Embed Size (px)
Citation preview

Chemosphere 84 (2011) 657–663
Contents lists available at ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Effects of the pharmaceuticals gemfibrozil and diclofenac on biomarkerexpression in the zebra mussel (Dreissena polymorpha) and their comparisonwith standardised toxicity tests
Brian Quinn a,⇑, Wiebke Schmidt a, Kathleen O’Rourke b, Robert Hernan b
a Irish Centre for Environmental Toxicology, Galway-Mayo Institute of Technology, Dublin Road, Galway, Irelandb Shannon Aquatic Toxicity Laboratory, Enterprise Ireland, Shannon, Co. Clare, Ireland
a r t i c l e i n f o a b s t r a c t
Article history:Received 14 January 2011Received in revised form 7 March 2011Accepted 19 March 2011Available online 13 April 2011
Keywords:GemfibrozilDiclofenacZebra musselBiomarkerToxicity
0045-6535/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.chemosphere.2011.03.033
⇑ Corresponding author. Tel.: +353 91 742515; fax:E-mail address: [email protected] (B. Quinn).
Pharmaceuticals, including the lipid regulator gemfibrozil and the non-steroidal anti-inflammatory drugdiclofenac have been identified in waste water treatment plant effluents and receiving waters throughoutthe western world. The acute and chronic toxicity of these compounds was assessed for three freshwaterspecies (Daphnia magna, Pseudokirchneriella subcapitata, Lemna minor) using standardised toxicity testswith toxicity found in the non-environmentally relevant mid mg L�1 concentration range. For the acuteendpoints (IC50 and EC50) gemfibrozil showed higher toxicity ranging from 29 to 59 mg L�1 (diclofenac47–67 mg L�1), while diclofenac was more toxic for the chronic D. magna 21 d endpoints ranging from10 to 56 mg L�1 (gemfibrozil 32–100 mg L�1). These results were compared with the expression of sev-eral biomarkers in the zebra mussel (Dreissena polymorpha) 24 and 96 h after exposure by injection toconcentrations of 21 and 21,000 lg L�1 corresponding to nominal concentrations of 1 and 1000 lg L�1.Exposure to gemfibrozil and diclofenac at both concentrations significantly increased the level of lipidperoxidation, a biomarker of damage. At the elevated nominal concentration of 1000 lg L�1 the biomark-ers of defence glutathione transferase and metallothionein were significantly elevated for gemfibrozil anddiclofenac respectively, as was DNA damage after 96 h exposure to gemfibrozil. No evidence of endocrinedisruption was observed using the alkali-labile phosphate technique. Results from this suite of biomark-ers indicate these compounds can cause significant stress at environmentally relevant concentrations act-ing primarily through oxidation pathways with significant destabilization of the lysosomal membraneand that biomarker expression is a more sensitive endpoint than standardised toxicity tests.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Pharmaceuticals are one of the number of emerging environ-mental contaminants receiving growing attention within the sci-entific literature. The data presently available confirm anenvironmental risk for both human and veterinary pharmaceuti-cals, with the ecotoxicity data for pharmaceuticals proven to bein the range of measured environmental concentrations (EEA,2010). Pharmaceuticals represent a diverse class of compoundsthat have several features in common: biologically active by de-sign; specific mode of action necessary to carry out their therapeu-tic action; relatively persistent and resistant to inactivation ordegradation; produced and consumed in vast quantities annually(Crane et al., 2006; Fent et al., 2006; Santos et al., 2010). Humanpharmaceuticals are principally introduced into the aquatic envi-
ll rights reserved.
+353 91 742500.
ronment via treated municipal effluent (Fent et al., 2006) wherethey can be found in both effluents and receiving environmentsat concentrations in the low lg L�1 to high ng L�1 range (Daughtonand Ternes, 1999; Metcalfe et al., 2003; Gomez et al., 2007). Envi-ronmental risk assessment (ERA) studies on pharmaceuticals showthat 95% were not readily biodegradable according to the OECDtest 301 (EEA, 2010). It has been highlighted (Santos et al., 2010)that throughout the literature there is a clear predominance ofstudies on the non-steroidal anti-inflammatory drugs (NSAIDs)and blood lipid lowering agents, as these are among the most com-monly found drugs in the aquatic environment.
NSAIDS act by inhibiting one or both isoforms of the cyclooxy-genase enzymes COX-1 or COX-2, involved in the synthesis of pros-taglandins (Gagné et al., 2005). Diclofenac is thought to be themost toxic member of this group who’s effect is known to occurby damaging renal and gastrointestinal tissue across several verte-brate taxa (Haap et al., 2008) and who’s accidental exposure has re-sulted in the near extinction of Asian vulture populations (Oakset al., 2004). For this reason diclofenac was highlighted by the

658 B. Quinn et al. / Chemosphere 84 (2011) 657–663
European Environment Agency (EEA, 2010) and several authors(Fent et al., 2006; Letzel et al., 2009) as being of particular environ-mental concern. Expression of a cyclooxygenase enzyme similar tohuman COX-2 has been found in fish (Santos et al., 2010) and hasbeen investigated in mollusc bivalves (Gagné et al., 2008). Severalstudies demonstrate that diclofenac has a low acute toxicity, asmeasured EC50 concentrations are much higher than measuredconcentrations in the environment (Santos et al., 2010). Howeverthe PEC for a realistic worst case scenario of 140 ng L�1 was calcu-lated by Letzel et al. (2009), a concentration that has been observedin surface waters and is close to the LOEC of 1–5 lg L�1 for rainbowtrout (Schwaiger et al., 2004; Triebskorn et al., 2004) and NOEC of0.5 lg L�1 for brown trout (Hoeger et al., 2005) indicating the po-tential for this drug to cause real concern in the environment.
Another group of pharmaceuticals causing concern are theblood lipid lowering agents. Within this group the fibrates act bybinding to nuclear peroxisome proliferators-activated receptors(PPAR a, b and c) and regulating the expression of several lipid reg-ulatory proteins (Staels et al., 1998). Fibrates have been shown tostimulate fatty acid uptake, conversion of acetyl-CoA derivativesand catabolism by the beta oxidation pathway (Staels et al.,1998) and are also involved in glycolysis, glutathione metabolism,redox balance and inflammatory processes (Canesi et al., 2007).Gemfibrozil has been identified as a member of this group com-monly found in the environment but with relatively little toxicitydata. Gemfibrozil induced antioxidant defence enzymes includingglutathione peroxidise and glutathione-S-transferase (GST) ingoldfish (Carassius auratus) (Mimeault et al., 2006) and can biocon-centrate in blood and reduce plasma testosterone levels (Mimeaultet al., 2005).
It has been observed by several authors (Crane et al., 2006; Fentet al., 2006; Santos et al., 2010) that most published aquatic toxic-ity data and risk assessment for pharmaceuticals are based onshort term acute studies, despite the recognition that exposure ismost likely to be continuous at low concentrations of effluent fromSTW. It is for this reason that in the current study we have decidedto investigate both the acute and chronic effects of gemfibrozil anddiclofenac in standardised toxicity tests using the freshwaterorganisms Pseudokirchneriella subcapitata, Lemna minor and Daph-nia magna and to compare these results with a biomarker assess-ment in the zebra mussel (Dreissena polymorpha). Bivalvesexhibit many of the basic physiological characteristics found invertebrates, contributing to their promotion as potential surro-gates to vertebrates for toxicity testing. Bivalves show a capacityto biotransform organic chemicals through phase I and phase IIreactions (Livingstone, 1998) necessary for the elimination by bio-transformation of pharmaceuticals. Bivalves also represent sensi-tive organisms for the action of fibrates in the environment(Canesi et al., 2007) and have shown reduced COX activity afterexposure to the NSAID ibuprofen (Gagné et al., 2005). Zebra mus-sels have been used to investigate the effects of organic com-pounds including endocrine disrupting compounds (Quinn et al.,2004, 2006) and pharmaceuticals (Binelli et al., 2009; Paroliniet al., 2010).
In the present study biomarkers of defence (glutathione trans-ferase (GST) and metallothionein (MT)) were investigated as theGST enzyme is important in drug conjugation/detoxification andMT is a marker of oxidative stress and is involved in scavengingreaction oxygen species (ROS). The biomarker of damage lipid per-oxidation (LPO) is associated with oxidative stress resulting in theoxidation of polyunsaturated lipids and could be a downstream ef-fect of both COX-2 inhibition or peroxisome proliferators where b-oxidation occurs. Although DNA strand breaks are indicative ofgenetic damage they have also been associated with oxidativedamage. The potential endocrine disrupting effects was investi-gated using alkali-labile phosphate (ALP) technique to measure
vitellin-like proteins as gemfibrozil and diclofenac have been re-ported to affect testosterone (Mimeault et al., 2005) and vitello-genin (Hong et al., 2007) levels respectively in fish. Thesebiomarkers were investigated in the zebra mussel and comparedto standardised toxicity test.
2. Materials and methods
2.1. Sampling
Zebra mussels, D. polymorpha, were collected in April 2009 nearKillinure Point Marina, Glasson, Co., Westmeath, Ireland on LoughRee (53�2705400N 7�5502000W) and acclimatised for 3 d in de-chlorinated tap water at 10 �C. In two separate exposuresgemfibrozil and diclofenac was injected into the posterior adductormuscle of groups of 14 mussels using a sterile 50 lL microsyringewith a 22G needle. Taking into account the average volume ofhemolymph that can be withdrawn from zebra mussels of this size(2–3 cm) is �200 lL (Byrne and Dietz, 2006) each mussel was in-jected with 10 lL of gemfibrozil or diclofenac dissolved in DMSOto give a nominal concentration of 1000 lg L�1 (from a 2.1 mg/100 mL DMSO stock solution taking into account a dilution factorof 21) and 1 lg L�1 (initial stock diluted 1/1000) as per Canesiet al., 2007. Exposures were performed in triplicate with controland solvent control animals being injected with 10 lL of de-chlori-nated water and solvent (DMSO) respectively. After injection mus-sels were left stand for 10 min and were placed in glass tankscontaining 10 L of de-chlorinated tap water at an average temp of6.9 and 13 �C for gemfibrozil and diclofenac exposures respectively,with the water in each tank changed every 24 h. After 24 and 96 hthree mussels were randomly selected from each of the three expo-sure and control tanks (n = 9). Each animal was measured (shelllength in centimetres) and weighed (whole animal weight ingrams) to determine its condition factor (CF) (Fultons condition fac-tor = W/L3). After removal of the byssal thread the visceral masswas dissected and immediately frozen at�80 �C for later biomarkeranalysis.
2.2. Tissue preparation and biochemical analysis
The dissected visceral mass was allowed to thaw on ice. Eachmussel was sexed using the squash technique and thoroughlyhomogenized in a 10 mM Hepes-NaOH buffer containing 100 mMNaCl, 0.1 mM dithiothreitol, 0.1 mM EDTA, pH 7.4 at 4 �C at a 1:5weight:volume ratio. These homogenised samples were frozen at�80 �C for analysis of DNA damage, lipid peroxidation (LPO) andtotal protein determination (Bradford, 1976). The remaininghomogenate was centrifuged at 12,000g for 30 min at 4 �C. Thesupernatant (S12) was aliquoted and frozen for later analysis ofglutathione transferase activity (Livingstone), metallothionein(MT) and alkali-labile phosphate (ALP). These biomarkers werealso normalised against the total protein content in the superna-tant (Bradford, 1976). A description of the techniques used is givenbelow. All techniques were adapted from those previously de-scribed by (Blaise et al., 2002).
2.3. Glutathione transferase activity (GST)
A sample of 50 lL of supernatant (S12) was added to 200 lL of1 mM GSH and 1 mM 1-chloro-2,4-dinitrobenzene in a buffer of10 mM Hepes-NaOH, pH 6.5 containing 125 mM NaCl (based onthe method developed by (Boryslawskyj et al., 1988)). The rate ofincrease of absorbance based on the appearance of the glutathioneconjugate was measured at 340 nm at time 0, 5, 10, 20 and 30 min.A blank substituting homogenization buffer for S12 was included.

B. Quinn et al. / Chemosphere 84 (2011) 657–663 659
Enzyme activity was calculated by the formation of product/(min �mg protein) and expressed as lmoles mg�1 protein.
2.4. Metallothionein (MT)
MT was measured using an adapted spectrophotometric meth-od developed by (Viarengo et al., 1997). A 500 lL S12 sample wasmixed with 500 lL of ethanol (95%) containing 8% chloroform andcentrifuged at 6000g for 10 m in at 2 �C. A 700 lL subsample ofsupernatant (S6) was mixed with 50 lL RNA (1 mg mL�1), 10 lLHCl 6 M and 1.2 mL of cold (4 �C) ethanol, mixed by inversionand placed at �80 �C for 25 min. This mixture was centrifuge at6000g for 10 min at 2 �C. The supernatant was removed and thepellet re-suspended with 87% ethanol:1% chloroform (300 lL)and centrifuged a second time at 6000g for 1 min at 2 �C. Afterthe removal of the supernatant, the pellet was re-suspended with150 lL of 0.25 M NaCl and 150 lL HCl 0.2 M containing 4 mMEDTA. Three hundred microliter of Ellman’s reagent (dith-ionitrobenzoate 0.4 mM, 2 M Nacl and 0.2 M Tris base, pH 8) wasadded, and absorbance of a 250 lL sample in a 96 well plate mea-sured at 412 nm after 5–10 min. A blank (re-suspension buffer)and standards of Glutathione (GSH) were included in each runand results expressed as nanomoles of thiols or MT-equivalentsper milligram of homogenate supernatant proteins.
2.5. DNA damage (strand breaks)
DNA strand breaks were quantified using a fluorescence tech-nique adapted from the alkaline precipitation assay (Olive, 1988).A 25 lL homogenate sample was mixed with 200 lL of 2% SDS con-taining 10 mM EDTA, 10 mM tris-base and 40 mM NaOH for 1 min.Two hundred microliter of 0.12 M KCl was added and the solutionheated at 60 �C for 10 min, mixed by inversion and cooled at 4 �Cfor 30 min. This mixture was then centrifuged at 8000g for 5 minat 4 �C. Fifty microliter of the supernatant was added to 150 lLof Hoescht dye (1 lg mL�1 diluted with buffer containing 0.4 MNaCl, 4 mM sodium cholate and 0.1 M Tris-acetate, pH 8.5–9) ina 96 well plate and mixed for 5 min. Fluorescence was measuredusing 360 nm (excitation) and 450 nm (emission) filters againstblanks containing identical constituents, with 25 lL homogenisa-tion (Hepes) buffer replacing the homogenate. Salmon sperm geno-mic DNA standards (Sigma) were added for DNA calibration andthe results expressed as DNA lg mg�1 protein.
2.6. Lipid peroxidation (LPO)
Lipid peroxidation was measured by the production of malonal-dehyde in the mussel homogenate by the thiobarbituric acid meth-od (Wills, 1987). In a 1.5 mL eppendorf tube, 150 lL homogenatewas mixed with 300 lL of 10% TCA containing 1 mM FeSO4 and150 lL of 0.67% thiobarbituric acid, heated to 70–80 �C for10 min and centrifuged (10,000g for 10 s) to remove the precipi-tate. Two hundred microliter was added to a 96 well plate andthe fluorescence measured at 516 excitation/600 nm emission.Blanks and standards of tetramethoxypropane were prepared inhomogenization (Hepes) buffer. Results were expressed as lmolesof thiobarbituric acid reactants per milligram of homogenateprotein.
2.7. Vitellin-like proteins
The vitellogenic (Vn) properties of the S12 supernatant weremeasured using the indirect alkali-labile phosphate (ALP) tech-nique (Blaise et al., 1999). A 100 lL sample was mixed with54 lL acetone (35% final concentration) for 5–10 min and centri-fuged 10,000g for 5 min. The supernatant was removed and the
pellet dissolved in 50 lL 1 M NaOH and mixed for 30 min at60 �C. Total phosphate was measured using a colorimetric phos-phomolybenum method (Stanton, 1968). To a 20 lL sample125 lL H2O, 5 lL 100% TCA, 25 lL of molybdate reactive and25 lL ascorbate 1% were added, mixed for 10 min and using a spec-trophotometer absorbance read at 815 nm and 444 nm. Rainbowtrout vitellogenin was used as a positive control for the assay withblanks having NaOH substituted for the sample. Vn levels were ex-pressed as lmoles of alkali-labile phosphate (ALP) per milligram ofsupernatant protein.
2.8. Standardised toxicity tests
D. magna acute & chronic toxicity tests used were ISO6341:1996/Cor.1:1998: ‘Water quality – Determination of the inhi-bition of the mobility of D. magna Straus (Cladocera, Crustacea)’and BS 6068-5.28:2000; ISO 10706:2000: ‘Water quality – Deter-mination of long term toxicity of substances to D. magna Straus’tests respectively. The freshwater algae test was ISO 8692:2004:‘Water quality – Freshwater algal growth inhibition test with uni-cellular green algae’ (P. subcapitata) and freshwater Plant toxicitytest ISO 20079:2005: ‘Water quality – Determination of the toxiceffect of water constituents and waste water to duckweed (L. min-or) – Duckweed growth inhibition test’. Solvent 0.2% DMSO wasused at each concentration and control.
2.9. Statistical treatment
Three mussels were taken per exposure tank at 24 h and 96 hfor biomarker analysis (n = 9 for each time period). Data are ex-pressed as the mean (n = 9) ± the standard error. Data normalityand distribution (homogeneity of variances) was tested using theShapiro-Wilks and Levene‘s tests respectively. Where normalityand equal variance was demonstrated analysis of variance was per-formed and critical differences between groups appraised usingthe Bonferroni t-test. Following transformation if data were foundto fail one or other of these tests and were found to be not normallydistributed or nonhomogeneous and subsequently nonparametricanalysis of variance was performed using Kruskal–Wallis oneway ANOVA on ranks to investigate biomarker expression for eachtime period. The relationships between biomarker expression foreach drug was investigated using Spearman rank correlation. Allstatistics were undertaken using Statistica (Version 6). The methodof calculation used for each of the standardised toxicity tests isshown in Table 2.
3. Results
Mussels were found to be in good condition after injection andthroughout the exposure, with a low mortality rate (3 out of 156mussels exposed dying (1.9%)) observed. The condition factor (totalweight � length3) remained unchanged between exposure concen-trations and time for both compounds (Fig. 1f), with little change incondition seen throughout the exposures. Mussels were generallywell attached to the substrate and appeared to be in good condi-tion. Various parameters such as temperature, pH, oxygen, NO2,NO3 and NHþ4 were measured during the 96 h exposure. Slightchanges were observed, but all were within the required measure-ments (data not shown). The only anomaly observed occurred dur-ing the gemfibrozil exposure when the temperature increased by7.35 �C from 72 h to 96 h. However after analysis between temper-ature and each biomarker using Mann–Whitney rank sum test andKruskal–Wallis ANOVA using temperature as a factor, no effect wasseen.
(b)(a)
(c)
(e) (f)
(d)
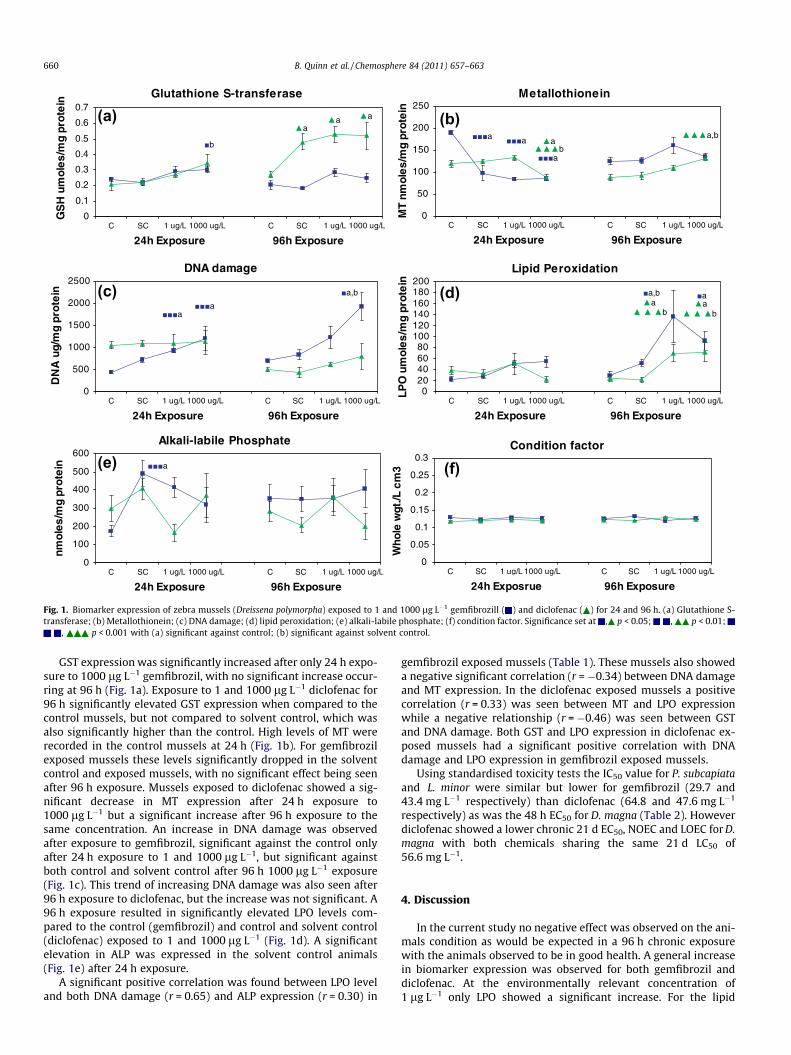
Fig. 1. Biomarker expression of zebra mussels (Dreissena polymorpha) exposed to 1 and 1000 lg L�1 gemfibrozill ( ) and diclofenac ( ) for 24 and 96 h. (a) Glutathione S-transferase; (b) Metallothionein; (c) DNA damage; (d) lipid peroxidation; (e) alkali-labile phosphate; (f) condition factor. Significance set at , p < 0.05; , p < 0.01;
, p < 0.001 with (a) significant against control; (b) significant against solvent control.
660 B. Quinn et al. / Chemosphere 84 (2011) 657–663
GST expression was significantly increased after only 24 h expo-sure to 1000 lg L�1 gemfibrozil, with no significant increase occur-ring at 96 h (Fig. 1a). Exposure to 1 and 1000 lg L�1 diclofenac for96 h significantly elevated GST expression when compared to thecontrol mussels, but not compared to solvent control, which wasalso significantly higher than the control. High levels of MT wererecorded in the control mussels at 24 h (Fig. 1b). For gemfibrozilexposed mussels these levels significantly dropped in the solventcontrol and exposed mussels, with no significant effect being seenafter 96 h exposure. Mussels exposed to diclofenac showed a sig-nificant decrease in MT expression after 24 h exposure to1000 lg L�1 but a significant increase after 96 h exposure to thesame concentration. An increase in DNA damage was observedafter exposure to gemfibrozil, significant against the control onlyafter 24 h exposure to 1 and 1000 lg L�1, but significant againstboth control and solvent control after 96 h 1000 lg L�1 exposure(Fig. 1c). This trend of increasing DNA damage was also seen after96 h exposure to diclofenac, but the increase was not significant. A96 h exposure resulted in significantly elevated LPO levels com-pared to the control (gemfibrozil) and control and solvent control(diclofenac) exposed to 1 and 1000 lg L�1 (Fig. 1d). A significantelevation in ALP was expressed in the solvent control animals(Fig. 1e) after 24 h exposure.
A significant positive correlation was found between LPO leveland both DNA damage (r = 0.65) and ALP expression (r = 0.30) in
gemfibrozil exposed mussels (Table 1). These mussels also showeda negative significant correlation (r = �0.34) between DNA damageand MT expression. In the diclofenac exposed mussels a positivecorrelation (r = 0.33) was seen between MT and LPO expressionwhile a negative relationship (r = �0.46) was seen between GSTand DNA damage. Both GST and LPO expression in diclofenac ex-posed mussels had a significant positive correlation with DNAdamage and LPO expression in gemfibrozil exposed mussels.
Using standardised toxicity tests the IC50 value for P. subcapiataand L. minor were similar but lower for gemfibrozil (29.7 and43.4 mg L�1 respectively) than diclofenac (64.8 and 47.6 mg L�1
respectively) as was the 48 h EC50 for D. magna (Table 2). Howeverdiclofenac showed a lower chronic 21 d EC50, NOEC and LOEC for D.magna with both chemicals sharing the same 21 d LC50 of56.6 mg L�1.
4. Discussion
In the current study no negative effect was observed on the ani-mals condition as would be expected in a 96 h chronic exposurewith the animals observed to be in good health. A general increasein biomarker expression was observed for both gemfibrozil anddiclofenac. At the environmentally relevant concentration of1 lg L�1 only LPO showed a significant increase. For the lipid
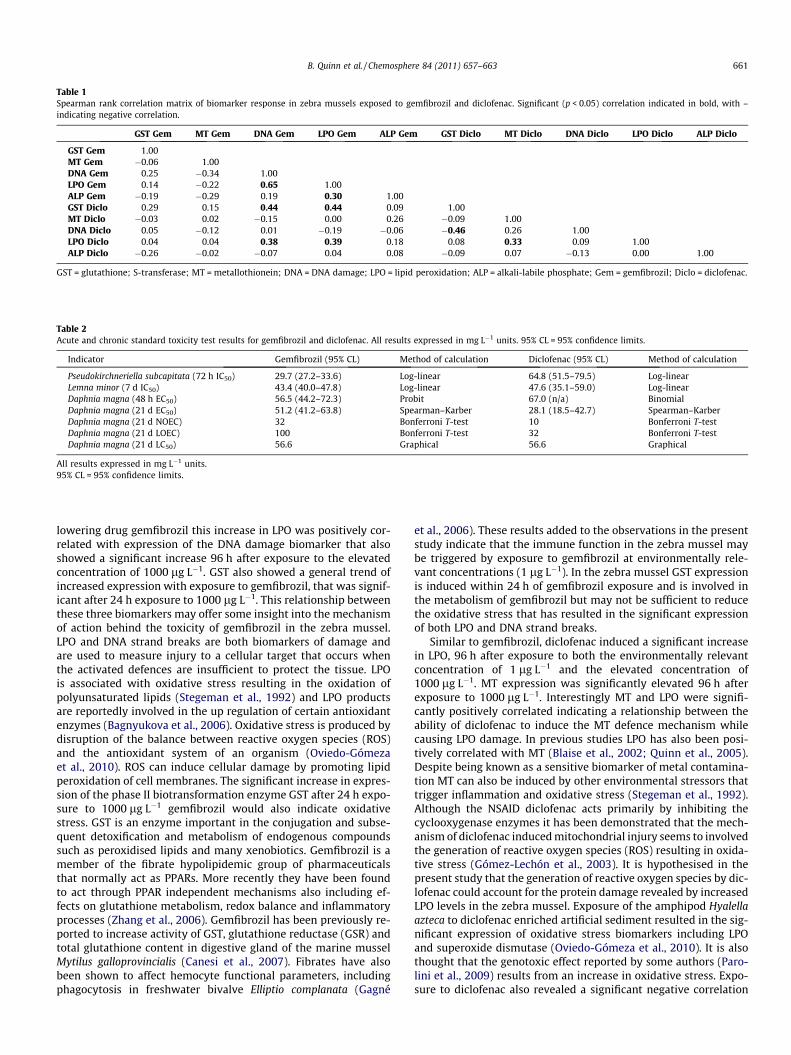
Table 1Spearman rank correlation matrix of biomarker response in zebra mussels exposed to gemfibrozil and diclofenac. Significant (p < 0.05) correlation indicated in bold, with –indicating negative correlation.
GST Gem MT Gem DNA Gem LPO Gem ALP Gem GST Diclo MT Diclo DNA Diclo LPO Diclo ALP Diclo
GST Gem 1.00MT Gem �0.06 1.00DNA Gem 0.25 �0.34 1.00LPO Gem 0.14 �0.22 0.65 1.00ALP Gem �0.19 �0.29 0.19 0.30 1.00GST Diclo 0.29 0.15 0.44 0.44 0.09 1.00MT Diclo �0.03 0.02 �0.15 0.00 0.26 �0.09 1.00DNA Diclo 0.05 �0.12 0.01 �0.19 �0.06 �0.46 0.26 1.00LPO Diclo 0.04 0.04 0.38 0.39 0.18 0.08 0.33 0.09 1.00ALP Diclo �0.26 �0.02 �0.07 0.04 0.08 �0.09 0.07 �0.13 0.00 1.00
GST = glutathione; S-transferase; MT = metallothionein; DNA = DNA damage; LPO = lipid peroxidation; ALP = alkali-labile phosphate; Gem = gemfibrozil; Diclo = diclofenac.
Table 2Acute and chronic standard toxicity test results for gemfibrozil and diclofenac. All results expressed in mg L�1 units. 95% CL = 95% confidence limits.
Indicator Gemfibrozil (95% CL) Method of calculation Diclofenac (95% CL) Method of calculation
Pseudokirchneriella subcapitata (72 h IC50) 29.7 (27.2–33.6) Log-linear 64.8 (51.5–79.5) Log-linearLemna minor (7 d IC50) 43.4 (40.0–47.8) Log-linear 47.6 (35.1–59.0) Log-linearDaphnia magna (48 h EC50) 56.5 (44.2–72.3) Probit 67.0 (n/a) BinomialDaphnia magna (21 d EC50) 51.2 (41.2–63.8) Spearman–Karber 28.1 (18.5–42.7) Spearman–KarberDaphnia magna (21 d NOEC) 32 Bonferroni T-test 10 Bonferroni T-testDaphnia magna (21 d LOEC) 100 Bonferroni T-test 32 Bonferroni T-testDaphnia magna (21 d LC50) 56.6 Graphical 56.6 Graphical
All results expressed in mg L�1 units.95% CL = 95% confidence limits.
B. Quinn et al. / Chemosphere 84 (2011) 657–663 661
lowering drug gemfibrozil this increase in LPO was positively cor-related with expression of the DNA damage biomarker that alsoshowed a significant increase 96 h after exposure to the elevatedconcentration of 1000 lg L�1. GST also showed a general trend ofincreased expression with exposure to gemfibrozil, that was signif-icant after 24 h exposure to 1000 lg L�1. This relationship betweenthese three biomarkers may offer some insight into the mechanismof action behind the toxicity of gemfibrozil in the zebra mussel.LPO and DNA strand breaks are both biomarkers of damage andare used to measure injury to a cellular target that occurs whenthe activated defences are insufficient to protect the tissue. LPOis associated with oxidative stress resulting in the oxidation ofpolyunsaturated lipids (Stegeman et al., 1992) and LPO productsare reportedly involved in the up regulation of certain antioxidantenzymes (Bagnyukova et al., 2006). Oxidative stress is produced bydisruption of the balance between reactive oxygen species (ROS)and the antioxidant system of an organism (Oviedo-Gómezaet al., 2010). ROS can induce cellular damage by promoting lipidperoxidation of cell membranes. The significant increase in expres-sion of the phase II biotransformation enzyme GST after 24 h expo-sure to 1000 lg L�1 gemfibrozil would also indicate oxidativestress. GST is an enzyme important in the conjugation and subse-quent detoxification and metabolism of endogenous compoundssuch as peroxidised lipids and many xenobiotics. Gemfibrozil is amember of the fibrate hypolipidemic group of pharmaceuticalsthat normally act as PPARs. More recently they have been foundto act through PPAR independent mechanisms also including ef-fects on glutathione metabolism, redox balance and inflammatoryprocesses (Zhang et al., 2006). Gemfibrozil has been previously re-ported to increase activity of GST, glutathione reductase (GSR) andtotal glutathione content in digestive gland of the marine musselMytilus galloprovincialis (Canesi et al., 2007). Fibrates have alsobeen shown to affect hemocyte functional parameters, includingphagocytosis in freshwater bivalve Elliptio complanata (Gagné
et al., 2006). These results added to the observations in the presentstudy indicate that the immune function in the zebra mussel maybe triggered by exposure to gemfibrozil at environmentally rele-vant concentrations (1 lg L�1). In the zebra mussel GST expressionis induced within 24 h of gemfibrozil exposure and is involved inthe metabolism of gemfibrozil but may not be sufficient to reducethe oxidative stress that has resulted in the significant expressionof both LPO and DNA strand breaks.
Similar to gemfibrozil, diclofenac induced a significant increasein LPO, 96 h after exposure to both the environmentally relevantconcentration of 1 lg L�1 and the elevated concentration of1000 lg L�1. MT expression was significantly elevated 96 h afterexposure to 1000 lg L�1. Interestingly MT and LPO were signifi-cantly positively correlated indicating a relationship between theability of diclofenac to induce the MT defence mechanism whilecausing LPO damage. In previous studies LPO has also been posi-tively correlated with MT (Blaise et al., 2002; Quinn et al., 2005).Despite being known as a sensitive biomarker of metal contamina-tion MT can also be induced by other environmental stressors thattrigger inflammation and oxidative stress (Stegeman et al., 1992).Although the NSAID diclofenac acts primarily by inhibiting thecyclooxygenase enzymes it has been demonstrated that the mech-anism of diclofenac induced mitochondrial injury seems to involvedthe generation of reactive oxygen species (ROS) resulting in oxida-tive stress (Gómez-Lechón et al., 2003). It is hypothesised in thepresent study that the generation of reactive oxygen species by dic-lofenac could account for the protein damage revealed by increasedLPO levels in the zebra mussel. Exposure of the amphipod Hyalellaazteca to diclofenac enriched artificial sediment resulted in the sig-nificant expression of oxidative stress biomarkers including LPOand superoxide dismutase (Oviedo-Gómeza et al., 2010). It is alsothought that the genotoxic effect reported by some authors (Paro-lini et al., 2009) results from an increase in oxidative stress. Expo-sure to diclofenac also revealed a significant negative correlation

662 B. Quinn et al. / Chemosphere 84 (2011) 657–663
between GST and DNA strand breaks, potentially indicating that anincrease in GST activity could reduce the amount of DNA strandbreaks.
During the present exposure significant differences were ob-served between the control (injected with de-chlorinated tapwater) and the solvent control (injected with solvent, DMSO) ashighlighted in the GST expression in diclofenac injected mussels.An unexpectedly high level of MT expression was also found inthe 24 h control group for both diclofenac and gemfibrozil expo-sures. The only significant increase in ALP expression was seenafter 24 h exposure to solvent control, indicating no endocrine dis-rupting effect on the zebra mussels under the current conditions.The zebra mussels had a normal sex ratio (M:F) of 51:49 and52:48 for gemfibrozil and diclofenac respectively.
In the current study we also investigated the toxic potential ofgemfibrozil and diclofenac, using both standardised toxicity testson three common freshwater test species. As far as the authorsare aware this is the first reported toxicity data for gemfibrozilon the freshwater duck weed L. minor with a calculated 7 d growthinhibition IC50 of 43.4 mg L�1. For diclofenac a similar result of47.6 mg L�1 7 d IC50 for L. minor was calculated, midway betweenthe previously reported values of 7.5 mg L�1 (Cleuvers, 2003) and148 mg L�1 (Kaza et al., 2007). Similarly our reported Daphnia48 h EC50 based on immobilisation of 56.5 mg L�1 for gemfibrozilis half the value reported by (Hernando et al., 2004) of 100 mg L�1
but similar to that found by (Zurita et al., 2007) of 42.5 mg L�1. Forthe freshwater green algae P. subcapitata our 72 h IC50 result of29.7 mg L�1 for gemfibrozil is comparable with the 15.19 mg L�1
previously reported (Isidori et al., 2007) using the same technique(ISO/DIS 8692 (1989)). For diclofenac our recorded 72 h IC50
(64.8 mg L�1) was higher but in the same order of magnitude asthe previously reported 96 h EC50 based on growth of 16.3 mg L�1
(Ferrari et al., 2004). So despite there being a few slight discrepan-cies as would be expected owing to biological variation, our acutetoxicity results generally correlate well with those previously pub-lished. As far as the authors are aware this is the first publication ofa D. magna 21 d EC50 (51.2 mg L�1), NOEC (32 mg L�1) and LOEC(100 mg L�1) results based on reproduction and LC50 (56.6 mg L�1)based on mortality for gemfibrozil. The 21 d D. Magna NOEC(10 mg L�1) and LC50 (56.6 mg L�1) results for diclofenac reportedhere correlate well with those previously published by (Hanet al., 2006) of 10 and 80.1 mg L�1 respectively.
Numerous publications have highlighted the lack of chronicecotoxicity data and the reliance on acute toxicity data usingstandardised toxicity tests that may not be appropriate for thelow concentrations of these drugs found in the environment (Bou-card, 2006; Crane et al., 2006; Fent et al., 2006). It has been shownin previous studies that the classical short term aquatic tests usingalgae, Daphnia and fish, may underestimate the toxicity of thepharmaceutical compounds tested (Henschel et al., 1997). As withall scientific experiments the results obtained depend upon thequestion being asked and perhaps standardised toxicity tests basedon acute endpoints are not asking the appropriate question. Whenapplied to EU directive 93/67/EEC, results from acute toxicity testare 1000 to 10,000 times above the reported concentrations foundin the environment indicating to regulators there is no potentialthreat to the environment. With the application of an assessmentfactor of 100 to the 21 d NOEC found for Daphnia in the currentstudy the chronic effects for gemfibrozil and diclofenac are seenat 320 and 100 lg L�1 respectively, still two orders of magnitudehigher than the concentrations found in the environment. Howeverbased on the current biomarker results a significant increase in LPOwas observed at the environmentally relevant (nominal) concen-tration of 1 lg L�1 for both compounds. This and other studies(Gagné et al., 2006; Canesi et al., 2007; Parolini et al., 2009;Oviedo-Gómeza et al., 2010) investigating biomarker responses
in non-standardised test species indicate that environmental con-centrations of numerous pharmaceuticals can affect various as-pects of the immune function, glycolysis, redox balance andperoxisomal function necessary for survival and growth.
Recently there have been recommendations for the inclusion ofbiomarkers in the assessment of the impact of pharmaceuticals inthe environment, particularly based on the drugs mode of actionwhose specific effects may be underestimated by standard toxicitytests (Boucard, 2006; Crane et al., 2006; Fent et al., 2006; Santoset al., 2010). The advantage of this method is that the biomarkersare very specific, but they may overlook unintended adverse reac-tions to the compound not related to this mode of actionparticularly with our limited understanding of the physiology ofnon-target organisms. For these reasons we have opted in thisstudy to use more integrative biomarkers that can be used to as-sess the general health status of an organism and are based onessential mechanism such as oxidative stress, biotransformation,DNA damage or reproduction and are therefore considered moreecologically relevant. The zebra mussel was seen to be an appropri-ate test species for these biomarker studies indicating that there isa significant biomarker response at environmentally relevant con-centrations of both gemfibrozil and diclofenac.
5. Conclusions
The current study adds further information on the acute andchronic toxicity for the lipid regulator gemfibrozil and the NSAIDdiclofenac using standardised acute toxicity tests.
Our results indicate that these standard tests may not be appro-priate to assess the potential toxicity of pharmaceuticals toanimals exposed in the environment as toxicity occurred in thenon-environmentally relevant mg L�1 concentration range. How-ever using the multi-biomarker approach it appears that zebramussels exposed to the environmentally relevant (nominal) con-centration of 1 lg L�1 of both gemfibrozil and diclofenac showedsigns of oxidative stress with elevated lipid peroxidation levels.At the higher concentration of 1000 lg L�1 the defence mechanismof GST and MT were induced by gemfibrozil and diclofenac respec-tively. This biomarker approach provides information not only intothe potential toxic effect of the compounds under investigation,but also offers an insight into their mechanism of action at envi-ronmentally relevant and elevated concentrations.
Acknowledgements
The authors acknowledge the funding of Ireland’s Environmen-tal Protection Agency (EPA) under the Science, Technology, Re-search and Innovation for the Environment (STRIVE) Programme2007–2013 and Kelly Moran and Tracy O’Shea for their assistance.
References
Bagnyukova, T.V., Chahrak, O.I., Lushchak, V.I., 2006. Coordinated response ofgoldfish antioxidant defenses to environmental stress. Aquat. Toxicol. 78, 325–331.
Binelli, A., Cogni, D., Parolini, M., Riva, C., Provini, A., 2009. Cytotoxic and genotoxiceffects of in vitro exposure to Triclosan and Trimethoprim on zebra mussel(Dreissena polymorpha) hemocytes. Comp. Biochem. Physiol. 150C, 50–56.
Blaise, C., Gagne, F., Pellerin, J., Hansen, P.D., 1999. Determination of vitellogenin-like properties in Mya arenaria hemolymph (Saguenay Fjord, Canada): apotential biomarker for endocrine disruption. Environ. Toxicol. 14, 455–465.
Blaise, C., Gagne, F., Pellerin, J., Hansen, P.D., Trottier, S., 2002. Molluscan shellfishbiomarker study of the Quebec, Canada, Saguenay Fjord with the soft-shellclam, Mya arenaria. Environ. Toxicol. 17, 170–186.
Boryslawskyj, M., Garrood, A.C., Pearson, J.T., Woodhead, D., 1988. Elevation ofglutathione-S-transferase activity as a stress response to organochlorinecompounds, in the freshwater mussel, Sphaerium corneum. Mar. Environ. Res.24, 101–104.
Boucard, T., 2006. Environment Agency Workshop on Chronic Aquatic EcotoxicityTesting of Human Pharmaceuticals. Environment Agency, Bristol. pp. 1–94.

B. Quinn et al. / Chemosphere 84 (2011) 657–663 663
Bradford, M., 1976. A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248–254.
Byrne, R.A., Dietz, T.H., 2006. Ionic and Acid-Base consequences of exposure toincreased salinity in the zebra mussel, Dreissena polymorpha. Biol. Bull. 211, 66–75.
Canesi, L., Lorusso, L.C., Ciacci, C., Betti, M., Regoli, F., Poiana, G., Gallo, G., Marcomini,A., 2007. Effects of blood lipid lowering pharmaceuticals (bezafibrate andgemfibrozil) on immune and digestive gland functions of the bivalve mollusc,Mytilus galloprovincialis. Chemosphere 69, 994–1002.
Cleuvers, M., 2003. Aquatic ecotoxicity of pharmaceuticals including the assessmentof combination effects. Toxicol. Lett. 142, 185–194.
Crane, M., Watts, C., Boucard, T., 2006. Chronic aquatic environmental risks fromexposure to human pharmaceuticals. Sci. Total Environ. 367, 23–41.
Daughton, C.G., Ternes, T.A., 1999. Pharmaceuticals and personal care products inthe environment: agents of subtle change? Environ. Health Perspect. 107, 31.
EEA, 2010. Pharmaceuticals in the Environment. Results of an EEA Workshop.European Environment Agency, p. 33.
Fent, K., Weston, A.A., Caminada, D., 2006. Ecotoxicology of humanpharmaceuticals. Aquat. Toxicol. 76, 122–159.
Ferrari, B., Mons, R., Vollat, B., Fraysse, B., Paxeus, N., Lo Giudice, R., Pollio, A., Garric,J., 2004. Environmental risk assessment of six human pharmaceuticals: are thecurrent environmental risk assessment procedures sufficient for the protectionof the aquatic environment? Environ. Toxicol. Chem. 23, 1344–1354.
Gagné, F., Berube, E., Fournier, M., Blaise, C., 2005. Inflammatory properties ofmunicipal effluents to Elliptio complanata mussels lack of effects from anti-inflammatory drugs. Comp. Biochem. Physiol. 141C, 332–337.
Gagné, F., Blaise, C., Fournier, M., Hansen, P.D., 2006. Effects of selectedpharmaceutical products on phagocytic activity in Elliptio complanatamussels. Comp. Biochem. Physiol. 143C, 179–186.
Gagné, F., Andre, C., Cejka, P., Hausler, R., Fournier, M., Blaise, C., 2008. Immunotoxiceffects on freshwater mussels of a primary-treated wastewater before and afterozonation: a pilot plant study. Ecotoxicol. Environ. Saf. 69, 366–373.
Gomez, M.J., Bueno, M.J.M., Lacorte, S., Fernandez-Alba, A.R., Aguera, A., 2007. Pilotsurvey monitoring pharmaceuticals and related compounds in a sewagetreatment plant located on the Mediterranean coast. Chemosphere 66, 993–1002.
Gómez-Lechón, M.J., Ponsoda, X., O’Connor, E., Donato, T., Jover, R., Castell, J.V.,2003. Diclofenac induces apoptosis in hepatocytes. Toxicol. in Vitro 17, 675–680.
Haap, T., Triebskorn, R., Köhler, H.-R., 2008. Acute effects of diclofenac and DMSO toDaphnia magna: immobilisation and hsp70-induction. Chemosphere 73, 353–359.
Han, G., Hur, H., Kim, S., 2006. Ecotoxicological risk of pharmaceuticals fromwastewater treatment plants in Korea, occurence adn toxicity to Daphniamagna. Environ. Toxicol. Chem. 25, 265–271.
Henschel, K.P., Wenzel, A., Diedrich, M., Fliedner, A., 1997. Environmental hazardassessment of pharmaceuticals. Regul. Toxicol. Pharm. 25, 220–225.
Hernando, M.D., Petrovic, M., Fernández-Alba, A.R., Barceló, D., 2004. Analysis byliquid chromatography-electrospray ionization tandem mass spectrometry andacute toxicity evaluation for [beta]-blockers and lipid-regulating agents inwastewater samples. J. Chromatogr. A 1046, 133–140.
Hoeger, B., Kollner, B., Dietrich, D.R., Hitzfeld, B., 2005. Water-borne diclofenacaffects kidney and gill integrity and selected immune parameters in browntrout (Salmo trutta f. fario). Aquat. Toxicol. 75, 53–64.
Hong, H.N., Kim, H.N., Park, K.S., Lee, S.K., Gu, M.B., 2007. Analysis of the effectsdiclofenac has on Japanese medaka (Oryzias latipes) using real-time PCR.Chemosphere 67, 2115–2121.
Isidori, M., Nardelli, A., Pascarella, L., Rubino, M., Parrella, A., 2007. Toxic andgenotoxic impact of fibrates and their photoproducts on non-target organisms.Environ. Int. 33, 635–641.
Kaza, M., Nalecz-Jewecki, G., Sawicki, J., 2007. The toxicity of selectedpharmaceuticals to the aquatic plant Lemna minor. Fresenius Environ. Bull. 16,524–531.
Letzel, M., Metzner, G., Letzel, T., 2009. Exposure assessment of the pharmaceuticaldiclofenac based on long-term measurements of the aquatic input. Environ. Int.35, 363–368.
Livingstone, D., 1998. The fate of organic xenobiotics in aquatic ecosystems:quantitative and qualitative differences in biotransformation by invertebratesand fish. Comp. Biochem. Physiol 120A, 43–49.
Metcalfe, C.D., Miao, X.S., Koenig, B.G., Struger, J., 2003. Distribution of acidic andneutral drugs in surface waters near sewage treatments plants in the lowerGreat Lakes, Canada. Environ. Toxicol. Chem. 22, 8.
Mimeault, C., Woodhouse, A., Miao, X.S., Metcalfe, C.D., Moon, T.W., Trudeau, V.L.,2005. The human lipid regulator, gemfibrozil bioconcentrates and reducestestosterone in the goldfish, Carassius auratus. Aquat. Toxicol. 73, 44–54.
Mimeault, C., Trudeau, V.L., Moon, T.W., 2006. Waterborne gemfibrozil challengesthe hepatic antioxidant defense system and down-regulates peroxisomeproliferator-activated receptor beta (PPARP) mRNA levels in male goldfish(Carassius auratus). Toxicology 228, 140–150.
Oaks, J.L., Gilbert, M., Virani, M.Z., Watson, R.T., Meteyer, C.U., Rideout, B.A.,Shivaprasad, H.L., Ahmed, S., Iqbal Chaudhry, M.J., Arshad, M., Mahmood, S., Ali,A., Ahmed Khan, A., 2004. Diclofenac residues as the cause of vulture populationdecline in Pakistan. Nature 427, 630–633.
Olive, P., 1988. DNA precipitation assay: a rapid and simple method for detectingDNA damage in mammalian cells. Environ. Mol. Mutagen. 11, 487–495.
Oviedo-Gómeza, D.G.C., Galar-Martínezb, M., García-Medinab, S., Razo-Estradab, C.,Gómez-Olivána, L.M., 2010. Diclofenac-enriched artificial sediment inducesoxidative stress in Hyalella azteca. Environ. Toxicol. Pharmacol. 29, 39–43.
Parolini, M., Binelli, A., Cogni, D., Riva, C., Provini, A., 2009. An in vitro biomarkerapproach for the evaluation of the ecotoxicity of non-steroidal anti-inflammatory drugs (NSAIDs). Toxicol. in Vitro 23, 935–942.
Parolini, M., Binelli, A., Cogni, D., Provini, A., 2010. Multi-biomarker approach for theevaluation of the cyto-genotoxicity of paracetamol on the zebra mussel(Dreissena polymorpha). Chemosphere 79, 489–498.
Quinn, B., Gagne, F., Costello, M., McKenzie, C., Wilson, J., Mothersill, C., 2004. Theendocrine disrupting effect of municipal effluent on the zebra mussel (Dreissenapolymorpha). Aquat. Toxicol. 66, 279–292.
Quinn, B., Gagne, F., Weber, J.P., Blaise, C., 2005. Ecotoxicological effects of a semi-submerged municipal dump (Castle harbour, Bermuda) on the Calico scallopArgopecten gibbus. Mar. Poll. Bull. 51, 534–544.
Quinn, B., Gagne, F., Blaise, C., Costello, M.J., Wilson, J.G., Mothersill, C., 2006.Evaluation of the lethal and sub-lethal toxicity and potential endocrinedisrupting effect of nonylphenol on the zebra mussel (Dreissena polymorpha).Comp. Biochem. Physiol. 142C, 118–127.
Santos, L.H.M.L.M., Araújo, A.N., Fachini, A., Pena, A., Delerue-Matos, C., Montenegro,M.C.B.S.M., 2010. Ecotoxicological aspects related to the presence ofpharmaceuticals in the aquatic environment. J. Hazard. Mater. 175, 45–95.
Schwaiger, J., Ferling, H., Mallow, U., Wintermayr, H., Negele, R.D., 2004. Toxiceffects of the non-steroidal anti-inflammatory drug diclofenac part 1:histopathological alterations and bioaccumulation in rainbow trout. Aquat.Toxicol. 68, 141–150.
Staels, B., Dallongeville, J., Auwerx, J., Schoonjans, K., Leitersdorf, E., Fruchart, J.-C.,1998. Mechanism of action of fibrates on lipid and lipoprotein metabolism.Circulation 98, 2088–2093.
Stanton, M.G., 1968. Colorimetric determination of inorganic phosphate in thepresence of biological material and adenosine triphosphate. Analyt. Biochem.22, 27–34.
Stegeman, J., Brouwer, M., Di Giulio, R., Forlin, L., Fowler, B., Sanders, B., Van Veld, P.,1992. Molecular responses to environmental contamination: enzyme andprotein systems as indicators of chemical exposure and effect. In: Huggett, R.,Kimerle, R., Mehrle, P., Bergman, H. (Eds.). Biomarkers: biochemical,physiological and histological markers of anthropogenic stress. A SpecialPublication of SETAC. Lewis Publishers, Chelsea, MI, pp. 235–335.
Triebskorn, R., Casper, H., Heyd, A., Eikemper, R., Kohler, H.R., Schwaiger, J., 2004.Toxic effects of the non-steroidal anti-inflammatory drug diclofenac part II.Cytological effects in liver, kidney, gills and intestine of rainbow trout(Oncorhynchus mykiss). Aquat. Toxicol. 68, 151–166.
Viarengo, A., Ponzano, E., Dondero, F., Fabbri, R., 1997. A simple spectrophotometricmethod for metallothionein evaluation in marine organisms: an application tomediterranean and Antarctic molluscs. Mar. Environ. Res. 44, 69–84.
Wills, E.D., 1987. Evaluation of lipid peroxidation in lipids and biologicalmembranes. In: Snell, K., Mullock, B. (Eds.), Biochemical Toxicology: APractical Approach. IRL Press, Washington, DC, pp. 127–152.
Zhang, X., Tanaka, N., Nakajima, T., Kamijo, Y., Gonzalez, F.J., Aoyama, T., 2006.Peroxisome proliferator-activated receptor [alpha]-independent peroxisomeproliferation. Biochem. Biophys. Res. Commun. 346, 1307–1311.
Zurita, J.L., Repetto, G., Jos, A., Salguero, M., Lopez-Artiguez, M., Camean, A.M., 2007.Toxicological effects of the lipid regulator gemfibrozil in four aquatic systems.Aquat. Toxicol. 81, 106–115.





























