Embed Size (px)
Citation preview

Vol. 35, No. 1INFECTION AND IMMUNITY, Jan. 1982, p. 256-2630019-9567/82/010256-08$02.00/0
Effects of Estradiol and Progesterone on Bacteroidesmelaninogenicus and Bacteroides gingivalis
KENNETH S. KORNMANt* AND WALTER J. LOESCHEDepartments ofMicrobiology and Oral Biology, University of Michigan, Ann Arbor, Michigan 48109
Received 17 April 1981/Accepted 27 August 1981
Bacteroides melaninogenicus subsp. intermedius increases in the subgingivalmicroflora during pregnancy. These studies evaluated direct interactions betweenhormonal steroids and oral Bacteroides species. Resting cell suspensions of purecultures of plaque organisms were incubated anaerobically with [ 4C]estradiol and[14C]progesterone. Uptake of labeled compound per microgram of bacterialprotein was determined by thin-layer chromatography and liquid scintillationcounting. B. melaninogenicus subsp. intermedius and B. melaninogenicus subsp.melaninogenicus took up 2.6 x 10-4 to 5.4 x 10-4 ,mol of estradiol orprogesterone per ,ug of cell protein. Minimal steroid uptake was observed with B.gingivalis and five other organisms. Uptake of steroids by B. melaninogenicussubsp. intermedius was temperature dependent and resulted in a labeled productas detected on thin-layer chromatography. Growth curves indicated that interme-dius and melaninogenicus subspecies of B. melaninogenicus but not B. gingivaliscould substitute progesterone or estradiol for vitamin K, an essential growthfactor. Growth of B. melaninogenicus subsp. intermedius in steroids was concen-tration dependent. Addition of fumarate to resting cells of B. melaninogenicussubspecies as well as B. gingivalis increased steroid uptake by 70 to 500% andresulted in the gas-liquid chromatographic detection of succinate. Cultures givenfumarate alone or steroids alone produced no succinate. Steroids appeared todirectly interact with the fumarate reductase system and foster the growth of B.melaninogenicus subsp. intermedius. This interaction may be of ecologicalsignificance.
In studies of the plaque flora associated withperiodontal disease, the proportions of certainbacterial species are greater than the propor-tions observed in gingival health (19, 26-28, 32,35). If periodontal disease involves increases inspecific components of the indigenous flora (17),the determination of the factors which allow orinitiate a shift in this microflora is essential to anunderstanding of the etiology of the disease.The subgingival bacterial flora of pregnant
individuals changes as pregnancy progresses(13). In the second trimester, a significant in-crease was observed in the ratio of anaerobic tofacultative bacteria and the proportional levelsof Bacteroides melaninogenicus subsp. interme-dius. In addition, uptake of [14C]estradiol and[14C]progesterone by dental plaque samples in-creased significantly during pregnancy and par-alleled the plaque proportions of B. melanino-genicus subsp. intermedius. This suggests thatendogenous steroids may influence the microbi-al ecology of the gingival sulcus.
In this communication, we report the identifi-
t Present address: Department of Periodonttics, Universityof Connecticut Health Center, Farmington, CN 06032.
cation of a role for steroid utilization in themetabolism of B. melaninogenicus subsp. inter-medius and B. asaccharolyticus (B. gingivalissp. nov. 6).
MATERIALS AND METHODSMicrobiological methods. Pure cultures ofActinomy-
ces naeslundii, A. viscosus, B. gingivalis, B. melanin-ogenicus subsp. intermedius, Capnocytophaga ochra-ceus, Fusobacterium nucleatum, and Streptococcussanguis were isolated from subgingival plaque samplesfrom a bacteriological study of pregnancy gingivitis. B.melaninogenicus subsp. melaninogenicus ATCCstrain 25845 was obtained from the American TypeCulture Collection. Each organism was grown anaero-bically to exponential phase in basal anaerobic broth(BAB; 30) with menadione (0.5 ,ug/ml, 2.9 ,uM), centri-fuged at 16,000 x g for 15 min, and washed two timesin reduced transport fluid (RTF; 18) without EDTA.Washed cell concentrates were assayed for protein(fluorescamine; 34) and diluted with RTF to 0.5 to 1.5,ug of protein per 50 pl.
[14C]steroid uptake. Fifty-microliter portions of thecell suspension were dispersed ultrasonically (Kontesmodel K88144, 5 s) and incubated anaerobically withradioactive estradiol and progesterone to evaluateuptake of these substrates by bacterial cell suspen-
256
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STEROID EFFECTS ON BACTEROIDES 257
sions. All steroid uptake assays were run three times,each in duplicate.
Radioactive estradiol and progesterone were ob-tained from Amersham/Searle as follows: [4-4C]estra-diol, specific activity of 56.7 mCi/mmol in benzene/ethanol; [4-14C]progesterone, specific activity of 58.8M mCi/mmol in benzene. Both steroids were taken todryness in a stream of nitrogen and diluted in ethanol(absolute) to a concentration of 1.82 x 10-3 ,mol/50 I1for estradiol and 3.40 x 10-3 Rmol/50 ,ul for progester-one.The assay was carried out in round-bottom plastic
microtiter plates (Falcon Plastics). Preliminary studies(data not shown) evaluated the effect of divalentcations, AMP, NAD, and NADH on the uptake ofestradiol and progesterone by Bacteroides species.The following components were found to give optimalsteroid uptake: 50 R1 of ["4C]estradiol (1.82 x 10-3,umol); 50 p1 of ['4C]progesterone (3.40 x 10-3 ,mol);and 50 p11 of RTF plus NADH (1.64 x 10-4 FM) plusMgCl2 (1.00 X 10-4 ,uM).The microtiter plate was covered with a self-adhe-
sive sealing tape, placed on a microtiter plate shaker,and taken into an anaerobic glove box (Coy Manufac-turing Co., Ann Arbor, Mich.) with an atmosphere of85% nitrogen, 5% carbon dioxide, and 10%o hydrogen.The assay was initiated by lifting the sealing tape andplacing 50 P1 of each sonified cell suspension, RTF, orheat-killed controls into the triplicate wells. The platewas resealed and vibrated at low speed for 10 s initiallyand again at 30 min.At 60 mnm, the sealing tape over each well was
punctured with a 10-pA Hamilton syringe, and 1 ,u ofthe cell suspension was removed by means of aCheney adapter and spotted at the origin of a silica gelthin-layer chromatography (TLC) sheet (Eastman Ko-dak 13179 without fluorescent indicator). The TLCplates were removed from the anaerobic chamber, andto each sample spot was added 1 pl each of unlabeledestradiol (2% wt/vol) and progesterone (2% wt/vol).Plates were developed vertically in chloroform-acetal-dehyde (80/20, vol/vol) to a solvent height of 100 mm.Steroids were visualized by means of a vanillin spray(0.5% vanillin in 1 part H2SO4 [1.0%/] and 4 partsabsolute ethanol; 29).The TLC spots with Rf values corresponding to the
estradiol and progesterone were cut out and placed inliquid scintillation vials with 10 ml of scintillation fluid(3% Biosolv [Beckman Instruments, Inc.] and 15 g ofOmnifluor [New England Nuclear Corp.] in 1 gallon[ca. 3.8 liters] of technical-grade toluene) and quanti-tated by means of a liquid scintillation counter (Beck-man LS 100). The quantities of [14C]estradiol and['4C]progesterone remaining in the bacterial suspen-sions were subtracted from the RTF controls to calcu-late the amount of substrate taken up by the bacterialsuspensions. The quantity of substrate uptake wascalculated per microgram of bacterial protein added tothe reaction well. Before the steroid uptake studieswere started the TLC procedure, was evaluated forquantitative recovery of [14C]estradiol and [14C]pro-gesterone. Since steroids are frequently destroyed byTLC procedures, the visualization reagents, scintilla-tion fluids, and developing solvents were tested foreffects on quantitation (data not included). The TLCprocedures eventually used were found to give optimalquantitative recovery.
Heat-killed controls were prepared by heating aportion of the cell suspension in a water bath at 80°Cfor 30 min. A culture plate was streaked with theheated suspension and incubated to confirm the lack ofviable cells. Attempts to determine steroid uptakeduring cell growth were not successful due to nonspe-cific oxidation-reduction of the estradiol and proges-terone when incubated for prolonged periods in thereduced medium.The effect of temperature on steroid uptake was also
evaluated. Microtiter plates were prepared withoutbacteria, sealed, and placed at 0°C for 2 h. The plateswere then taken into the anaerobic chamber, and thesealing tape was removed. Cell suspensions or RTFwere added to appropriate wells to initiate the reac-tion. The plates were then resealed with two layers ofadhesive plastic strips and immediately returned to0°C for 60 min before taking 1-IlI samples for the TLCassay of steroid uptake.
Autoradiographs using no-screen X-ray film (KodakNS54T) were prepared from the TLC sheets to deter-mine the location of the 14C label after the steroiduptake assay.
Gas-liquid chromatography. Succinate and fumaratewere sought for in the supernatants of the steroiduptake assays. Their methyl derivatives were preparedby the method of Holdeman et al. (10). A VarianAerograph series 2700 gas-liquid chromatograph wasused with a flame ionization detector. Temperatureswere maintained at 150°C in the column, 200°C in thedetector, and 175C at the injector port. Standardsuccinate and fumarate solutions were freshly methyl-ated and chromatographed on the day of sampleanalysis. Detection of fumarate and succinate stan-dards in the range of 2.0 to 7.0 pumol/100 pl offumarateand 0.2 to 0.7 pLmol/100 p1 of succinate was found to belinear.Growth studies. Vitamin K, or an analog such as
menadione, is essential to the growth of most strains ofB. melaninogenicus and B. asaccharolyticus (8, 16).Structural similarities with regard to electron transferbetween estradiol, progesterone, and vitamin K sug-gested the possibility that steroid hormones might beable to substitute for vitamin K compounds in thegrowth of pigmented Bacteroides species. Therefore,growth of these organisms was evaluated in thepresence of estradiol or progesterone and in the ab-sence of menadione in the medium.The following organisms were maintained in pure
culture on enriched Trypticase soy agar plates (31): B.gingivalis strains W, 165.5, and 208.1; B. melanino-genicus subsp. intermedius strains 155.6, 166.5, and167.4; and B. melaninogenicus subsp. melaninogeni-cus strain ATCC 25845. A single colony was subcul-tured to 5 ml of BAB and incubated until the culturereached an optical density of 0.30 at 540 nm asmeasured in a Spectronic 20 spectrophotometer(Bausch & Lomb, Inc.) present within the anaerobicchamber.
Cultures were Gram stained and streaked on en-riched Trypticase soy agar plates to confirm purity,and 0.1 ml was inoculated into the following media: (i)BAB containing 0.5 pg of menadione per ml (2.9 F.M);(ii) BAB without menadione; (iii) BAB without mena-dione plus estradiol, equimolar or twice (5.8 p,M) orthree times (8.7 ,uM) the concentration of menadionein BAB; and (iv) BAB without menadione plus proges-
VOL. 35, 1982
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

258 KORNMAN AND LOESCHE
terone, equimolar or twice or three times the concen-tration of menadione in BAB. All broth media werereduced and incubated within the anaerobic chamberfor 48 h before use.
Optical density at 540 nm was monitored until astationary phase was reached. At that time 0.1 ml wastransferred to a second tube of the same medium, andgrowth was again monitored. If growth occurred in thesecond transfer, then 0.1 ml of that stationary-phaseculture was used to inoculate a third tube of the samemedium, and growth was again monitored. This proce-dure was used to prevent influences on growth fromcarryover of nutrients present in the original 0.1-mlinoculum from BAB. In separate experiments, opticaldensity was related to dry weight by means of astandard curve derived for each organism. Growthcurves represent the mean (± standard deviation) ofthree separate studies.
RESULTSUptake of 14C-labeled steroids by pure cultures
of bacteria. Steroid uptake was consistently ob-served in strains of oral Bacteroides, with B.melaninogenicus subsp. intermedius and subsp.melaninogenicus taking up substantially moreestradiol and progesterone than B. gingivalis(Table 1). There was no detectable uptake ofeither [14C]estradiol or [14C]progesterone by C.ochraceus, F. nucleatum, or S. sanguis) Table1). The uptake of both steroids by A. viscosuswas similar to uptake observed with B gingivalis,whereas A. naeslundii took up progesterone butnot estradiol. Six different batches of strain155.6 of B. melaninogenicus subsp. intermediusconsistently showed steroid uptake, with themean uptake of the six experiments being 4.06 ±1.22 x 10-4 ,umol/4Lg of protein for estradiol and2.21 ± 0.30 x 10-4 ,umol/,ug of protein forprogesterone.The reaction mixtures from steroid uptake
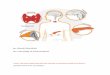
studies with B. melaninogenicus subsp. interme-
dius strain 155.6 were spotted on TLC sheetsand developed. The radioactivity in various spotswas demonstrated by autoradiography. Reac-tion mixtures with RTF instead of bacteria (Fig.1A) revealed labeled spots with Rf values of 0.69and 0.97 which were compatible with the loca-tions of progesterone and estradiol, respective-ly. Reaction mixtures B to G, to which wereadded increasing concentrations of bacteria,also revealed labeled spots for residual proges-terone and estradiol and at an Rf of 0.53 as in thecontrol. Additional labeled spots were evident atthe origin and with Rf values of 0.25 and 0.84.The labeling at the origin represents 14C-labeledcompounds which were cell bound, and the spotat Rf 0.25 appears to represent diffusion of cell-bound material which did not increase with theincrease in bacterial concentration from B to F.The labeled spot at Rf0.84 was not present in thecontrol and appears to be an unknown productof the labeled steroid utilization, since it in-creased with bacterial concentration. Of thetotal uptake, 26% was accountable in this un-known spot and 62% was located at the origin,apparently bound to the bacterial cells depositedwith the sample. Uptake of 14C-labeled steroidwas found to be temperature dependent. Uptakeof progesterone by viable B. melaninogenicussubsp. intermedius incubated at 0 and 37°C was9.6 and 20.9 times the uptake by heat-killed cellsat 37°C. Uptake of estradiol by viable cells at 0and 37°C was 16.7 and 32.9 times the uptake byheat-killed cells.
Utilization of steroids for growth ofB. melanin-ogenicus. B. gingivalis failed to grow in mediumdepleted of menadione (Table 2). The substitu-tion of estradiol or progesterone for menadionedid not support growth of B. gingivalis. B.melaninogenicus subsp. intermedius strains also
TABLE 1. Uptake of 14C-labeled estradiol and progesterone by pure cultures of oral bacteria
Organism Strain Uptakea of:
Estradiol Progesterone
A. naeslundii 401N 0 1.09 ± 0.22A. viscosus 378.5 0.79 ± 0.09 1.76 ± 0.28B. gingivalis w 0 0.41 ± 0.06
167.5 0.71 ± 0.08 1.76 ± 0.82208.1 0.42 ± 0.10 0.83 ± 0.44
B. melaninogenicus subsp.intermedius 155.6 2.15 ± 1.44 2.41 ± 0.42
166.5 3.35 ± 0.48 3.12 ± 0.81167.4 5.48 ± 0.68 2.48 ± 0.39
B. melaninogenicussubsp. melaninogenicus ATCC 25845 2.96 + 0.42 4.12 ± 1.70
C. ochraceus 146.1 0 0F. nucleatum 395.4 0 0S. sanguis 399.5 0 0
a Mean micromoles uptake x 10-4/microgram of cell protein ± standard deviation for triplicate samples.
INFECT. IMMUN.
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STEROID EFFECTS ON BACTEROIDES 259
* *e
A B c E F G
FIG. 1. Autoradiograph of TLC of "C-labeledsteriod uptake. (A) Standard steroid uptake system towhich was added estradiol, progesterone, and RTF,but no bacteria. (B-G) Standard steroid uptake systemto which was added estradiol, progesterone, RTF, andincreasing quantities of B. melaninogenicus subsp.intermedius strain 155.6.
did not grow in the absence of menadione.However, when estradiol (2.91 ,uM) was substi-tuted for menadione, growth approached thatobserved with menadione (Table 2 and Fig. 2). Atwofold increase in estradiol inhibited growthslightly, whereas a threefold increase markedlyreduced growth. Progesterone at 5.8 and 8.7 ,uMsubstituted for menadione in the growth of B.melaninogenicus subsp. intermedius, and no re-duction in growth was observed at the higherconcentrations (Table 2 and Fig. 3).The growth pattern of B. melaninogenicus
subsp. melaninogenicus was different from thatof the other subspecies, as this subspecies grewto some extent in the absence of menadione. The
substitution of estradiol (2.9 and 5.8 ,uM) stimu-lated growth, but not to extent seen with mena-dione. A higher concentration of estradiol (8.7,uM) inhibited growth. Progesterone stimulatedgrowth of this organism only at the highestconcentration.
Interaction of steroids with the fumarate sys-tem. The addition of fumarate to the assay foruptake of 14C-labeled steroids (Table 3) in-creased the uptake of estradiol and progesteroneby B. gingivalis and the subspecies of B. melan-inogenicus. Uptake of steroids by B. gingivalisstrain W was negligible but, with the addition offumarate, increased to a level intermediate be-tween those of the two subspecies ofB. melanin-ogenicus. When the supernatants of the uptakeassay were methylated and analyzed by gas-
liquid chromatography, the addition of fumarategave rise to trace amounts of succinate. Howev-er, fumarate plus steroids greatly enhanced suc-cinate production (Table 4).
DISCUSSIONChanges in endogenous steroid levels have
been associated with alterations in the vaginaltract microflora (6, 9, 14), the urinary tract flora(21, 25, 29), the gastrointestinal tract flora (1, 7),and the oral subgingival flora (13). Most investi-gators have attributed hormonally associatedmircobial shifts to steroid-induced tissue andimmunological changes and not to direct effectsof the steroids on bacteria. Studies of gram-positive organisms have shown that various ste-roids inhibit in vitro growth and enhance cellleakage (15, 36).Progesterone has also been reported to inhibit
growth of Neisseria gonorrhoeae and to have noeffect on the growth of 14 other facultative gram-
TABLE 2. Effect of menadione and steroids on growth of B. melaninogenicus and B. gingivalis
B. gingivalis B. melaninogenicus subsp. B. melaninogenicus subsp.intermedius melaninogenicus
BAB additivesDoubling Maximal Doubling Maximal Doubling Maximaltimea cel mass (%) time cell mass (%) time cel mass (%)
Menadione,2.9 ,uM 21 100 14 100 12 100
None NG NG 24 32.4Estradiol
2.9 ,uM NG 16 82.0 12 62.15.8 ,LM NG 20 28.3 12 62.88.7 ,uM NG NG 24 27.0
Progesterone2.9 FM NG 14 98.6 NG5.8 ,uM NG 12 102.6 NG8.7 FxM NG 9 99.3 12 148.0
a Hours required for cell mass, as measured by turbidity, to double during the exponential growth phase. NG,No growth.
b Maximal turbidity obtained over % h was converted to cell mass by means of standard curves relatingturbidity to dry weight for each organism. Growth in complete medium (BAB) plus 2.9 ,uM menadione was takenas 100o.
VOL. 35, 1982
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

260 KORNMAN AND LOESCHE
E 0.050
H ElIamm-NBAB3 minus msnadione
10 30 50 70 90HOURS
FIG. 2. Growth in complete medium (BAB which included 2.9 ,uM menadione), BAB without menadione, orBAB without menadione to which was added estradiol in equimolar, twice (5.8 FM), or three times (8.7 ,M) theconcentration of menadione in BAB. Data represent mean cell masses of strains 155.6, 166.5, and 167.4.
negative species (23). In the present study, B.melaninogenicus was found to have an affinityfor estradiol and progesterone which was notdemonstrated by other tested oral bacteria.Some aspects of steroid uptake may reflect
binding to the cell wall rather than actual utiliza-tion. Morse and Fitzgerald (23) observed thatbinding of [14C]progesterone was three timesgreater to Neisseria species which were meta-bolically sensitive to the steroid than to insensi-tive cells. In our study, the uptake of steroidsappeared to reflect an active metabolic process,
since the uptake was temperature dependent andresulted in the detection of an unknown labeledby-product with an Rf value intermediate be-tween those of progesterone and estradiol. Pre-liminary chromatographic studies (data notshown) suggest that this compound may be 5-,B-pregnane 3-20 dione which would result from5-1 reductase activity on progesterone. Furtheranalyses of this unknown product are in prog-ress.Mixed cultures of human fecal bacteria have
been shown to dehydroxylate or dehydrogenate
E~~~~
E 0.100
BAB minus menadione10 30 SO 70 90
HOURS
FIG. 3. Growth ofB. melaninogenicus subsp. intermedius in complete medium (BAB which included 2.9 ,uMmenadione), BAB without menadione, or BAB without menadione to which was added progesterone inequimolar, twice (5.8 ,uM), or three times (8.7.,uM) the concentration of menadione in BAB. Data represent meancell masses of strains 155.6, 166.5, and 167.4.
INFECT. IMMUN.
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STEROID EFFECTS ON BACTEROIDES 261
TABLE 3. Effect of fumarate on uptake of 14C-labeled estradiol and progesterone
Organism Fumarate added' Total steroid Steroid uptakeuptakeb fumarate+/fumarate-
B. melaninogenicus subsp.intermedius strain 155.6 4.56 ± 0.93 1.5
+ 6.70 ± 0.76B. gingivalis strain W 0.41 ± 0.03 21.4
+ 8.76 ± 1.07B. melaninogenicus subsp.
melaninogenicus strainATCC 25845 5.71 ± 1.40 2.2
+ 12.37 ± 1.50a Fumarate added to standard reaction mixture to final concentration of 0.5%.b Mean micromoles uptake of labeled estradiol and progesterone x 10-4/microgram of cell protein ± standard
deviation for triplicate samples.
deoxycorticosterone (2, 12), bile salts and an-drosterone (20), estrone and estradiol (11), andprogesterone (1). In addition, B. fragilis (V. C.Aries and M. J. Hill, Biochem. J. 119:37p-38p,1970) and B. thetaiotaomicron (24) have beenshown to exhibit 7-a-hydroxysteroid dehydro-genase activity in vitro, and pure cultures of B.fragilis appear capable of reducing estrone toestradiol (11).However, chemical changes resulting from
bacterial action and binding of a compound mayrepresent reactions which are not truly involvedin physiological processes. The hypothesis thatsteroids directly affect subspecies ofB. melanin-ogenicus is strengthened by the observation thatestradiol and progesterone can substitute forvitamin K compounds, an essential growth fac-
TABLE 4. Effect of steroids and fumarate onsuccinate detection
Steroids Fumarate SuccinateOrganism added' addedb detectedc
B. gingivalis strain W + - 0+ + 0.48_ - 0
_ + <0.01B. melaninogenicus
subsp. intermediusstrain 155.6 + - 0
+ + 0.52_ - 0
- + 0.03B. melaninogenicus
subsp.melaninogenicusstrain ATCC 25845 + - 0
+ + 0.61_ - 0
- + 0.04a Estradiol was added at 1.82 x 10-3 ,umol; proges-
terone was added at 3.77 x 10-3 ,umol.b Fumarate was added at 8.60 ,umol.c Micromoles/100-ILl sample as detected by gas-
liquid chromatography.
tor, in the metabolism of these organisms. Theability of the steroids to substitute for vitamin Kis compatible with earlier observations that theenzymes involved in vitamin K utilization by B.melaninogenicus appear to have broad substratespecificity (8, 16).
Steroids have not been previously reported toenhance bacterial growth. In fact, gonadal ste-roids have been shown to inhibit growth of avariety of gram-positive organisms (3, 36) aswell as N. gonorrhoeae (23). The various growtheffects of different concentrations of estradioland progesterone as substitutes for vitamin K inB. melaninogenicus are consistent with previousreports (16) of a narrow concentration range forvitamin K stimulation of B. melaninogenicus,with inhibition of growth observed at highervitamin K concentrations. Substrate inhibitionwas also reported for a hydroxysteroid dehydro-genase isolated from B. thetaiotaomicron, inwhich enzyme Km values were low and substan-tial inhibition was observed at steroid concentra-tions greater than 1 mM (24).
In the present study, fumarate, a terminalelectron acceptor in anaerobic respiration, in-creased steroid uptake by B. melaninogenicussubspecies and B. gingivalis and was reduced tosuccinate in the presence of estradiol and pro-gesterone. Naphthoquinones, such as vitamin Kor menadione, have been implicated as electroncarriers in the bacterial anaerobic reduction offumarate to succinate (33). In B. thetaiotaomi-cron, fumarate has been shown to markedlyincrease 7-a-hydroxysteroid dehydrogenase ac-tivity (24). Therefore, the effect of fumarate onthe uptake of estradiol and progesterone addsfurther support to the finding that steroids maysubstitute for vitamin K analogs in these micro-organisms. The fact that B. gingivalis does notroutinely produce succinate as an end product,yet is able to do so if given exogenous fumarateplus estradiol and progesterone, raises the possi-bility that this organism has a block in fumaratesynthesis.
VOL. 35, 1982
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

262 KORNMAN AND LOESCHE
In general, a role for estradiol and progester-one in the metabolism of B. melaninogenicusand B. gingivalis is suggested by four observa-tions: (i) there was a significant increase in theuptake of [14C]estradiol and [14C]progesteroneby dental plaque samples taken from preg-nant patients in the second trimester, and theincreased uptake paralleled increased plaqueproportions of B. melaninogenicus subsp. inter-medius (13); (ii) pure cultures of B. melanino-genicus and B. gingivalis took up 14C-labeledestradiol and progesterone; (iii) estradiol andprogesterone substituted for menadione, an es-sential growth factor, in the growth of B. melan-inogenicus subspecies; and (iv) the uptake ofestradiol and progesterone was increased by theaddition of fumarate, with fumarate apparentlybeing reduced to succinate in the presence ofsteroids.These findings strongly suggest that estradiol
and progesterone are involved in the fumaratereductase system of subspecies of B. melanino-genicus and B. gingivalis. In addition, the inter-changeability of menadione and gonadal steroidraises the possibility that estrogens or progester-one, which are readily available in the gingivalsulcular fluid (4), may be a primary growthfactor in vivo for B. melaninogenicus. Steroidhormones therefore appear to have the potentialfor altering the subgingival microbial ecology bydirectly influencing the metabolic pathways ofB. melaninogenicus.
Several mucosal diseases, including periodon-titis and urinary tract infections, are known to becaused by bacteria but have been frequentlyassociated with altered hormone levels. Theidentification of the potential for endogenoussteroids to directly induce a shift in the normalmicroflora may provide new insights to theassociation of periodontal disease with condi-tions characterized by altered steroid levels.
ACKNOWLEDGMENTSThis investigation was supported by Public Health Service
research grants DE 03011 and DE 02731 from the NationalInstitute of Dental Research. K.S.K. was a recipient of PublicHealth Service fellowship DE 00013 from the same institute.We thank Tom Ruprecht for his assistance in this project.
LITERATURE CITED1. Adlercreutz, H., F. Martin, P. Jarvenpaa, and T. Fotsls.
1979. Steroid absorption and enteroheoptic recycling con-traception. J. Steroid Biochem. 20:201-223.
2. Bokkenheuser, V. D., J. Winter, and W. G. Kelly. 1978.Metabolism of biliary steroids by human fecal flora. Am.J. Clin. Nutr. 31:S221-S226.
3. Casas-Campfllo, C., D. Balandrano, and A. Galrza. 1961.Antimicrobial properties of 21,-dimethoxy progesteroneand other progesterone analogues. J. Bacteriol. 81:366-375.
4. Cimasoni, G. 1974. The crevicular fluid. Monogr. OralSci. 3:1-74.
5. Coykendall, A. L., F. S. Kaczmarek, and J. Slots. 1980.
Genetic heterogeneity in Bacteroides asaccharolyticus(Holdeman and Moore 1970). Finegold and Barnes 1977(Approved Lists 1980) and proposal for two new species:Bacteroides gingivalis sp. nov. and Bacteroides macacaecomb. nov. Int. J. Syst. Bacteriol. 30:559-564.
6. Crulkahank, R., and A. Shanran. 1934. The biology ofthe vagina in the human subject. II. The bacterial flora andsecretion of the vagina in relation to glycogen in thevaginal epithelium. J. Obstet. Gynaecol. Br. Emp. 41:208-266.
7. Drasar, B. S., and M. J. HUI. 1974. Human intestinal flora.Academic Press, Inc., New York.
8. Gibbons, R. J., and J. B. MacDonald. 1960. Hemin andvitamin K compounds as required factors for the cultiva-tion of certain strains of Bacteroides melaninogenicus. J.Bacteriol. 80:164-169.
9. Goplerud, C. P., J. J. Ohm, and R. P. Galsk. 1976.Aerobic and anaerobic flora of the cervix during pregnan-cy and the puerperium. Am. J. Obstet. Gynecol. 126:858-868.
10. Holdeman, L. V., E. P. Cato, and W. E. C. Moore (ed.).1977. Anaerobe laboratory manual, 4th ed. Virginia Poly-technic Institute and State University, Blacksburg.
11. Jarvenpaa, P., T. Kosunen, T. Fotasi, and H. Adkrcreutz.1980. In vitro metabolism of estrogens by isolated intesti-nal micro-organisms and by human fecal microflora. J.Steroid Biochem. 13:345-349.
12. Kelly, W. G., 0. DeLeon, J. Winter, and V. D. Bokken-heuser. 1977. Exchange of hydrogen at C-21 during dehy-droxylation of deoxycorticosterone by mixed cultures ofhuman fecal flora. J. Steroid Biochem. 8:73-75.
13. Kornman, K. S., and W. J. Loesche. 1980. The subgingivalmicrobial flora during pregnancy. J. Periodontal Res.15:111-112.
14. Larsen, B., A. J. Markovetz, and R. P. Galsk. 1976.Quantitative alterations in the genital microflora offemalerats in relation to the estrous cycle. J. Infect. Dis.134:486-489.
15. Lester, G., and 0. Hetcher. 1966. Effects of deoxycortico-sterone on growth of microorganisms. J. Bacteriol.92:627-635.
16. Lev, M. 1959. The growth-promoting activity of com-pounds of vitamin K group and analogues for a rumenstrain of Fusiformis nigescens. J. Gen. Microbiol. 20:697-703.
17. Loesche, W. J. 1976. Chemotherapy of dental plaqueinfections. Oral Sci. Rev. 9:65-107.
18. Loesche, W. J., R. Hockett, and S. A. Syed. 1972. Thepredominant cultivable flora of tooth surface plaque re-moved from institutionalized subjects. Arch. Oral Biol.17:1311-1325.
19. Loesche, W. J., and S. A. Syed. 1978. Bacteriology ofhuman experimental gingivitis II. Effect of plaque andgingivitis score. Infect. Immun. 21:830-839.
20. MacDonald, I. A., G. R. Webb, and D. E. Mahony. 1978.Fecal hydroxysteroid dehydrogenase activities in vegetar-ian Seventh-Day Adventists, control subjects, and bowelcancer patients. Am. J. Clin. Nutr. 31:S233-S238.
21. Marrie, T. J., C. A. Swantee, and M. Harden. 1980.Aerobic and anaerobic urethral flora of healthy females invarious physiological age groups and of females withurinary tract infections. J. Clin. Microbiol. 11:654-659.
22. Marshall, S. and J. Linfoot. 1977. Influence of hormoneson urinary tract infection. Urology 9:675-678.
23. Morse, S. A., and T. J. Fitzgerald. 1974. Effect of proges-terone on Neisseria gonorrhoeae. Infect. Immun.10:1370-1377.
24. Sherrod, A., and P. B. Hyleman. 1977. Partial purificationand characterization of NAD-dependent 7-alpha-hydroxy-steroid dehydrogenase from Bacteroides thetaiotaomi-cron. Biochim. Biophys. Acta 486:351-358.
25. Silk, M., and D. B. Perez-Vorela. 1970. Effects of oralcontraceptives on urinary bacterial growth rate. Invest.Urol. 8:239-245.
INFECT. IMMUN.
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STEROID EFFECTS ON BACTEROIDES 263
26. Slots, J. 1977. Microflora in the healthy gingival sulcus inman. Scand. J. Dent. Res. 85:274-254.
27. Slots, J. 1977. The predominant cultivable microflora ofadvanced periodontitis. Scand. J. Dent. Res. 85:114-121.
28. Slots, J., D. Moenbo, J. Ian_ek, and A. Frandsen.1978. Microbiota of gingivitis in man. Scand. J. Dent. Res.86:174-181.
29. StaUl, E. 1969. Thin layer chromatography, 2nd ed. Aca-demic Press, Inc., New York.
30. Syed, S. A. 1980. Characteristics ofBacteroides asacchar-olyticus from dental plaques of beagle dogs. J. Clin.Microbiol. 11:522-526.
31. Syed, S. A., M. Svanberg, and G. Svanberg. 1980. Thepredominant cultivable dental plaque flor of beagle dogswith gingivitis. J. Periodontal Res. 15:123-136.
32. Tanner, A. C. R., C. Haffer, G. T. Bratthal, R. A.Viconti, and S. S. Socransky. 1979. A study of the
bacteria associated with advancing periodontitis in man.J. Clin. Periodontics 6:278-307.
33. Thaner, R. K., K. Jn n, and K. Decker. 1977.Energy conservation in chemotrophic anaerobic bacteria.Bacteriol. Rev. 41:100-180.
34. Udenfrlend, S., S. Stein, and P. Bobkn. 1972. Fluoresca-mine: a reagent for assay of amino acids, peptides,protein, and primary amines in the picomole range. Sci-ence 178:871-872.
35. Van Palensten-Hddeman, W. H. 1975. Total viable countand differential counts of vibrio (Campylobacter) spur-torum, Fusobacterium nucleatum, Selenomonas sputi-gena, Bacterioides ochraceus, and Veilloneila in the in-flamed and non-inflamed human gingival crevice. J.Periodontal Res. 10:294-301.
36. Yots, W. W. 1967. In vivo and in vitro action of norethin-drone on staphylococci. J. Bacteriol. 94:1353-1358.
VOL. 35, 1982
on February 7, 2020 by guest
http://iai.asm.org/
Dow
nloaded from