal atmospheric conditions, the ARA of NFB
varied from 25 to 44 nmol C2H4 culture−1 h−1 (Table 7). In
contrast, all of the NFB tested in this experiment showed
stimulated ARA under reduced oxygen concentrations. The
amount of oxygen in the reduced oxygen conditions was
268 µmol (equivalent to 67% of removed oxygen from the
vial) and the most ARA under such conditions was exhib-
ited by Klebsiella sp. strain R4 (120 nmol C2H4 culture−1 h−1).
In the other NFB listed in Table 7, increased NFA was
observed, ranging from 65 to 110 nmol C2H4 culture−1 h−1.
The ARA of NFB in the cell-free filtrate fluids of each
group of indigenous microbes was found not to be stimu-
lated (data not shown), compared to the ARA measured
under the reduced oxygen conditions.
Discussion
We previously reported the existence of NFB (K. pnue-
moniae, K. oxytoca, E. cloacae, and P. agglomerans) in dif-
ferent parts of sago palm40). In this study we again isolated
NFB belonging to the same species from sago samples. The
sequence similarity of our previously isolated NFB (Kleb-
siella sp. strain RD3, Klebsiella sp. strain RD4, Klebsiella
strain sp. RH2, Klebsiella sp. strain RH1, Enterobacter sp.
strain RA1 and Pantoea sp. strain RA2) and the strains iso-
lated in this study showed 99–100% homology among K.
pneumoniae strains, 98.4 to 100% among E. cloacace
strains and 100% among K. oxytoca strains and P. agglom-
erans strains. This suggested the generality of the identified
species colonizing sago palm.
In an attempt to confirm the existence of isolated NFB in
the plant tissues of sago palm, PCR-DGGE patterns target-
ing the nifH gene were compared between the isolated
strains and the community DNA samples from which the
strains were isolated. The presence of several dominant
bands directly reflected the existence of NFB in the sago
root and starch (Fig. 2). Out of four strains tested (K. pnue-
moniae, P. agglomerans, E. cloacae and K. oxytoca), identi-
cal band positions of the former three strains were detected
in the root and starch samples, suggesting that these NFB
may be colonizing the sago root and starch. The nifH
sequences from the excised DGGE bands (Fig. 2., band “d”,
“e” and “l”) were identical with those from pure culture
NFB (Fig. 2., band “A”, “B” and “D”), further supporting
that these NFB are truly colonizing the sago root and starch.
The occurrence of non-identical bands in DGGE (Fig. 2.
band “a”, “b” and “c” in sago root and “g” and “h” and “i”
in sago starch) as compared to the tested strains indicated
that there may exist some other NFB that were not isolated
by the culture-based approach in this study. PCR amplifi-
cation of nifH sequences, followed by their separation
through DGGE, has been used to examine the complexity
Table 6. Effect of co-culture with hemicellulose-degrading bacteria on nitrogen-fixing activity (acetylene-reducing activity)
Strain
Acetylene-reducing activity (nmol C2H4 culture−1 h−1)
Single cultureculture with:
Agrobacterium sp. strain HMC1 Flexibacter sp. strain HMC2
Pantoea sp. strain R1 0.1±0.5a) 31±15* 38±13*
Enterobacter sp. strain R2 0.1±0.1 28±14** 28±2.7**
Klebsiella sp. strain R3 0.1±0 16±13** 26±3.1**
Klebsiella sp. strain R4 0 30±13** 25±4.1**
a) Mean values in triplicate.
Table 7. Effect of oxygen concentration on nitrogen-fixing activity
(acetylene-reducing activity)
Strain
Acetylene-reducing activity
(nmol C2H4 culture−1 h−1)
Normal Reduced
Pantoea sp. strain R1 41±5.3a) 85±19*
Enterobacter sp. strain R2 26±17 97±8.4**
Klebsiella sp. strain R3 42±0.4 110±23*
Klebsiella sp. strain R4 25±0.5 120±4.4**
Enterobacter sp. strain S1 41±16 72±2.3*
Klebsiella sp. strain S2 33±15 65±3.6**
Pantoea sp. strain S3 41±0.7 81±8.7**
Pantoea sp. strain M 36±4.9 72±14**
Klebsiella sp. strain T1 40±16 95±2.7**
Klebsiella sp. strain T2 44±1.5 72±7.9**
a) Mean values in triplicate.

SHRESTHA et al.68
and stability of the diazotroph assemblage found in the
Spartina rhizosphere35). Several other studies4,6,7,34,38,41,45,47)
also showed that the analysis of partial nifH gene sequences
can provide information on the phylogeny and composition
of a diazotrophic community.
In our study, the nitrogenase activity of the consortium
of NFB with indigenous bacteria was remarkably increased
in all the groups of the consortium (Table 4). Many con-
sortium-based studies have suggested stimulated
ARA19,20,23,26,37,48). The evidence presented here also supports
the hypothesis that nitrogen fixation in a naturally occurring
bacterium is usually affected by interactions with other
microbes44). The reduction in the concentration of oxygen in
the culture medium by indigenous dominant bacteria could
result in anaerobic condition that could be preferred by
NFB. In contrast, it is known that the presence of growth
factor(s) in the cell filtrate of a bacterium can promote the
growth and function of the accompanying microbes42).
Therefore, the mechanisms whereby indigenous bacteria
enhance nitrogen fixation were estimated by measuring the
ARA of NFB (i) at a reduced oxygen level and (ii) in cell-
free filtrate fluids from a 48-h broth culture of indigenous
bacteria. Increased NFA at the reduced oxygen level (67%
reduction) suggested that the sago nitrogen fixers favor low
oxygen levels for high NFA (Table 7), like in the papers by
van de Broek et al.46) and Zlotnikov et al.48), who found that
most diazotrophs decreased their NFA under aerobic con-
ditions. Minamisawa et al.29) reported that Clostridium sp.
was able to grow and fix nitrogen after the accompanying
bacteria had eliminated the oxygen by respiration. No or
very weak ARA of NFB was detected in cell-free filtrates of
indigenous bacteria, suggesting that enhanced NFA through
the metabolites by indigenous bacteria may not be the case.
This result further suggested that sago NFB might favor low
oxygen levels for expressing high NFA. Haahtela et al.10)
reported that anaerobic conditions were required for maxi-
mum nitrogenase activity of facultatively anaerobic NFB
like K. pnuemoniae and E. agglomerans. It is known that
dinitrogen-fixing bacteria grow best in the presence of other
heterotrophic bacteria which may stimulate the nitrogen-fix-
ing bacteria and in turn receive required N compounds44).
Zlotnikov et al.48) showed a binary association of Bacillus
firmus E3 (promoter) and Klebsiella terrigena E6 (nitrogen
fixer), both of which were isolated from the rhizosphere of
Dactylus glomerata: 23.7 (single culture) to 111.2 (dual cul-
ture) nmol C2H4 culture−1 h−1. Holguin et al.20) reported that
the NFA that occurred in the rhizosphere of mangrove
plants was probably not the result of the individual nitro-
gen-fixing strains, but the sum of interactions between
members of the rhizosphere community.
In this study, a consortium of starch-degrading bacteria,
Bacillus sp. strain B1 plus NFB isolated from the sago
starch, showed markedly increased nitrogenase activity in
N-free medium containing starch as a sole C source (Table
5). This result suggested that Bacillus sp. strain B1
degraded starch into cellobiose or glucose, which could be
utilized by NFB for nitrogen fixation. In a different experi-
ment, we measured the ARA of NFB in glucose and cellobi-
ose. The activity varied from 4.2 to 9.1 and 0 to 2.1 nmol
C2H4 culture−1 h−1 in glucose and cellobiose, respectively
(Table 2), suggesting that glucose was preferably utilized by
NFB. The degradation of starch might have a beneficial
effect on the nitrogen fixation system of sago palm. This
was supported by the result that the population numbers of
NFB and Bacillus sp. strain B1 in single cultures were
increased by dual culture with Bacillus sp. strain B1 and
NFB, respectively.
The increased NFA in the consortium of hemicellulose-
degrading microbes, Agrobacterium sp. strain HMC1, and
Flexibacter sp. strain HMC2 plus NFB in hemicellulose-
based N-free medium, suggested that these microbes
degraded hemicellulose and released products like glucose,
galactose, mannose, and cellobiose which may be further
utilized by NFB. Flexibacter spp. are common soil and
freshwater saprophytes species, and none have been defined
as pathogens28). The genus Flexibacter spp. requires com-
plex media for good growth and digests polysachharides
like cellulose, agar, and chitin28). In our study, the consor-
tium of Agrobacterium sp. strain HMC1 and Flexibacter sp.
strain HMC2 plus NFB showed enhanced NFA when mea-
sured in semi-solid N-free hemicellulose media. That there
was no nitrogenase activity of pure NFB in hemicellulose
media also indicated that the hemicellulose was not utilized
by NFB alone for nitrogen fixation (Table 3). Hemicellulose
which constitutes the most abundant constituent of plants
especially in the roots24), is made up of water-insoluble
polymers consisting of various hexoses and pentoses17) and
can be degraded by bacteria, actinomycetes, and fungi24).
According to our study, the degradation of hemicellulose by
indigenous microbes existing in the natural habitat of sago
palm, could have a beneficial effect on NFA in sago palm.
The consortium of Cellvibrio sp. (hemicellulose-degrading
bacteria) plus Clostridium butyricum, studied in hemicellu-
lose medium as a form of crude xylan, showed a significant
increase in NFA21). Halsall et al.13) reported that straw
(hemicellulose major component) could not be utilized as
an energy source for nitrogen fixation by pure culture NFB,
but Halsall and Gibson11) found enhanced NFA using straw

Microbial Consortium Enhance N-fixation 69
as a carbon source in a consortium of Azospirillum spp.
(NFB) and Cellulomonas gelida.
No enhanced nitrogenase activity was observed in the
consortium of NFB plus Burkholderia sp. strain BT1 and
NFB plus Paenibacillus sp. strain P1, suggesting that col-
laborative nitrogen fixation using pectin might not occur in
the sago palm. Pectin is a complex of polymers present in
the cell wall of higher plants24) and the mucilage found in
root tips consists of polygalactoruonic acid, a pectin-like
mucus8). However, the study of carbon utilization ability
suggested that neither pectin and nor its monomer (polyga-
lactouronic acid) were utilized by NFB for nitrogen fixation
(Table 3). Hence, we might conclude from our laboratory
experiment that in the sago palm’s natural environment
there might be little possibility of utilizing pectin as a
substrate for NFA by NFB living in a consortium or as indi-
viduals. However, Khammas and Kaiser23) reported that
consortiums of different Azospirillum spp. with Bacillus
polymyxa or B. subtilis enabled the efficient utilization of
pectin as carbon and energy sources for nitrogen fixation.
In most previous studies about consortiums of NFB and
non-NFB, the origin and habitats were not clear9,11,13,14) and
therefore, whether beneficial interactions occur or not in
natural microhabitats remains unkown. In our study, all of
the NFA of different Enterobacteriaceae strains and of
different indigenous microbes used in the consortium study
was isolated from the same sago palm. Therefore, the bene-
ficial interactions between these microbes may occur in the
sago palm’s natural microhabitats.
Our overall results suggested the importance of bacterial
associations and this collaborative fixation of nitrogen by
NFB and indigenous bacteria could enable them to sustain
high levels of nitrogen acquisition in natural environments.
In this study, some of the genera of the indigenous nitrogen-
fixing bacteria associated with sago palm have not been iso-
lated (the bands a, b, c, h, i in Fig. 2). It is well known that
there are many uncultivable microorganisms in the natural
environment22,39). In addition, no efforts were made to iso-
late slow growing or anaerobic microorganisms in this
study, that could play an important role in some beneficial
microbial interactions22). Further study is, therefore, neces-
sary to evaluate their contribution to the nitrogen fixation
process in sago palm.
Acknowledgements
Archana Shrestha is the recipient of a scholarship from the
Jinnai International Study Exchange Program. We would like
to express special thanks to the sponsors of that program.
References
1) Adachi, K., Y. Takahata and Constancio A. Asis, Jr. 2004. Occur-
rence of diazotrophic endophytes in different portions of sweet
potato stems. Microbes Environ. 20: 40–44.
2) Atlas, R.M. and R. Bartha. 1993. Interactions among microbial
populations, pp. 37–38. In B. Brady and D. Lisa (eds.), Microbial
ecology: fundamentals and applications. The Benjamin/Cum-
mings Publishing Company, Inc., California, C.A.
3) Baldani, J.L., V.L.D. Baldani, L. Seldin and J. Doberenier. 1986.
Characterization of Herbaspirillum seropediacecae gen. nov., sp.
nov., a root-associated nitrogen-fixing bacteria. Int. J. System.
Bacteriol. 36: 86–93.
4) Brown, M.M., M.J. Friez and C.R. Lovell. 2003. Expression of
nifH genes by diazotrophic bacteria in the rhizosphere of short
form of Spartina alterniflora. FEMS Microbiol. Ecol. 43: 411–417.
5) Chulavatanol, M. 2002. Starch utilization in Asia, pp. 9–14. In K.
Kainuma, M. Okazaki, Y. Toyoda and J.E. Cecil (eds.), New
frontiers of sago palm studies. Universal Academy Press, Tokyo,
Japan.
6) Diallo, M.D., A. Willems, N. Vloemans, S. Cousin, T.T. Vande-
kerchove, P. Lajudie, M. Neyra, W. Vyverman, M. Gillis and
K.V. Gucht. 2004. Polymerase chain reaction denaturing gradient
gel electrophoresis analysis of the N2-fixing bacterial diversity in
soil under Acacia tortilis ssp. raddiana and Balanites aegyptiaca
in the dryland part of Senegal. Environ. Microbiol. 6: 400–415.
7) Deslippe, J.R., K.N. Egger and G.H.R. Henry. 2005. Impacts of
warming and fertilization on nitrogen-fixing microbial communi-
ties in the Canadian high arctic dwarf shrubs. FEMS Micrbiol.
Ecol. 53: 41–50.
8) Elsas, D.J., J.T. Trevor and E.M.H. Wellington. 1997. The rhizo-
sphere as a habitat for soil microorganisms, pp. 21–23. In J.D.V.
Elsas, J.T. Trevor and E.M.H. Wellington (eds.), Modern soil
microbiology. Marcell Dekker Inc, New York. N.Y.
9) Groppa, M.D., M.S. Zawoznik and M.L. Tomaro. 1998. Effect of
co-inoculation with Bradyrhizobium japonicum and Azospirillum
brasilense on soybean plants. Eur. J. Soil. Biol. 34: 75–80.
10) Haahtela, K.T., K. Kari and V. Sundman. 1983. Nitrogenase
activity of root associated, cold climate Azozpirillum, Entero-
bacter, Klebsiella and Pseudomonas species during growth on
various sources and at various partial pressures of oxygen. Appl.
Environ. Microbiol. 45: 563–570.
11) Halsall, D.M. and A.H. Gibson. 1986. Comparison of two Cellu-
lomonas strains and their interaction with Azospirillum brasilense
in degradation of wheat straw and associated nitrogen fixation.
Appl. Environ. Microbiol. 51: 855–861.
12) Halsall, D.M. and A.H. Gibson. 1989. Nitrogenase activity of a
range of diazotrophic bacteria on straw, straw breakdown prod-
ucts and related compounds. Soil Biol. Biochem. 21: 291–298.
13) Halsall, D.M., G.L. Turner and A. Gibson. 1985. Straw and xylan
untilization by pure cultures of nitrogen-fixing Azospirillum spp.
Appl. Environ. Microbiol. 49: 423–428.
14) Harper, S.H.T. and J.M. Lynch. 1984. Nitrogen fixation by cellu-
lolytic communities at aerobic-anaerobic interfaces in straw. J.
Appl. Bacteriol. 57: 131–137.
15) Harayadi. 2002. The current status and future prospect of sago
palm in Java, pp. 37–42. In K. Kainuma, M. Okazaki, Y. Toyoda
and J.E. Cecil (eds.), New frontiers of sago palm studies. Univer-
sal Academy Press, Tokyo.
16) Hasan, F.H. 2002. Agronomic practice in cultivating the sago
palm, Metroxylan sagu, Rottb-the Sarawak experience, pp. 37–
42. In K. Kainuma, M. Okazaki, Y. Toyoda and J.E. Cecil (eds.),

SHRESTHA et al.70
New frontiers of sago palm studies. Universal Academy Press,
Tokyo.
17) Hattori, T. 1973. Plant growths and microbes in the soil, pp. 315–
346. In A.D. McLaren (ed.), Microbial life in the soil, an intro-
duction. Marcel Dekker Inc, New York, N.Y.
18) Heuer, H., M. Krsek, P. Baker, K. Smalla and E.M. Wellington.
1997. Analysis of actinomycetes communities by specific ampli-
fication of genes encoding 16S rRNA and gel-electrophoretic
separation in denaturing gradients. Appl. Environ. Microbiol. 63:
3233–3241.
19) Holguin, G., M.A. Guzman and Y. Bashan. 1992. Two new nitro-
gen-fixing bacteria from the rhizosphere of mangrove trees: their
isolation, identification and in vitro interaction with rhizosphere
Staphylococcus sp. FEMS Microbiol. Ecol. 101: 207–216.
20) Holguin, G. and Y. Bashan. 1996. Nitrogen fixation by Azospiril-
lum brasilense Cd is promoted when co-cultured with a man-
grove rhizopshere bacterium (Staphyloccocus sp.). Soil Biol. Bio-
chem. 28: 1651–1660.
21) Jensen, H.L. 1941. Nitrogen fixation and cellulose decomposition
by soil microorganisms. III. Department of Bacteriology, Univer-
sity of Sydney.
22) Kamagata, Y. and H. Tamaki. 2005. Cultivation of uncultured
fastidious microbes. Microbes Environ. 2: 85–91.
23) Khammas, K.M. and P. Kaiser. 1992. Pectin decomposition and
associated nitrogen fixation by mixed cultures of Azospirillum
and Bacillus species. Can. J. Microbiol. 38: 794–797.
24) Lack, A.J. and D.E. Evans. 2001. The plant cell wall, pp. 6–7. In
B.D. Hames (ed.), Instant notes: plant biology. BIOS Scientific
Publishers Limited, U.K.
25) Lane, D.J. 1991. 16S/23S rRNA sequencing, pp. 115–175. In E.
Stackebrandt and M. Goodfellow (eds.), Nucleic acid techniques
in bacterial systematics. John Wiley and Sons, New York, N.Y.
26) Lippi, D., I. Cacciari, T. Pietrosanti and W. Pietrosanti. 1992.
Interactions between Azospirillum and Arthrobacter in diaz-
otrophic mixed culture. Symbiosis. 12: 107–114.
27) Lovell, C.R., Y.M. Piceno, J.M. Quattro and C.E. Bagwell. 2000.
Molecular analysis of diazotroph diversity in the rhizosphere of
the smooth cordgrass, Spartina alterniflora. Appl. Environ.
Microbiol. 66: 3814–3822.
28) Madigan, T.M., J.M. Martinko and J. Parker. 1984. Evolutionary
microbiology and microbial diversity: Prokaryotic diversity: bac-
teria, pp. 431–432. In G. Carlson, S.L. Snavely and D.A.
Wescherl (eds.), Brock biology of microorganisms. Pearson Edu-
cation, New Jersey, N.J.
29) Minamisawa, M., K. Nishioka, T. Miyaki, B. Ye, T. Miyamoto,
M. You, A. Saito, M. Saito, W.L. Barraquio, N. Teaumroong, T.
Sein and T. Sato. 2004. Anaerobic nitrogen-fixing consortia con-
sisting of clostridia isolated from gramineous plants. Appl. Envi-
ron. Microbiol. 70: 3096–3102.
30) Miyashita, K. 1992. Dojyobiseibutsujikkenhou, pp. 163–172. In
Nihon dojobiseibutsukenkyuukai (ed.), Yokendo, Tokyo (in Japa-
nese).
31) Morimoto, S., K. Togami, N. Ogawa, A. Hasebe and T. Fujii.
2005. Analysis of a bacterial community in 3-chlorobenzoate-
contaminated soil by PCR-DGGE targeting the 16S rRNA gene
and benzoate 1, 2-dioxygenase gene (benA). Microbes Environ.
20: 151–159.
32) Muthukumarasamy, R., I. Cleenwerckb, G. Revathi, M. Vadi-
velu, D. Janssens, B. Hoste, K.U. Gumc, K.D. Park, C.Y. Son, T.
Sa and J. Caballero-Mellado. 2005. Natural association of Glu-
conacetobacter diazotrophicus and diazotrophic Acetobacter per-
oxydans with wetland rice. Syst. Appl. Microbiol. 28: 277–286.
33) Oates, G.C. 2002. Sago starch production in Asia and Pacific-
problems and prospects, pp. 27–36. In K. Kainuma, M. Okazaki,
Y. Toyoda and J.E. Cecil (eds.), New frontiers of sago palm stud-
ies. Universal Academy Press, Tokyo, Japan.
34) Ohkuma, M., S. Noda, R. Usami, K. Horikoshi and T. Kudo.
1996. Diversity of nitrogen fixation genes in the symbiotic intes-
tinal microflora of the termite Reticultermes speratus. Appl.
Environ. Microbiol. 62: 2747–2752.
35) Piceno, Y.M. and C.R. Lovell. 2002. Stability in natural bacterial
communities: nutrient addition effects on rhizosphere diazotroph
assemblage composition. Microb. Ecol. 39: 32–40.
36) Rennie, R.J. 1981. A single medium for isolation of acetylene-
reducing (dinitrogen-fixing) bacteria from soils. Can. J. Micro-
biol. 27: 8–24.
37) Rojas, A., G. Holguin, B.R. Glick and Y. Bashan. 2001. Syner-
gism between Phyllobacterium sp. (N2-fixer) and Bacillus
licheniformis (P-solubilizer), both from a semiarid mangrove
rhizosphere. FEMS Microbiol. Ecol. 35: 181–187.
38) Saito, A. and K. Minamisawa. 2006. Evaluation of nitrogen-
fixing ability of endophytic clostridia based on acetylene reduc-
tion and reverse transcription-PCR targeting the nifH transcript
and ribosomal RNA. Microbes Environ. 21: 23–35.
39) Sekiguchi, Y. 2006. Yet-to-be cultures microorganisms relevant
to methane fermentation. Microbes Environ. 21: 1–15.
40) Shrestha, A., K. Toyota, Y. Nakano, M. Okazaki, M.A. Quevedo,
A.B. Loreto, A.M. Mariscal and E.I. Abayon. 2006. Nitrogen-
fixing activity in various parts of sago palm (Metroxylan sagu)
and characterization of aerobic nitrogen-fixing bacteria coloniz-
ing the sago palm. Sago Palm. 14: 20–32.
41) Soares, R.A., L.F.W. Roesch, G. Zannatta, F.A.O. Camargo and
L.M.P. Passaglia. 2006. Occurrence and distribution of nitrogen-
fixing bacterial community associated with oat (Avena sativa)
assessed by molecular and microbiological techniques. Appl.
Soil. Ecol. 33: 221–234.
42) Tanaka, Y., S. Hanada, H. Tamaki, K. Nakamura and Y. Kama-
gata. 2005. Isolation and identification of bacterial strains pro-
ducing diffusible growth factors(s) for Catellibacterium necta-
riphilum strain AST4T. Microbes Environ. 20: 110–116.
43) Tarimo, A., Y. Takamura, H. Runkulatile and K. Osozawa. 2002.
Introducing the sago palm (Metroxylan sagu L.) to Tanzania, pp.
69–74. In K. Kainuma, M. Okazaki, Y. Toyoda and J.E. Cecil
(eds.), New frontiers of sago palm studies. Universal Academy
Press, Tokyo, Japan.
44) Tyler, M.E., J.R. Milan, R.L. Smith, S.C. Schank and D.A.
Zuberer. 1979. Isolation of Azospirillum from diverse geographic
regions. Can. J. Microbiol. 25: 693–697.
45) Ueda, T., Y. Suga, N. Yahiro and T. Matsuguchi. 1995. Remark-
able N-fixing bacterial diversity detected in rice roots by molecu-
lar evolutionary analysis of nifH gene sequences. J. Bacteriol.
177: 1414–1417.
46) van de Broek, A., V. Keijers and J. Vanderleyden. 1996. Effect of
oxygen on the free-living nitrogen fixation activity and expres-
sion of the Azopirillum brasilense nifH gene in various plant-
associated diazotrophs. Symbiosis. 21: 25–40.
47) Zehr, J.P., M. Mellon, S. Braun, W. Litaker, T. Stepper and H.W.
Pearl. 1995. Diversity of heterotrophic nitrogen fixation genes in
a marine cyanobacterial mat. Appl. Environ. Microbiol. 61:
2527–2532.
48) Zlotnikov, A.K., N.Y. Shapovalova and A.A. Makarov. 2001.
Association of Bacillus firmus E3 and Klebsiella terrigena E6
with increased ability for nitrogen fixation. Soil Biol. Biochem.
33: 1525–1530.