Embed Size (px)
DESCRIPTION
123
Citation preview

J Comp Physiol B (2008) 178:515–527DOI 10.1007/s00360-007-0244-6
123
ORIGINAL PAPER
Parasitism of an insect Manduca sexta L. alters feeding behaviour and nutrient utilization to inXuence developmental success of a parasitoid
S. N. Thompson · R. A. Redak
Received: 27 August 2007 / Revised: 5 December 2007 / Accepted: 16 December 2007 / Published online: 15 January 2008© Springer-Verlag 2008
Abstract The eVects of macronutrient balance on nutrientintake and utilization were examined in Manduca sexta lar-vae parasitized by Cotesia congregata. Insects fed an artiW-cial diet having constant total macronutrient, but withvaried ratios of protein and carbohydrate, with altered dietconsumption in response to excesses and deWciencies of theindividual macronutrients. Bivariate plots of protein andcarbohydrate consumption for non-parasitized larvae demon-strated a curvilinear relationship between points of nutrientintake for the various diets, and the larvae grew best oncarbohydrate-biased diets. The relationship was linear forparasitized larvae with the growth uniform across diets. Onprotein-biased diets, the larvae regulated the nitrogen con-tent, containing similar amounts of nitrogen regardless ofconsumption. EYciency of nitrogen conversion in non-para-sitized larvae was greatest on carbohydrate-biased diets,while nitrogen conversion by parasitized larvae was great-est with intermediate nutrient ratios. Accounting for carbo-hydrate consumption, the lipid content decreased as dietarycarbohydrate increased, but parasitized larvae containedsigniWcantly less lipid. The total biomass of parasites deve-loping in individual host larvae was positively correlatedwith host protein consumption, but the individual parasiteswere similar in size. Parasitism inXuences host nutrientconsumption in a manner that achieves uniform hostgrowth under diverse nutritional regimes, thereby con-straining blood nutrient concentrations within limitssuitable for parasite growth and development.
Keywords Nutrient utilization · Nutrition · Manduca sexta · Cotesia congregata · Parasitism
Introduction
Parasitism is the symbiotic association or “durable interac-tion” where one organism, the parasite, lives at the expenseof another, the host (Roberts and Janovy 2005; Combes2001). Parasitic associations are common among insects,involving principally insects parasitized by other insectscalled parasitoids (Askew 1971; Eggleton and Gaston1990). The immature or larval stages are parasitic, whileadult parasitoids are free-living. There are an estimated600,000 species of insect parasitoids, predominantly in theorder Hymenoptera, of which approximately 60,000 spe-cies are described (Eggleton and Belshaw 1992). Parasiticinsects are thus among the most successful and plentifulorganisms known (Hearaty 2007).
The host relationships of many insect endoparasites arecomplex, involving dramatic changes in host behaviour andphysiology (Quickie 1997) that are frequently mediated byparasitoid-derived factors including venoms and polydnavi-ruses, the latter an integral component of the parasitoidgenome (Federici and Bigot 2003). Suppression of the hostimmune system (Amaya et al. 2005; Beckage 1998) andendocrine-based alterations of host development (Beckageand Gelman 2001; Edwards et al. 2001; Kelly et al. 1998)are perhaps the best characterized physiological eVects.Collectively, manifestations of parasitism often are consid-ered manipulation of the host by the parasite (Cole et al.2002; Jones et al. 1986; Espagne et al. 2005; Thompson1990), a reXection of Richard Dawkin’s (1999) “extendedphenotype”, where the phenotype of the parasitized host isexpressed to the beneWt of the parasite rather than the host
Communicated by I.D. Hume.
S. N. Thompson (&) · R. A. RedakDepartment of Entomology, University of California, Riverside, CA 92521, USAe-mail: [email protected]; [email protected]

516 J Comp Physiol B (2008) 178:515–527
123
(Combes 2001). Other than the intuitively obvious advan-tage provided by immunosuppression, however, there islimited evidence in support of this paradigm of adaptivebeneWt regarding the modiWed traits of parasitized insects.Indeed, the “adaptive” signiWcance inferred in many casesfor the altered behaviour of parasitized animals fail to fulWllthe minimal criteria necessary to support that conclusion(Poulin 1995). Because nourishment is essential for suc-cessful parasite growth and development, investigations ofhost feeding and nutrition, and of how nutrients are parti-tioned between host and parasite to inXuence parasite suc-cess, may provide new insights into the ultimate role andcosts of parasite-induced alterations of host behaviour andphysiology.
Cotesia congregata is a gregarious braconid parasite oflarvae of the sphingid moth Manduca sexta. Adult femaleparasitoids deposit large numbers of eggs into individualhost larvae. After hatching, parasite larvae develop in thehost’s haemocoel, feeding on the blood (haemolymph).When mature, parasitoids emerge from the host, undergothe last larval moult and subsequently form pupae fromwhich develop adults of the next generation of parasitoids.The parasitic relationship of C. congregata with M. sexta isvery suitable for examining nutritional interactions becausethe number of parasite larvae developing in individual hostlarvae is highly variable, from a few to hundreds of para-sites representing in extreme cases one-quarter the entiremass of the parasite/host complex (Alleyne and Beckage1997; Thompson et al. 2005a).
Employing a geometric analysis of feeding (Simpsonand Raubenheimer 1993a, b; Raubenheimer and Simpson1997, 1999), we recently examined the eVects of dietarymacronutrient ratio (protein:carbohydrate or P:C) on nutri-ent intake and growth over the fourth and Wfth stadia ofM. sexta, comparing parasitized larvae with normal non-parasitized insects (Thompson et al. 2005a). Normal insectsgrew best on a diet consisting of equal amounts of proteinand carbohydrate. Further, when oVered a choice of variousdiets having diVerent P:C ratios, these larvae consistentlydisplay an intake target ratio, the ratio of protein to carbo-hydrate required for optimal growth, of approximately 1:1(Thompson and Redak 2005). When normal larvae wereoVered individual diets where the P:C ratio was displacedfrom 1:1, insects responded by increasing or decreasingconsumption to compensate for deWciencies and excessesof protein or carbohydrate in a manner predictable for ageneralist feeder (Simpson and Raubenheimer 1993a, b;Thompson and Redak 2005). In contrast, parasitized insectsresponded to macronutrient imbalance by consuming lessdiet regardless of whether protein or carbohydrate was thedeWcient nutrient. The total number and biomass ofparasites developing in individual host larvae was corre-lated with host haemolymph nutrient levels (protein, amino
acids and trehalose), which, in turn, depended upon nutri-ent intake and dietary nutrient balance (Thompson et al.2005a, b).
A signiWcant Wnding of the above investigations was thatthe eVects of dietary macronutrient balance on growth andhaemolymph nutrient levels in parasitized M. sexta and per-haps, therefore, the numbers of C. congregata developingin host larvae are not simply due to diVerences in nutrientintake between diets. We speculated that dietary nutrientratio may inXuence nutrient utilization. Studies by othershave established that conversion eYciency in some insectsvaries with dietary macronutrient balance (Raubenheimerand Simpson 1999; Zanotto et al. 1993, 1997; Lee et al.2002).
EVects of parasitism and interactions between parasitismand macronutrient balance on nutrient utilization by lepi-dopteran larvae are relatively unknown. While results ofsome investigations indicate that insect parasites them-selves have high eYciencies of food utilization, theseinvestigations failed to consider how parasite developmentis inXuenced by the eVects of parasitism on host nutrientintake and nutrient availability to developing parasites(Slansky and Scriber 1985; Slansky 1986; Thompson1999). Ultimately, insect parasites obtain nourishmentthrough the feeding activities of their hosts. Parasitedevelopment, therefore, depends on a tritrophic interactionwith all trophic levels inXuencing the Wnal outcome.Furthermore, understanding how parasitism aVects hostnutrient intake and utilization is critical for assessing theadaptive consequences of changes in host behaviour andphysiology to successful parasite development.
Here, we investigate the eVects of variation in dietarymacronutrient levels on nutrient intake and utilization bynormal and parasitized M. sexta larvae, and in the case ofparasitized insects, distinguish nutrient utilization directedtoward host mass gain from that directed towards the devel-oping parasite. Based on earlier Wndings, we predict thatchanges in post-ingestive regulation of nutrient utilizationoccur in parasitized larvae; however, because growth of lar-vae is low and uniform with dietary nutrient ratio, wehypothesize that host nutrient utilization is signiWcantlyreduced by parasitism and that host growth and parasitedevelopment have higher relative energy costs than is thecase with growth and development of normal non-para-sitized insects.
Materials and methods
Insect culture
M. sexta was reared on a wheat germ-based artiWcial diet(Bell and Joachim 1976), as described (Thompson and

J Comp Physiol B (2008) 178:515–527 517
123
Redak 2005a). Second instar M. sexta larvae were parasit-ized by C. congregata in 3 l glass jars containing 50–100adult parasitoids of mixed sex. The parasitized host larvaewere isolated and placed individually on the rearing diet.All larvae were housed in an incubator at 28°C with a 16 hlight/8 h dark non-diapausing, long-day photocycle. Theparasites emerged from the hosts during the last or Wfth sta-dium. Emerging second instar parasite larvae moult into thethird stadium and immediately pupate. Pupae were col-lected from the surface of the host and held in plastic cups,approximately 150/cup, and placed in 3 l glass jars. Theemerged adult parasitoids were provided with water andhoney.
Rearing on experimental diets
M. sexta were developmentally synchronized as pharatefourth instar larvae (Baker et al. 1987) and were superpara-sitized two to four times (Alleyne 1995). Larvae weresuperparasitized to obtain large parasite burdens, or num-bers of parasites developing in individual host larvae,thereby maximizing the nutritional impact of parasitism.Normal (non-parasitized) and parasitized larvae were thenmaintained on the rearing diet until the end of the fourthstadium. Pharate Wfth instar larvae were placed on chemi-cally deWned fat-free experimental artiWcial diets. The arti-Wcial diet was a simpliWed formulation of the chemicallydeWned diet described by Ahmad et al. (1989). Casein andsucrose were the sole sources of digestible protein and car-bohydrate, respectively, and the basic formulation con-tained 60 g/l of each macronutrient. Other ingredientsincluded: ascorbic acid, inorganic salts, cholesterol andvitamins. The nutrients were purchased principally fromNutritional Biochemicals (Cleveland, OH, USA) and Bio-serve (Frenchtown, NJ, USA). The pH of the diet wasadjusted with KOH to approximately 6.5. Larvae were notconditioned prior to feeding on the experimental diets andbegan feeding after moulting to the Wfth stadium. Theexperimental diets varied in protein and carbohydrate asdescribed below and the larvae were maintained in an incu-bator under the conditions outlined above.
Feeding protocol
Groups of ten larvae were fed on one of nine experimentaldiets for 3–8 days after moulting to the Wfth stadium. Thelarvae were allowed to feed until cessation of feeding dur-ing the wandering phase. All diets had the same combinedor total level of macronutrient, protein plus carbohydrate(120 g/l of diet), but the following ratios of casein (g/l P)and sucrose (g/l C), respectively: 100P:20C, 90P:30C,80P:40C, 70P:50C, 60P:60C, 50P:70C, 40P:80C, 30P:90Cand 100P:20C. This variation in dietary nutrient ratio nar-
rowed what was employed during our earlier investigations(Thompson et al. 2005a, b), which included extreme “path-ological” ratios where insects failed to grow and develop.Diets were prepared in a 2% agar solution having anapproximate 6.3:1 ratio of agar solution to total dry ingredi-ents.
The larvae were fed individual pieces of diet approxi-mately 4 £ 2.5 £ 1 cm and weighing about 10 g in 130 mlplastic cups with snap-cap lids. They were housed in anincubator maintained at 28°C. In the case of non-parasit-ized insects, diet consumption was measured after 2, 3 and5 days, and at the time they stopped feeding to enter intothe wandering stage in preparation for pupation. Diet con-sumption by parasitized larvae was similarly measured at 2,3 and 5 days, and at the time they stopped feeding andparasite larvae began emerging. The diet was replaced asnecessary.
Estimation of growth and nutrient consumption
At the end of the feeding period, larvae were dissected toremove undigested food from the gut. In the case of para-sitized larvae, the dissected hosts were examined under amicroscope and the parasites were collected from the hae-mocoel. The carcasses of the individual M. sexta larvaeincluding their emptied guts were dried in an oven atapproximately 75°C for 72 h and weighed. The parasitesfrom parasitized larvae were counted and those collectedfrom each host larvae were dried together as above. TheWnal dry mass was used as the measure of growth. For sta-tistical purposes, the initial dry mass of the experimentallarvae was estimated after determining the average dry/wetmass ratio of 10 larvae sampled from the initial insect popu-lation. These larvae were dried as above to determine theirwater content. The average dry/wet mass ratio was calcu-lated and applied to the wet mass of larvae used in theexperiments. The diet remaining at the end of the feedingtrials was dried as above and the total consumption calcu-lated as the diVerences in dry mass of the remaining dietplus that of the gut contents and the dry mass of the totaldiet given to larvae calculated from the known dry masscontent of the fresh diet (Thompson et al. 2005a). Proteinand carbohydrate intake were then calculated from theknown casein and sucrose content of the diet consumed.
Fat and nitrogen analyses
We estimated the contributions of protein and carbohydrateto larval growth by analyzing the larvae for nitrogen and fatcontent. The dried carcasses of M. sexta larvae and thecombined parasites collected from individual parasitizedhosts were powdered with a mortar and pestle. Each samplewas divided approximately in half. Fat was extracted from

518 J Comp Physiol B (2008) 178:515–527
123
one portion and nitrogen analyzed in the second. Fat wasextracted by Wltering each sample three times with approxi-mately 10–15 volumes of petroleum ether (35–60°C). Thesolvent was evaporated from the combined Wltrates and thefat determined gravimetrically with a microbalance.
Nitrogen was determined with a Thermo-Finnigan nitro-gen/carbon analyzer (Model: Flash EA1112) in accordancewith the method outlined in the operating manual (Chapter8: Analytical Methods, Analytical method for the NC-soils,NC-sediments and NC-Wlters conWguration, pp 184–186).
Geometric representation and analysis of data
The geometric analysis of Raubenheimer and Simpson(1997, 1999) was employed to examine the eVects of die-tary macronutrient ratio on growth, nutrient consumptionand nutrient utilization by M. sexta larvae. In this analyticalapproach, protein and carbohydrate consumption are shownas the two axes of a single bivariate plot, with consumptionof the two nutrients indicated by points along linear trajec-tories or nutrient rails deWned by the nutrient ratios of theindividual diets. We measured diVerences in feedingresponse between individual diets by determining thelength or distance along the nutrient rails to the points ofnutrient intake at 2 and 3 days and until the termination offeeding at the wandering stage for normal larvae and at 2, 3and 5 days and until the end of feeding by parasitized lar-vae. Nutrient rail distance was calculated according to Rau-benheimer and Simpson (2003) using the Pythagoreantheorem where rail distance is the hypotenuse of a right tri-angle formed by carbohydrate consumption and proteinconsumption in the bivariate plot.
The response of larvae to macronutrient imbalance, thatis, the feeding response of larvae to dietary nutrient ratiosthat deviate from the intake target ratio is further suggestedby the relationship between the points of nutrient intakeacross the nutrient rails for the diVerent diets. The shape ofthe line connecting the intake points, referred to as a nutri-ent intake array, varies depending upon the larval responseto excesses and deWciencies in macronutrients (Raubenhei-mer and Simpson 2003). The intake target ratio for Wfthinstar M. sexta larvae, demonstrated experimentally, isapproximately 1:1 protein to carbohydrate, (Thompson andRedak 2005).
Statistics
The eVects of dietary macronutrient ratio and parasitism ongrowth and nutrient intake by M. sexta were principallyanalyzed by analysis of variance (ANOVA) and analysis ofcovariance (ANCOVA) using initial mass as the primarycovariate to account for diVerences in initial mass betweenthe experimental host larvae (Raubenheimer and Simpson
1992; Horton and Redak 1993). Analyses were also con-ducted using additional covariates and multiple covariatesas indicated. Unless otherwise indicated, ANCOVA dataare presented as least square means § SE (standard error).Normality and homogeneity of variance were establishedwith the Shapiro–Wilk “W” test and normal probabilityplots. Preliminary analyses determined that the data met theassumptions of analysis of variance.
Accelerated failure time analysis was employed toexamine the eVects of diet on the end time for feeding bynormal and parasitized larvae (Fox 2001; Lee et al. 2002).
All statistics were performed using SAS (version 9.02,2003; SAS Institute Inc., Cary, NC, USA).
Results
EVects of dietary nutrient ratio and parasitism by C. congregate on nutrient consumption and larval development time of M. sexta
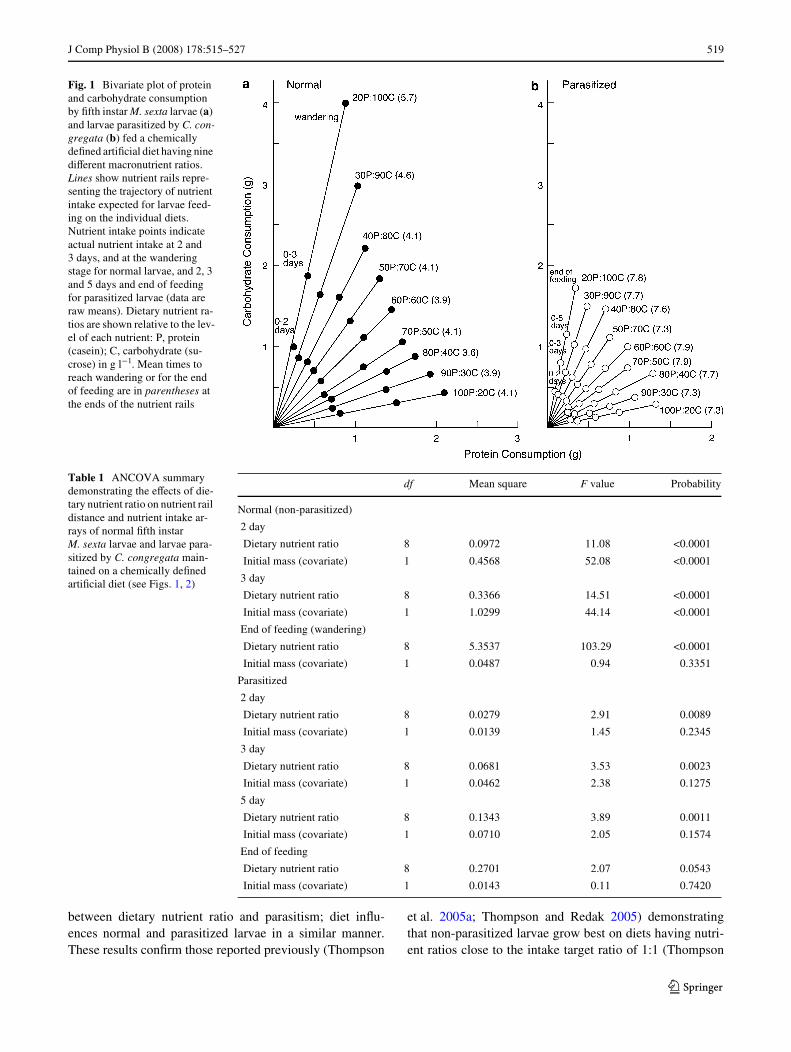
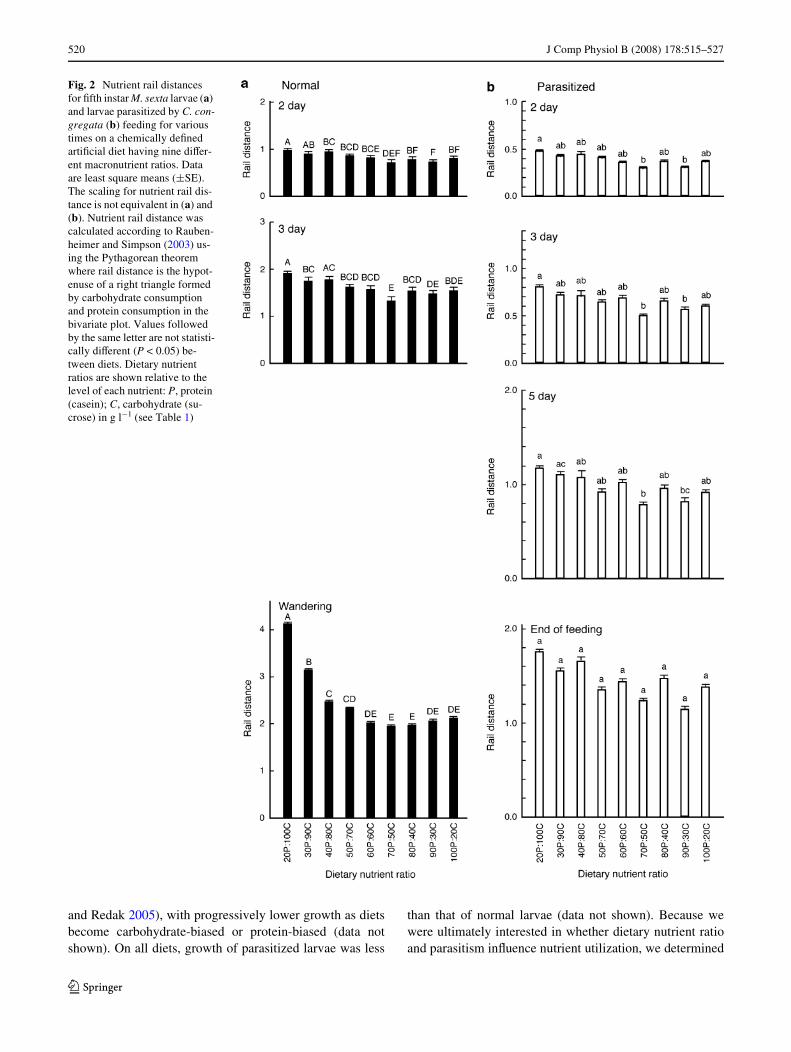
Cumulative consumption of protein and carbohydrate byM. sexta larvae on the various diets is illustrated as a stan-dard bivariate plot in Fig. 1. For normal and parasitized lar-vae, nutrient rail distance, the distance along the nutrientrail from the origin to the point of nutrient intake, wasaVected by dietary nutrient ratio throughout the feedingperiod (Table 1). The feeding response of larvae toexcesses in carbohydrate and protein were not equal.Analysis of nutrient rail distance demonstrated that larvaegenerally consume much more on diets having excesscarbohydrate with nutrient ratios less than 1, than was thecase with diets having nutrient ratios greater than 1 andequivalent excesses of protein (Fig. 2). This was mostapparent with normal larvae feeding to the end, where theactual times of feeding were longer on the most carbohy-drate-biased diet.
Failure time analysis established a signiWcant eVect ofparasitism on overall feeding time (X2 = 580.10, df = 1,P < 0.0001). Parasitized larvae fed for signiWcantly longertimes than normal larvae (see Fig. 1). Dietary nutrient ratiohad a signiWcant eVect on the time to the wandering stage bynormal larvae (X2 = 372.71, df = 8, P < 0.0001), with larvaeon the C20:100S diet feeding for the longest duration. Thediet, however, did not aVect the time to end of feeding byparasitized insects (X2 = 4.99, df = 8, P = 0.7582).
EVects of dietary nutrient ratio and parasitism by C. congregata on growth of M. sexta
The Wnal mass of M. sexta larvae was signiWcantly aVectedby dietary nutrient ratio (F = 6.27, P < 0.0001) and para-sitism (F = 584.45, P < 0.0001). There was no interaction
J Comp Physiol B (2008) 178:515–527 519
123
between dietary nutrient ratio and parasitism; diet inXu-ences normal and parasitized larvae in a similar manner.These results conWrm those reported previously (Thompson
et al. 2005a; Thompson and Redak 2005) demonstratingthat non-parasitized larvae grow best on diets having nutri-ent ratios close to the intake target ratio of 1:1 (Thompson
Fig. 1 Bivariate plot of protein and carbohydrate consumption by Wfth instar M. sexta larvae (a) and larvae parasitized by C. con-gregata (b) fed a chemically deWned artiWcial diet having nine diVerent macronutrient ratios. Lines show nutrient rails repre-senting the trajectory of nutrient intake expected for larvae feed-ing on the individual diets. Nutrient intake points indicate actual nutrient intake at 2 and 3 days, and at the wandering stage for normal larvae, and 2, 3 and 5 days and end of feeding for parasitized larvae (data are raw means). Dietary nutrient ra-tios are shown relative to the lev-el of each nutrient: P, protein (casein); C, carbohydrate (su-crose) in g l¡1. Mean times to reach wandering or for the end of feeding are in parentheses at the ends of the nutrient rails
Table 1 ANCOVA summary demonstrating the eVects of die-tary nutrient ratio on nutrient rail distance and nutrient intake ar-rays of normal Wfth instar M. sexta larvae and larvae para-sitized by C. congregata main-tained on a chemically deWned artiWcial diet (see Figs. 1, 2)
df Mean square F value Probability
Normal (non-parasitized)
2 day
Dietary nutrient ratio 8 0.0972 11.08 <0.0001
Initial mass (covariate) 1 0.4568 52.08 <0.0001
3 day
Dietary nutrient ratio 8 0.3366 14.51 <0.0001
Initial mass (covariate) 1 1.0299 44.14 <0.0001
End of feeding (wandering)
Dietary nutrient ratio 8 5.3537 103.29 <0.0001
Initial mass (covariate) 1 0.0487 0.94 0.3351
Parasitized
2 day
Dietary nutrient ratio 8 0.0279 2.91 0.0089
Initial mass (covariate) 1 0.0139 1.45 0.2345
3 day
Dietary nutrient ratio 8 0.0681 3.53 0.0023
Initial mass (covariate) 1 0.0462 2.38 0.1275
5 day
Dietary nutrient ratio 8 0.1343 3.89 0.0011
Initial mass (covariate) 1 0.0710 2.05 0.1574
End of feeding
Dietary nutrient ratio 8 0.2701 2.07 0.0543
Initial mass (covariate) 1 0.0143 0.11 0.7420
520 J Comp Physiol B (2008) 178:515–527
123
and Redak 2005), with progressively lower growth as dietsbecome carbohydrate-biased or protein-biased (data notshown). On all diets, growth of parasitized larvae was less
than that of normal larvae (data not shown). Because wewere ultimately interested in whether dietary nutrient ratioand parasitism inXuence nutrient utilization, we determined
Fig. 2 Nutrient rail distances for Wfth instar M. sexta larvae (a) and larvae parasitized by C. con-gregata (b) feeding for various times on a chemically deWned artiWcial diet having nine diVer-ent macronutrient ratios. Data are least square means (§SE). The scaling for nutrient rail dis-tance is not equivalent in (a) and (b). Nutrient rail distance was calculated according to Rauben-heimer and Simpson (2003) us-ing the Pythagorean theorem where rail distance is the hypot-enuse of a right triangle formed by carbohydrate consumption and protein consumption in the bivariate plot. Values followed by the same letter are not statisti-cally diVerent (P < 0.05) be-tween diets. Dietary nutrient ratios are shown relative to the level of each nutrient: P, protein (casein); C, carbohydrate (su-crose) in g l¡1 (see Table 1)
J Comp Physiol B (2008) 178:515–527 521
123
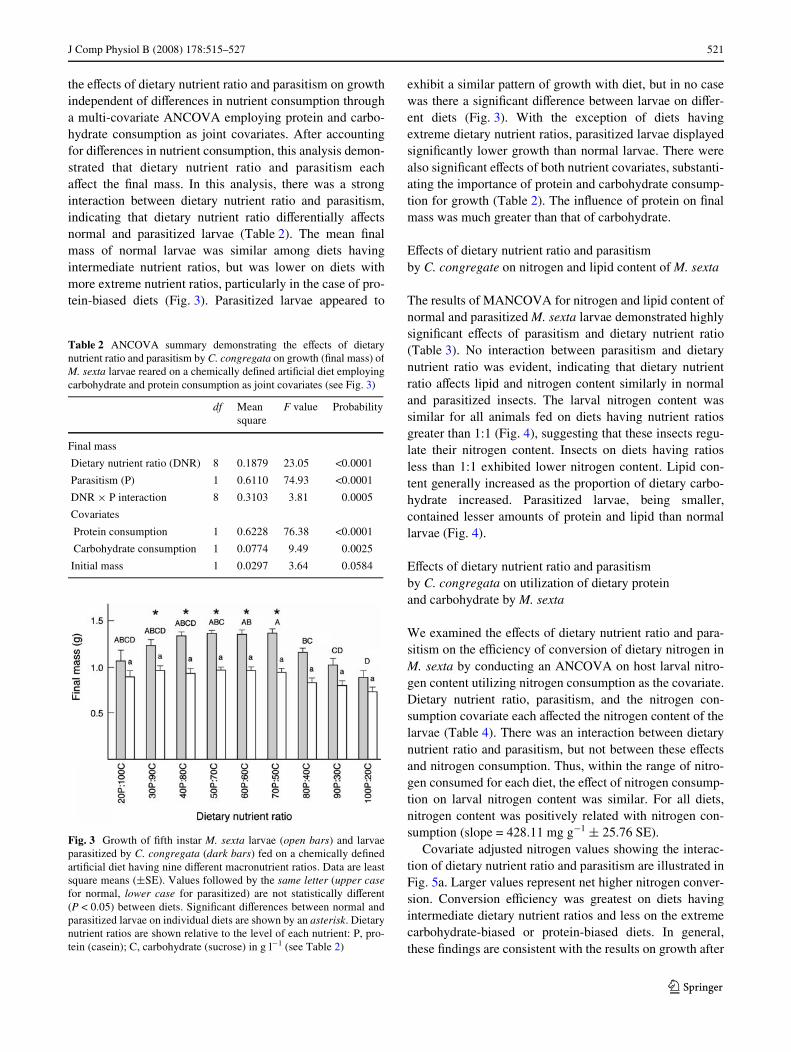
the eVects of dietary nutrient ratio and parasitism on growthindependent of diVerences in nutrient consumption througha multi-covariate ANCOVA employing protein and carbo-hydrate consumption as joint covariates. After accountingfor diVerences in nutrient consumption, this analysis demon-strated that dietary nutrient ratio and parasitism eachaVect the Wnal mass. In this analysis, there was a stronginteraction between dietary nutrient ratio and parasitism,indicating that dietary nutrient ratio diVerentially aVectsnormal and parasitized larvae (Table 2). The mean Wnalmass of normal larvae was similar among diets havingintermediate nutrient ratios, but was lower on diets withmore extreme nutrient ratios, particularly in the case of pro-tein-biased diets (Fig. 3). Parasitized larvae appeared to
exhibit a similar pattern of growth with diet, but in no casewas there a signiWcant diVerence between larvae on diVer-ent diets (Fig. 3). With the exception of diets havingextreme dietary nutrient ratios, parasitized larvae displayedsigniWcantly lower growth than normal larvae. There werealso signiWcant eVects of both nutrient covariates, substanti-ating the importance of protein and carbohydrate consump-tion for growth (Table 2). The inXuence of protein on Wnalmass was much greater than that of carbohydrate.
EVects of dietary nutrient ratio and parasitism by C. congregate on nitrogen and lipid content of M. sexta
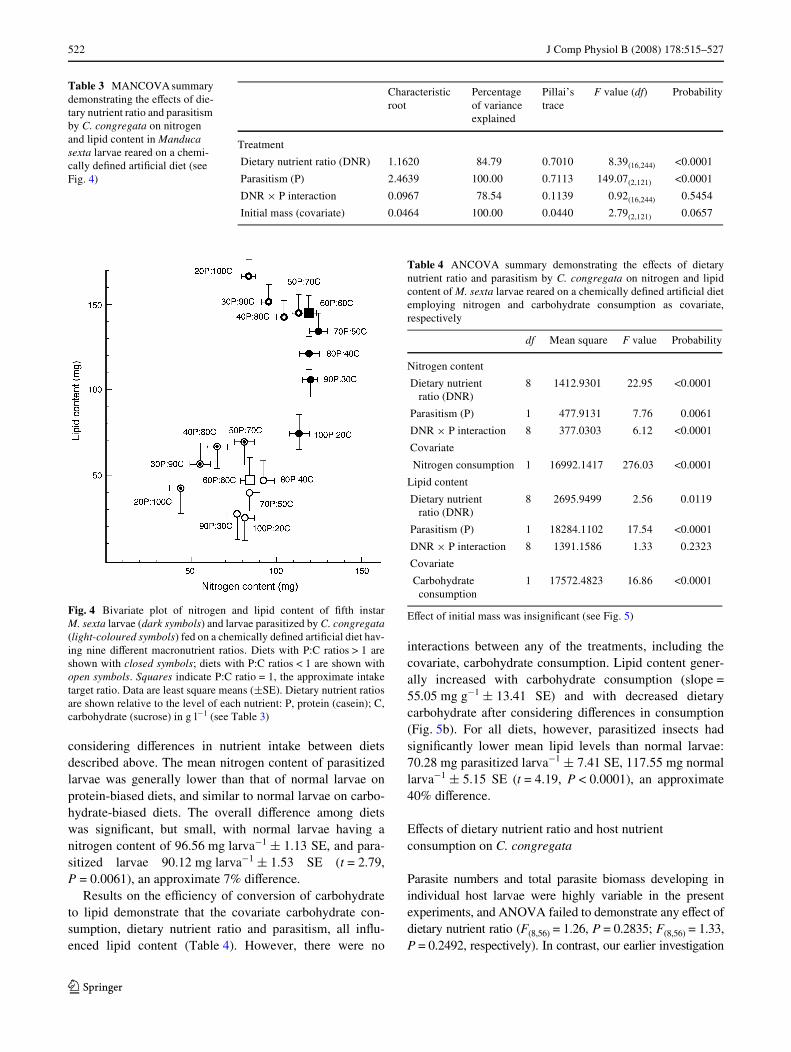
The results of MANCOVA for nitrogen and lipid content ofnormal and parasitized M. sexta larvae demonstrated highlysigniWcant eVects of parasitism and dietary nutrient ratio(Table 3). No interaction between parasitism and dietarynutrient ratio was evident, indicating that dietary nutrientratio aVects lipid and nitrogen content similarly in normaland parasitized insects. The larval nitrogen content wassimilar for all animals fed on diets having nutrient ratiosgreater than 1:1 (Fig. 4), suggesting that these insects regu-late their nitrogen content. Insects on diets having ratiosless than 1:1 exhibited lower nitrogen content. Lipid con-tent generally increased as the proportion of dietary carbo-hydrate increased. Parasitized larvae, being smaller,contained lesser amounts of protein and lipid than normallarvae (Fig. 4).
EVects of dietary nutrient ratio and parasitism by C. congregata on utilization of dietary protein and carbohydrate by M. sexta
We examined the eVects of dietary nutrient ratio and para-sitism on the eYciency of conversion of dietary nitrogen inM. sexta by conducting an ANCOVA on host larval nitro-gen content utilizing nitrogen consumption as the covariate.Dietary nutrient ratio, parasitism, and the nitrogen con-sumption covariate each aVected the nitrogen content of thelarvae (Table 4). There was an interaction between dietarynutrient ratio and parasitism, but not between these eVectsand nitrogen consumption. Thus, within the range of nitro-gen consumed for each diet, the eVect of nitrogen consump-tion on larval nitrogen content was similar. For all diets,nitrogen content was positively related with nitrogen con-sumption (slope = 428.11 mg g¡1 § 25.76 SE).
Covariate adjusted nitrogen values showing the interac-tion of dietary nutrient ratio and parasitism are illustrated inFig. 5a. Larger values represent net higher nitrogen conver-sion. Conversion eYciency was greatest on diets havingintermediate dietary nutrient ratios and less on the extremecarbohydrate-biased or protein-biased diets. In general,these Wndings are consistent with the results on growth after
Table 2 ANCOVA summary demonstrating the eVects of dietarynutrient ratio and parasitism by C. congregata on growth (Wnal mass) ofM. sexta larvae reared on a chemically deWned artiWcial diet employingcarbohydrate and protein consumption as joint covariates (see Fig. 3)
df Mean square
F value Probability
Final mass
Dietary nutrient ratio (DNR) 8 0.1879 23.05 <0.0001
Parasitism (P) 1 0.6110 74.93 <0.0001
DNR £ P interaction 8 0.3103 3.81 0.0005
Covariates
Protein consumption 1 0.6228 76.38 <0.0001
Carbohydrate consumption 1 0.0774 9.49 0.0025
Initial mass 1 0.0297 3.64 0.0584
Fig. 3 Growth of Wfth instar M. sexta larvae (open bars) and larvaeparasitized by C. congregata (dark bars) fed on a chemically deWnedartiWcial diet having nine diVerent macronutrient ratios. Data are leastsquare means (§SE). Values followed by the same letter (upper casefor normal, lower case for parasitized) are not statistically diVerent(P < 0.05) between diets. SigniWcant diVerences between normal andparasitized larvae on individual diets are shown by an asterisk. Dietarynutrient ratios are shown relative to the level of each nutrient: P, pro-tein (casein); C, carbohydrate (sucrose) in g l¡1 (see Table 2)
522 J Comp Physiol B (2008) 178:515–527
123
considering diVerences in nutrient intake between dietsdescribed above. The mean nitrogen content of parasitizedlarvae was generally lower than that of normal larvae onprotein-biased diets, and similar to normal larvae on carbo-hydrate-biased diets. The overall diVerence among dietswas signiWcant, but small, with normal larvae having anitrogen content of 96.56 mg larva¡1 § 1.13 SE, and para-sitized larvae 90.12 mg larva¡1 § 1.53 SE (t = 2.79,P = 0.0061), an approximate 7% diVerence.
Results on the eYciency of conversion of carbohydrateto lipid demonstrate that the covariate carbohydrate con-sumption, dietary nutrient ratio and parasitism, all inXu-enced lipid content (Table 4). However, there were no
interactions between any of the treatments, including thecovariate, carbohydrate consumption. Lipid content gener-ally increased with carbohydrate consumption (slope =55.05 mg g¡1 § 13.41 SE) and with decreased dietarycarbohydrate after considering diVerences in consumption(Fig. 5b). For all diets, however, parasitized insects hadsigniWcantly lower mean lipid levels than normal larvae:70.28 mg parasitized larva¡1 § 7.41 SE, 117.55 mg normallarva¡1 § 5.15 SE (t = 4.19, P < 0.0001), an approximate40% diVerence.
EVects of dietary nutrient ratio and host nutrient consumption on C. congregata
Parasite numbers and total parasite biomass developing inindividual host larvae were highly variable in the presentexperiments, and ANOVA failed to demonstrate any eVect ofdietary nutrient ratio (F(8,56) = 1.26, P = 0.2835; F(8,56) = 1.33,P = 0.2492, respectively). In contrast, our earlier investigation
Fig. 4 Bivariate plot of nitrogen and lipid content of Wfth instarM. sexta larvae (dark symbols) and larvae parasitized by C. congregata(light-coloured symbols) fed on a chemically deWned artiWcial diet hav-ing nine diVerent macronutrient ratios. Diets with P:C ratios > 1 areshown with closed symbols; diets with P:C ratios < 1 are shown withopen symbols. Squares indicate P:C ratio = 1, the approximate intaketarget ratio. Data are least square means (§SE). Dietary nutrient ratiosare shown relative to the level of each nutrient: P, protein (casein); C,carbohydrate (sucrose) in g l¡1 (see Table 3)
Table 3 MANCOVA summary demonstrating the eVects of die-tary nutrient ratio and parasitism by C. congregata on nitrogen and lipid content in Manduca sexta larvae reared on a chemi-cally deWned artiWcial diet (see Fig. 4)
Characteristic root
Percentage of variance explained
Pillai’s trace
F value (df) Probability
Treatment
Dietary nutrient ratio (DNR) 1.1620 84.79 0.7010 8.39(16,244) <0.0001
Parasitism (P) 2.4639 100.00 0.7113 149.07(2,121) <0.0001
DNR £ P interaction 0.0967 78.54 0.1139 0.92(16,244) 0.5454
Initial mass (covariate) 0.0464 100.00 0.0440 2.79(2,121) 0.0657
Table 4 ANCOVA summary demonstrating the eVects of dietarynutrient ratio and parasitism by C. congregata on nitrogen and lipidcontent of M. sexta larvae reared on a chemically deWned artiWcial dietemploying nitrogen and carbohydrate consumption as covariate,respectively
EVect of initial mass was insigniWcant (see Fig. 5)
df Mean square F value Probability
Nitrogen content
Dietary nutrient ratio (DNR)
8 1412.9301 22.95 <0.0001
Parasitism (P) 1 477.9131 7.76 0.0061
DNR £ P interaction 8 377.0303 6.12 <0.0001
Covariate
Nitrogen consumption 1 16992.1417 276.03 <0.0001
Lipid content
Dietary nutrient ratio (DNR)
8 2695.9499 2.56 0.0119
Parasitism (P) 1 18284.1102 17.54 <0.0001
DNR £ P interaction 8 1391.1586 1.33 0.2323
Covariate
Carbohydrate consumption
1 17572.4823 16.86 <0.0001
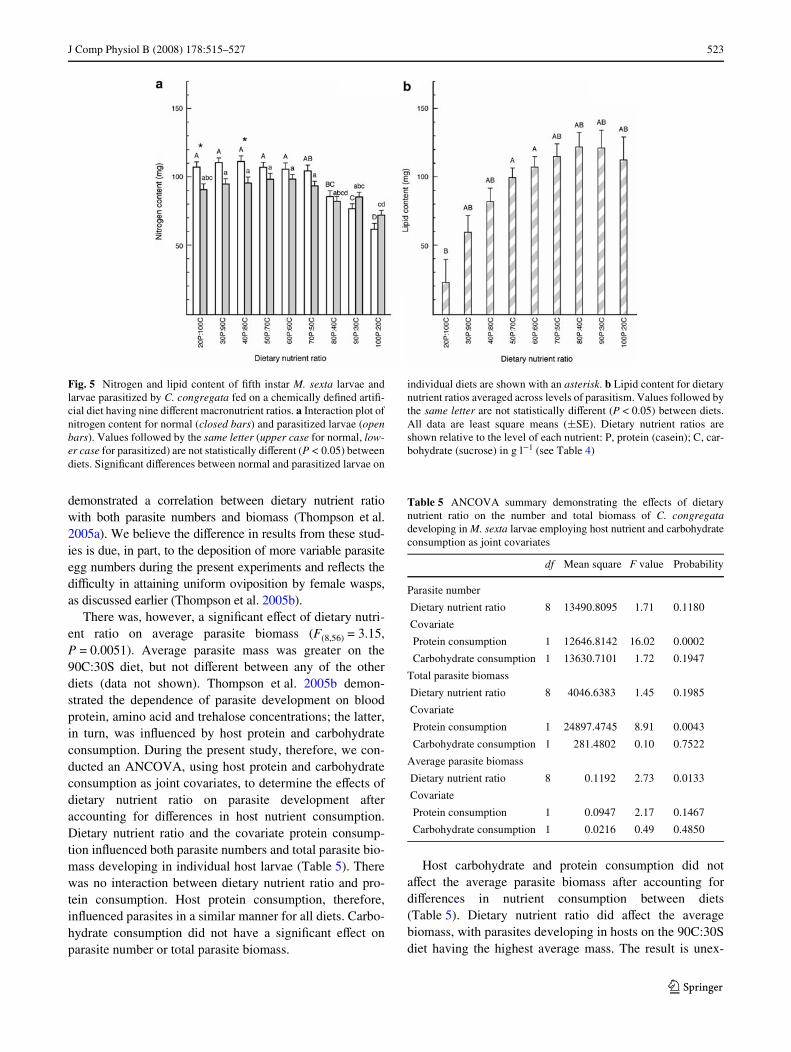
J Comp Physiol B (2008) 178:515–527 523
123
demonstrated a correlation between dietary nutrient ratiowith both parasite numbers and biomass (Thompson et al.2005a). We believe the diVerence in results from these stud-ies is due, in part, to the deposition of more variable parasiteegg numbers during the present experiments and reXects thediYculty in attaining uniform oviposition by female wasps,as discussed earlier (Thompson et al. 2005b).
There was, however, a signiWcant eVect of dietary nutri-ent ratio on average parasite biomass (F(8,56) = 3.15,P = 0.0051). Average parasite mass was greater on the90C:30S diet, but not diVerent between any of the otherdiets (data not shown). Thompson et al. 2005b demon-strated the dependence of parasite development on bloodprotein, amino acid and trehalose concentrations; the latter,in turn, was inXuenced by host protein and carbohydrateconsumption. During the present study, therefore, we con-ducted an ANCOVA, using host protein and carbohydrateconsumption as joint covariates, to determine the eVects ofdietary nutrient ratio on parasite development afteraccounting for diVerences in host nutrient consumption.Dietary nutrient ratio and the covariate protein consump-tion inXuenced both parasite numbers and total parasite bio-mass developing in individual host larvae (Table 5). Therewas no interaction between dietary nutrient ratio and pro-tein consumption. Host protein consumption, therefore,inXuenced parasites in a similar manner for all diets. Carbo-hydrate consumption did not have a signiWcant eVect onparasite number or total parasite biomass.
Host carbohydrate and protein consumption did notaVect the average parasite biomass after accounting fordiVerences in nutrient consumption between diets(Table 5). Dietary nutrient ratio did aVect the averagebiomass, with parasites developing in hosts on the 90C:30Sdiet having the highest average mass. The result is unex-
Fig. 5 Nitrogen and lipid content of Wfth instar M. sexta larvae andlarvae parasitized by C. congregata fed on a chemically deWned artiW-cial diet having nine diVerent macronutrient ratios. a Interaction plot ofnitrogen content for normal (closed bars) and parasitized larvae (openbars). Values followed by the same letter (upper case for normal, low-er case for parasitized) are not statistically diVerent (P < 0.05) betweendiets. SigniWcant diVerences between normal and parasitized larvae on
individual diets are shown with an asterisk. b Lipid content for dietarynutrient ratios averaged across levels of parasitism. Values followed bythe same letter are not statistically diVerent (P < 0.05) between diets.All data are least square means (§SE). Dietary nutrient ratios areshown relative to the level of each nutrient: P, protein (casein); C, car-bohydrate (sucrose) in g l¡1 (see Table 4)
Table 5 ANCOVA summary demonstrating the eVects of dietarynutrient ratio on the number and total biomass of C. congregatadeveloping in M. sexta larvae employing host nutrient and carbohydrateconsumption as joint covariates
df Mean square F value Probability
Parasite number
Dietary nutrient ratio 8 13490.8095 1.71 0.1180
Covariate
Protein consumption 1 12646.8142 16.02 0.0002
Carbohydrate consumption 1 13630.7101 1.72 0.1947
Total parasite biomass
Dietary nutrient ratio 8 4046.6383 1.45 0.1985
Covariate
Protein consumption 1 24897.4745 8.91 0.0043
Carbohydrate consumption 1 281.4802 0.10 0.7522
Average parasite biomass
Dietary nutrient ratio 8 0.1192 2.73 0.0133
Covariate
Protein consumption 1 0.0947 2.17 0.1467
Carbohydrate consumption 1 0.0216 0.49 0.4850
524 J Comp Physiol B (2008) 178:515–527
123
plained, and if this group were excluded from the analysisthere would be no eVect of diet. Thus, the results weregenerally similar to those reported previously where meanaverage biomass was unaVected by dietary nutrient ratio(Thompson et al. 2005b).
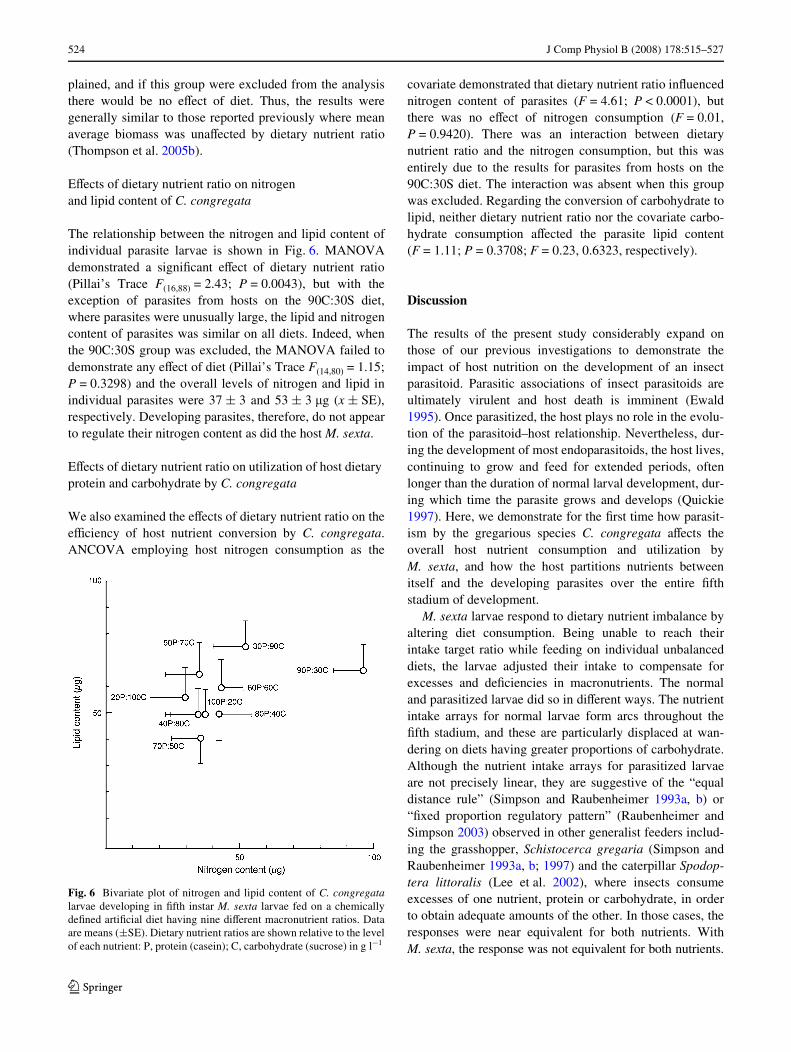
EVects of dietary nutrient ratio on nitrogen and lipid content of C. congregata
The relationship between the nitrogen and lipid content ofindividual parasite larvae is shown in Fig. 6. MANOVAdemonstrated a signiWcant eVect of dietary nutrient ratio(Pillai’s Trace F(16,88) = 2.43; P = 0.0043), but with theexception of parasites from hosts on the 90C:30S diet,where parasites were unusually large, the lipid and nitrogencontent of parasites was similar on all diets. Indeed, whenthe 90C:30S group was excluded, the MANOVA failed todemonstrate any eVect of diet (Pillai’s Trace F(14,80) = 1.15;P = 0.3298) and the overall levels of nitrogen and lipid inindividual parasites were 37 § 3 and 53 § 3 �g (x § SE),respectively. Developing parasites, therefore, do not appearto regulate their nitrogen content as did the host M. sexta.
EVects of dietary nutrient ratio on utilization of host dietary protein and carbohydrate by C. congregata
We also examined the eVects of dietary nutrient ratio on theeYciency of host nutrient conversion by C. congregata.ANCOVA employing host nitrogen consumption as the
covariate demonstrated that dietary nutrient ratio inXuencednitrogen content of parasites (F = 4.61; P < 0.0001), butthere was no eVect of nitrogen consumption (F = 0.01,P = 0.9420). There was an interaction between dietarynutrient ratio and the nitrogen consumption, but this wasentirely due to the results for parasites from hosts on the90C:30S diet. The interaction was absent when this groupwas excluded. Regarding the conversion of carbohydrate tolipid, neither dietary nutrient ratio nor the covariate carbo-hydrate consumption aVected the parasite lipid content(F = 1.11; P = 0.3708; F = 0.23, 0.6323, respectively).
Discussion
The results of the present study considerably expand onthose of our previous investigations to demonstrate theimpact of host nutrition on the development of an insectparasitoid. Parasitic associations of insect parasitoids areultimately virulent and host death is imminent (Ewald1995). Once parasitized, the host plays no role in the evolu-tion of the parasitoid–host relationship. Nevertheless, dur-ing the development of most endoparasitoids, the host lives,continuing to grow and feed for extended periods, oftenlonger than the duration of normal larval development, dur-ing which time the parasite grows and develops (Quickie1997). Here, we demonstrate for the Wrst time how parasit-ism by the gregarious species C. congregata aVects theoverall host nutrient consumption and utilization byM. sexta, and how the host partitions nutrients betweenitself and the developing parasites over the entire Wfthstadium of development.
M. sexta larvae respond to dietary nutrient imbalance byaltering diet consumption. Being unable to reach theirintake target ratio while feeding on individual unbalanceddiets, the larvae adjusted their intake to compensate forexcesses and deWciencies in macronutrients. The normaland parasitized larvae did so in diVerent ways. The nutrientintake arrays for normal larvae form arcs throughout theWfth stadium, and these are particularly displaced at wan-dering on diets having greater proportions of carbohydrate.Although the nutrient intake arrays for parasitized larvaeare not precisely linear, they are suggestive of the “equaldistance rule” (Simpson and Raubenheimer 1993a, b) or“Wxed proportion regulatory pattern” (Raubenheimer andSimpson 2003) observed in other generalist feeders includ-ing the grasshopper, Schistocerca gregaria (Simpson andRaubenheimer 1993a, b; 1997) and the caterpillar Spodop-tera littoralis (Lee et al. 2002), where insects consumeexcesses of one nutrient, protein or carbohydrate, in orderto obtain adequate amounts of the other. In those cases, theresponses were near equivalent for both nutrients. WithM. sexta, the response was not equivalent for both nutrients.
Fig. 6 Bivariate plot of nitrogen and lipid content of C. congregatalarvae developing in Wfth instar M. sexta larvae fed on a chemicallydeWned artiWcial diet having nine diVerent macronutrient ratios. Dataare means (§SE). Dietary nutrient ratios are shown relative to the levelof each nutrient: P, protein (casein); C, carbohydrate (sucrose) in g l¡1

J Comp Physiol B (2008) 178:515–527 525
123
As reXected by increased nutrient rail distance, normal lar-vae on diets having excess carbohydrate, dietary nutrientratios less than 1:1, consumed much more diet than was thecase with diets having equivalent excesses of protein. Thisdemonstrates that larvae respond more strongly to deWcien-cies in protein, than in carbohydrate, and suggests that lar-vae may have greater diYculty processing excess proteinthan excess carbohydrate. For normal larvae, this was alsoreXected by diVerences in the Wnal mass between diets,where larvae clearly grew best on diets having dietarynutrient ratios close to 1:1, the intake target ratio, with thelowest mean growth on the most protein-biased diet. Thenutrient rail distances for parasitized larvae were moreuniform between diets, but there still was a slight biastowards increased consumption on diets having highercarbohydrate.
The Wnding that growth of parasitized larvae, althoughlower than that of non-parasitized insects, was uniform withvariations in dietary nutrient ratio, consistent with theresults for nutrient consumption and with the results of ourearlier investigation demonstrating that parasitized larvaefail to regulate macronutrient intake and fail to display aspeciWc intake target ratio when oVered dietary choices(Thompson and Redak 2005). We suggest that absence of aspeciWc intake target accompanied by decreased and uni-form growth allows parasitized larvae to accept a broadrange of nutritional conditions that would be unacceptableto normal larvae. This, in turn, may enable parasitizedinsects to feed on areas within plants that are less nutrition-ally suitable and not preferred by normal larvae, therebydecreasing competition and providing an adaptive advan-tage for survival of parasitized larvae and successful devel-opment of parasites. This hypothesis is yet to be tested. Thealtered feeding response and reduced growth of parasitizedlarvae fulWll a number of the criteria outlined by Poulin(1995) for being adaptive. They are complex, show signs ofpurposeful design and have arisen in several lineages ofhosts and insect parasites. The alternate hypothesis thatreduced growth is merely a pathogenic response to para-sitism, however cannot be excluded until a Wtness beneWtfor the parasite can be demonstrated.
Despite the responses of normal and parasitized M. sextato dietary nutrient ratio, our results clearly indicate that lar-vae display post-ingestive regulation of protein utilization.On diets having dietary P:C ratios greater than 1:1 (protein-biased diets), larvae display high conversion eYcienciesand contain a similar amount of nitrogen regardless of theamount of nitrogen ingested. In this regard, the results ofM. sexta are similar to those reported for larvae of S. litto-ralis (Lee et al. 2002). Further, our results are consistentwith the eVects of dietary nutrient ratio on nutrient con-sumption, where larvae displayed diVerential responses todiets having excess protein or excess carbohydrate as dis-
cussed above. Excess nitrogen is likely eliminated throughincreased uric acid production as suggested for other spe-cies (Lee et al. 2002; Zanatto et al. 1993). Also, when car-bohydrate intake is very low, the deaminated proteinsubstrate also contributes to net carbohydrate synthesis andthe formation of trehalose through gluconeogenesis(Thompson et al. 2003). Unlike the case of S. litoralis (Leeet al. 2002), we did not observe a dramatic reduction ofnitrogen conversion eYciency on the extreme protein-biased diet; S. littoralis, however, displayed higher proteinconsumption on protein-biased diets than did M. sexta.Despite increased consumption on carbohydrate-biaseddiets, M. sexta larvae generally have lower protein contentas the dietary proportion of protein decreases.
Lipid content of both normal and parasitized larvae gen-erally increased as the proportion of dietary carbohydrateincreased. An important Wnding in these investigations wasthat after accounting for diVerences in carbohydrate con-sumption among diets, the lipid content of parasitized lar-vae was dramatically less than that of normal larvae.Although nitrogen content of parasitized larvae was alsoless after diVerences in nitrogen consumption between dietswas considered, the diVerence was slight. We suggest thatthe low level of host lipid accumulation represents the prin-cipal energetic cost of supporting parasite development.Although previous investigations suggest that one mecha-nism enabling insects to eliminate excess carbohydrate isthrough increased respiration (Zanotto et al. 1993, 1997;Lee et al. 2002), a diVerence in respiration between parasit-ized and normal larvae is unlikely to contribute to the varia-tion observed in lipid levels. Studies by others failed todemonstrate any eVect of parasitism on CO2 production byM. sexta during the feeding phase of the Wfth stadium(Alleyne et al. 1997).
Lipid synthesis and storage in insects occur principallyin the perivisceral fat body, an organ surrounding that gutand responsible for regulating much of intermediary meta-bolism (Keeley 1985; Haunerland and Shirk 1995). Previousstudies suggest that parasitism signiWcantly aVects fat bodydiVerentiation and function in M. sexta larvae. Dahlmanand Green (1981) reported that perivisceral fat body is sub-stantially diminished in parasitized larvae, a Wnding sup-ported by our own observations (unpublished), andconsistent with decreased lipid accumulation. Metabolicstudies with parasitized M. sexta demonstrate a shift of die-tary carbohydrate away from lipid synthesis into trehalose,sometimes accompanied by an elevation in blood trehaloselevel (Thompson 1993). Further, there occurs a simulta-neous increase in gluconeogenesis, presumably in theperipheral subcutaneous fat body, which is induced in para-sitized larvae under nutritional conditions that fail to stimu-late de novo carbohydrate synthesis in normal larvae(Thompson 2001; Thompson et al. 2002). Considering

526 J Comp Physiol B (2008) 178:515–527
123
these metabolic alterations, it is not surprising that dietarycarbohydrate consumption is of lesser importance than pro-tein consumption. Indeed, the present results conWrm thoseof our previous investigations indicating that host proteinconsumption is the principal factor supporting successfulparasite development.
In contrast to the results with host larvae, C. congregatado not appear to regulate either their nitrogen or lipid con-tent. The larvae had similar nitrogen and lipid levelsregardless of the host diet. This undoubtedly reXects thegenerally uniform size of both the parasitized hosts andindividual parasites, and the fact that host blood proteinnitrogen and trehalose levels that serve to nourish parasitesare maintained within a relatively narrow range over thevaried nutritional conditions employed in the present inves-tigation (Thompson et al. 2005a, b). The depressed growthof parasitized hosts and their resultant uniform size likelyhave physiological as well as ecological relevance for suc-cessful parasite development.
In conclusion, M. sexta larvae respond to variations indietary nutrient ratio and nutrient imbalances by alteringtheir diet consumption, reXecting potential trade-oVsbetween growth and the costs of the physiologicalresponses to excesses and deWciencies in macronutrients.Regulation of feeding behaviour in response to dietarynutrient composition is complex, involving modulation ofperipheral chemoreception, nutrient-allelochemical inter-actions, feedback through blood chemistry, as well asfeeding experience and learning (Simpson and White1990; Simpson and Raubenheimer 1993a, b; 1996; Glen-dinning and Slansky 1994; Thompson et al. 2001; Thomp-son and Redak 2007). How these mechanisms areintegrated with nutrient availability and the insect’s nutri-tional state is poorly understood. Recently, authors havespeculated about the involvement of biogenic amines, neuro-peptides and other hormones in feeding regulation (Rogerset al. 2004; Cohen et al. 1988, 2002; Thompson et al.2001; Bede et al. 2007). The present study demonstratesthat parasitism by C. congregata perturbs the mechanismsthat regulate feeding by M. sexta, and, moreover, appearsto do so in a manner that contributes to the nourishment ofthe parasites. Studies on parasitized insects provide aunique opportunity to examine the essential roles of vari-ous mechanisms in eVecting feeding. Previous investiga-tions on parasitized caterpillars demonstrate changes inresponsiveness to allelochemicals and in nutrient-allelo-chemical interactions (Slansky 1992; Singer et al. 2004;Thompson and Redak 2007). Changes in hormone titersare also well documented in parasitized insects (Adamoand Shoemaker 2000; Beckage and Gelman 2001). Futureinvestigations will attempt to establish physiological cor-relates more directly with feeding behaviour and nutrientintake.
References
Adamo S, Shoemaker KL (2000) EVects of parasitism on octopaminecontent of the central nervous system of Manduca sexta; a possi-ble mechanism underlying host behavioural change. Can J Zool78:1580–1587
Ahmad IM, Waldbauer GP, Friedman S (1989) A deWned artiWcial dietfor the larvae of Manduca sexta. Entomol Exp Appl 53:189–192
Alleyne M (1995) Alterations in host growth and metabolism of Mand-uca sexta larvae parasitized by Cotesia congregata, MS Thesis.University of California, Riverside
Alleyne M, Beckage NE (1997) Parasitism-induced eVects on hostgrowth and metabolic eYciency in tobacco hornworm larvae par-asitized by Cotesia congregata. J Insect Physiol 43:407–424
Alleyne M, Chappell MA, Beckage NE (1997) EVects of parasitism bythe braconid wasp Cotesia congregata on metabolic rate in hostlarvae of the tobacco hornworm, Manduca sexta. J Insect Physiol43:143–154
Amaya KE, Asgari S, Jung R, Hongskula M, Beckage NE (2005) Par-asitization of Manduca sexta larvae bya the parasitoid wasp Cote-sia congregata induces an impaired host immune response.J Insect Physiol 51:505–512
Askew RR (1971) Parasitic insects. Elsevier, New YorkBaker FC, Tsai LW, Reuter CC, Schooley DA (1987) In vivo Xuctua-
tions of JH, JH acid, and ecdysteroid titer, and JH esterase activ-ity, during development of Wfth stadium Manduca sexta. InsectBiochem 17:989–996
Beckage NE (1998) Modulation of immune responses to parasitoids bypolydnaviruses. Parasitology 116:57–64
Beckage NE, Gelman DB (2001) Parasitism of Manduca sexta by Cotesiacongregata: a multitude of disruptive endocrine eVects. In: EdwardsJP, Weaver RJ (eds) Endocrine interactions of insect parasites andpathogens. Biosis ScientiWc Publishers, Oxford, pp 59–81
Bell RA, Joachim RG (1976) Techniques for rearing laboratory colo-nies of tobacco hornworms and pink bollworms. Ann EntomolSoc Am 69:365–373
Bede JC, McNeil JN, Tobe SS (2007) The role of neuropeptides in cat-erpillar nutritional ecology. Peptides 28:185–196
Cohen RW, Friedman S, Waldbauer GP (1988) Physiological controlof nutrient self-selection in Heliothis zea larvae: the role of sero-tonin. J Insect Physiol 34:935–940
Cohen RW, Mahoney DA, Can HD (2002) Possible regulation of feed-ing behavior in cockroach nymphs by the neurotransmitter octo-pamine. J Insect Behav 15:37–50
Cole TJ, Beckage NE, Tan FF, Srinivasan A, Ramaswamy SB (2002)Parasitoid-host endocrine relations: self-reliance or co-optation?Insect Biochem Mol Biol 32:1673–1679
Combes C (2001) The ecology and evolution of intimate interactions.University of Chicago Press, Chicago
Dahlman DL, Green JR (1981) Larval hemolymph protein patterns intobacco hornworms parasitized by Apanteles congregatus. AnnEntomol Soc Am 74:130–133
Dawkins R (1999) Host phenotypes and parasite genes. In: The extend-ed phenotype/the long reach of the gene. Oxford University Press,Oxford, 336pp
Edwards JP, Weaver RJ, Marris GC (2001) Endocrine changes in lep-idopteran larvae: potential challenges to parasitoid developmentand survival. In: Edwards JP, Weaver RJ (eds) Endocrine interac-tions of insect parasites and pathogens. Biosis ScientiWc Publish-ers, Oxford, pp 1–32
Eggleton P, Gaston KJ (1990) Parasitoid species and assemblages—convenient deWnitions or misleading compromises. Oikos59:417–421
Eggleton P, Belshaw R (1992) Insect parasitoids: an evolutionary over-view. Philos Trans R Soc Lond B 337:1–20

J Comp Physiol B (2008) 178:515–527 527
123
Espagne E, Douris V, Lalmanach G, Provost B, Cattolico, Lesobre J,Kurata S, Iatrou K, Drezen J-M, Huguet E (2005) A virus essen-tial for insect host-parasite interactions encodes cystatins. J Virol79:9765–9776
Ewald PW (1995) The evolution of virulence: a unifying link betweenparasitology and ecology. J Parasitol 81:659–669
Federici BA, Bigot Y (2003) Origin and evolution of polydnavirusesby symbiogenesis of insect DNA viruses in endoparasitic wasps.J Insect Physiol 49:419–432
Fox GA (2001) Failure-time analysis: studying times to events andrates at which events occur. In: Scheiner SM, Gurevitch J (eds)Design and analysis of ecological experiments. Oxford UniversityPress, New York, pp 235–266
Glendinning JI, Slansky F (1994) Interactions of allelochemicals withdietary constituents: eVects on deterrency. Physiol Entomol19:173–186
Hearaty JM (2007) Parasitoid biodiversity and insect pest manage-ment. In: Foottit R, Adler P (eds) Insect biodiversity: science andsociety. Blackwell ScientiWc, Oxford (in press)
Haunerland NH, Shirk PD (1995) Regional and functional diVerentia-tion in the insect fat body. Annu Rev Entomol 40:121–145
Horton DR, Redak RA (1993) Further comments on analysis of covari-ance in insect dietary studies. Entomol Exp Appl 69:263–275
Jones D, Jones G, Rudnicka M, Click A, Reck-Melleczewen V, IwayaM (1986) Pseudoparasitism of host Trichoplusia ni by Chelonusspp. as a new model system for parasite regulation of host physi-ology. J Insect Physiol 32:315–328
Keeley LL (1985) Physiology and biochemistry of the fat body. In: KerkutGA, Gilbert LI (eds) Comprehensive insect physiology, biochemis-try and pharmacology, vol 3. Permagon, Oxford, pp 211–248
Kelly TJ, Gelman DB, Reed DA, Beckage NE (1998) EVects of para-sitization by Cotesia congregata on the brain-prothoracic glandaxis of its host, Manduca sexta. J Insect Physiol 44:323–332
Lee KP, Behmer ST, Simpson SJ, Raubenheimer D (2002) A geomet-ric analysis of nutrient regulation in the generalist caterpillar Spo-doptera littoralis (Boisduval). J Insect Physiol 48:655–665
Poulin R (1995) Adaptive change in the hehaviour of parasitized ani-mals: a critical review. Int J Parasitol 25:1371–1383
Quickie DLJ (1997) Parasitic wasps. Chapman and Hall, LondonRaubenheimer D, Simpson SJ (1992) Analysis of covariance: an alter-
native to nutritional indices. Entomol Exp Appl 62:221–231Raubenheimer D, Simpson SJ (1997) Integrative models of nutrient
balancing: application to insects and vertebrates. Nutr Res Rev10:151–179
Raubenheimer D, Simpson SJ (1999) Integrating nutrition: a geomet-rical approach. Entomol Exp Appl 91:67–82
Raubenheimer D, Simpson SJ (2003) Nutrient balancing in grasshop-pers: behavioural and physiological correlates of dietary breadth.J Exp Biol 206:1669–1681
Roberts LS, Janovy J Jr (2005) Gerald D Schmidt and Larry S Roberts,Foundations of parasitology, 7th edn. McGraw-Hill, Boston
Rogers SM, Matheson T, Sasaki K, Kendrick K, Simpson SJ, BurrowsM. (2004) Substantial changes in central nervous system neuro-transmitters and neruomodulators accompany phase change in thelocust. J Exp Biol 207:3603–3617
Simpson SJ, Raubenheimer D (1993a) A multi-level analysis of feed-ing behaviour: the geometry of nutritional decisions. Philos TransR Soc 342:381–402
Simpson SJ, Raubenheimer D (1993b) The central role of haemolymphin the regulation of nutrient intake in insects. Physiol Entomol18:395–403
Simpson SJ, Raubenheimer D (1996) Feeding behaviour, sensoryphysiology and nutritional feedback: an integrative model. Ento-mol Exp App 80:55–64
Simpson SJ, White P (1990) Associative learning and locust feeding:evidence for a “learned hunger” for protein. Anim Behav 40:506–513
Singer MS, Carriere Y, Theuring C, Hartmann T (2004) Disentanglingfood quality from resistance against parasitoids: diet choice by ageneralist caterpillar. Amer Nat 164: 423–429
Slansky F Jr (1986) Nutritional ecology of endoparasitic insects andtheir hosts: an overview. J Insect Physiol 32:255–261
Slansky F (1992) Allelochemical–nutrient interactions in herbivorenutritional ecology. In: Rosenthal GA, Berenbaum MR (eds) Her-bivores: their interactions with secondary plant metabolites. Aca-demic Press, New York, pp 135–174
Slansky F Jr, Scriber JM (1985) Food consumption and utilization. In:Kerkut GA, Gilbert LI (eds) Comprehensive insect physiology,biochemistry and pharmacology, vol 4. Permagon, Oxford,pp 87–163
Thompson SN (1990) Physiological alterations during parasitism andtheir eVects on host behaviour. In: Barnard CJ, Behnke JM (eds)Parasitism and host behaviour. Taylor and Francis, London,pp 64–94
Thompson SN (1999) Nutrition of entomophagous insects and otherarthropods. In: Bellows TS, Fisher TW (eds) Handbook of biolog-ical control. Academic Press, New York, pp 594–652
Thompson SN (1993) Redirection of host metabolism and eVects onparasite nutrition. In: Beckage NE, Thompson SN, Federici BA(eds) Parasites and pathogens of insects, vol 1. Academic Press,New York, pp 125–144
Thompson SN (2001) Parasitism enhances the induction of glucogen-esis by the insect, Manduca sexta L. Int J Biochem Cell Biol33:163–173
Thompson SN, Redak RA (2005) Feeding behaviour and nutrientselection in an insect Manduca sexta L. and alterations induced byparasitism. J Comp Physiol A 191:909–923
Thompson SN, Redak RA (2007) Nicotine moderates the eVects ofmacronutrient balance on nutrient intake by parasitized Manducasexta L. J Comp Physiol B 177:375–391
Thompson SN, Borchardt DB, Wang L-W (2003) Dietary nutrient lev-els regulate protein and carbohydrate intake, gluconeogenic/gly-colytic Xux and blood trehalose level in the insect Manduca sextaL. J Comp Physiol B 173:149–163
Thompson SN, Redak RA, Borchardt DB (2002) The glucogenic re-sponse of a parasitized insect Manduca sexta L. is partially medi-ated by diVerential nutrient intake. Biochim Biophys Acta1571:138–150
Thompson SN, Redak RA, Wang L-W (2001) Altered dietary nutrientintake maintains metabolic homeostasis in parasitized larvae ofthe insect Manduca sexta L. J Exp Biol 204:4065–4080
Thompson SN, Redak RA, Wang L-W (2005a) Nutrition interacts withparasitism to inXuence growth and physiology of the insect Mand-uca sexta L. J Exp Biol 208:611–623
Thompson SN, Redak RA, Wang L-W (2005b) Host nutrition deter-mines blood nutrient composition and mediates parasite develop-mental success: Manduca sexta L. parasitized by Cotesiacongregata (Say). J Exp Biol 208:625–635
Zanotto FP, Simpson SJ, Raubenheimer D (1993) The regulation ofgrowth by locusts through post-ingestive compensation for varia-tion in the levels of protein and carbohydrate. Physiol Entomol18:425–434
Zanotto FP, Gouveia SM, Simpson SJ, Raubenheimer D, Calder PC(1997) Nutritional homeostasis in locusts: is there a mechanismfor increased energy expenditure during carbohydrate overfeed-ing? J Exp Biol 200:2437–2448














![arXiv:1802.02678v1 [q-bio.NC] 8 Feb 2018 · Hebbian plasticity. ... edge, we have constructed an end-to-end computational model of the Manduca sexta moth olfactory](https://img.pdfslide.us/doc/110x75/5b0475ff7f8b9a6c0b8dd8a4/arxiv180202678v1-q-bionc-8-feb-2018-plasticity-edge-we-have-constructed.jpg)















