Embed Size (px)
Citation preview

Acta physiol. scand. 1979. 105. 461-468 From the Department of Physiology, University of Goteborg, Sweden
Effects of vasoactive intestinal polypeptide on blood flow, motility and fluid transport in the gastrointestinal tract of the cat
BY
STEFAN EKLUND, MATS JODAL, OVE LUNDGREN and ANDERS SJOQVIST
Received 18 September 1978
Abstract
EKLUND, S., M. JODAL, 0. LUNDGREN and A. SJOQVIST. Effects of vasoactive intestinal polypeptide on blood flow, motility and fluid transport in the gastrointestinal tract of the cat. Acta physiol. scand. 1979. 105. 461-468.
The effect of close intraarterial infusions of vasoactive intestinal polypeptide (VIP) on gastric motility, intestinal fluid transport and colonic motility were studied in the cat. Regional blood flow was also fol- lowed in all experiments. In the stomach VIP produced a gastric relaxation and a blood flow increase. The motility response was similar to that observed when eliciting the vago-vagal reflex relaxation by dis- tending the esophagus. In the small intestine a hyperemia and a decrease of net water uptake was observed. When infusing small amounts of VIP a decrease of net water uptake was seen without any change of in- testinal blood flow. Large amounts of VIP produced a transient secretory state in the small intestine. In the colon a hyperemia was seen immediately upon starting the infusion of the drug. After 2-3 min of in- fusion a contraction of the colon was apparent. The administration of atropine to the animal did not significantly affect any of the responses produced by VIP. The results are discussed in relation to VIP as a possible neurotransmitter in the gastrointestinal tract.
Vasoactive intestinal polypeptide (VIP), a compound with a large spectrum of biological actions, was originally isolated from the gastrointestinal tract (Said and Mutt 1970). The molecular mass of VIP exceeds 3 000 d and it is composed of 28 amino acids. The sequence of the amino acids is fully established and it resembles glucagon and secretin. These well established hormones produce biological effects similar to VIP. It has therefore been labelled a “candidate hormone”, together with several other peptides recently isolated from the gastrointestinal tract (Grossman et al. 1974).
Immunohistochemical methods for studying the tissue localization of VIP have, how- ever, demonstrated that most, if not all, VIP is localized to nervous tissue not only in the brain but also in peripheral organs (see e.g. Larsson et al. 1976). Furthermore, immuno- histochemical and biochemical techniques show that VIP is predominantly localized to synaptosomes (Giachetti et al. 1977, Larsson 1977, Emson et al. 1978), suggesting that VIP may be a neurotransmitter. This proposal was further strengthened by the demonstration of a VIP release from the alimentary canal upon direct electric stimulation of the vagal and
461

462 STEFAN EKLUND, MATS JODAL, OVE LUNDGREN AND ANDERS SJOQVIST
pelvic nerves, and also when various types of reflexes are induced via these nerves (Fahren- krug et al. 1978 a, b).
One of the criteria for a neurotransmitter is that it should elicit the same response in a n organ when administered intravascularly as when released from nervous endings. How- ever, there is a paucity of data concerning the effects in uiuo of intravascularly administered VIP on gastrointestinal function, particularly with respect to motility. Thus, in the present study the effect of intravascularly infused VIP on motility, blood flow and water absorption in the gastrointestinal canal in situ was investigated.
Methods Animals and general operative procedures
The experiments were performed on 14 cats, anaesthetized with chloralose (50 mg/kg b.wt.) after induction with ether. The cats had been deprived of food for at least 24 h with free access to water. They exhibited no obvious signs of gastrointestinal disease.
The following procedures were common to all types of experiments performed in this study: After opening the abdomen by a midline incision, the great omentum and the spleen were extirpated. The femoral artery was cannulated after hepariniration (3-5 mg/kg b.wt. i.v.) to record mean arterial blood pressure by means of a Statham pressure transducer (model P23AC). All measured parameters (see below) were recorded on a Grass polygraph. The splanchnic nerves were cut bilaterally.
A slow intravenous infusion of a glucose solution containing bicarbonate (see below) was started at the time of the induction of anaesthesia and then continued throughout the experiment. This infusion has previously been shown to maintain arterial pH at a normal level, despite varying degrees of operative trauma (Haglund and Lundgren 1972).
Experiments on the stomach
A loose ligature was placed around the cardia without damaging the vagal nerve trunks. A wide-bore plastic catheter was introduced into the stomach via the pylorus through a duodenostomy and connected to a volume transducer for continuous recording of changes in gastric volume. The stomach and the volume recording system was filled with isotonic saline and arranged in such a way that the intragastric pressure could be set at, and continuously adjusted to. the desired level, usually 2 cm above that at the pylorus.
Gastric blood flow was registered aia a catheter in the splenic vein connected to a drop recorder unit operating an ordinate writer. All other veins draining the stomach were ligated. Intra-arterial infusions of VIP to the stomach was made into the hepatic artery in a retrograde direction. A vago-vagal receptive relaxation of the stomach was elicited reflexly by distension of the esophagus (Abrahamsson and Jansson 1969). This was achieved by filling a small balloon, placed in the proximal part of the esophagus, with 20 ml saline.
Experiments on the small intestine
An 8-10 cm long jejunal segment was isolated with its blood supply intact. The colon and remaining parts of the small intestine were extirpated. The jejunal segment was so chosen that its arterial supply was derived from one single main branch of the superior mesenteric arteri and La. infusions of VIP were in most ex- periments performed via an adjacent branch. Total venous outflow from the jejunal segment and its lymph nodes was registered uia a catheter in the superior mesenteric vein, as described above.
Net intestinal transport of fluid was measured with the method described in detail by Jodal el al. (1975), where the intestinal lumen was perfused with a body-warm Krebs-glucose solution (see below) at a rate of 1 ml/min in a recirculating system. Changes in motility and/or net fluid transport were registered by a volume transducer connected to the system via a T-tube.
Experiments on the colon
In these experiments the small intestine was extirpated and the inferior mesenteric artery was divided. This procedure has been shown not to decrease blood flow to the feline large bowel (Hulten et d. 1969) which is amply supplied uia the superior mesenteric artery. The distal part of the colon was extirpated,
VIP AND CI FUNCTION 463
GASTRIC BLOOD
ml/min xlOOg
TIME .min 1 I I I I I
FLOW, i:q - VIP-infusion
0' 5 10 15 20 25 30 35
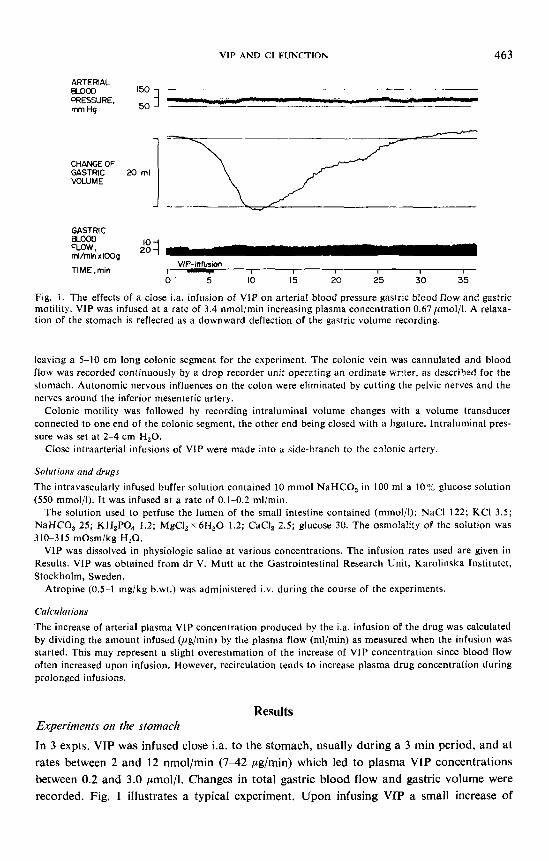
Fig. I . The effects of a close i.a. infusion of VIP on arterial blood pressure gastric blood flow and gastric motility. VIP was infused at a rate of 3.4 nmol/min increasing plasma concentration 0.67pmol/l. A relaxa- tion of the stomach is reflected as a downward deflection of the gastric volume recording.
leaving a 5-10 cm long colonic segment for the experiment. The colonic vein was cannulated and blood flow was recorded continuously by a drop recorder unit operating an ordinate writer, as described for the stomach. Autonomic nervous influences on the colon were eliminated by cutting the pelvic nerves and the nerves around the inferior mesenteric artery.
Colonic motility was followed by recording intraluminal volume changes with a volume transducer connected to one end of the colonic segment, the other end being closed with a ligature. Intraluminal pres- sure was set at 2 4 cm H,O.
Close intraarterial infusions of VIP were made into a side-branch to the colonic artery.
Solutions and drugs
The intravascularly infused buffer solution contained 10 mmol NaHCO, in 100 ml a 10% glucose solution (550 mmol/l). It was infused at a rate of 0.1-0.2 ml/min.
The solution used to perfuse the lumen of the small intestine contained (mmol/l): NaCl 122; KCI 3.5; NaHCO, 25; KH,PO, 1.2; MgCI, x 6H,O 1.2; CaCI, 2.5; glucose 30. The osmolality of the solution was 310-315 mOsm/kg H 2 0 .
VIP was dissolved in physiologic saline at various concentrations. The infusion rates used are given in Results. VIP was obtained from dr V. Mutt at the Gastrointestinal Research Unit, Karolinska Institutet, Stockholm, Sweden.
Atropine (0.5-1 mg/kg b.wt.) was administered i.v. during the course of the experiments.
Calculations
The increase of arterial plasma VIP concentration produced by the i.a. infusion of the drug was calculated by dividing the amount infused (,ug/min) by the plasma flow (ml/min) as measured when the infusion was started. This may represent a slight overestimation of the increase of VIP concentration since blood flow often increased upon infusion. However, recirculation tends to increase plasma drug concentration during prolonged infusions.
Results Experiments on the stomach
In 3 expts. VIP was infused close i.a. to the stomach, usually during a 3 min period, and at rates between 2 and 12 nmol/min (7-42 pg/min) which led to plasma VIP concentrations between 0.2 and 3.0 pmol/l. Changes in total gastric blood flow and gastric volume were recorded. Fig. 1 illustrates a typical experiment. Upon infusing VIP a small increase of

464 STEFAN EKLUND, MATS JODAL, OVE LUNDGREN AND ANDERS SJOQVIST
ARTERIAL BLOOD PRESSURE, rnrn Hg 5o
CHANGE OFGASTRIC rnl ] VOLUME
Fig. 2. The effects of distending the esophagus on arterial blood pressurd, gastric blood flow and gastric motility. The experiment
animal as illustrated in Fig. I . Esophageal distension
GASTRIC BLOOD FLOW,rnl/rnin x lOOg
TIME,rnin I. I I
- was performed on the same
0 5 10 15
gastric blood flow was observed concomitant to a pronounced increase in gastric volume, depicted as a downward deflection in the recording. Blood flow increased somewhat during the period of constant VIP infusion and returned to control fairly quickly after the infusion. The changes in gastric volume exhibited a characteristic, fairly fast increase during the period of drug infusion, while the return to control volume was quite slow, lasting 20-30 min. A closely similar change of gastric volume could be elicited reflexly by distending a balloon in the esophagus, as illustrated in Fig. 2 taken from the same expt. No significant change in gastric blood flow was observed upon esophageal distension.
In 2 expts. an attempt was made to estimate the increase of arterial plasma VIP con- centration necessary to produce a gastric relaxation of the same magnitude as that elicited by distension. It was found to be 0.3 and 0.5 ,umol/l plasma.
The motility effects observed upon VIP infusions were similar in magnitude before and after giving atropine to the animal.
Experiments on the small intestine
The effects of intraarterial VIP infusions on blood flow and net water uptake in the small intestine were studied in 7 cats. VIP was infused at a rate of 0.001-7.1 nmol/min (0.003-24 ,ug/min) increasing the plasma VIP concentration 0.4-1 900 nmol/l. The high infusion rates were usually maintained for only 3 min to study the VIP effects on blood flow. At low rate infusions the experimental runs were prolonged for 10-15 niin to allow recordings of VIP effects on net fluid transport rate in the intestine. Plasma concentrations of VIP were cal- culated without taking into consideration recirculation of the peptide (see Methods). Fig. 3 illustrates a representative experiment, showing that blood flow is slightly increased upon VIP infusion while the rate of intestinal water uptake is reduced as indicated by dotted lines.
The lowest increase of arterial plasma VIP concentration able to decrease intestinal net fluid uptake was 2.0 nmol/l, which did not significantly increase total intestinal blood flow. At high rates of infusion, increasing plasma VIP concentrations above 0.5 ,umol/l, a net intestinal secretion was observed, continuing also after ended infusion with a slow return to control rate of absorption first about 30 min after the infusion period. With regard to

VIP AND GI FUNCTION 465
INTESTINAL 25 -___ W D R D W . 50 ml/min x lOOg
VIP-infusion TIME,mln I - 1 1 I I
NET RATE OF WATER I 1 1 I f I
ABSORPTION. 0.500 0.285 0.500 ml/mhxKX)cm* se4u!3dwrfocs
0 5 10 15 20 25 x) 35
Fig. 3. The effects of a close infusion of VIP on arterial blood pressure intestinal blood flow and intestinal net water transport. The drug was infused at a rate of 0.017 nmol/min increasing plasma concentration 4.9 nmol/l. The upward deflections of the recording of perfusate volume were produced by adding 1 rnl perfusate to the volume transducer.
intestinal vasodilatation it was calculated that jejunal blood flow increased 40-50 % above control when increasing arterial plasma VIP concentrations between 0.1 5 and 0.45 ,umol/l.
Atropine administration had no appreciable effects on the intestinal responses to VIP.
Experiments on the colon
In 5 expts. VIP was infused i.a. to the colon while colonic blood flow and motility were followed. The infusions were performed at rates varying between 1.7 and 7.1 nmol/min (5.8-24.0 ,ug/min), increasing plasma VIP concentrations between 0.35 and 4.6 ,umol/l. A representative expt. is shown in Fig. 4. Upon starting the VIP infusion a marked blood flow increase immediately ensued, but subsided after about half a minute and levelled off at a moderate flow elevation maintained throughout the infusion. After about 2 min a contraction of the colonic musculars occurred, reflected as an increased fluid volume in the volume transducer. When the VIP infusion was stopped blood flow returned rapidly to control while the colonic volume increase returned to control first 5-10 min later.
Fig. 4. The effects of a close i.a. infusion of VIP on arterial blood pressure, colonic blood flow and colonic motility. VIP was in- fused at a rate of 3.4 nmol/rnin increasing plasma concentration 0.72 prnol/l. A contraction of the colon is reflected as an upward deflection of the colonic volume recording.
ARTERIAL BLOOD 150 PRESSURE,mm Hg 3
50
CHANGE OF COLONIC VOLUME, ml
0
COLONIC BLOOD FLOW,ml/min x 1OOg
TIME ,min VIP- infusion
0 5 10 15 I I I

466 STEFAN EKLUND, MATS JODAL, OVE LUNDGREN AND ANDERS SJOQVIST
To quantitate the VIP effects on colonic blood flow the increase of arterial plasma VIP concentration needed to produce as initial doubling of blood flow (Fig. 4) was determined. It varied between 1 and 4 pmol/l plasma.
Atropine had no effect on the VIP induced changes in colonic blood blow and motility. In 3 of the 5 colon expts. isoprenaline and papaverin were infused i.a. in the same way as for VIP. Vasodilatations of the same magnitude as with VIP were then produced but colonic contractions were never observed; if anything, minor relaxations occurred.
Discussion
In the present study the effects of i.a. infused VIP on various gastrointestinal functions have been investigated. In the stomach it was possible to demonstrate that VIP relaxed both the vascular smooth muscle cells and the gastric muscularis propria. Relaxatory effects of VIP on gastric smooth muscle has been reported in vifro (Piper, Said and Vane 1970). In contrast to the vascular effects the gastric relaxatory response to VIP under in vivo conditions was very longlasting returning to control first 20-40 min after ended infusion. This response closely mimics the receptive relaxation elicited by direct stimulation of the efferent vagal ‘relaxatory’ fibres (Martinson 1965) or by their reflex activation by way of distending the esophagus (Abrahamsson and Jansson 1969; Fig. 2). In the small intestine VIP induced a vasodilatation and a decrease of net water transport. At high VIP concentration intestinal secretion was recorded. These observations are in agreement with earlier reports (Blood flow: Said and Mutt 1970, Thulin and Olsson 1973, Kachelhoffer et al. 1974; Net water transport: Berbezat and Grossman 1971, Krejs et al. 1978). Finally, in the colon a hyperemia and a delayed contraction was recorded upon VIP infusion. These two colonic responses are similar to those induced by pelvic nerve stimulation in animals given atropine (Hulten and Jodal 1969).
In an earlier report from this laboratory (Fahrenkrug et al. 1978 b) it was shown that VIP was released from the gastrointestinal tract when eliciting three different non-adrener- gic, non-cholinergic neurogenic responses, i.e. the vagally mediated gastric receptive relaxa- tion (Martinson 1965), the intestinal vasodilatation produced by mechanical stimulation of the intestinal mucosa (Biber et al. 1971) and the colonic hyperemia evoked by the pelvic nerves (HultBn ef al. 1969). From these and other observations it was proposed that VIP is a neurotransmitter in the gastrointestinal tract. The present results strengthen this con- clusion by showing that the intravascular infusion of the proposed neurotransmitter evokes physiological effects closely similar to those seen when the mentioned non-adrenergic, non-cholinergic nervous mechanisms of the alimentary canal are activated.
In the study by Fahrenkrug et al. (1978 b) the increase of venous plasma VIP concentra- tions, accompanying the induction of the different nervous effects, were determined. In the present study the arterial VIP concentrations necessary for producing the same gastro- intestinal effects were estimated. A comparison of the plasma VIP concentrations in the two experimental situations reveals that the nervous release of VIP into the blood stream was only 1/10 000 of the arterial VIP concentrations needed to induce quantitatively similar physiological effects upon intraarterial VIP administration. This large difference between

VIP AND GI FUNCTION 467
plasma concentrations is, indeed, to be expected if VIP is a neurotransmitter in the gastro- intestinal tract. VIP is a fairly large, water soluble compound suggesting that its capillary permeability is fairly low (Landis and Pappenheimer 1963). Thus VIP, when released topi- cally from nerve endings in the tissues, will initially show a much restricted tissue distribu- tion. The compound will then be locally dispersed by diffusion and probably only to a small extent reach the blood stream due to the hindrance offered by the capillary walls. Further- more, potent mechanisms are likely to exist at synaptic clefts to degrade the VIP molecule and/or bring it back into the nerve endings. Similarly, the low capillary permeability of VIP necessitates a high rate of intraarterial infusion of the compound to reach the same VIP concentration at the effector cell as that induced by the nervous release. Such principles hold, for instance, for the blood concentration of neurogenically released noradrenaline and intraarterially infused noradrenaline when given in concentration to match the neuro- genic effects (Ljung 1970).
None of the nervous effects described above were studied with regard to their effects on transepithelial fluid transport. In the present study it was observed that one could induce a decrease of net fluid absorption in the small intestine at plasma VIP concentrations well below those inducing a vasodilatation. In fact, the increase of arterial plasma VIP concentra- tion that diminished net fluid uptake was only 5-10 times greater than the increase of venous plasma concentration observed upon mechanical stimulation of the intestinal mucosa (Fahrenkrug et af. 1978 b). It seems therefore reasonable to conclude that VIP profoundly affects intestinal net fluid transport rate at very low plasma concentrations, possibly even at such plasma concentrations seen during physiological circumstances. This conclusion is corroborated by the findings reported from studies on isolated intestinal epithelial cells indicating the presence of membrane receptors with a very high affinity to VIP (Dupont et af. 1978, Laburthe et af. 1978).
This research was supported by grants from the Swedish Medical Research Council (14X-2855), from the Swedish Society for Medical Sciences, from Harald and Greta Jeanssons Fund, from Wilhelm and Martha Lundgrens Fund, from Magnus Bergvalls Stiftelse and from the Faculty of Medicine, University of Gote- borg.
References ABRAHAMSSON, H. and G . JANSSON, Elicitation of reflex vagal relaxation of the stomach from pharynx
and esophagus in the cat. Acra physiol. scand. 1969. 77. 172-178. BARBEZAT, G. 0. and M. 0. GROSSMAN, Intestinal secretion stimulation by peptides. Science 1971. 174.
422-423. B I B E R , B., 0. LUNDGREN and J. SVANVIK, Studies on the intestinal vasodilatation observed after mechanical
stimulation of the mucosa of the gut. Acra physiol. scand. 1971. 82. 177-190. DUPONT, C., M. LABURTHE and G. ROSSELIN, Isolation of epithelial cells of human colon and denionstra-
tion of the action of VIP. Scand. J. Gastroenterol. 1978. 13. Suppl. 49: 51. EMSON, P. C., J . FAHRENKRUG, 0. B. SCHAFFALITZKY DE MUCKADELL, T. M. JESSELL and L. L. IVERSEN,
Vasoactive intestinal polypeptide (VIP): vesicular localization and potassium evoked release from rat hypothalamus. Brain Res. 1978. 143. 174-178.
FAHRENKRUG, J . , H. GALBO, J. J. HOLST and 0. B. SCHAFFALITZKY DE MUCKADELL, Influence of the auto- nomic nervous system on the release of vasoactive intestinal polypeptide from the porcine gastrointestinal tract. J . Physiol. (Lond.) 1978. 280. 405422.
FAHRENKRUG, J. , U. HAGLUND, M. JODAL, 0. LUNDGREN, L. OLBE ana 0. B. SCHAFFALITZKY DE MUCKA- DELL, Nervous release of vasoactive intestinal polypeptide in the gastrointestinal tract of cats: possible physiological implications. J . Physiol. (Lond.) In press.

468 STEFAN EKLUND, MATS JODAL, OVE LUNDGREN AND ANDERS SJOQVIST
GiAcHETTI, A,, F. KONIGES and S. I . SAID, VIP localization in synoptosomes of rat brain. Fed. Proc. 1977. 36. 951.
HAGLUND, U. and 0. LUNDGREN, Reactions within consecutive vascular sections of the small intestine of the cat during prolonged hypotension. Acfa physiol. scand. 1972. 84. 151-163.
H U L T ~ N , L. and M. JODAL, Extrinsic nervous control of colonic motility. Acta physiol. scand. 1969. Suppl. 335: 21-38.
HULTEN, L., M. JODAL and 0. LUNDGREN, Extrinsic nervous control of colonic blood flow. Acta physiol. scond. 1969. Suppl. 335: 39-50.
JODAL, M., D.-A. HALLBACK, J. SVANVIK and 0. LUNDGREN, A method for the continuous study of net water transport in the feline small bowel. Acta phvsiol. scand. 1975. 95. 441-447.
KACHELHOFFER, J., M. R. ELOY, A. PoirssE, D. HOHMATTER and J. F. GRENiER, Mesenteric vasomotor effects of vasoactive intestinal polypeptide study on perfused isolated canine jejunal loops. Pflug. Arch. ges. Physiol. 1974. 352. 3746.
KREJS, G. J., R. M. BARKLEY, N. W. READ and 1. S. FORDTRAN, Intestinal secretion induced by vasoactive intestinal polypeptide. J . din. Inuest. 1978. 61. 1337-1345.
LABURTHE, M., M. TOUSSET, C. BOISSARD, G . CHEVALIER, A. ZwEieAuM and G. ROSSELIN, Vasoactive intestinal peptide (VIP) stimulates cyclic AMP accumulation in human malignant intestinal cell lines in cultures. Srand. J. GastrornteroL 1978. 13. Suppl. 49: 107.
LARSSON, L.-I., Ultrastructural localization of a new neuronal peptide (VIP). Hisrochem. 1977. 54. 173-176. LARSSON, L.-I., J. FAHRENKRUG, 0. SCHAFFALITZKY DE MUCKADELL, F. SUNDLER, R. HAKANSSON and
J. F. REHFELD, Localization of vasoactive intestinal polypeptide (VIP) to central and peripheral neurons. Proc. nut. Acurl. Sci. 1976. 73. 3197-3200.
LJUNG, B., Nervous and myogenic mechanisms in the control of a vascular neuroeffector system. An integrated survey of a series of studies on the portal vein. Acta physiul. scand. 1970. Suppl. 349. 33-68.
MARTINSON, J., Studies on the efferent vagal control of the stomach. Acta phvsiol. scand. 1965. 65. Suppl. 255.
PIPER, P. J., S. 1. SAID and J. R. VANE, Effects on smooth muscle preparations of unidentified vasoactive peptides from intestine and lung. Nature (Lond.) 1970. 225. 1144-1 146.
SAID, S. I. and V. MUTT, Polypeptide with broad biological activity: Isolation from small intestine. Science 1970. 169. 1217-1218.
SAID, S . I. and V. MUTT, Potent peripheral and splanchnic vasodilator peptide from normal gut. Nature (Lond.) 1970. 225. 863-864.
THULIN, L. and P. OLSSON, Effects of intestinal peptide mixture G2 and vasoactive intestinal peptide VIP on splanchnic circulation in the dog. A d a chir. scantl. 1973. 139. 691-697.