Embed Size (px)
Citation preview

Camp. Bochem. Phy&L, 1971, Vat. 45A, pp. 871 to 880. Pergamon Press. printed in Great Britain
EFFECTS OF THE PARASITE, APANTELES CONGREGATUS, ON RESPI~TION OF TOBACCO HORNWOR~,
~AN~UCA SEXTA LARVAE*
DOUGLAS L. DAHLMAN and FREDDIE HERALD
Department of Entomology, University of Kentucky, Lexington, Kentucky 40506
(lieceiwed 10 May 1971)
Abstract-l. Oxygen consumption of diet-reared Mandwa sexta larvae changes in a repeated and predictable pattern during development of each stadium.
2. The first four instars of unparasitized larvae respire as a direct function of their weight, slope = 1.
3. The highly significant deviation from a slope of 1 (slope 6 t) found in the fifth stadium is attributed to the production of low-metabolizing tissue by the developing larvae.
4. Parasites have a damping effect on the normal cycle of Or consumption with active feeding phases consuming less and inactive phases (during apolysis and ecdysis) consuming more Oa than expected.
INTRODUCTION
METABOLIC rate is influenced by such factors as the presence of parasites, changes in temperature, chemical inhibitors, hormone concentration and type of diet (Zwicky SC Wigglesworth, 1956; Duerr, 1967; Skelton & Hunter, 1970). The relationship between an organism’s metabolic rate and weight can be mathe- matically expressed by the equation
M = aI@,
where M is oxygen consumption per unit time, W is weight and a and b are empirically determined constants. This equation is more usually written in the form
log IM= loga+b log w.
Oxygen consumption has long been used as an index of metabolic rate. The importance of this relationship, especially the slope of the line b, was considered by Duerr (1967, and references cited therein), and more recently Schroeder &
* The investigation reported in this paper (No. 71-7-52) is in connection with a project of the Kentucky Agricultural Experiment Station and is published with approval of the Director.
871

872 DOUGLAS L. DAHLMAN AND FREDDIE HERALD
Dunlap (1970) reviewed aspects related to immature stages of holometabolous insects.
Von Bertalanffy (1957) grouped organisms into three classes based on the
exponent b: (1) those whose metabolic rate followed the surface law, b = #; (2) those whose metabolic rate was linearly related to weight, b = 1; and (3) those
whose metabolic rate was intermediate between types 1 and 2, 8 < b < 1. Growing insect larvae and hemimetabolous insects exhibit exponential growth, and accord-
ing to von Bertalanffy 11957, 1960) they belong to the second metabolic type, i.e. b = 1.
Zwicky & Wigglesworth (1956) reported that oxygen consumption of each instar of the hempteran Rhodnius prolixus changed in a predictable pattern during de- velopment and this pattern was repeated in each instar. Changes in oxygen con-
sumption were related to protein synthesis and ecdysis. Sluss (1968) reported that adult male convergent lady beetles, Hipp~damia convergens, parasitized by the braconid wasp, Plaits cocci~eZl~, consumed less oxygen than unp~asitized
beetles. Dahlman (1969) demonstrated that a number of physiological parameters associated with the tobacco hornworm, Manduca sexta, larvae undergo a cycle
which is repeated, with minor variations, in each stadium. These patterns are altered when internal parasites are present in the hornworm larvae.
Body-weight of the tobacco hornworm increases many 100 times during larval development, with about 85 per cent of the total weight attained during the last
instar. Fat body, which serves as a depot for materials to be used during meta- morphosis, makes a significant contribution to the increase in weight of the fifth instar. It is also known that fat body development is greatly retarded and levels of
blood trehalose are also greatly depleted in hornworm larvae parasitized by the braconid wasp, Apanteles congregatus (Dahlman, 1970). Sluss (1968) observed morphological lesions in the fat body of the coccinellid, H. converge-m, following parasitism by a braconid wasp and the fat body of the tobacco budworm, Heliothis virescens, appeared to be the only tissue outside of the hemolymph that was noticeably affected following parasitism by the braconid wasp, C~diochiles nigriceps, up to the time the parasite began to consume all internal tissues (Vinson & Barras, 1970).
Work reported in this paper was undertaken (1) to determine whether there was a repeated cyclic pattern of oxygen consumption in each instar of hornworm larvae, (2) to determine the influence of an internal parasite on such a pattern, if it did exist and (3) to examine the influence of large amounts of non- or Iow-metaboli- zing tissue on total oxygen consumption in relation to total weight.
MATERIALS AND METHODS
Hornworm larvae were reared on synthetic diet following procedures described by Hoffman et al. (1966) at 27°C and 16 hr of light. Parasitization was accomplished by placing newly hatched larvae (less than 12 hr old) on a tobacco plant which was placed in a cage containing A. congregutus wasps. After 48 hr of exposure to the wasps, the larvae were transferred and held on the synthetic diet until they were used.

EFFECTS OF PARASITE ON RESPIRATION OF TOBACCO HORNWORM 873
The period of development of each instar was divided into phases which were described in detail by Dahlman (1969). Basically the phases were as follows: phase I, period between apolysis (the freeing of the epidermal cells from the old cuticle) and ecdysis as defined by Jenkin & Hinton (1966); phase II, period of initiation of feeding shortly after ecdysis; phase III, larvae actively feeding and half of normal instar growth completed; phase IV, maximum instar size has been reached but the larvae is still feeding and actively moving about.
A Warburg constant volume respirometer was used to measure oxygen consumption at 28°C. Techniques similar to those described by Umbreit et al. (1957) were followed. All tests used individual hornworms, with the exception of tests with first instars which em- ployed groups of five larvae. Each homworm was used only once. For testing, a larva weighing less than O-6 g was placed inside a 23-ml Warburg vessel. A screen cap prevented contact of the freely moving larvae with 0.5 ml of 5% KOH in the center well. Larvae weighing more than 0.6 g were placed in specially constructed 35-ml flasks which contained 0.5 ml of 5% KOH in a screen-covered center well. The Warburg vessels were fitted to the manometers, inserted into a 28°C water-bath and allowed to equilibrate for 15 min before collection of oxygen consumption data was started. The time between readings ranged from every 15 min for first and second instars to every 2 min for fifth instars. At least four consecutive readings with similar corrected changes in manometer fluid height were re- quired as a criterion for acceptance of data.
Upon completion of the test, dry weights of unparasitized larvae were determined by placing them in a drying oven at 105°C until a constant weight was obtained. Parasitized larvae were killed in boiling water and preserved in 70% alcohol. The larvae were dissected and counts of the total number of parasites were made to determine a possible correlation between parasite number and oxygen consumption. No larvae were used from which parasites had emerged.
Possible error due to oxygen uptake occurring in the feces was partially eliminate by the pre-test fasting period of l-2 hr. However, it was established that oxygen uptake by the feces was insignificant for the period of time involved in the observations. This confirmed information reported by Schroeder & Dunlap (1970).
RESULTS
A significant increase (P ,< O-05) in body-weight usually occurred between successive phases in both parasitized and unparasitized larvae (Table 1). Excep- tions occurred during the period of apolysis and ecdysis when the weight did not increase significantly from the preceding phase or even significantly decreased. With the exception of phase I, weights for the first three instars of parasitized larvae were generally si~ific~tly greater (P< O-05) than unparasitized Iarvae, Weights of phase I parasitized larvae were always either significantly less (P = 0.05) or not different from weights of unparasitized larvae. Weights of parasitized and unparasitized fourth and fifth instars were generally not significantly different. Two exceptions were (1) phase XII, fourth instar, when parasitized larvae were, for some unexpl~ned reason, much heavier (P = O-001), and (2) phase IV, fifth instar, when parasitized larvae weighed signicantly less (P = 04331) than unparasitized larvae.
There were significant differences (P< 0.05) in total oxygen consumption between successive phases of homworm larvae (Table 2). Unparasitized phase I was always significantly lower (P < 0405) than either the preceding or following phase, and there was usually no significant increase in oxygen ~o~umption between

TABLE
I-M
EA
N~E
T
WEI
GH
T OF
VA
RIO
US
STAGES
OF LARVAE
OF THE TOBACCO
HORNWORM,M.
sext
a
P
valu
e le
vel
of
Un
par
asit
ized
P
aras
itiz
ed
sign
ific
ance
(t
-tes
t)
+ S
tan
dar
d
+ S
tan
dar
d
Inst
ar a
nd
W
eigh
t N
o.
of
erro
r of
W
eigh
t N
o.
of
erro
r of
p
has
e $
Un
par
asit
ized
vs
. tm
g)
larv
ae
mea
n
(mg)
la
rvae
m
ean
p
aras
itiz
ed
u
z
1 IV
13
*0**
16
2.
9 6.
4t
20
0.3
0.00
1 $
21
6.8*
* 13
0.
2 7*
0**
19
0.2
> 0
*500
%
2 II
21
*7t
15
1.4
19.1
**
20
1.0
0.20
0 r
2 II
I 19
.4**
20
0.
8 28
.7 *
13
3.
2 o*
oos
tl
2 IV
31
.6**
17
1.
2 50
*7t
10
10.6
0.
050
G
31
36.5
**
11
I.5
30.8
**
7 2.
1 0*
050
E
3 II
67
.4*
* 20
7.
0 12
2.8t
17
13
.0
0.00
1 $
3 II
I lo
l~o*
* 18
7.
1 14
@0*
* 28
6.
4 0.
001
3 IV
19
1.7t
%
19
17
.3
458.
5 *
* 6
40.1
O
*OO
l u
41
178.
1**
13
9.4
157-
o**
17
6.7
o*os
o 2
4 II
26
1.6*
* 18
25
.6
277*
9* *
10
31
.0
> o
*soo
B
4
III
453*
4**
14
45.0
90
5.2
t 14
84
.8
O*O
Ol
u
4 IV
10
50*5
t
15
40.9
90
7.9
* i;j
10
83
.1
0.20
0 51
10
76.4
t 20
25
.2
1195
*5t
16
85.3
0.
200
5 II
10
30*8
**
F 20
58
.3
1023
.9*
17
120.
5 >
0~
500
5 II
I 14
44*9
**
19
51.4
13
90*0
+*
16
115.
6 >
0.5
00
$
5 IV
71
21.2
18
17
4.4
1995
.5
19
128.
7 O
*OO
l
* S
ign
ific
ant
at P = 0
.05
leve
l of
pro
bab
ilit
y fr
om
valu
e d
irec
tly
foll
owin
g it
in
th
e co
lum
n
(t-t
est)
. **
S
ign
ific
ant
at P = 0
.01
leve
l of
pro
bab
ilit
y fr
om v
alu
e d
irec
tly
foll
owin
g it
in
th
e co
lum
n
(t-t
est)
. t
Not
si
gnif
ican
t at
P = 0.05 leve
l of
pro
bab
ilit
y fr
om v
alu
e d
irec
tly
foll
owin
g it
in
th
e co
lum
n
(t-t
est)
. :
Des
crib
ed
in M
ater
ials
an
d M
eth
ods
sect
ion
.

TA
BL
E ~
-ME
AN
O
2 C
ON
SU
ME
D/h
r per
LA
RV
AE
OF
VA
RIO
US
ST
AG
ES
OF
TO
BA
CC
O HO
RN
WO
RM
, M.
sext
a
P
valu
e le
vel
of
Unp
aras
itiz
ed
Par
asit
ized
si
gnif
ican
ce
(t-t
est)
f S
tand
ard
+ S
tand
ard
Inst
ar a
nd
O9
cons
umed
N
o.
of
9 er
ror
of
0,
cons
umed
N
o. o
f er
ror
of
Unp
aras
itiz
ed
vs.
phas
e +’
$/
hr
per
larv
ae
larv
ae
mea
n
$/hr
pe
r la
rvae
la
rvae
m
ean
para
siti
zed
8 +u
I IV
21
**
16
1.4
6t
20
0.5
0.00
1 21
6*
* 13
0.
6 7*
* 19
0.
6 0.
200
g
2 II
26
t 15
2.
9 37
t 20
3.
0 0.
025
Yi
2 II
I 29
**
20
2.3
41*
13
5.6
0*05
0 8
2 IV
55
**
17
3.1
76t
10
16.4
0.
200
31
34**
11
5.
9 40
**
7 2.
7 >
0*50
0 B
3 II
11
2t
20
11.5
13
8f
17
14.3
2
0.20
0 5!
3
III
141*
* 18
9.
2 15
1**
28
10.1
>
0*50
0 ti
3
IV
264*
* 19
21
.0
574*
* 6
49.9
0.
001
41
180*
* 13
$
5.0
155*
* 17
13
.2
0.20
0 8
4 II
33
1**
18
28.1
38
2**
10
49.4
04
00
4 II
I 85
8*
14
110.
2 81
4T
14
73.2
>
0~50
0 :
4 IV
11
48**
15
75
.6
907t
10
56
.9
0*05
0 E
51
751*
* 20
32
.3
875t
16
45
.2
0.05
0 Q
5
II
1419
t 20
65
.5
1193
t 17
15
6.2
0.20
0 5
III
1512
**
19
138.
3 14
23 t
16
12
1.7
g
5 IV
39
94
18
> 0*
500
335.
2 17
32t
19
100.
1 0.
001
2 g
* S
igni
fica
nt a
t P
=
0.0
5 le
vel
of p
roba
bili
ty
from
val
ue d
irec
tly
foll
owin
g it
in
the
colu
mn
(t-t
est)
. z
** S
igni
fica
nt
at P
=
0.
01
leve
l of
pro
babi
lity
fr
om v
alue
dir
ectl
y fo
llow
ing
it i
n th
e co
lum
n (t
-tes
t).
t N
ot s
igni
fica
nt
at P
=
0.
05 l
evel
of
prob
abil
ity
from
val
ue d
irec
tly
foll
owin
g it
in
the
colu
mn
(t-t
est)
. $
Des
crib
ed
in M
ater
ials
and
Met
hod
s se
ctio
n.

876 DOUGLAS L. DAHLMAN AND FREDDIE HERALD
phases II and III, although weight significantly increased. This pattern was not as
pronounced in parasitized larvae, but, oxygen consumption by phase I was usually lower and there was usually no significant increase in oxygen consumption between phases II and III. It differed in that no significant increase between successive
phases was found from phase III, fourth instar to the end of larval development. A plot of oxygen consumption/unit weight shows a regular cyclic pattern in
unparasitized larvae and less regular pattern in parasitized larvae (Fig. 1). A
decrease in oxygen consumption in phase I is the most common feature of this pattern.
FIG. 1. Oxygen consumption &l/mg) by parasitized (a) and unparasitized (0) M. sexta during larval development.
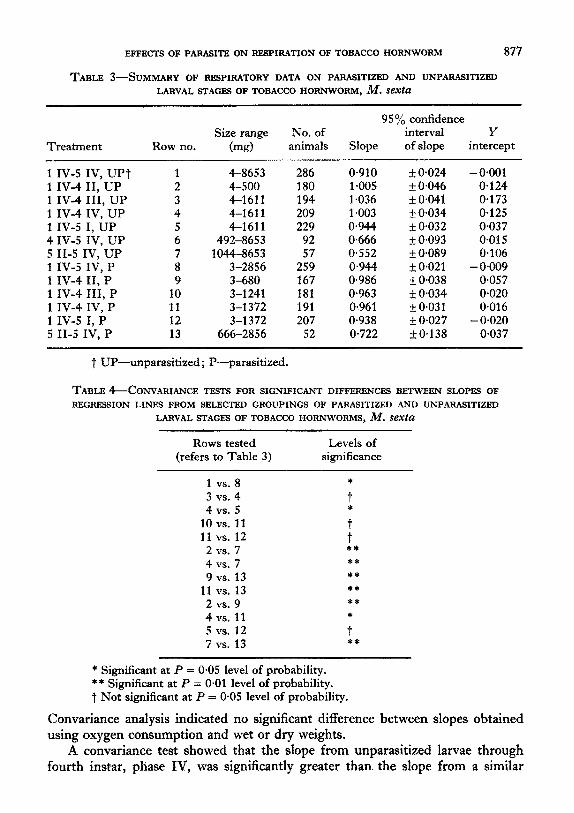
The logarithmic relationship between oxygen consumption/hr and wet weight was calculated by least square regression analysis using the model : log Y = log X+ /3 log X+log E. Table 3 contains information on slopes, Y-intercepts and con- fidence intervals of selected groupings of parasitized and unparasitized larvae. Results of covariance tests for differences between the slopes of these lines are given in Table 4. It was assumed that both parasitized and unparasitized larvae came from the same population because a null hypothesis test for common variance (a2) was true.
Least squares regression analysis, using dry weight instead of wet weight, was also run on first instar, phase IV through fifth instar, phase IV unparasitized larvae.
EFFECTS OF PARASITE ON RESPIRATION OF TOBACCO HORNWORM 877
TABLE 3-SUMMARY OF RESPIRATORY DATA ON PARASITIZED AND UNPARASITIZED
LARVAL STAGES OF TOBACCO HORNWORM, M. SeXtU
Treatment
1 IV-5 IV, UP7 1 IV-4 II, UP 1 IV-4 III, UP 1 IV-4 IV, UP 1 IV-5 I, UP 4 IV-5 IV, UP 5 II-5 IV, UP 1 IV-5 IV, P 1 IV-4 II, P 1 IV-4 III, P 1 IV-4 IV, P 1 IV-5 I, P 5 II-5 IV, P
95 o/0 confidence Size range No. of interval Y
Row no. (mg) animals Slope of slope intercept
1 4-8653 286 0.910 -t o-024 - 0.001 2 4-500 180 1.005 zf 0.046 0.124 3 4-1611 194 I.036 + 0.041 0.173 4 4-1611 209 I.003 zt 0.034 0.125 5 4-1611 229 0.944 rt 0.032 0.037 6 492-8653 92 0.666 F 0,093 0.015 7 1044-8653 8 3-2856
2:; 0.552 rt 0.089 0,106 0.944 zt 0.021 - 0,009
9 3-680 167 O-986 rt 0.038 0.057 10 3-l 241 181 0.963 rf: 0.034 0.020 11 3-1372 191 0.961 zk 0.031 0.016 12 3-l 372 207 0.938 E!Z 0.027 - 0.020 13 666-2856 52 0.722 rt 0.138 0.037
1 UP-unparasitized ; P-parasitized.
TABLE $_-CONVARIANCE TESTS FOR SIGNIFICANT DI FFRRFNCES BETWEE SLOPES OF
RRGRFSION LINES FROM SELECTED GROUPINGS OF PARASITIZED AND UNPARASITIZED
LARVAL STAGES OF TOBACCO HORNWORMS, M. S.?XtU
Rows tested Levels of (refers to Table 3) significance
1 vs. 8 3 vs. 4 4 vs. 5
10 vs. 11 11 vs. 12 2 vs. 7 4 vs. 7 9 vs. 13
11 vs. 13 2 vs. 9 4 vs. 11 5 vs. 12 7 vs. 13
;
; t ** ** ** ** **
: **
* Significant at P = 0.05 level of probability. ** Significant at P = 0.01 level of probability. t Not significant at P = 0.05 level of probability.
Convariance analysis indicated no significant difference between slopes obtained using oxygen consumption and wet or dry weights.
A convariance test showed that the slope from unparasitized larvae through fourth instar, phase IV, was significantly greater than the slope from a similar

878 DOUGLAS L. DAHLMAN AND FREDDIE HERALD
group of parasitized larvae (Table 4; 4 vs. 11) and that the slope obtained from active feeding fifth instars was significantly less than the slope from a similar group
of parasitized larvae (Table 4; 7 vs. 13). Figure 2 illustrates two distinct rates of oxygen consumption during different periods of unparasitized larval development.
A similar phenomenon existed in parasitized larvae, and the data in Tables 3 and 4 show the gradual decrease in slope (i.e. Table 3, rows 2-5 and 9-12) as the result of
lower rates of oxygen uptake by older larvae.
Wet weight, g/individual
FIG. 2. Relationship of oxygen consumption (ml/hr) during two different periods of unparasitized M. sextu larval development. Length of line is not indicative of
range in weights, see Table 3.
No significant correlation was found between the number of parasites present
in the host (the number ranged from 20 to more than 300) and the rate of oxygen consumption/mg wet weight.
DISCUSSION
A mature second instar parasite is 4.5-5.0 mm long and has an average diameter of 1.2 mm (Fulton, 1940). A conservative estimate of its volume would be 6.4 mm3. On this basis, 100-200 mature parasites make up between 35 and 60 per cent of the total volume of fifth instar, phase IV parasitized hornworms. Thus, the actual weight of the host was much less than that reported. The significantly greater larval weight of parasitized second and third instars may be explained by the presence of the internal parasites. However, temporary nutritional benefits derived from the plant during the 4%hr parasitization period may also be re- sponsible for the significant difference because control larvae were not placed on the plants.
The cyclic pattern of total oxygen consumption and oxygen consumed/mg tissue by unparasitized larvae illustrates the considerable variation between different

EFFECTS OF PARASITE ON RESPIRATION OF TOBACCO HORNWORM 879
physiological phases and emphasizes the importance of knowing the physiological condition of the test organism. Decreased oxygen consumption in phase I may result from decreased activity of this phase when compared with the active feeding phases II, III and IV.
Hornworm larvae reach phase II, second instar approximately 3 days after the mid-point of exposure to wasps. The peak oxygen consumption/mg found at this point in development could coincide with the time of the hatching of the parasite eggs, which have an incubation period of 3 days at 29-30°C (Fulton, 1940). The presence of parasites seemed to have a damping effect on the normal cycle with parasitized phase I consuming more oxygen than expected and the active phases II, III and IV consuming less oxygen than expected.
Von Bertalanffy’s hypothesis (1957,196O) for holometabolous insect larvae does not hold for the entire larval stage of either parasitized or unparasitized tobacco hornworms because the slopes of the regression lines for the combinations of all phases of parasitized and unparasitized larvae are less than 1 (Table 3 ; rows 1 and 8). However, when only the data from 1st through fourth stadia are examined, the regression of larval oxygen consumption on weight does give a slope of 1 (Table 3, rows 24; Fig. 2) and supports von Bertalanffy’s hypothesis. The slope of a re- gression line calculated for unparasitized larvae from fourth instar, phase IV through fifth instar, phase IV, is 0.67 (Table 3, row 6; Fig. 2), the exponent pre- dicted for organisms whose metabolic rate follows the surface law. The actual transition begins in fourth instar, phase III, and the metabolic rate steadily declines, as indicated by the decrease in slope of the regression lines (Table 3, rows 3-7). This coincides precisely with the rapid development of fat body in unparasitized larvae and supports the suggestion of Zuethen (1953) that the ob- served depression of metabolic rates with increased size is the result of the “new” tissue being formed having a greater proportion of non-metabolically active components.
A similar phenomena was observed in parasitized larvae, but the decrease in slope was not as great (Table 3, rows 10-13). Fat body is poorly or not at all developed in fifth instar parasitized larvae (D~lm~, 1970) and the reduction in metabolic rate which was observed probably resulted from the build-up of storage tissues in the parasite larvae which in turn was reflected in total oxygen consump- tion of the parasite-host combination.
REVERENCES DAHLMAN D. L. (1969) Haemofymph specific gravity, soluble total protein and total solids of
plant-reared, normal, and parasitized diet-reared tobacco homworm larvae. J. Insect Physiol. 15, 2075-2084.
DAHIMAN D. L. (1970) Trehalose levels in parasitized and nonparasitized tobacco hom- worm, vodka se&a, larvae. Amt. ent. Sot. Am. 63, 615-617.
DUERR F. G. (1967) Changes in the size-metabolic rate relationship of Lymnaeu stugdis ;;y_;; Say produced by digenetic trematode parasitism. Comp. Biochem. Physiol. XI,
FULTON B. B. (1940) The homworm parasite, Apanteles congregatus Say and the hyper- parasite, Hypop~~~al~ tabacum (Fitch). Ann. ent. Sot. Am. 33, 2X-244.

880 DOUGLAS L. DAHLMAN AND FREDDIE HERAI.D
HEMMINGSEN A. M. (1960) Energy metabolism as related to body size and respiratory surfaces, and its evolution. Rep. Steno. Meml. Hosp. 9, (Part 2), l-110.
HOFFMAN J. D., LAWSON F. R. & YAMAMOTO R. T. (1966) Tobacco hornworms. In Insect Colonization and Muss Reproduction (Edited by SMITH C. N.), pp. 479-486. Academic Press, New York.
JENKIN P. M. & HINTON H. E. (1966) Apolysis in Arthropod moulting cycles. Nature, Lond. 211, 871.
SCHROEDER L, & DUNLAP D. G. (1970) Respiration of cecropia moth (~~~ZO~~OY~ cecropia L.) larvae. Corn@ &o&972. P~ysioZ. 35, 953-957.
SKELTON T. E. & HUNTER P. E. (1970) Flight and respiration responses in houseflies following topical application of sterilization levels of Tepa. Ann. ent. Sot. Am. 63, 770-773.
SLUSS R. (1968) Behavioral and anatomical responses to the convergent lady beetle to parasitism by Perilitus coccinellue (Schrank) (Hymenoptera: Braconidae). J. Invert. Path. 10, 9-27.
UMBREIT W. W., BURRIS R. & SXUJFFER J. (1957) Munometric Tec?miques, 3rd edn. Burgess, Minneapolis.
VINSON S. B. & Barras D. J. (1970) Effects of the parasitoid, Ca~dioc~~Zes nigriceps, on growth, development, and tissues of HeZiothis virescens. J. Insect Physiol. 16, 1329-1338.
VON BERTALANFFY L. (1957) Quantitative laws in metabolism and growth. Q. Rev. Biol. 32, 217-231.
VON BERTALANFFY L. (1960) Principles and theory of growth. In Fundamental Aspects of Normal and Malignant Growth (Edited by NOWINSKI W. W.), pp. 137-259. Elsevier, Amsterdam.
ZEUTHEN E. (1953) Oxygen uptake as reiated to body size in organisms. Q. Rev. Biol. 28, 1-12.
ZWICKY K. & WIGGLESWORT~ V. B. (1956) Th e course of oxygen consumption during the moulting cycle of Rhodnius proZixus Stol (Hemiptera). Proc. R. Ent. Sot. Lond. (A) 31, 153-160.
Key Word Index-Tobacco hornworm; Manduca sexta; Apanteles congregatus; Sphin- gidae ; Braconidae ; respiration; host-parasite interaction ; growth ; fat body.
![arXiv:1802.02678v1 [q-bio.NC] 8 Feb 2018 · Hebbian plasticity. ... edge, we have constructed an end-to-end computational model of the Manduca sexta moth olfactory](https://img.pdfslide.us/doc/110x75/5b0475ff7f8b9a6c0b8dd8a4/arxiv180202678v1-q-bionc-8-feb-2018-plasticity-edge-we-have-constructed.jpg)







![Manduca sexta · 2017. 8. 28. · lipophorin (Lp) and lipid transport particles (LTPs) are responsible for the intercellular transportation of choles-terol and lipids in insects [5-7]](https://img.pdfslide.us/doc/110x75/60907ad3b500a22d98078c8f/manduca-sexta-2017-8-28-lipophorin-lp-and-lipid-transport-particles-ltps.jpg)




















