Embed Size (px)
Citation preview

Comp. Biochenr. Physiol., 1975, Vol. 51A, pp. 863 to 813. Pergamon Press. Printed in Great Britain
EMBRYONIC TEMPERATURE PACIFIC TREEFROG,
ADAPTATIONS OF H YLA REGILLA
THE
HERBERT A. BROWN
Department of Biology, Western Washington State College, Bellingham, WA 98225, U.S.A.
(Received 20 May 1974)
Ah&act-l. The embryonic temperature adaptations of the Pacific treefrog, Hyla regilla, from southern California and western Washington are described. Each population represents a different subspecies, and both belong to the “thermal group” of H. regilia.
2. In western Washington the water temperatures of breeding pools during embryonic development may range from @5 to 20°C and embryonic development requires about 3 weeks.
3. Tbe embryonic temperature tolerance of Washington embryos is 628°C and California embryos, 6-2936°C.
4. The developmental rate-temperature curves cross with Washington embryos faster at low temperatures and California embryos faster at high temperatures.
5. Development of California embryos at high temperatures (29-31*X) shows inhibition of developmental rate and much variation in tolerance. There may be selection for increased heat resistance in this population.
6. Compared to many cold and warm adapted frogs of North America, H. regilla seems generalized in its embryonic temperature adaptations. It is a eurytopic treefrog, one found in a wide variety of habitats that are thermally diverse.
INTRODUCTION
ALTHOUGH there has been a recent upsurge of interest in the comparative reproductive biology of anuran amphibians (see Dent, 1968; Salthe & Duellman, 1973; Salthe & Mecham, 1974) much of the information about physiological adaptation of embryos to temperature is based on study of only a few families, mainly the Ranidae (true frogs) and Bufonidae (toads). There has been little information available concerning the embryonic temperature tolerance and developmental rate in treefrogs of the family Hylidae (see Bachmann, 1969). There are a few brief reports on developmental temperature tolerance: Ballinger & McKinney (1966) report pre- liminary experiments on Acris crepitans, Pseudacris clarki and Hyla cinerea from Texas; and Hubbs et al. (1963) studied Pseudacris streckerifrom Texas. Also, studies on the embryonic developmental rate of hylids are scanty. Volpe et al. (1961) described the development of Louisiana Hyla avivoca under laboratory conditions. Pettus & Angleton (1967) compared developmental rate (as time to hatching at different constant temperatures) between two populations of Pseudacris streckeri from northern Colorado. Zweifel (1968) studied embryonic temperature tolerance and developmental rate of Hyla arenicolor from southeastern Arizona.
The Pacific treefrog, Hyla regilla, is one of the most abundant and widespread amphibians of western North America. It ranges from southern
British Columbia to the tip of Baja California and from the Pacilic coast to western Montana and eastern Nevada. It is found in a variety of habitats ranging from wet coniferous forest, or even alpine meadows, to meandering streams in deserts (Cunningham & Mullally, 1956; Stebbins, 1966). Considering this extensive geographical and eco- logical distribution, sinificant geographic variation should be expected in this treefrog, and this has been shown in numerous studies.
Jameson et al. (1966) studied geographic variation of morphological characters in H. regilla and suggested that ten subspecies could be identified and organized into three groups: (1) thermal forms, (2) pluvial forms and (3) mountain forms. Further- more, they suggested that the results of genetic, developmental, physiological and ecological studies should be interpreted after an understanding of the taxonomy and past historical distribution of this treefrog were established.
Already further information on geographic variation in H. regilla has heen gathered by studying color polymorphism (Resnick & Jameson, 1963; Jameson & Pequegnat, 1971), morphology (Calhoon & Jameson, 1970; Vogt & Jameson, 1970; Jameson et al., 1973), metabolism (Jameson et al., 1970), water balance (Jameson, 1966), heat tolerance (CTM) of adults (Brattstrom, 1968) and mating calls (Snyder & Jameson, 1965; Ball & Jameson, 1966).
863

864 HERBERT A. BROWN
The present study reports on the embryonic temperature adaptations of H. regilla by comparing the temperature tolerance and developmental rate between a northern and southern population of this treefrog.
MATERIALS AND METHODS
All adult treefrogs used in this study were collected from their breeding pools during February through April at two localities: (1) San Jacinto Valley (near Lakeview), Riverside County, California, and (2) Nooksack Vallev (near Ferndale). Whatcom County, Washington. Foilowing the taxond&c study of James& et al. (1966) the southern California population represents H. r. hypochondriaca and the western Washington population, H. r. regilla, and both subspecies are members of the “thermal group” of H. regillu.
Ovulation was induced by injecting either preserved frog (female Runapipiens) pituitary glands or Antuitrin-S (chorionic gonadotropin, human, from Parke-Davis Co.) into freshly collected mature females (Rugh, 1962). After a lapse of 12-14 hr, the egg masses were directly stripped into a sperm suspension prepared by macerating a pair of testes in 20ml of 10% Ringer’s solution (IS- 1 SOC).
Embryos were cultured in glass dishes containing 3OOml of filtered pond water. About twenty-five eggs, in small clusters of three or four, were kept in each container. When the fertilized eggs were in first cleavage the culture bowl was transferred to a constant tempera- ture incubator (water-bath, forced air incubator or refrigerator).
The analysis of embryonic temperature adaptations follows the methods of Zweifel (1968). Temperature tolerance was determined by exposing fertilized eggs at first cleavage to constant high or low temperatures (+0*2”C). Lethal temperatures are detined as those below which (lower limiting) or above which (upper limiting) fewer than 50 per cent of the embryos develop normal external morphology. Rates of embryonic development were determined by staging the embryos according to Gosner (1960): stage 3 (first cleavage) considered as zero time and stage 20 (gill circulation) as the end point.
RESULTS
1. Embryonic development in natural pools
The breeding season of H. regilla, considering its entire range, extends from January to July (Stebbins, 1966). In southern California reproduction of low- land populations may begin in early January and continue intermittently until May, but the greatest reproductive activity generally occurs during Feb- ruary through April. In western Washington reproduction begins in late February and continues to late May with greatest activity in March and April. Thus, despite the latitudinal separation, both populations of this study have a similar breeding season.
The significance of experimental results of temperature tests on amphibian embryos must be
evaluated in relation to the micro-climatic tempera- tures that developing embryos experience in the natural spawning pools (Zweifel, 1968). The work of Herreid & Kinney (1967) is a model study of amphibian development examined from an eco- logical point of view.
I studied the development of H. regiZIa eggs in natural pools in western Washington by determining the rate of embryonic development and measuring water temperatures near developing egg masses. During the 2 years of study, the treefrogs appeared at one temporary pool on 20 February 1968 and 5 March 1969, and egg deposition began im- mediately. This temporary pool had a bottom of mud and gravel and was approximately 10 x 30 m with a maximum depth of 30 cm. I found most egg masses attached to submerged grass at the shallow margins of the pool. The data of embryonic develop- ment in the temporary pool (Fig. 1) were collected by repeated sampling of marked egg masses during a period of 4 weeks. In 1968 I placed maximum- minimum thermometers near the egg masses at a depth of about 15 cm and measured the range of water temperatures that embryos experienced during their development.
The developmental rate of eggs laid early in the breeding season of 1968 and 1969 (Fig. 1) was similar, although the embryos hatched at a later date in 1969. Average time from first cleavage (stage 3) to gill circulation (stage 20) was about 3 weeks (20 days in 1968 and 23 days in 1969). The cleavage period (stages 3-10) was 5-6 days, and gastrulation (stages lO-13), 4-5 days, and organogenesis (stages 13-20), about 12 days. Storer (1925) studied the reproduction of H. regifla in natural pools near Berkeley, California, and found that embryonic development to the hatching stage requires about 2 weeks.
The microclimatic temperatures in the temporary pool where the embryos were developing ranged from a low of 0.5”C to a high of 2@2”C. The average minimum temperature was 49°C and the average maximum, 13.7”C; an average pool tempera- ture of 9°C was estimated from the median value of maximum-minimum recordings. In the laboratory, development of eggs from first cleavage to gill circulation at a constant temperature of 9°C required 519 hr or 21.6 days. Thus, there is close agreement in developmental rate between eggs developing in natural pool conditions and those experimental groups reared at constant temperature in the laboratory. Apparently, the fluctuating temperatures in the pool did not significantly accelerate or retard development of eggs when compared to the laboratory group.
I did not study the embryonic development of H. regiila eggs in natural pools in California, but there are data on water temperatures associated with chorusing and egg deposition. In the San Jacinto Valley I found chorusing treefrogs from December to

Tern~ra~u~ and development of treefrog 865
Fig, 1. Embryonic development of Washington H. regillu eggs in a temporary pool. show total variation in developmental stages numbered according to Gosner (1960).
Vertical lines
April with water temperatures ranging from 8 to development at high temperatures, but Cunningham 19*4”C. Brattstrom & Warren (1955) report that & Mullally (1956) “found recently laid eggs in males in southern California do not chorus at streams where the Iowest water temperatures were water temperatures below 9*8’C and that egg Iayin8 above 20°C”. Water temperatures above 20°C seem occurs only at 12-15°C. However, Cunningham & to inhibit chorusing (Brattstrom & Warren, 1955) Mullally (1956) found males calling at 4°C and and, presumably, this may reduce other breeding amplexing pairs at 8-9.7% and developing eggs in activity (e.g. amplexus and egg laying). This be- water of 8*1-14~S’C. Snyder & Jameson (1965) havioral response by adults probably reduces the report males calling at 4@C in rnoun~~s of exposure of developing eggs to phys~olo~~ly southern California. There are no specific data on damaging high temperatures.
100 - . l . l * l . l . . . . 0’ .n 00 * 0 .
D . . D
90 - 0 * e.
l
IO - I
\
i
01
,. *. ~._ . ,,,,,,,,,,,‘,,,, ~:,_;,~
0 2 4 6 8 IO 12 14 I6 16 20 22 24 26 28 30 32
Temperature, “C
Fig. 2. Embryonic temperature tolerance of H. regih eggs: 0, California; 0, WaKington.

866 HERBERT A, BROWN
2. Embryonic temperature toferance
The percentage of the embryos developing normally at each of the experimental temperatures is shown in Fig. 2; the data for California embryos were obtained from twenty females, and the data for Washington embryos from sixteen females. The range of temperatures that permits a high percentage of normal development extends from about 8 to 27°C for Washington embryos, and about 8-29°C for California embryos.
While the data of low temperature tolerance are not complete, it appears that 6°C is the lower limiting temperature for normal development in each population. For the Washington embryos, normal development (90 per cent viability in nine experi- ments) was observed at 7.3”C, but at 4°C cleavage of embryos from four experiments was irregular and all embryos cytolyzed in the early blastuia stages. For the California embryos, viability in five experiments at 5°C ranged between 16 and 74 per cent, at 7*7”C viabiIity ranged between 43 and 77 per cent and at 9°C development was completely normal in all embryos.
The analysis of temperature tolerance is more complete at high temperatures than low temperatures (Fig. 2), and the results show that California embryos have a higher upper limiting temperature and greater intrapopulation variability than Wash- ington embryos. This variation of high temperature tolerance in a single population is unusual, and its reIationship to developmental rates at high tempera- tures will be described in another section of these results.
The upper limiting temperature for normal development of Washington embryos is about 28°C. At 30 and 29°C average viability was only 15 per cent (range, 3-30 per cent). At 27.5%
Table 1. Temperature and embryonic development of H. regiliu from northwestern Washington
Table 2. Temperature and embryonic development of H. regi/h from southern California
7.7
8.8
12.0
I‘.3
15.2
16.5
20.0
24.7
27.0
28.1
29.0
29.6
30.0
30.5
31.0
31.5
2 R18.0 2 0.00 818.0-arf3.0 1.22
5 665.‘ & 19.40 hi?.S-h9*.3 1.50
8 272.5 + 16.15 259.8-303.5 3.67
3 L7R.8) 2.02 176.X-MO.” 5.59
5 158.6 + 2.01 156.5-161.0 6.30
4 129.9 + 0.49 1?9.3-133.8 7.70
17 80.3, 3.“5 75.*- 83.8 12.48
1s 52.91 1.90 48.7- 55.6 18.90
4 46.3 2 cl.!*9 4X.?- 46.7 21.60
9 43.85 O.R9 42.G 44.8 22.83
7 43.1, I.?7 41.s 45.0 25.20
9 43.9 f 1.35 42.h 46.2 22.78
8 44.8 + 0.32 44.3- 45.2 22.32
9 ‘5.4 * “.2? 44.5 47.0 22.02
2 44.8 2 (1.00 46.5 45.0 22.32
2 44.5 + 0.00 44.5 44.5 22.47
viability was 62 per cent (range, 23-97 per cent). At 26.3°C per cent normal development was always high (average, 82 per cent).
In the California population there was si~i~~t variation in developments response to high tempera- tures, but despite this variation the results suggest that 29.6% is a good estimate of the upper limiting temperature for normal development. At 29°C normal development ranged between 80 and 100 per cent in all experiments. However, over a wide temperature interval (29.6, 30, 30.5 and 31.5”C) viability ranged from zero to 100 per cent. These last results are unusual since amphibian embryos generally show that “the change from lethal to non- lethal [temperatures] takes place rather abruptly” (Zweifel, 1968). Finally, at higher temperatures of 32-4 and 32*6”C development was always abnormal, never showing development beyond early cleavage stages.
3. Rate of embryonic development
Developmental rates of H. regilla embryos were determined at constant temperatures between 7.3 and 31+‘C (Tables 1 and 2), and a plot of the rate- temperature curve (Fig. 3) for each population is typical of amphibian development (~chrna~, 1969). The rate is a linear function of temperature in the range between 13 and 25°C for Washington embryos and between 15 and 28°C for California embryos. Outside these temperature intervals developmental rate deviates from a straight line.

Temperature and development of treefrog 867
25 1 I I / I
25- -
.20 -
Fig. 3. Embryonic developmental rate of H. regilla eggs from California and Washington.
This deviation is most conspicuous at high tempera- tures (28-32°C). California embryos show a marked inflexion of the rate curve beginning at 29°C whore the most rapid rate occurs; at higher temperatures there is a progressive decrease in developmental rate with the values at 296-315°C similar to those at 27 or 28°C. While the upper limiting temperature for this California population is 29*6”C, the rate values at higher temperatures were taken from samples where normal development ranged between 6 and 100 per cent. The Washington rate curve appears temperature independent over the range between 27 and 30°C. However, the upper limiting temperature for this population is 28”C, and the rate values at 29 and 30°C represent samples where normal development was below 50 per cent. Thus, some California embryos may show resistance to damage by high temperatures, but these high temperatures will inhibit developmental rate. In comparison, Washington embryos at high tempera- tures show thermal damage and inhibition of developmental rate.
The developmental rate of embryos from the two populations is graphically compared in Fig. 4 by using the procedure of Moore (1949) where the difference in developmental rate appears as the per cent acceleration or per cent retardation with the Washington population serving as the horizontal base line. Thus, Washington embryos have a more rapid developmental rate at low temperatures, and California embryos a more rapid developmental rate at high temperatures. At the lowest temperature of 7*3’C development of Washington embryos from first cleavage (stae 3) to gill circulation (stage 20)
Fig. 4. Comparison of developmental rate between Washington (horizontal line) and California (diagonal
line) H. regilla.
requires 789 hr (32.9 days). The most rapid develop- ment took place at 29°C where California embryos complete development in 43.1 hr (1.8 days). At 19°C both populations have the same developmental rate which requires about 89 hr (3.7 days).
Embryonic development of California H. regilla was observed for times required to reach eighteen specific developmental stages at several constant temperatures (Table 3); the age in hours accompany- ing a given stage, numbered according to Gosner (1960), refers to time elapsed since first cleavage (stage 3). From these data the relative amount of time to complete four major phases of embryonic development was calculated: the period of cleavage (stages 3-10) was 19 per cent; gastrulation (stages 10-f3), 17 per cent; neurulation (stages 13-17), 21 per cent; and organogenesis (stages 17-20), 43 per cent. These values are similar to those calculated for embryonic development of the tailed frog, Ascuphus truei (Brown, 1975).
4. Development of embryos at high temperatures
The development of embryos at critical tempera- tures near the upper limiting temperature may cause the retardation or complete arrest of development (Kiie & Kinne, 1962; Zweifel, 1968; Bachmann, 1969). Thus, Zweifel (1968), referring to amphibian embryos, cautioned that “the time to gill circulation may actually increase slightly at the highest tempera- tures tolerated, which does not necessarily mean that the development of the embryo as a whole has been retarded”. Although the present study did not attempt to analyze the phenomenon of develop- mental arrest or retardation at high temperatures in detail, it seems worthwhile to describe more care- fully the relationship between developmental rate
868 HERBERT A. BROWN
Table 3. Elapsed time (hr) to reach specific develop- mental stages in H. rzgilla from southern California
Two-cell (3) * 0 0 0 0 0
Four-cell (4) 7. 1 1 1 0.5
Light-cell (5) 4 3 2 1.5 1
16-cell (6) 5 4 3 2 1.3
32-.x11 (7) 9 7 6 3 2
Mid-cleavage (a) 12 10 9 6 4
Late-cleavage (9) 23 20 11 9 6
Dorsal lip (IO) 42 33 15 11 a
Nid-gastrula (11) 49 41 21 14 11
Late patlula (12) 56 La 26 16 13
Neural pl.¶Ce (13) 80 61 31 20 16
Neural folds (14) 96 66 35 24 18
Rooeation (15) 118 12 37 25 19
Neural tube (16) 131 80 40 27 21
Tsil-bud (17) 145 88 46 32 26
Muscular response (18) 179 116 52 38 32
Heart boar (19) 218 140 70 51 43
Gill circ"lation (20) 273 157 80 53 45
*Numbers refer to developmental stages of Gosner (1960).
and viability at high temperatures in the eggs of H. regillu from southern California.
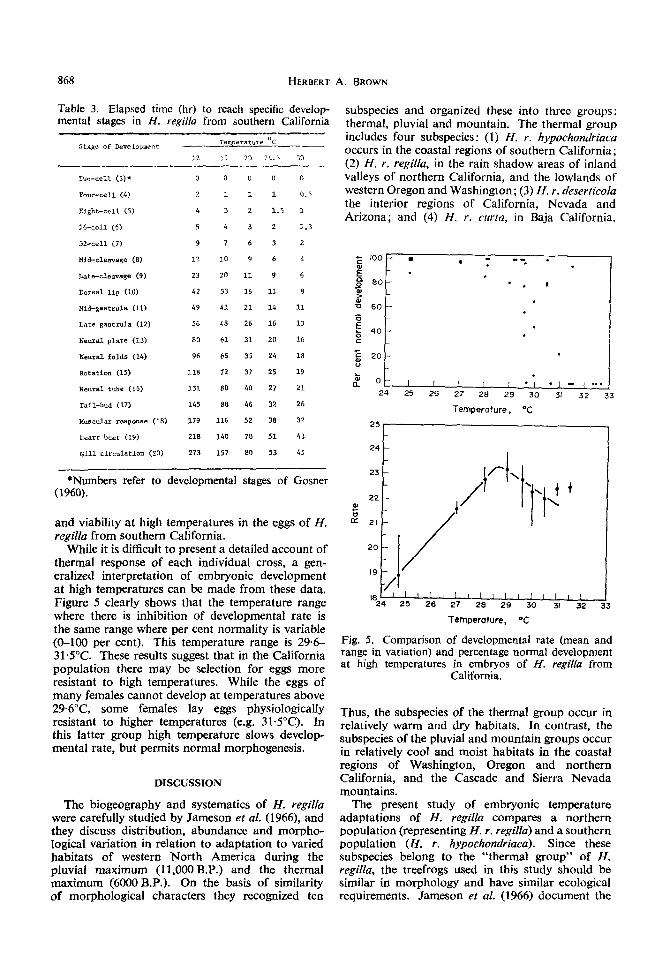
While it is difficult to present a detailed account of thermal response of each individual cross, a gen- eralized interpretation of embryonic development at high temperatures can be made from these data. Figure 5 clearly shows that the temperature range where there is inhibition of developmental rate is the same range where per cent normality is variable (O-100 per cent). This temperature range is 29.6- 31.5”C. These results suggest that in the California population there may be selection for eggs more resistant to high temperatures. While the eggs of many females cannot develop at temperatures above 29.6”C, some females lay eggs physiologically resistant to higher temperatures (e.g. 31.5”C). In this latter group high temperature slows develop- mental rate, but permits normal morphogenesis.
DISCUSSION
The biogeography and systematics of H. regilla were carefully studied by Jameson et al. (1966), and they discuss distribution, abundance and morpho- logical variation in relation to adaptation to varied habitats of western North America during the pluvial maximum (11,000 B.P.) and the thermal maximum (6000 B.P.). On the basis of similarity of morphological characters they recognized ten
subspecies and organized these into three groups: thermal, pluvial and mountain. The thermal group includes four subspecies: (1) H. r. hypochondriaca occurs in the coastal regions of southern California; (2) H. r. regilla, in the rain shadow areas of inland valleys of northern California, and the lowlands of western Oregon and Washington; (3) H. r. deserticola the interior regions of California, Nevada and Arizona; and (4) H. Y. curra, in Baja California.
E 100” .
E I : --. - . .
g 60- .
. . I
. .
. .
.
;i; . a. O I I 1 I . . _ . . .
24 25 26 27 28 29 30 31 32 33
Temperature, “C
24
t I i
Temperature, OC
Fig. 5. Comparison of developmental rate (mean and range in variation) and percentage normal development at high temperatures in embryos of H. regi[la from
California.
Thus, the subspecies of the thermal group occur in relatively warm and dry habitats. In contrast, the subspecies of the pluvial and mountain groups occur in relatively cool and moist habitats in the coastal regions of Washington, Oregon and northern California, and the Cascade and Sierra Nevada mountains.
The present study of embryonic temperature adaptations of H. regilla compares a northern population (representing H. r. regilla) and a southern population (H. r. hypochondriaca). Since these subspecies belong to the “thermal group” of H. regilla, the treefrogs used in this study should be similar in morphology and have similar ecological requirements. Jameson et al. (1966) document the

Temperature and development of treefrog 869
morphological similarity, and the present study shows that these populations have a similar breeding season and embryonic thermal requirements. Both populations breed in the late winter and spring months in relatively cool water. The temperature tolerance range is only slightly different: northern embryos, 6-28”C, and southern embryos, 6-29.6%. Comparison of developmental rates shows that northern embryos have faster rates at low tempera- tures and southern embryos faster rates at high temperatures. The direction of these differences suggest that northern embryos are cold adapted and southern embryos warm adapted. It is possible that more extreme differences in embryonic temperature adaptations will be found when montane subspecies (H. r. cascadae and H. r. sierrae) and desert sub- species (H. r. deserticola and H. r. curta) are studied. Compared to the results of the present study, montane populations may possess even lower limiting temperatures and faster developmental rates at low temperature, and desert populations higher limiting temperatures and faster develop- mental rates at high temperature. Perhaps the small differences in temperature adaptation between the northern and southern embryos of the present study are a consequence of gene flow from surrounding populations of different temperature adaptation. Thus, in southern California embryos (H. r. hypochondriaca) the faster developmental rate at high temperatures and higher upper limiting temperature may be due to gene flow from warm adapted populations (H. r. curta and H. r. deserticola) of nearby desert areas. Likewise, the faster developmental rate at low temperatures of western Washington embryos (H. r. regilla) may be due to gene flow from cold adapted montane popu- lations (H. r. cascadae).
The results of temperature tests on H. regilla are more meaningful when compared to temperature data on embryos of other anuran amphibians. Some species are highly specialized and possess very narrow limits of temperature tolerance. Ascaphus truei and Rana aurora inhabit the same region as H. regilla in the Pacific Northwest, and the last two
species may breed in the same pond. The lower and upper limiting temperatures of A. truei is 5 and 185°C (Brown, 1975) and R. aurora, 4 and 21°C (Licht, 1971). Bufo valliceps from southern Texas has the highest values of lower and upper limiting temperatures, 18 and 35°C (Volpe, 1957). The limiting temperatures of H. regilla, 6 and 29_6”C, suggest that this treefrog is not specialized for cold nor warm temperatures. Compared to other treefrog species (Table 4), H. regilla has a broad range of temperature tolerance, and this characteristic correlates with its abundance and its wide ranging geographic and ecologic distribution.
The thermal tolerance limits of Washington H. regilla embryos, 6-28”C, are very broad compared to the water temperatures in the pools during the early part of the breeding season. The upper limiting temperature (28°C) is much higher than the mean maximum temperature (13.7%) of the water. However, the lower limiting temperature (6°C) is close to the mean minimum temperature (4.9”C). In general, these very low water temperatures are not lethal unless they are maintained for many days. Schechtman & Olson (1941) placed eggs of H. regilla (from southern California) in a refrigerator at - 5 to - 7°C and found that the eggs frozen in ice overnight could continue to develop normally when thawed at room temperature (2124°C). Also, they found that blastulae, gastrulae, neurulae and tailbud stages stored for 8 days at 4-5°C can develop normally when brought to room tempera- ture. The high upper limiting temperature of 28°C may show a better relationship to water tempera- ture during the later part of the breeding season (May and June). Volpe (1957) found no difference in response to experimental temperatures of B. valliceps embryos derived from early breeding (April-early June) and late breeding (late June- August) toads. Thus, the broad range of tempera- ture tolerance in H. regilla may correlate with the maximum and minimum water temperatures for the entire breeding season (February-June).
This study of development of California embryos at high temperatures provides unusual results: (1)
Table 4. Representative data of embryonic temperature tolerance of North American treefrogs (Hylidae)
specie*
Ella regilla (Washington)
H. reailla (California)
H. arenicalor (Arirona)
H. cinerea (Texas)
Peeudscris streckeri (Texas)
P. clerk1 (Texas)
ACCTI. crellitanns (Texas)
Limiting Temp ‘C
LOWar upper Reference_
6 26 Present study
6 29.6 Present study
413 s30.5 Zveffel (1968)
%20 34-3s Ballingar 6 WcKinney (1966)
g(1) 27 "ubbs et al. (1963) --
5.543 35-39 gallinger 6 HcKimeY (1966)
1s 34 Ballinga 6 McKimey (1966)

870 HERBERT A. BROWN
the most rapid developmental rate occurs at 29°C; (2) there is inhibition of developmental rate at temperatures above 29°C; (3) the upper limiting temperature for normal development of most embryos is 29~6°C; but (4) some embryos show normal development at high temperatures (29.6- 31*5X), and their developmental rate is inhibited. These data contrast to the conclusions of Bachmann (1969): “Developmenta rate [in amphibiansJ is a linear function of temperature throughout the range of lOO”/, survival”, and temperatures causing inhibition of developmental rate usually induce severely abnormal embryos.
Two previous studies on the relationship between temperature and embryonic development of anuran eggs provide results relevant to the present discus- sion. Zweifel (1968) found significant inhibitjon of developmental rate of Bufu punctatus embryos at high temperatures: at 33.8”C embryonic develop- ment required 21 hr but at 34.3”C it required 26.5 hr. Dasgupta & Grewal (1968, 1970) studied embryonic temperature tolerance and developmental morpho- Iogy of Diana c~u~~~~y~~c~~~ from northern India. This population has lower and upper limiting temperatures of 22 and 31*5X, respectively, but a population from southern India has a lower limiting temperature of 17-18’C. The northern population (presumably warm adapted) is unusual because it shows genetic polymorphism regarding development at high temperatures. Frogs with vertebral fusion (due to a single dominant gene) comprise about 10 per cent of the natural population, and these frogs can tolerate higher temperatures during embryonic development than frogs with normal vertebral morphology. These examples support the data on embryonic development of H. regilra and iIiustrate the possible variability when eggs develop at high temperatures near the upper limiting temperature. The finding of inhibition of developmental rate should be expected, but it is seldom found in developmental studies of amphibian eggs. The polymorphic heat resistance shown by Dasgupta & Grewal (1968) and the present study are exceptional, although they might be expected in populations where environmental selection favors mutants with increased heat resistance.
The thermal tolerance and developmental rate of California embryos deserve more thorough study, but these preliminary results suggest that there may be selection for increased heat resistance of embryos in this population. This development probably involves evolutionary adaptation of enzymes to temperature. Somero (1969) found that zones of thermal insensitivity in the metabolism of aquatic poikilotherms are in part due to changes in enzyme- substrate affinity over a range of temperatures. Somero concluded that sharp decreases in enzyme- substrate affmity which frequently occur at the extremes of an organism’s habitat temperature may be important in establishing thermal tolerance
limits for the organism. The work of Lovtrup (1961) has been concerned with embryological develop- ment and enzyme synthesis at different tem~ratures in eggs of Rana plaryrrhina, and studies of this type are required for an understanding of the mechanism of evolutionary temperature adaptation in the embryos of H. regilla.
The physiological and ecological adaptations of amphibian embryos to temperature have been reviewed in two recent and comprehensive studies. Bachmann (1969) compared the developmental rate of twenty anuran species representing six families, but this includes only one treefrog (Hyla Diridis from Italy). Zweifel (1968) studied temperature adaptations in eight anuran species representing four families; data on temperature tolerance and deveIop- mental rate of one treefrog (H. uren~c~~o~) are presented. In the United States treefrogs of the family Hylidae are represented by about twenty-nine species belonging to five genera but, despite this
250
I Hyla regilla (Cotif.) 6-29.6’C
2 Hylo arenicolor 13-31°C
200 3 Bufo cognatus 16-33.5”C
4 Scophiopus couchi 155-34*C
5 Rana pipiens 12-3l5 “C
150 z
Ii i=
100
20 25 30
Temperature, ‘C
Fig. 6. A comparison of the rate of embryonic develop- ment between warm adapted species of North American anurans. The curves represent the time between stages 3 and 20 at several temperatures. Data for H. regilra (present study); X. arenicolor, B. cognatus, S. couchi and
R. pipiens (Zweifel, 1968).
abundance of species, there is almost a total absence of information on developmental rate of treefrog embryos, and it is not possible to make comparisons within this family.
Rates of embryonic development in representatives of five families of North American anuran
Temperature and development of treefrog 871
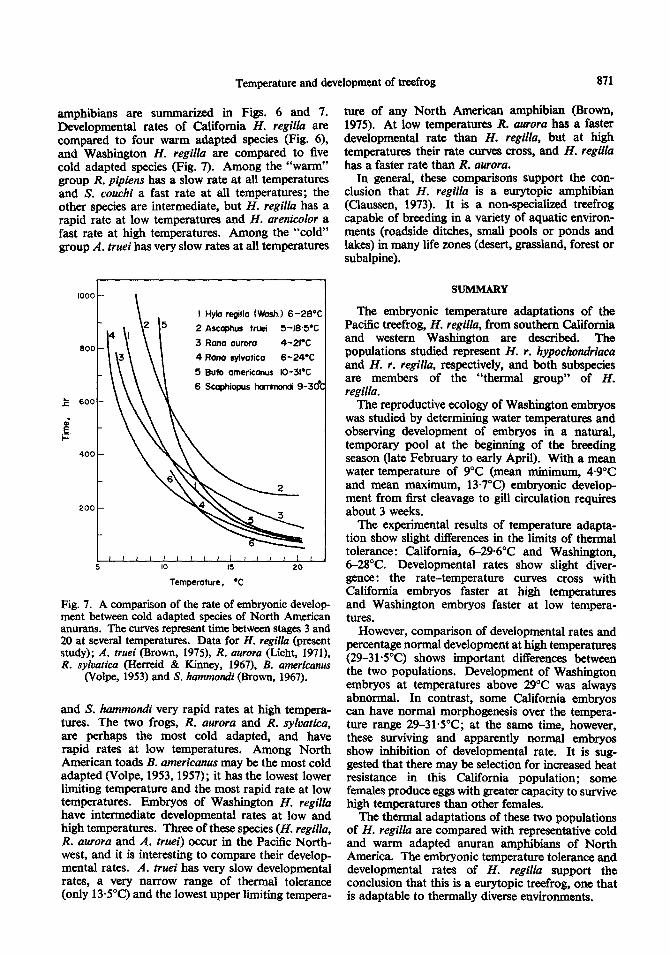
amphibians are summarized in Figs. 6 and 7. Developmental rates of California H. regilla are compared to four warm adapted species (Fig. 6), and Washiion H. regilla are compared to five cold adapted species (Fig. 7). Among the “warm” group R. pipiens has a slow rate at all temperatures and S. couchi a fast rate at all temperatures; the other species are intermediate, but H. regilla has a rapid rate at low temperatures and H. arenicolor a fast rate at high temperatures, Among the “cold” group A. truei has very slow rates at all temperatures
I Hyb regilla Wash.) 6-26’C
Temperature, ‘C
Fig. 7. A comparison of the rate of embryonic develop- ment between cold adapted species of North American anurans. The curves represent tune between stages 3 and 20 at several temperatures. Data for H. regillu (present study); A. truei (Brown, 1975), R. aurora (Licht, 1971), R. sylvatica (Herreid & Kinney, 1967), B. americanus
(Volpe, 1953) and S. hammondi (Brown, 1967).
and S. hammondi very rapid rates at high tempera- tures. The two frogs, R. aurora and R. sylvatica, are perhaps the most cold adapted, and have rapid rates at low temperatures. Among North American toads B. americanus may be the most cold adapted (Volpe, 1953,1957); it has the lowest lower limiting temperature and the most rapid rate at low temperatures. Embryos of Washington H. regilla have intermediate developmental rates at low and high temperatures. Three of these species (H. regilla, R. aurora and A. truei) occur in the Pacific North- west, and it is interesting to compare their develop- mental rates. A. truei has very slow developmental rates, a very narrow range of thermal tolerance (only 135°C) and the lowest upper limiting tempera-
ture of any North American amphibian (Brown, 1975). At low temperatures R. aurora has a faster developmental rate than H. regilla, but at high temperatures their rate curves cross, and H. regild has a faster rate than R. aurora.
In general, these comparisons support the con- clusion that H. regilla is a eurytopic amphibian (Claussen, 1973). It is a non-specialized treefrog capable of breeding in a variety of aquatic environ- ments (roadside ditches, small pools or ponds and lakes) in many life zones (desert, grassland, forest or subalpine).
SUMMARY
The embryonic temperature adaptations of the Pacific treefrog, H. regilla, from southern Caliiomia and western Washington are described. The populations studied represent H. r. hypochondriaca and H. r. regilla, respectively, and both subspecies are members of the “thermal group” of H. regilla.
The reproductive ecology of Washington embryos was studied by determining water temperatures and observing development of embryos in a natural, temporary pool at the beginning of the breeding season (late February to early April). With a mean water temperature of 9°C (mean miniium, 4.9% and mean maximum, 13*7”C) embryonic develop- ment from first cleavage to gill circulation requires about 3 weeks.
The experimental results of temperature adapta- tion show slight differences in the limits of thermal tolerance: California, 6-29*6”C and Washington, 6-28°C. Developmental rates show slight diver- gence: the rate-temperature curves cross with California embryos faster at high temperatures and Washington embryos faster at low tempera- tures.
However, comparison of developmental rates and percentage normal development at high temperatures (29-31*5”C) shows important differences between the two populations. Development of Washington embryos at temperatures above 29°C was always abnormal. In contrast, some California embryos can have normal morphogenesis over the tempera- ture range 29-31*5”C; at the same time, however, these surviving and apparently normal embryos show inhibition of developmental rate. It is sug- gested that there may be selection for increased heat resistance in this California population; some females produce eggs with greater capacity to survive high temperatures than other females.
The thermal adaptations of these two populations of H. regilla are compared with representative cold and warm adapted anuran amphibians of North America. The embryonic temperature tolerance and developmental rates of H. regild support the conclusion that this is a eurytopic treefrog, one that is adaptable to thermally diverse environments.

872 HERBERT A. BROWN
Acknowledgements-I wish to thank Dr. Rodolfo Ruibal and Dr. Mack Dugger of the Department of Life Sciences, University of California, Riverside, for provid- ing me with the opportunity to study embryos of the Pacific treefrog in southern California.
REFERENCES
BACHMANN K. (1969) Temperature adaptations of amphibian embryos. Am. Nat. 103, 115-130.
BALL R. W. & JAME~~N D. L. (1966) Premating isolating mechanisms in sympatric and allopatric HyIu regilla and HyIa caIiforniae. Evolution 20, 533-551.
BALLINGER R. E. & MCKINNEY C. 0. (1966) Develop- mental temperature tolerance of certain anuran species. J. exp. Zool. 161, 21-28.
BRATISTROM B. H. (1968) Thermal acclimation in anuran amnhibians as a function of latitude and altitude. Camp. Biochem. Physiol. 24, 93-l 11.
BRATTSTROM B. H. & WARREN J. W. (1955) Observations on the ecology and behavior of the Pacific treefrog, HyIa regilla. Copeia 1955, 18 1-191.
BROWN H. A. (1967) Embryonic temperature adaptations and genetic compatibility in two allopatric populations of the spadefoot toad, Scaphiopus hammondi. Evolution 21, 742-761.
BROWN H. A. (1975) Temperature and development of the tailed frog, Ascaphus truei. Comp. Biochem. Physiol. SOA, 397-405.
CALHOUN R. E. & JAME~~N D. L. (1970) Canonical correlation between variation in weather and variation in size in the Pacific tree frog, HyIa regilla, in southern California. Copeia 1970, 124-134.
CLAU~~EN D. L. (1973) The thermal relations of the tailed frog, Ascaphus truei, and the Pacific treefrog, HyIa regiIIa. Comp. Biochem. Physiol. 44A, 137-153.
CUNNINGHAM J. D. & MULLALLY D. P. (1956) Thermal factors in the ecology of the Pacific treefrog. Herpeto- Iogica 12, 68-79.
DASGUPTA S. & GREWAL M. (1968) The selective advantage of temperature tolerance among the progeny of frogs with vertebral fusions. Evolution 22, 87-93
DA~GUPTA S. & GREWAL M. (1970) Inheritance of vertebral fusion in the skipper frog. J. Hered. 61, 174-176.
DENT J. N. (1968) Survey of amphibian metamorphosis. In Metamorphosis (Edited by ETKIN W. & G~BERT L.), pp. 271311. Appleton-Century-Crofts, New York.
GOSNER K. L. (1960) A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16, 183-190.
HERREID C. F. & KINNEY S. (1967) Temperature and development of the wood frog, Rana syhatica, in Alaska. Ecology 48, 579-590.
HUEIBS C., WRIGHT T. & CUELLAR 0. (1963) Develop- mental temperature tolerance of central Texas popula- tions of two anuran amphibians, Bufo valliceps and Pseudacris streckeri. Southwest. Nat. 8, 142-149.
JAME~ON D. L., MACKEY J. P. & RICHMOND R. C. (1966) The systematics of the Pacific tree frog, HyIa regiIIa. Proc. Calif. Acad. Sci. 33, 551-620.
JAME~~N D. L. & PEQUEGNAT S. (1971) Estimation of relative viability and fecundity of color polymorphisms in anurans. Evolution 25, 180-194.
JAMESON D. L., TAYLOR W. & MOUNTJOY J. (1970) Metabolic and morphological adaptation to hetero- geneous environments by the Pacific tree toad, Hy/a regilla. Evolution 24, 75-89.
KINNE 0. & KINNE E. M. (1962) Rates of development in embryos of a cyprinodont fish exposed to different temperature-salinity-oxygen combinations. Can. J. ZooI. 40, 231-253.
LICHT L. E. (1971) Breeding habits and embryonic thermal requirements of the frogs, Runa aurora aurora and Rana pretiosa pretiosa, in the Pacific Northwest. Ecology 52, 116-l 24.
LOVTRUP S. (1961) Morphological development and chemical differentiation during amphibian embryo- genesis at low temperatures. J. exp. ZooI. 147, 227- 232.
MOORE J. A. (1949) Geographical variation of adaptive characters in Rana pipiens Schreber. Evolution 3, l-24.
PETNS D. & ANGLETON G. M. (1967) Comparative reproductive biology of montane and Piedmont chorus frogs. Evolution 21, 500-507.
RE~NICK L. E. & JAME~~N D. L. (1963) Color poly- morphism in Pacific tree frogs. Science, Wash. 142, 1081-1083.
RUGH R. (1962) Experimental EmbryoIogy, 3rd Edn. Burgess, Minneapolis, Wisconsin.
SALTHE S. N. & DUELLMAN W. E. (1973) Quantitative constraints associated with reproductive mode in anurans. In Evolutionary Biology of the Anurans (Edited by VIAL J.), pp. 229-249. University of Missouri Press, Columbia.
SALTHE S. N. & MECHAM J. S. (1974) Reproductive and courtship patterns. In PhysioIogy of the Amphibia (Edited by LOFTS B.), Vol. 2, pp. 309-521. Academic Press, New York.
SCHECHTMAN A. M. Jc OLSON J. B. (1941) Unusual temperature tolerance of an amphibian egg (HyIu regiIIa). EcoIogy 22,409-410.
SNYDER W. F. & JAME~~N D. L. (1965) Multivariate geographic variation of mating call in populations of the Pacific tree frog (HyIa regilla). Copeia 1%5, 128-141.
SOMERO G. N. (1969) Enzymic mechanisms of tempera- ture compensation: immediate and evolutionary effects of temperature on enzymes of aquatic poikilo- therms. Am. Nat. 103, 517-530.
STEBBINS R. C. (1966) A FieId Guide to the Western ReptiIes and Amphibians. Houghton-Mifflin, Boston, Mass.
STORER T. I. (1925) A synopsis of the amphibia of California. Univ. Calif. PubI. ZooI. 27, l-342.
VOGT T. & JAMEX~N D. -L. (1970) Chronological correla- JAMESON D. L. (1966) Rate of weight loss of tree frogs at tion between change in weather and change in morph-
various temDeratures and humidities. Ecoloav 47. ology of the Pacific tree frog in southern California. 605-613. =
__ I Copeia 1970, 135-144.
JAME~ON D. L., MACKEY J. P. & ANDERSON M. (1973) VOLPE E.P . (1953) Embryonic temperature adaptations Weather, climate, and the external morphology of and relationships in toads. Physiol. Zoiil. 26, 344- Pacific tree toads. Evolution 27,285-302. 354.

Temperature and development of treefrog 873
VOLPE E. P. (1957) Embryonic temperature tolerance ZWEIFEL R. G. (1968) Reproductive biology of anurans and rate of development in Bz& valliceps. Physiol. of the arid southwest, with emphasis on adaptation ZoH. 3@,164-176. of embryos to ~rn~~tu~. Bull. Am. Mur. Nat. Hist.
VOLPE E. P., W~KI!NS M. A. & DOBIE J. L. (1961) 140,144.
Embryonic and larval development of Hyla nvivocu. Key Word Index-Embryonic temperature adaptations; Copeia 1961, 340-349. limiting temperatures; developmental rate; amphibians;
anurans : Hyia regilla.





























