Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, July 1973, p. 316-322Copyright 0 1973 American Society for Microbiology
Vol. 115, No. 1Printed in U.SA.
Electron Micrograph Study of the Asci andAscospores of Metschnikowia Kamienski
L. T. TALENS,1 M. W. MILLER, AND MARY MIRANDADepartment of Food Science and Technology, University of California, Davis, California 95616
Received for publication 2 April 1973
Internal and surface structures of asci and ascospores were studied bytransmission electron microscopy (TEM) and by scanning electron microscopy(SEM) to establish the character and number of ascospores within the ascus ofMetschnikowia krissii. Enzyme digestion with snail gut enzymes and SEMexamination suggested the presence of a single ascospore enclosed in a thicksheath of epiplasmic materials. Two closely associated ascospores without anepiplasmic sheath were clearly distinguishable from asci of M. bicuspidata var.chathamia when similarly treated. Ultramicrotomy and TEM establishedconclusively that M. krissii produced a single ascospore per ascus. Neither SEMnor TEM revealed any morphological detail of the ascospores of taxonomicsignificance.
Metschnikoff (7) first described and namedMonospora bicuspidata, a yeast-like fungusparasitizing the fresh-water crustaceanDaphnia magna. The organism reproduced bymultilateral budding and formed club-shapedasci containing single, needle-like ascosporeswith both ends pointed. Kamienski (1) observeda similar type of yeast in the brine shrimpArtemia salina and renamed the genusMetschnikowia.
Several species and varieties ofMetschnikowia are presently recognized (6, 8).These species are differentiated on the basis ofphysiology and manner of sexual reproduction,including shape and number of ascospores perascus. Treatment of the ascus with enzymesobtained from snail gut demonstrated the pres-ence of two closely appressed spores in M.bicuspidata var. chathamia when it was ex-amined by light microscopy (8). When the sameenzyme treatment was used on sporulatingsuspensions derived from M. krissii, some ob-servations suggested that this species might alsoproduce two spores per ascus. Additional evi-dence, by means of electron microscopy, shallbe necessary to clarify these observations.
MATERIALS AND METHODSThe culture of M. krissii used in this study (UCD,
FS&T 61-31) was the type strain (CBS 4823) isolatedfrom sea water in La Jolla, Calif. The culture wastransferred to a slant of V-8 juice-agar medium (6)
1Present address: Department of Veterinary Microbiology,School of Veterinary Medicine, University of California,Davis, Calif. 95616.
and incubated at 20 C for 2 weeks. M. bicuspidatavar. chathamia (UCD 67-2) was the type strain (CBS6011) isolated from a fresh-water pond on ChathamIsland, New Zealand. The culture was incubated ondiluted V-8 juice-agar medium (1:20, vol/vol) for 10days at 12 C to obtain ascospores. Light microscopyexaminations of these cultures were made to ascertainat what point the yeast cells were at maximumsporulation.
For scanning electron microscopy, surface growthof a sporulating culture was scraped from the agarslant and suspended in 50 mM phosphate buffer (pH7.0) containing 1- to 2-mM mercaptoethanol. After20 min the cells were washed twice with 20-mM phos-phate buffer (pH 7.0), which was used in all subse-quent washes. The cells were suspended in 0.5 mlof buffer to which was added 0.25 ml of a 1:10 dilu-tion of a preparation from the digestive tract of thegarden snail Helix pomatia. The suspension was in-cubated for 2 h (or until enough ascospores were re-leased) at 37 C, at which time 0.05 ml of ribonuclease(Calbiochem, Los Angeles, Calif; 2 mg/ml) wasadded. Incubation was continued for 2 more h at 37 C,and then 0.05 ml of Pronase (Calbiochem; 2 mg/ml)was added. After 1 h, again at 37 C, the sample waswashed three times in buffer. Subsequent specimenfixation, dehydration, and preparation for scanningelectron microscopy (SEM) examination was by themethods of Talens et al. (9).
Specimen preparation for transmission electronmicroscopy (TEM) also were by the methods ofTalens et al. (9).
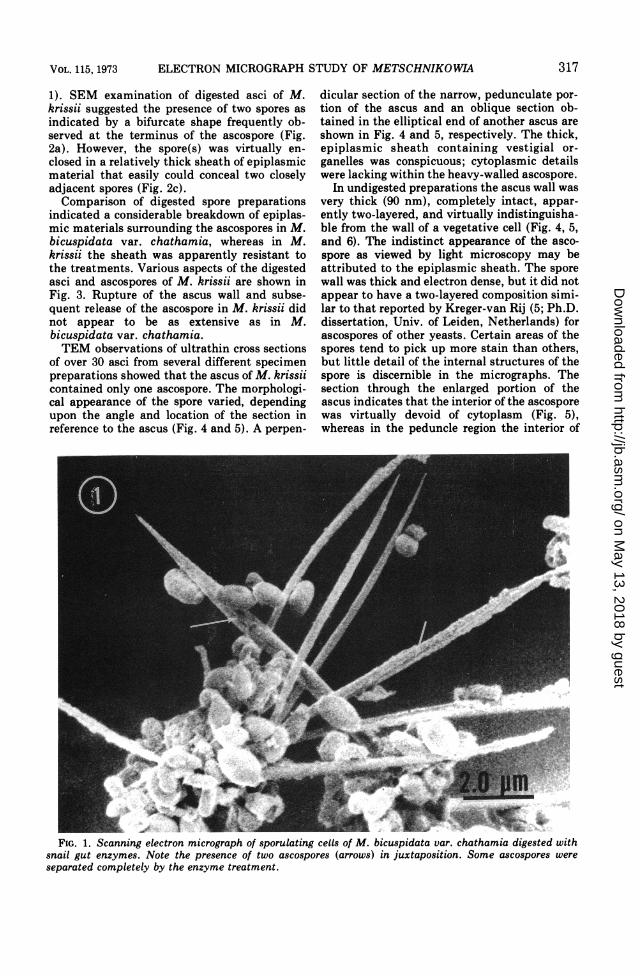
RESULTSDigested asci of M. bicuspidata var.
chathamia showed two, frequently closely ap-pressed, but distinguishable, ascospores (Fig.
316
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
ELECTRON MICROGRAPH STUDY OF METSCHNIKOWIA
1). SEM examination of digested asci of M.krissii suggested the presence of two spores asindicated by a bifurcate shape frequently ob-served at the terminus of the ascospore (Fig.2a). However, the spore(s) was virtually en-closed in a relatively thick sheath of epiplasmicmaterial that easily could conceal two closelyadjacent spores (Fig. 2c).Comparison of digested spore preparations
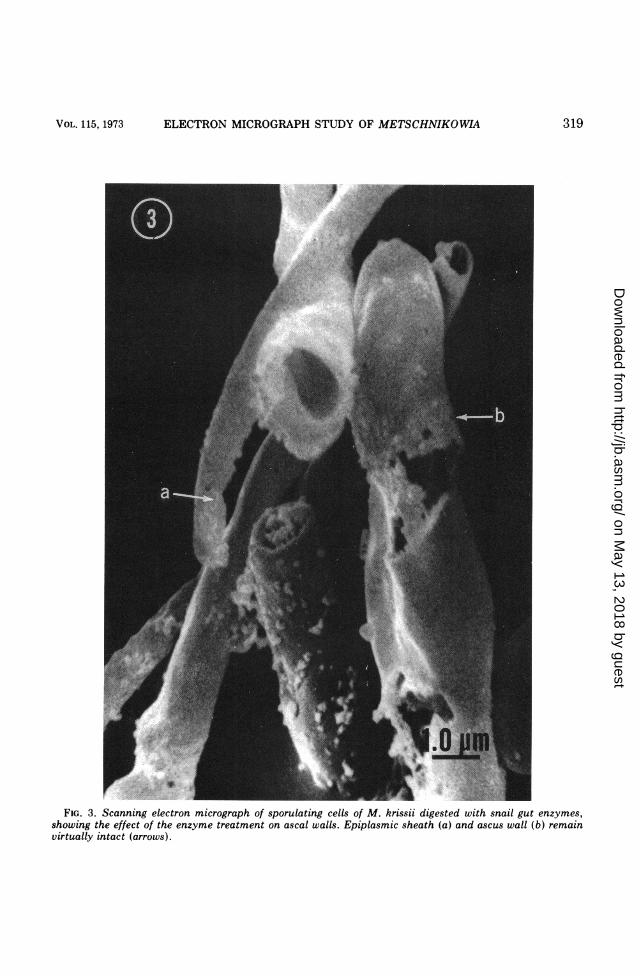
indicated a considerable breakdown of epiplas-mic materials surrounding the ascospores in M.bicuspidata var. chathamia, whereas in M.krissii the sheath was apparently resistant tothe treatments. Various aspects of the digestedasci and ascospores of M. krissii are shown inFig. 3. Rupture of the ascus wall and subse-quent release of the ascospore in M. krissii didnot appear to be as extensive as in M.bicuspidata var. chathamia.TEM observations of ultrathin cross sections
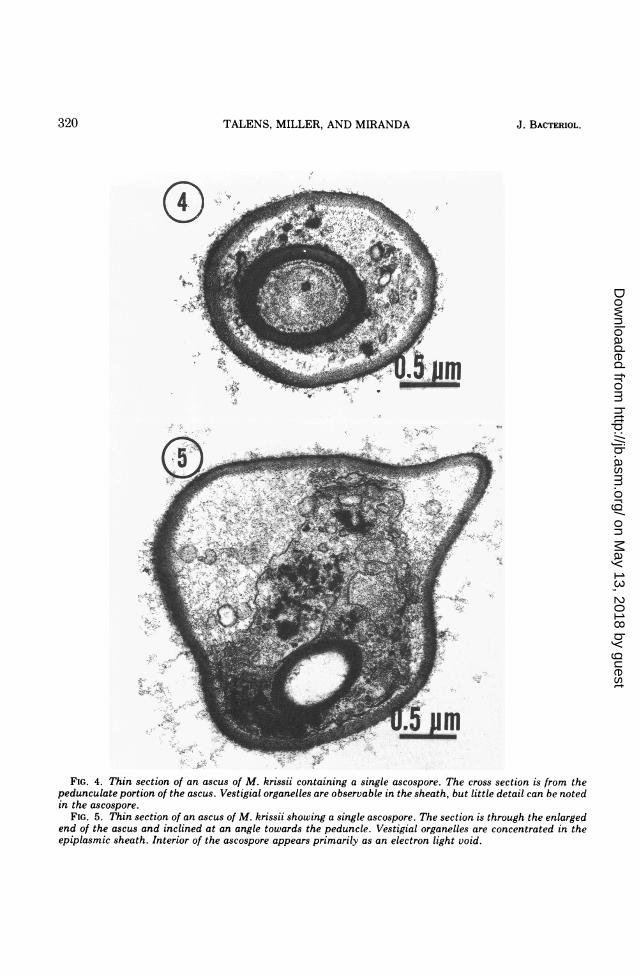
of over 30 asci from several different specimenpreparations showed that the ascus of M. krissiicontained only one ascospore. The morphologi-cal appearance of the spore varied, dependingupon the angle and location of the section inreference to the ascus (Fig. 4 and 5). A perpen-
dicular section of the narrow, pedunculate por-tion of the ascus and an oblique section ob-tained in the elliptical end of another ascus areshown in Fig. 4 and 5, respectively. The thick,epiplasmic sheath containing vestigial or-ganelles was conspicuous; cytoplasmic detailswere lacking within the heavy-walled ascospore.
In undigested preparations the ascus wall wasvery thick (90 nm), completely intact, appar-ently two-layered, and virtually indistinguisha-ble from the wall of a vegetative cell (Fig. 4, 5,and 6). The indistinct appearance of the asco-spore as viewed by light microscopy may beattributed to the epiplasmic sheath. The sporewall was thick and electron dense, but it did notappear to have a two-layered composition simi-lar to that reported by Kreger-van Rij (5; Ph.D.dissertation, Univ. of Leiden, Netherlands) forascospores of other yeasts. Certain areas of thespores tend to pick up more stain than others,but little detail of the internal structures of thespore is discernible in the micrographs. Thesection through the enlarged portion of theascus indicates that the interior of the ascosporewas virtually devoid of cytoplasm (Fig. 5),whereas in the peduncle region the interior of
FIG. 1. Scanning electron micrograph of sporulating cells of M. bicuspidata var. chathamia digested withsnail gut enzymes. Note the presence of two ascospores (arrows) in juxtaposition. Some ascospores wereseparated completely by the enzyme treatment.
317VOL. 115, 1973
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TALENS, MILLER, AND MIRANDA
FIG. 2. Scanning electron micrograph of sporulating cells of M. krissii digested with snail gut enzymes. Note(a) the bifurcate end of the ascospore, (b) remnants of the ascus structure, and (c) the epiplasmic sheathimmediately surrounding the ascospores.
the spore was filled with cytoplasmic material(Fig. 4).
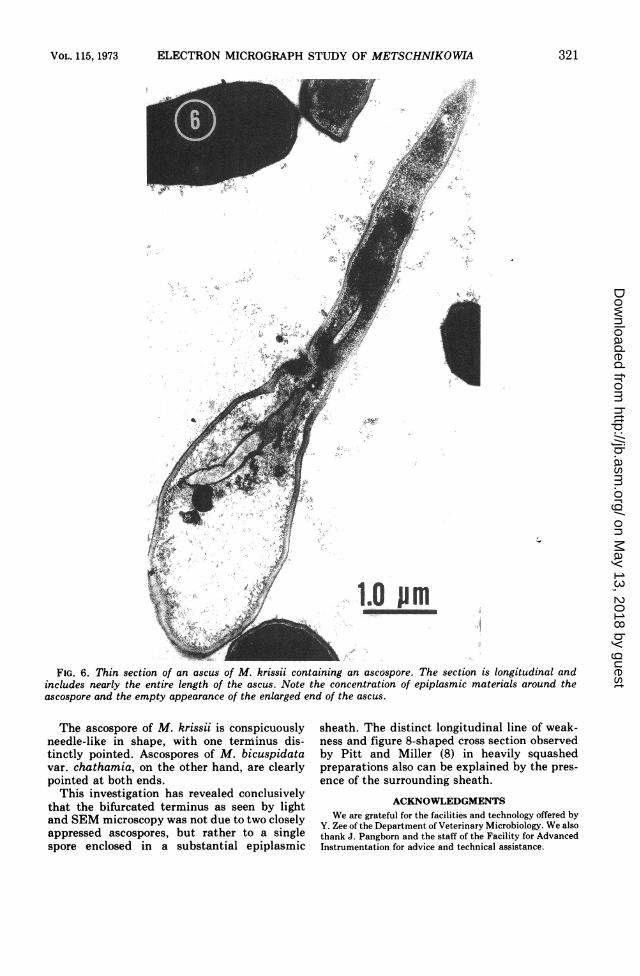
Figure 6 shows a rarely observed, longitudinalsection through the ascus and ascospore. Themajor portion of the enlarged end of the ascusappears to be less electron dense than does thearea surrounding the ascospore, which can beattributed to the epiplasmic sheath. The sporeis somewhat curved and the visible terminusappears to be blunt. The obscured terminusappears to taper gradually which would resultin a single-pointed terminus as is observed bylight microscopy.
DISCUSSIONLittle attention was given to the submicro-
scopic structure of yeast until Kawakami (2)and Kawakami and Nehira (3, 4) publishedelectron micrographs of the various ascosporesof yeast with some reference to their possibletaxonomic usefulness. Although it has beenknown for many years that spores of yeastexhibit considerable variation in shape, surfaceas well as internal structures could reveal de-tails which could lead to further characteriza-tion and conceivably to speciation, and such
structural details would be valuable in studiesof yeast species with similar ascospore shapes asobserved by light microscopy.The investigations by Kreger-van Rij (5;
Ph.D. dissertation, Univ. of Leiden, Nether-lands) have been a significant step in thedirection of the possible use of spore ultrastruc-ture in yeast taxonomy. Particular attentionhas been given to the spore wall, which showsconsiderable variation as compared with theinternal structures, which have disclosed toolittle detail to be useful.The ascospore wall of M. krissii that was
examined by TEM did exhibit varying electrondensity. Whether this represents the presence ofmore than one wall layer, as reported for manyyeast spores by Kreger-van Rij, or whether thepreparatory treatments are responsible for thedifferential electron stain uptake is not certain.No distinguishing surface details can be recog-nized because the spore wall is smooth, presum-ably through its entire length.Demonstration of the internal details of the
ascospore has been unsuccessful. Due to thelength and narrow diameter of the ascospore,the ideal section of the spore through its entirelength is virtually impossible to obtain.
318 J. BACTERIOL.
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
ELECTRON MICROGRAPH STUDY OF METSCHNIKOWIA
FIG. 3. Scanning electron micrograph of sporulating cells of M. krissii digested with snail gut enzymes,showing the effect of the enzyme treatment on ascal walls. Epiplasmic sheath (a) and ascus wall (b) remainvirtually intact (arrows).
319VOL. 115, 1973
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
TALENS, MILLER, AND MIRANDA
o.-
FIG. 4. Thin section of an ascus of M. krissii containing a single ascospore. The cross section is from thepedunculate portion of the ascus. Vestigial organelles are observable in the sheath, but little detail can be notedin the ascospore.
FIG. 5. Thin section of an ascus of M. krissii showing a single ascospore. The section is through the enlargedend of the ascus and inclined at an angle towards the peduncle. Vestigial organelles are concentrated in theepiplasmic sheath. Interior of the ascospore appears primarily as an electron light void.
320 J. BACTERIOL.
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
ELECTRON MICROGRAPH STUDY OF METSCHNIKOWIA
'V.
sl j | ! .s.S-8 S j~~~~.1r'>
S;t L l.Ollm~~~~~~~~~-
FIG. 6. Thin section of an ascus of M. krissii containing an ascospore. The section is longitudinal andincludes nearly the entire length of the ascus. Note the concentration of epiplasmic materials around theascospore and the empty appearance of the enlarged end of the ascus.
The ascospore of M. krissii is conspicuouslyneedle-like in shape, with one terminus dis-tinctly pointed. Ascospores of M. bicuspidatavar. chathamia, on the other hand, are clearlypointed at both ends.
This investigation has revealed conclusivelythat the bifurcated terminus as seen by lightand SEM microscopy was not due to two closelyappressed ascospores, but rather to a singlespore enclosed in a substantial epiplasmic
sheath. The distinct longitudinal line of weak-ness and figure 8-shaped cross section observedby Pitt and Miller (8) in heavily squashedpreparations also can be explained by the pres-ence of the surrounding sheath.
ACKNOWLEDGMENTSWe are grateful for the facilities and technology offered by
Y. Zee of the Department of Veterinary Microbiology. We alsothank J. Pangborn and the staff of the Facility for AdvancedInstrumentation for advice and technical assistance.
321VOL. 115, 1973
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TALENS, MILLER, AND MIRANDA
LITERATURE CITED
1. Kamienski, T. 1889. Notice preliminaire sur la nouvelleespece de Metschnikowia (Monospora Metschn.). Trav.Soc. Imp. Natural. (St. Petersburg) 30:363-364.
2. Kawakami, N. 1958. Electron microscopy of fungi. V. Themorphological study of the spore of non-pellicle group inthe genus Debaryomyces and the classification of thegenus Debaryomyces. J. Electronmicrosc. 7:35-38.
3. Kawakami, N., and T. Nehira. 1958. Electron-microscopyof fungi. IV. The morphological study of the spore ofpellicle group in the genus Debaryomyces. J. Electron-microsc. 7:33-34.
4. Kawakami, N., and T. Nehira. 1959. Electron-microscopyof fungi. IX. Intracellular structures of Rhodotorulaglutinis and Nadsonia fulvescens and their relation tothe physiological characters and taxonomic affinity. J.
Ferment. Technol. 37:125-132.5. Kreger-van Rij, N. J. W. 1966. Some features of yeast
ascospores observed under the electron microscope. InYeasts, Proc. Sec. Symp. Yeasts. Vydavatel'stvo Sloven-skej Akademe Vied, Bratislava, 1969.
6. Lodder, J. 1970. The yeasts, a taxonomic study. North-Holland Publishing Co., Amsterdam.
7. Metschnikoff, E. 1884. Uber eine Sprosspilzkrankeit derDaphnien. Beitrag zur Lehre uber den Kampf derPhagocyten gegen Krankheitserreger. Arch. Pathol.Anat. Physiol. Klin. Med. Virshows 96:177-195.
8. Pitt, J. I., and M. W. Miller. 1970. Speciation in the yeastgenus Metschnikowia. Antonie van Leeuwenhoek J.Microbiol. Serol. 36:357-381.
9. Talens, L. T., M. Miranda, and M. W. Miller. 1973.Electron micrographic study of bud formation in Met-schnikowia krissii. J. Bacteriol. 114:413-423.
322 J. BACTERIOL.
on May 13, 2018 by guest
http://jb.asm.org/
Dow
nloaded from













![Preparation Physicochemical Characterization and Catalytic ... · modified polymeric Catalysts [18] (Table 2). 3.3. High Resolution Scanning Electron Micrograph SEM at various stages](https://img.pdfslide.us/doc/110x75/5f1fdc0fc7f36e47270b0f24/preparation-physicochemical-characterization-and-catalytic-modified-polymeric.jpg)















