Embed Size (px)
Citation preview

11
In Vivo NMR, Applications, 31P
Ruth M Dixon and Peter Styles, University of Oxford, UK
& 2010 Elsevier Ltd. All rights reserved.
This article is reproduced from the previous edition, volume 2,
pp 851–857, & 1999, Elsevier Ltd., with an updated Further
Reading section supplied by the Editor.
Symbols
J12
coupling constant
T1
longitudinal relaxation timeT2
transverse relaxation timeIntroduction
The phosphorus nucleus has occupied centre stage in thedevelopment of in vivo NMR spectroscopy for twoimportant reasons. First, the NMR properties of thenucleus are well suited to in vivo NMR (100% naturalabundance, reasonable sensitivity and adequate but notexcessive chemical shift). Second, key metabolites, par-ticularly those involved in the production of energy,contain phosphorus and are present in substantial con-centrations in living tissue. It is for these reasons that thefirst NMR spectra of living cells (a suspension of ery-throcytes), and later of excised frog muscle, wereobtained using 31P NMR. In both instances, resonanceswere observed from adenosine triphosphate (ATP) andinorganic phosphate, the position of the latter being anindicator of intracellular pH. Additional peaks were seenfrom 2, 3-diphosphoglycerate (2, 3-DPG) in red bloodcells and creatine phosphate in skeletal muscle. Despitethe comparative simplicity of 31P NMR tissue spectra,the technique offers unique insight into the regulation ofcritical energetic pathways. The early studies of ex vivo
samples were soon extended to in vivo examination ofintact animals and later to humans, experiments whichwere made possible by the development of large super-conducting magnets, the use of local surface coil probesand the invention of a variety of techniques for localizingthe source of the NMR signal.
Techniques for 31P Spectroscopy In Vivo
Signal Acquisition and Spatial Localization
Perhaps the most critical step in the development of in
vivo spectroscopy was the realization that it was notnecessary to surround the ‘sample’ with a conventionalNMR probe, but that a small local coil (a surface coil)
could be placed against an animal to preferentiallyreceive signals from the tissues adjacent to this coil. Thesurface coil has two advantages, namely excellent sensi-tivity close to the coil and inherent localizing propertiesowing to the rapid fall-off of the magnetic flux away fromthe coil. However, when deep tissues are to be investi-gated, additional localization is required. In choosing anappropriate technique for obtaining 31P NMR spectrafrom a live animal or man, careful consideration must begiven to the spatial resolution and signal-to-noise ratiowhich can reasonably be expected. As the phosphorusnucleus has only 6% of the sensitivity of the proton, itfollows that these expectations will be somewhat moremodest than for 1H spectroscopy. The usual approach isto adopt the least demanding methodology appropriatefor any particular application. As a consequence, in manyinstances simple surface coil localization is used to giveadequate localization and provide the best signal-to-noise ratio in short acquisition times. When this simpleapproach is inadequate, a range of single and multiplevoxel methods can be considered, but care must be takento choose methods which work well at short echo timesto avoid excessive T2 relaxation and minimize problemsdue to the J modulation of peaks such as adenosinetriphosphate.
Dealing first with the single voxel approach, imageselected in vivo spectroscopy (ISIS) is frequently themethod of choice because the spins are kept pre-dominantly along the z axis, thus minimizing T2 and Jcoupling effects. Other localizing schemes (e.g. stimu-lated echo acquisition mode spectroscopy (STEAM) andpoint resolved spectroscopy (PRESS)) which are com-monly used for proton spectroscopy can give reasonableresults, but are more sensitive to the aforementionedproblems.
Where multivoxel localization is advantageous, che-mical shift imaging techniques can be applied in one, twoor three dimensions, but relatively poor spatial resolutionis obtainable and long acquisition times are usuallyrequired.
Finally, it will often be advantageous to implementhybrid protocols in which, for example, two dimensionsmight be defined by an ISIS sequence, and phase encodedchemical shift imaging (CSI) implemented along the

Figure 1 31P spectra of human organs (a) calf muscle, (b)
heart, (c) liver, (d) brain. Peak assignments: 1. PME, 2. Pi, 3.
PDE, 4. PCr, 5. ATP, 6. 2, 3-DPG. Reproduced with permission
of Academic Press from Fu, Schroeder, Jones, Santini, and
Gorenstein (1998) Journal of Magnetic Resonance 77: 577–582.
In Vivo NMR, Applications, 31P 1113
third axis, thus producing a set of spectra that representslices through a well-defined column of tissue.
Proton Decoupling
The quality of in vivo 31P spectra can be improved by theuse of proton decoupling. Two benefits are available. Someof the phosphorus spectral lines are significantly broadenedby scalar coupling to hydrogen. Irradiation of the protonsduring acquisition removes this interaction and sharpensthe spectra. The effect is particularly advantageous in thephosphomonoester (PME) and phosphodiester (PDE)regions of the spectrum where conventional acquisitionwill fail to resolve, for example, glycerophosphocholine andglycerophosphoethanolamine. Together with the increasedresolution, proton decoupling also increases the signal-to-noise ratio due to peak narrowing. A further increase insignal is available by low power decoupling during theinterpulse relaxation period. Here, the nuclear Overhausereffect (NOE) can result in improved sensitivity of up toabout 50% owing to interactions between the phosphorusnuclei and neighbouring protons.
Magnetisation Transfer
Under certain conditions, rates of chemical exchange canbe measured using saturation- or inversion-transfermethods. These work by magnetically labelling onechemical species and observing the transfer of thismagnetic signature to a second compound which iscoupled by chemical exchange. For the technique to beviable, there needs to be significant exchange during atime scale which is of the order of the longitudinalrelaxation time T1. Such reactions include the synthesisand hydrolysis of ATP (by measuring exchange betweenATP and inorganic phosphate) and the fast exchangecatalysed by the creatine kinase enzyme.
The In Vivo 31P NMR Spectrum
Figure 1 shows spectra from human skeletal muscle,heart, liver and brain. Each contains the three peaks fromATP and one from inorganic phosphate. ATP showsthree signals due to the different chemical environmentsof its three phosphorus atoms. These signals are furthersplit by 31P–31P J-coupling, but this splitting is notalways detectable in vivo owing to the relatively broadlines. These peaks are normally designated ‘ATP’ but alsocontain minor contributions from other nucleotides, suchas guanosine triphosphate, NADH, NADP, UDP-sugarsand ADP. The signal which contains the least con-tamination from other compounds is the one from thecentral (beta) phosphate, which appears at the upfieldend of the spectrum, and so this peak is generally usedfor the measurement of ATP levels. In addition, spectra
from muscle, heart and brain contain a prominent peakfrom creatine phosphate (PCr). This compound is absentfrom liver. Liver and brain spectra also contain sub-stantial peaks designated as phosphomonoester (PME)and phosphodiester (PDE). These regions of the spectracan contain metabolites involved in membrane metabo-lism and also phosphorylated sugars involved in glyco-lysis. An important feature of the 31P spectrum is that theposition of the inorganic phosphate (Pi) peak is pHdependent, thus providing a sensitive marker of intra-cellular acidity/alkalinity.
Biological Relevance of the Peaks
Adenosine triphosphate (ATP)
ATP is the universal energy carrier in all cells. Whenenergy is required for cell functions such as preservingionic gradients, muscular contraction, etc., the terminalphosphate is cleaved by ATPase enzymes to form ade-nosine diphosphate (ADP) and inorganic phosphate, anenergy-producing reaction:
ATPþH2O-ADPþ Pi
ATP is resynthesized by one of several pathways, mostusually by ATP synthesis using energy obtained fromoxidative or anaerobic metabolism.
Inorganic phosphate (Pi)
This compound may be considered as the waste productof ATP hydrolysis or conversely as a substrate for ATPsynthesis. It may also play a role in regulating the supplyof energy via oxidative metabolism in the cells’ mito-chondria. At physiological pH, Pi exists in two ionicforms in fast equilibrium via the reaction:
H2PO4�"HPO4
2� þHþ

Figure 2 A typical time course for changes in PCr and pH
during exercise and recovery in the human forearm muscle.
1114 In Vivo NMR, Applications, 31P
This reaction has a pKa of 6.75, and so the relative con-centrations of Pi
� and Pi2� vary with the pH. Owing to
the fast exchange between these two species, the positionof the Pi peak in the spectrum reflects the weightedaverage of the two chemical shifts, and is thus a directmeasure of pH.
Creatine phosphate (PCr)
This molecule is present in abundance in muscle, heartand brain cells. Through the fast equilibrium reactioncatalysed by creatine kinase, PCr buffers the ATP con-centration by phosphorylating ADP, the reaction being:
PCrþ ADPþHþ"ATPþ Cr
PCr is depleted during sudden energetic demands suchas vigorous muscle contraction.
Adenosine diphosphate (ADP)
ADP is a substrate for ATP production, and a product ofits hydrolysis. ADP stimulates respiration (oxidativemetabolism) – breakdown of ATP increases ADP whichin turn activates the processes which provide energy forATP synthesis. In tissues such as muscle, much of theADP is bound to macromolecules and therefore invisibleto NMR. However, the free (and therefore metabolicallyactive) ADP, usually present in only micromolar con-centrations, may be deduced using the creatine kinaseequilibrium (see above).
Phosphomonoesters (PME)
Certain intermediaries in the glycolytic pathway such asglucose and fructose phosphates resonate in this region ofthe spectrum and are sometimes detected. More usually,the PME peak comprises the lipid precursors phos-phocholine and phosphoethanolamine.
Phosphodiester (PDE)
This region also contains compounds involved in mem-brane metabolism, specifically glycerophosphocholine(GPC) and glycerophosphoethanolamine (GPE).
Broad resonances
In addition to the sharp peaks that have just been men-tioned, there will often be underlying broad ‘humps’ inthe phosphorus spectrum, arising from phosphorusmoieties that are immobile or tumbling very slowly.Phospholipid head groups in cell membranes produce abroad and asymmetric signal centred under the PDE partof the spectrum. Bone also produces a broad resonance,several kilohertz wide, which is easily visible in surfacecoil spectra of the head, but is not usually present inlocalized spectra due to its very short T2.
Applications in Skeletal Muscle
Skeletal muscle is a tissue that is particularly suited tostudy by 31P NMR. From a technical standpoint, muscleis an ideal target because it exists close to the bodysurface and is not obscured by any other pool of phos-phorus-containing metabolites. From a biochemicalstandpoint, muscle is especially interesting because itsenergetic demands vary by at least two orders of mag-nitude (from rest to exercise) and so it provides anexample of exquisite metabolic control which can bestudied non-invasively. Finally, muscle can be stressedreadily by exercise within the NMR instrument, thusfacilitating the real-time investigation of the dynamics ofenergetic regulation.
Animal studies of skeletal muscle often employ directstimulation of the sciatic nerve to induce contraction andassociated metabolic response. Basic metabolic controlcan be investigated using such a model, as can a widerange of disease-related processes. Genetically abnormalanimals provide models of diseases such as musculardystrophy and 31P NMR spectroscopy provides a tool forinvestigating metabolism, pH regulation and possibletherapeutic interventions.
In a typical human examination, a subject will per-form an exercise regime within the magnet, perhapssqueezing a handgrip or pressing down a loaded pedal.The NMR signals are usually collected using a surfacecoil placed over the exercising muscle and a series ofspectra are collected to follow the metabolic state before,during and after the exercise. A typical time course isshown in Figure 2. The several metabolite concentra-tions and pH are then used to quantitatively assess themetabolic response to the stress, and, by using models ofmetabolic control, unique insight can be gained into the

In Vivo NMR, Applications, 31P 1115
complementary processes of mitochondrial respirationand anaerobic metabolism.
Muscle Metabolism
Figure 3 is a block diagram of the main energy-produ-cing pathways in skeletal muscle (and other tissues,except that the creatine kinase pathways may not bepresent). In resting muscle, ATP is synthesized only byoxidative metabolism, but during exercise glycolysis(anaerobic metabolism) and PCr breakdown via thecreatine kinase reaction provide additional ATP pro-duction. Although only three compounds are directlydetected by 31P NMR, others may be deduced by makingassumptions based on data derived from biopsy analysis.In particular, an estimate of free [ADP] is obtained usingassumed values of ATP and total creatine concentrations,and applying the known value for the creatine kinaseequilibrium constant.
In using these data to investigate metabolic controlin vivo, two periods within an exercise regime are ofparticular importance. At the start of exercise, energy isproduced primarily by glycolysis and a breakdown ofcreatine phosphate. The latter is measured directly whilethe former produces lactate which is reflected in the pH.At the start of recovery, glycolysis stops, the energydemands fall dramatically and oxidative metabolismrestores the PCr. Using models for proton efflux andmetabolic control by ADP, it is possible to assess therelative contributions of the various energy pathways inboth normal and diseased muscle.
Diseases of Skeletal Muscle
In man, the understanding and quantification of muscleenergetics have facilitated the study and, sometimes,diagnosis of a wide range of muscle diseases. Often thesestem from inherited genetic abnormalities, including thedystrophies, mitochondrial myopathies and so-called
Figure 3 A schematic diagram of energy metabolism in
skeletal muscle, heart and brain. The creatine kinase pathway is
absent in liver.
energy storage diseases. Muscle can also be affected byarterial disease and systemic abnormalities secondary to,for example, renal and cardiac failure. Many studies havelooked at these phenomena and monitored the responseto training or therapeutic intervention.
Applications in the Heart
Basic NMR research of the heart has concentrated onstudies of the organ ex vivo (the excised heart, perfusedwith physiological buffer, can be maintained in a viableand beating condition for many hours). In vivo studies inboth animals and man are more difficult – the heart is amoving target, is filled with blood and overlaid withskeletal muscle which gives a similar 31P NMR signature.The approaches that have been applied in animal systemsoften involve exposing the organ and suturing a surfacecoil to the heart wall, thus facilitating the measurementof localized metabolic events. In man, single voxel ISIS issometimes used, or chemical shift imaging, usually as ahybrid experiment (see above) where spectra areobtained from parallel slabs of tissue. The biochemistryof the heart is in many ways similar to skeletal muscle.However, the dynamic range of energetic demand is lessextreme than in muscle and in normal controls it isdifficult to induce significant changes in the 31P spectrumfrom the heart. In the diseased heart, the main findingshave been that cardiac energetics in the resting subjectare substantially stable until the onset of heart failure,when PCr falls significantly. In patients with coronaryartery disease, metabolic levels can be modulated byexercise. As an alternative to exercise, the positive ino-trope atropine-dobutamine has been administered tonormal subjects, and at high cardiac output a reduction inthe PCr:ATP ratio has been observed.
Intracellular pH is difficult to measure because the Pi
peaks can be obscured by 2, 3-DPG in the blood.However, it is possible to overcome this problem usingproton decoupling to sharpen the resonances, or satura-tion transfer to identify the intracellular Pi
unambiguously.
Applications in Brain
Using animal models, many disorders of the brain havebeen studied, including the effects of ischaemia (e.g.models of stroke and birth asphyxia), systemic dis-turbances (e.g. hepatic encephalopathy) and geneticabnormalities (e.g. models of Duchenne dystrophy).Spatial localization is usually kept to a minimum – oftenthe scalp is removed to avoid contamination by signalsfrom the cranial musculature, and a single surface coil isused to obtain the 31P NMR signal either from the wholebrain or, by appropriate placing of the coil,

1116 In Vivo NMR, Applications, 31P
predominantly from a single hemisphere. Almostinvariably it is the energy metabolism of the brain whichis of interest.
There have been many studies using 31P NMR toassess disease processes in man. Perhaps the most inter-esting have been the investigation of birth asphyxia inneonates where energy metabolite levels and intracel-lular pH give direct measures of brain damage andprovide a good prognostic index. Other disorders whichhave been investigated include stroke, multiple sclerosis,epilepsy, hydrocephalus, tumours, dementias and sys-temic encephalopathies. In all cases there have beenreports of altered energy metabolism (reduced PCr orincreased Pi) and, in multiple sclerosis, changes havebeen observed in the broad phospholipid peak. The pHdata are somewhat more variable – acidic values havebeen reported in acute stroke and hydrocephalus, butalkaline values have been observed in chronic stroke andbrain tumours. One of the main limitations in performing31P NMR spectroscopy in the human brain is that thespatial resolution is inadequate for interrogating focallesions of moderate size. Nevertheless the global picturesthat have emerged give insight into the way that brainenergetics respond in disease.
Applications in Liver
The liver is relatively simple to study in vivo by 31Pspectroscopy. On a macroscopic level it is homogeneous,and localization is generally straightforward. Sincehepatocytes do not express creatine kinase, the livercontains no detectable phosphocreatine and so thedegree of contamination of the liver spectra by signalsfrom overlying muscle can be readily assessed (and insome cases corrected for).
Localization strategies have ranged from simple sur-face coil detection (with a hard 1801 inversion pulse atthe surface to suppress the muscle signal) throughFourier series window approaches and ISIS to one-, two-or three-dimensional chemical shift imaging. Protondecoupling and NOE irradiation have occasionally beenemployed, but the technical requirements and powerdeposition considerations have limited these to a fewcases. In animal studies, localization methods are oftenavoided by exposing the liver and using a surface coilplaced directly on the organ.
Liver Metabolism
The liver’s role is to maintain chemical homeostasis andit can, in certain circumstances, be stressed by means of ametabolic load. One example has been to administer anoral fructose load which causes minimal spectral changesin normal subjects but, at very low dosage, results in anincrease in the fructose-1-phosphate peak in patients
with fructose intolerance. Other metabolic stresses havebeen administered to investigate galactose intolerance,glycogen storage diseases and cirrhosis. The rat liver(in situ or perfused) has been studied to elucidate detailsof carbohydrate and fatty acid metabolism, sometimes bya combination of 13C and 31P NMR. The control of pHin the liver has also been studied.
Liver Diseases
Some common diseases that involve the liver have alsobeen studied by 31P NMR. These include alcoholic liverdisease, hepatitis, cirrhosis of various etiologies and livertumours. The findings have often allowed the diseased liverto be distinguished from the healthy one, but the changeshave not usually been specific to a particular disease andthe interpretation of the changes is often speculative.Overall, these studies have shown that the major long-termchanges in liver spectra involve the PME and the PDEsignals, which probably signify changes in phospholipidmetabolism (see below) while the energetics of the liver(assessed from the Pi:ATP ratio and the pH) remainsnormal until a very late stage of disease. Another area thatshows promise is in assessing the transplanted liver, eitherbefore or after transplantation.
Spectral Interpretation
The PME and the PDE signals in the liver spectrum areeach composed of several components, which cannotgenerally be resolved, except with the use of protondecoupling. Aqueous extracts show that the PME signalcontains primarily phosphoethanolamine and phos-phocholine, with contributions from 2, 3-DPG fromblood and other minor components such as AMP, glu-cose-6-phosphate, sn-glycerol-1-phosphate and 3-phos-phoglycerate. The PDE signal (at the low magnetic fieldstrengths used for whole-body studies) is mostly com-posed of signals from phospholipid headgroups, broa-dened by chemical shift anisotropy and proton–phosphorus dipolar coupling, and also contains signalsfrom GPE and GPC. These can be resolved by protondecoupling as can the PME signal, but this has rarelybeen undertaken.
Applications in Cancer
From the first 31P NMR studies of tumours in vivo, thedifferences between tumour spectra and those from othertissues were obvious. The tumour spectra were domi-nated by the PME signal, and the pH was found to behigher than in most other tissues. While these findingshad potential use in distinguishing cancerous from nor-mal tissue, it was hoped that subtle differences in thespectra would also help in determining the type or grade
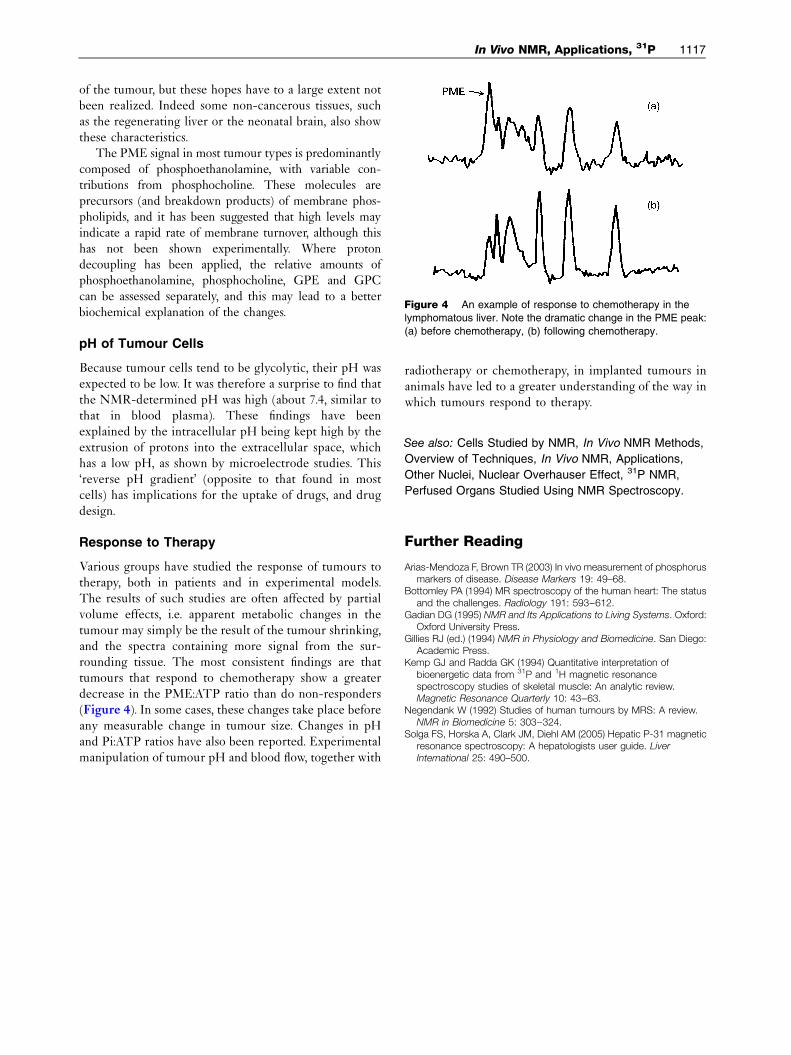
Figure 4 An example of response to chemotherapy in the
lymphomatous liver. Note the dramatic change in the PME peak:
(a) before chemotherapy, (b) following chemotherapy.
In Vivo NMR, Applications, 31P 1117
of the tumour, but these hopes have to a large extent notbeen realized. Indeed some non-cancerous tissues, suchas the regenerating liver or the neonatal brain, also showthese characteristics.
The PME signal in most tumour types is predominantlycomposed of phosphoethanolamine, with variable con-tributions from phosphocholine. These molecules areprecursors (and breakdown products) of membrane phos-pholipids, and it has been suggested that high levels mayindicate a rapid rate of membrane turnover, although thishas not been shown experimentally. Where protondecoupling has been applied, the relative amounts ofphosphoethanolamine, phosphocholine, GPE and GPCcan be assessed separately, and this may lead to a betterbiochemical explanation of the changes.
pH of Tumour Cells
Because tumour cells tend to be glycolytic, their pH wasexpected to be low. It was therefore a surprise to find thatthe NMR-determined pH was high (about 7.4, similar tothat in blood plasma). These findings have beenexplained by the intracellular pH being kept high by theextrusion of protons into the extracellular space, whichhas a low pH, as shown by microelectrode studies. This‘reverse pH gradient’ (opposite to that found in mostcells) has implications for the uptake of drugs, and drugdesign.
Response to Therapy
Various groups have studied the response of tumours totherapy, both in patients and in experimental models.The results of such studies are often affected by partialvolume effects, i.e. apparent metabolic changes in thetumour may simply be the result of the tumour shrinking,and the spectra containing more signal from the sur-rounding tissue. The most consistent findings are thattumours that respond to chemotherapy show a greaterdecrease in the PME:ATP ratio than do non-responders(Figure 4). In some cases, these changes take place beforeany measurable change in tumour size. Changes in pHand Pi:ATP ratios have also been reported. Experimentalmanipulation of tumour pH and blood flow, together with
radiotherapy or chemotherapy, in implanted tumours inanimals have led to a greater understanding of the way inwhich tumours respond to therapy.
See also: Cells Studied by NMR, In Vivo NMR Methods,
Overview of Techniques, In Vivo NMR, Applications,
Other Nuclei, Nuclear Overhauser Effect, 31P NMR,
Perfused Organs Studied Using NMR Spectroscopy.
Further Reading
Arias-Mendoza F, Brown TR (2003) In vivo measurement of phosphorusmarkers of disease. Disease Markers 19: 49–68.
Bottomley PA (1994) MR spectroscopy of the human heart: The statusand the challenges. Radiology 191: 593--612.
Gadian DG (1995) NMR and Its Applications to Living Systems. Oxford:Oxford University Press.
Gillies RJ (ed.) (1994) NMR in Physiology and Biomedicine. San Diego:Academic Press.
Kemp GJ and Radda GK (1994) Quantitative interpretation ofbioenergetic data from 31P and 1H magnetic resonancespectroscopy studies of skeletal muscle: An analytic review.Magnetic Resonance Quarterly 10: 43--63.
Negendank W (1992) Studies of human tumours by MRS: A review.NMR in Biomedicine 5: 303--324.
Solga FS, Horska A, Clark JM, Diehl AM (2005) Hepatic P-31 magneticresonance spectroscopy: A hepatologists user guide. LiverInternational 25: 490–500.
















![using 31P NMR - NREL · some background on the use of 31P NMR for analysis of bio-oils [1]. 2. Scope 2.1 This procedure has been optimized for the quantification of hydroxyls (-OH)](https://img.pdfslide.us/doc/110x75/5f0f5f5e7e708231d443d59e/using-31p-nmr-some-background-on-the-use-of-31p-nmr-for-analysis-of-bio-oils-1.jpg)












