Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Acta Oecologica 49 (2013) 83e91
Contents lists available
Acta Oecologica
journal homepage: www.elsevier .com/locate/actoec
Original article
Effects of sea level rise on mangrove Avicennia population growth,colonization and establishment: Evidence from a field survey andgreenhouse manipulation experiment
Weizhi Lu a, Luzhen Chen a,*, Wenqing Wang a, Nora Fung-Yee Tamb, Guanghui Lin a,c,**
aKey Laboratory of the Ministry of Education for Coastal and Wetland Ecosystems, College of the Environment and Ecology, Xiamen University, Xiamen,Fujian 361005, ChinabDepartment of Biology and Chemistry, City University of Hong Kong, Tat Chee Avenue, Kowloon, Hong KongcKey Laboratory of the Ministry of Education for Earth System Modelling, Center for Earth System Science, Tsinghua University, Beijing 100084, China
a r t i c l e i n f o
Article history:Received 24 July 2012Accepted 12 March 2013Available online 10 April 2013
Keywords:Accelerated sea level riseColonizationEstablishmentGrowthMangrovesWater use efficiency
* Corresponding author. Tel.: þ86 592 2181430; fax** Corresponding author. Key Laboratory of the Minand Wetland Ecosystems, College of the Environmenversity, Xiamen, Fujian 361005, China. Tel.: þ8662797230.
E-mail addresses: [email protected] (L. C(G. Lin).
1146-609X/$ e see front matter � 2013 Elsevier Mashttp://dx.doi.org/10.1016/j.actao.2013.03.009
a b s t r a c t
Mangroves will either face longer tidal inundation or retreat landwards in response to on-going accel-erated sea level rise. However, little is known about the growth, regeneration or colonization ofmangrove seedlings under the different tidal inundation regimes associated with accelerated sea levelrise. In the present study, a field survey and a greenhouse mesocosm experiment were conducted toevaluate possible effects of accelerated sea level rise on colonization, establishment and seedling growthof a mangrove pioneer species, Avicennia marina. Avicennia populations at different elevations of theintertidal zone on Xiamen Island in Fujian Province, China showed distinctive patterns in both seedlingdensity and plant age. The seedlings at lower elevations had less annual biomass accumulation andpopulation productivity, but higher shoot to root ratios, suggesting that elevation has positive effects onseedling growth. The greenhouse mesocosm experiment with 1-year-old A. marina seedlings utilized fiveinundation periods (0, 2, 4, 6 and 12 h in a semidiurnal tidal cycle) and two inundation depths (rootimmersion and canopy immersion). Both inundation period and depth exerted significant and negativeeffects on biomass accumulation, photosynthetic rate, leaf electron transportation and water use effi-ciency. However, the negative effects of canopy immersion were more profound than root immersion.Canopy immersion exacerbated the effects of prolonged inundation, with no seedlings surviving undercanopy immersion at the 12-hour treatment. These results suggest that at lower elevations with highersea level, canopy immersion will have greater negative effects on seedling colonization, establishmentand early growth of A. marina. This finding is instrumental in predicting the future dynamics of mangroveforests under increasing sea levels.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
Mangrove ecosystems are very sensitive to global changesbecause they are located at the interface between land and sea(Duke et al., 1998; Day et al., 2008). Of all climate change-relatedeffects, sea level rise is the most serious threat to mangroves(Gilman et al., 2008; Chin et al., 2010). Global sea level rise had been
: þ86 592 2181430.istry of Education for Coastalt and Ecology, Xiamen Uni-10 62797230; fax: þ86 10
hen), [email protected]
son SAS. All rights reserved.
occurring for more than 20,000 years, however rates of sea levelrise appear to be increasing rapidly due to anthropogenic impacts(Erlandson, 2012). Coastal areas of China are especially vulnerableto accelerated sea level rise and associated coastal flooding becauseof low topography and socioeconomic factors such as highlydeveloped economies and dense populations (Yin et al., 2012). Insoutheast China, mangroves are also affected by accelerated sealevel rise. If sediment surface elevations do not keep pace with sealevel rise, mangroves will either retreat landwards or face longperiods of tidal inundation (Gilman et al., 2008; Krauss et al., 2008;Abel et al., 2011). If mangroves retreat landwards, there will beincreased opportunity for seedling regeneration and colonization(Panapitukkul et al., 1998; Abel et al., 2011). The natural regenera-tion of a forest plays an important role in community expansion,dispersion and maintenance, as well as in maintaining community

Fig. 1. (A) Location of field survey sample plots. E1 to E5 indicate the five different elevations of the plots: 105, 95, 65, 55, and 45 cm above mean sea level, respectively. (B)Experimental set-up for the greenhouse manipulation experiment showing the tidal tanks used for different immersion treatments. Hollow arrows indicate water flowing betweentanks B to E, water retreating back to the water container and water reentering the tanks from the water container; h indicates inundation period (hour); RI: root immersion, CI:canopy immersion; and the dotted line indicates water level.
W. Lu et al. / Acta Oecologica 49 (2013) 83e9184
stability and succession (Martinkova et al., 2011). Studies on theregeneration and colonization of mangrove forests at different el-evations can provide new insights into how mangrove seedlingsrespond to tidal inundation and accelerated sea level rise.
Previous studies on the potential impact of accelerated sea levelrise on mangrove forests focused on growth, physiology,morphology, anatomy and reproduction, and mainly utilizedmanipulation experiments with water inundation treatments(Ellison and Farnsworth, 1997; Naidoo et al., 1997; Kitaya et al.,2002; Chen et al., 2005; He et al., 2007; Hoppe-Speer et al.,2011). These studies revealed that, with prolonged inundation,mangroves allocated more relative biomass to abovegroundstructures or grew taller as mechanisms for avoiding tidal inun-dation. However, these studies focused mainly on the effect of tidalinundation periods (Ellison and Farnsworth, 1993; Buckley et al.,1998; Kitaya et al., 2002; Chen et al., 2005; Krauss et al., 2008). Inactuality, water inundation depth as well as inundation period willsimultaneously increase with accelerated sea level rise, and thesetwo factors may have different effects on the physiology andgrowth of different mangrove species due to different distributionsin the intertidal zone. Previous research focused either on inun-dation depth or inundation period, and thus do not provide acomplete picture for mangroves in different intertidal zones andwith different canopy heights, in terms of their potential sensitivityto accelerated sea level rise. Only a few studies were conducted toinvestigate mangrove seedling colonization and establishmentunder accelerated sea level rise scenario (Ye et al., 2004; McKeeet al., 2007; Di Nitto et al., 2008; Hickey and Bruce, 2010), andstudies of population structure and productivity responses (e. g.
forest density, tree height, biomass increment) were rare. Thus,there remain many knowledge gaps about the growth, regenera-tion and colonization of mangrove seedlings as sea level rise ac-celerates in response to global warming.
In this study, we investigated the following questions: (1) howdo mangrove population structure and productivity change withaccelerated sea level rise? (2) Is inundation depth or inundationperiod correlated to the observed negative effects of tidal inunda-tion on mangrove seedling growth and colonization? We hypoth-esized that (1) the growth, population structure and productivitywould be devastated with the accelerated sea level rise; (2) inun-dation depth exerts a stronger influence on seedlings facingaccelerated sea level rise.
Avicennia marina was selected in this study because it is awidespread mangrove species with a large distribution in SouthAsia and Australia (Duke, 1991; Lin, 1999). Compared with othermangrove species, A. marina occupy more seaward habitats andexhibit relatively higher waterlogging tolerance, achieving highersurvival over a wider tidal range (Ashford and Allaway, 1995; Lin,1999; He et al., 2007; Xiao et al., 2009). In this study, mangrovegrowth and biomass were investigated in a field survey to deter-mine the effects of accelerated sea level rise on population struc-ture and productivity. The separate and interactive effects ofinundation period and depth on the growth and leaf physiology ofA. marina seedlings were investigated in greenhouse manipulationexperiments. The greenhouse investigation focused only on seed-lings since they are more likely to be immersed if sea levels meetthe projection of rising at least 30 cm over current levels by 2100(IPCC, 2007; Kemp et al., 2011).

W. Lu et al. / Acta Oecologica 49 (2013) 83e91 85
2. Materials and methods
2.1. Description of study sites
The study sites for the population survey were located in anabandoned fish pond, adjacent to a mudflat on the northwest coastof Xiamen Island, Fujian Province of China (24�320 N, 118�060 E,Fig.1A). The pond area was greater than 1 ha, and had been un-dergoing natural succession for more than 10 years. The mangroveforest was dominated by A. marina with most of individuals lessthan 10 years old. The mean annual temperature in Xiamen wasestimated to be 20.7 �C and the mean annual rainfall was 1387 mmaccording to observations of the Xiamen Meteorological Observa-tory (Chen, 2005). The tidal regime at the site was regular semi-diurnal with an average tidal range of 3.96 m, and a high waterdepth of 5.39 m and low water depth of 1.43 m (Chen, 2005).
2.2. Field survey and sampling
The field sample plots were divided into five classes according totidal elevation,where Plot E1 represented the highest elevation, andPlot E5 represented the lowest elevation. For eachplot,wemeasuredwater depth at tidal flood slack periods, and then calculated eleva-tions according to tidal prediction tables. Elevations were 105, 95,65, 55, and 45 cm above mean sea level for Plot E1 through E5,respectively. For each elevation plot except E4, four replicate sub-plots were surveyed (Fig. 1A). Because of the area limitation, PlotE4 had only three replicates. After determining the number of treesin each plot (Plot E1 to E5), all treeswithin the plotswere tagged andthe population density was calculated. In addition, tree height, stembase diameter and canopy breadth were also measured.
One standard tree in each plot was selected and harvested. Thestem, leaves, flowers and roots of each sampled tree were separated.We determined the age of each tree roughly according to stem rings.Although some research indicates that A. marina do not have annualrings (Tomlinson, 1995), other sources point out that in subtropicalmangroves, stem rings are coincident with annual rings (Yu et al.,2007). In our rough estimate for tree age, we accepted stem ring asthe annual ring. Biomasswasdetermined afterovendrying for 72hat60 �C after constant weights were attained. Annual biomass incre-ment (ABI), population productivity (PP) and biomass ratios of shootto root (S/R) were calculated according to the following equations.
Annual biomass increment ðABIÞ ¼ Total BiomassTree Age
Population productivity ðPPÞ ¼ Total plant biomassForest area� Tree age
Shoot=root ratio�SR
�¼ Aboveground biomass
Belowground biomass
2.3. Soil measurements
In the field survey, five soil samples were collected in each plotto measure the soil pH, salinity, bulk density, organic carbon andsoil texture. Only one soil sample was measured in Plot E5 becauseof the high homogeneity revealed in our preliminary experiment.Soil pH and salinity were measured by portable pH/salinity indi-cator (Thermo Electron Corporation, USA). Bulk density wasdetermined by the volumetric ring method (Blake, 1965). Volu-metric ring samples were collected using 90 cm3 sharpened rings at0e20 cm depth. Samples were oven-dried at 80 �C for 72 h and the
dry weight was used to calculate soil bulk density. Subsamples ofsoil were air-dried, ground and sieved to a fine power for thedetermination of total organic carbon (TOC) and soil texture. TOCwas determined on acidified samples by standard methods using aLeco induction furnace (Blakemore, 1987). The clay, silt and sandpercentage of soil samples were determined using the plummetbalance method (Hutton, 1955).
2.4. Seedling preparation for greenhouse manipulation experiment
Healthy and mature viviparous propagules of A. marina werecollected from Zhangjiang Estuary Mangrove Nature Reserve inZhangzhouCountry, Fujian Province, China (23�550 N,117�240 E). Thepropagules were grown in a greenhouse at Xiamen University,which had an average daily temperature of 24 � 8 �C and relativehumidity of 80 � 10% during the treatment days. The mid-dayphotosynthetic active radiation (PAR) was about 700 mmol pho-tons m�2 s�1, during the sunny treatment days which weremeasured from 10:00 to 16:00. The propagules were irrigated everytwo days with artificial seawater with a salinity of 15&, similar tothe field mangrove areas salinity of 17&. The artificial seawater wasprepared by diluting seawater collected from the west coast ofXiamen, with salinity ranging from 22 to 28&, with local tap water.When the seedlings developed the first two pairs of leaves, andheight was 16.4 � 0.2 cm (mean and standard error, n ¼ 20), theywere transplanted to pots and placed in the tidal tank systemdescribed below.
2.5. Experimental design of greenhouse studies
According to our field observations, water inundation durationin mangrove habitats in Southern China is generally less than 6-hour in a tidal cycle. The inundation period during a single tidalcycle varies depending on the elevation of the intertidal area. Thefour inundation periods for the selected treatments were 0, 2, 4,and 6-hour during a semidiurnal tidal cycle. A 12-hour inundationtreatment was also included as the fifth inundation period tosimulate the scenario that seedlings were inundated throughout anentire semidiurnal tidal cycle, which represented the longest waterinundation. Each inundation period treatment had two differentwater inundation depths e root immersion (RI) and canopy im-mersion (CI) e to simulate current conditions and future acceler-ated sea level rise scenarios for mangroves at low and mid-intertidal zones. All seedlings were inundated twice daily exceptthe 0-hour inundation treatment.
To simulate tidal inundation conditions with different inunda-tion depths and periods, a tide tank system using six plastic tanks(65 cm � 50 cm � 50 cm) was constructed as Fig. 1B, also refer toChen et al. (2005). Tank A held artificial seawater throughout thetidal cycle to simulate the 12-hour inundation. PVC pipes connectedTank B through Tank E. The total time to fill each tank with artificialseawater to the predetermined water level was two hours. Tank B,which simulated the 6-hour inundation filled with artificialseawater from the water container first. When Tank B was full, thewater overflow spilled intoTank C, and so on. When Tank E was full,all of thewater in Tanks B, C, D and E discharged back into thewatercontainer via timer-controlled pumps. The seawater in the tidaltank system was renewed every two weeks to reduce contamina-tion. To achieve two different inundation depths, seedlings sub-jected to the RI treatment were elevated by a 30 cm high platformwhile those subjected to the CI treatment were placed in the bot-tom of the tank (Fig. 1B). There were 10 pots of seedlings in eachtidal tank, with half on the platform (RI) and the other half on thebottom of the tank (CI). For the RI treatment, water submerged theseedling to the junction between the stem and the root, while for

Table 1Topsoil physical and chemical properties and vegetation structure of A. marina population in the field plots.
Samplegroup
Elevation(cm)
pH value Soil salinity(&)
Soil bulkdensity (g cm�3)
Organic carboncontent (%)
Soil texturetype
Seedlingheight (cm)
Canopybreadth (cm2)
Vegetationcover (%)
E1 105 6.81 � 0.04c 19.42 � 1.42c 1.13 � 0.03b 3.25 � 0.09b Silt clay 83.2 � 4.74a 559 � 36.8a 5.59 � 0.74aE2 95 6.96 � 0.01b 25.4 � 1.19b 1.17 � 0.02b 3.49 � 0.11b Silt clay 91.0 � 3.73a 498 � 67.6a 1.97 � 0.10bE3 65 6.98 � 0.03b 31.1 � 0.36a 1.08 � 0.01b 3.44 � 0.13b Silt clay 39.7 � 3.60b 116 � 23.8b 1.69 � 0.25bE4 55 7.12 � 0.03a 20.5 � 1.68c 1.25 � 0.07a 2.91 � 0.05a Clay 48.7 � 2.39b 115 � 4.42b 0.24 � 0.08cE5 45 7.06 25.1 1.22 2.48 Silt clay 30 41.6 0.01
Fig. 2. Changes in seedling density (A) and age (B) of A. marina under different tidalelevation in the field. A fitted curve is shown, where R2 ¼ 0.53 and 0.88 for seedlingdensity and seedling age respectively, both p < 0.001. E1 to E5 indicate the fivedifferent elevations of the survey plots, which were 105, 95, 65, 55, and 45 cm abovemean sea level, respectively.
W. Lu et al. / Acta Oecologica 49 (2013) 83e9186
the CI treatment, the whole seedling, including the canopy wascompletely submerged (Fig. 1B). Each inundation treatment hadthree replicates. Each pot, with a dimension of 13-cm height and12-cm diameter, contained one seedling with two pairs of matureand fully expanded leaves. All seedlings remained in the green-house for 100 days. After 100 days of treatment, each seedling inTanks B, C, D, E and F had four to five pairs of leaves, however, thoseunder the CI treatment in Tank A were all dead at the end of theexperiment. This indicated that they could not stand the 12-hourinundation period combined with canopy immersion.
2.6. Measurements form the greenhouse manipulation experiment
2.6.1. Biomass and growthAll seedlings were harvested at the end of the greenhouse
manipulation experiments. After separating the leaves, stems androots, the fresh weight of each part was determined using anelectronic balance (Sartorius BS, Goettingen, Germany). The sam-ples were then oven-dried to constant weight at 60 �C for 72 h andre-weighed. Seedling height was measured using a ruler. The totalleaf area was calculated using the weighing paper cut-outs method(Fisher and Honda,1977). Specific leaf area (SLA) was defined as leafarea per unit dry weight.
2.6.2. Leaf gas exchangeFor the greenhouse experiment, photosynthetic rates (Pn) and
transpiration rates (Tr) of mature leaves were measured using aportable infrared gas analyzer (LI-6400, Li-cor, Inc., Lincoln, USA) after76 days treatments (on 17 December, 2008). In order to access theleaves of plants under the CI treatment, water was drained from thetanks until the leaves were above the water. Gas exchange measure-ments were made on fully expanded leaves, which were conductedbetween 9:00 am and 11:00 am. The PAR was fixed at1000 mmol m�2 s�1 by a blue/red light (LI-6400-02 LED light, Li-cor,Inc.) source fitted to the leaf chamber and an ambient CO2 concen-tration was set at 380 � 2 mmol mol�1. Instantaneous water-use effi-ciency (WUEi) was calculated from the ratio of Pn:Tr, and stomatallimitation (Ls) was defined as 1 � Ci/C0, with Ci representing inter-cellular CO2 concentration and C0 for atmospheric CO2 concentration.Three mature leaves for each replicate were chosen for the mea-surement of Pn and Tr, and the mean values were used for statisticalanalyses.
2.6.3. Chlorophyll fluorescenceChlorophyll fluorescence was measured at night for the cultured
seedlings with a field-portable, pulse amplitude, modulated fluo-rometer (PAM-2100, Walz, Effeltrich, Germany). All measurementswere taken midway between the base and the tip of mature leaves.Rapid light curves (RLCs)were generatedwith the PAM-2100 (Whiteand Critchley, 1999; Naidoo et al., 2010). RLCs were plots of electrontransport rate (ETR) versus actinic irradiances, applied for 10 s. ThePAR was set for the following light levels: 25, 50, 100, 150, 250, 500,1000 and 1200 mmol m�2 s�1. The potential maximum electrontransport rate (ETRmax), efficient use of light quantum (a) and photo-inhibition (b) was calculated using the equation: ETR ¼ ETRmax$ð1�
e�a$PAR=ETRmax Þ$e�b$PAR=ETRmax (modified from Platt et al. (1980)). Thehalf-saturation light intensity (Ik) was calculated by ETRmax/a.
2.7. Statistical analysis
Data were examined for normality and homoscedasticity of re-sidual variances. Repeated measures analysis of variance was usedto compare field survey measures of ABI, PP, and S/R ratios changeover elevation. For the greenhouse measurements, mean andstandard error (SE) values of three replicates were calculated foreach of all measured parameters. A parametric two-way analysis ofvariance (ANOVA) was used to test significant differences amonginundation periods and between inundation depths. If the differ-ence was significant at p � 0.05, a post-hoc test was employed todetermine exactly where the difference was. The assumptions ofANOVA, including normal distribution and homogenous variance ofthe data, were tested. A multivariate ANOVA using the generallinear model was used to demonstrate the interactive effects be-tween inundation depth and period on various photosynthetic gasexchange parameters, growth and chlorophyll fluorescence. Linearregression analysis was used to evaluate the relationships betweenLs andWUEi. The correlation between density and seedling agewasalso evaluated for the field investigation. All statistical analyseswere performed with SPSS version 13.0 (SPSS Inc., Chicago, USA).
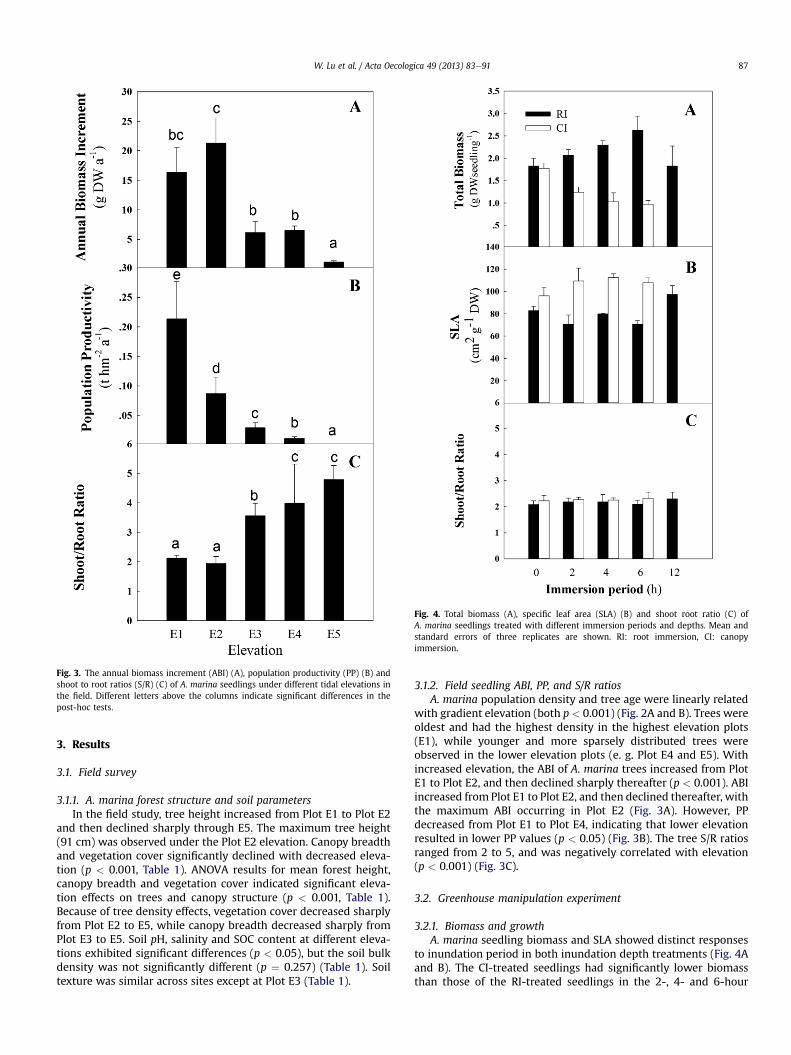
Fig. 3. The annual biomass increment (ABI) (A), population productivity (PP) (B) andshoot to root ratios (S/R) (C) of A. marina seedlings under different tidal elevations inthe field. Different letters above the columns indicate significant differences in thepost-hoc tests.
Fig. 4. Total biomass (A), specific leaf area (SLA) (B) and shoot root ratio (C) ofA. marina seedlings treated with different immersion periods and depths. Mean andstandard errors of three replicates are shown. RI: root immersion, CI: canopyimmersion.
W. Lu et al. / Acta Oecologica 49 (2013) 83e91 87
3. Results
3.1. Field survey
3.1.1. A. marina forest structure and soil parametersIn the field study, tree height increased from Plot E1 to Plot E2
and then declined sharply through E5. The maximum tree height(91 cm) was observed under the Plot E2 elevation. Canopy breadthand vegetation cover significantly declined with decreased eleva-tion (p < 0.001, Table 1). ANOVA results for mean forest height,canopy breadth and vegetation cover indicated significant eleva-tion effects on trees and canopy structure (p < 0.001, Table 1).Because of tree density effects, vegetation cover decreased sharplyfrom Plot E2 to E5, while canopy breadth decreased sharply fromPlot E3 to E5. Soil pH, salinity and SOC content at different eleva-tions exhibited significant differences (p < 0.05), but the soil bulkdensity was not significantly different (p ¼ 0.257) (Table 1). Soiltexture was similar across sites except at Plot E3 (Table 1).
3.1.2. Field seedling ABI, PP, and S/R ratiosA. marina population density and tree age were linearly related
with gradient elevation (both p < 0.001) (Fig. 2A and B). Trees wereoldest and had the highest density in the highest elevation plots(E1), while younger and more sparsely distributed trees wereobserved in the lower elevation plots (e. g. Plot E4 and E5). Withincreased elevation, the ABI of A. marina trees increased from PlotE1 to Plot E2, and then declined sharply thereafter (p < 0.001). ABIincreased from Plot E1 to Plot E2, and then declined thereafter, withthe maximum ABI occurring in Plot E2 (Fig. 3A). However, PPdecreased from Plot E1 to Plot E4, indicating that lower elevationresulted in lower PP values (p < 0.05) (Fig. 3B). The tree S/R ratiosranged from 2 to 5, and was negatively correlated with elevation(p < 0.001) (Fig. 3C).
3.2. Greenhouse manipulation experiment
3.2.1. Biomass and growthA. marina seedling biomass and SLA showed distinct responses
to inundation period in both inundation depth treatments (Fig. 4Aand B). The CI-treated seedlings had significantly lower biomassthan those of the RI-treated seedlings in the 2-, 4- and 6-hour
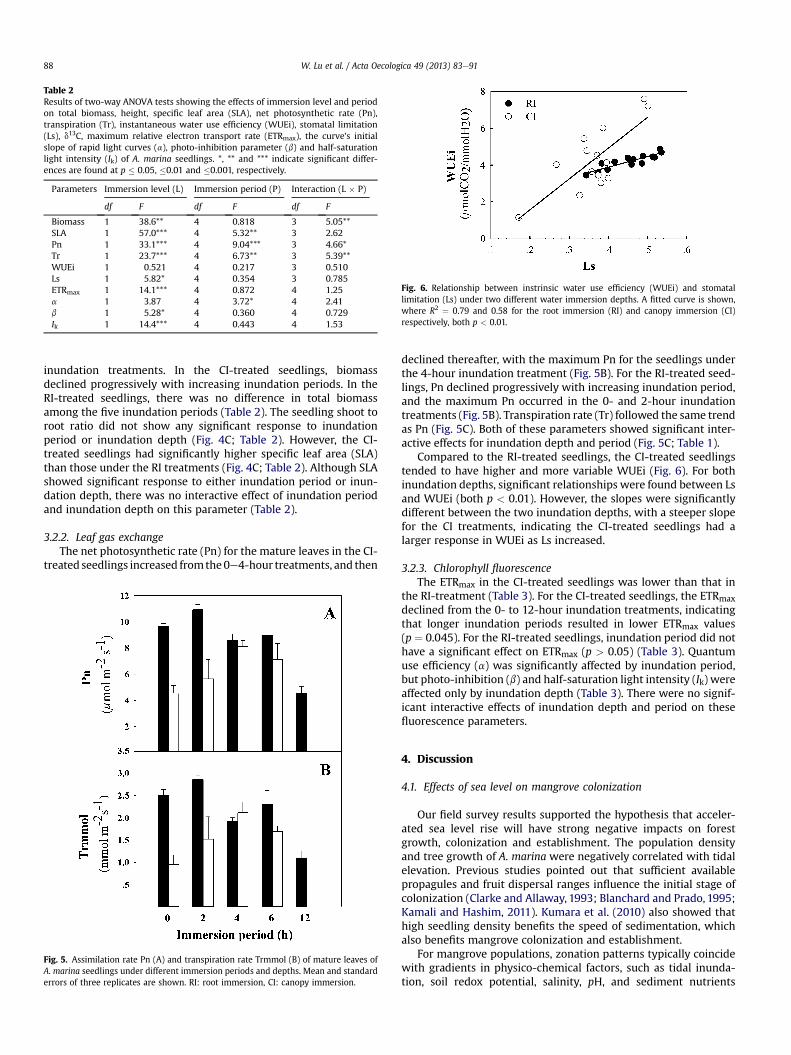
Table 2Results of two-way ANOVA tests showing the effects of immersion level and periodon total biomass, height, specific leaf area (SLA), net photosynthetic rate (Pn),transpiration (Tr), instantaneous water use efficiency (WUEi), stomatal limitation(Ls), d13C, maximum relative electron transport rate (ETRmax), the curve’s initialslope of rapid light curves (a), photo-inhibition parameter (b) and half-saturationlight intensity (Ik) of A. marina seedlings. *, ** and *** indicate significant differ-ences are found at p � 0.05, �0.01 and �0.001, respectively.
Parameters Immersion level (L) Immersion period (P) Interaction (L � P)
df F df F df F
Biomass 1 38.6** 4 0.818 3 5.05**SLA 1 57.0*** 4 5.32** 3 2.62Pn 1 33.1*** 4 9.04*** 3 4.66*Tr 1 23.7*** 4 6.73** 3 5.39**WUEi 1 0.521 4 0.217 3 0.510Ls 1 5.82* 4 0.354 3 0.785ETRmax 1 14.1*** 4 0.872 4 1.25a 1 3.87 4 3.72* 4 2.41b 1 5.28* 4 0.360 4 0.729Ik 1 14.4*** 4 0.443 4 1.53
Fig. 6. Relationship between instrinsic water use efficiency (WUEi) and stomatallimitation (Ls) under two different water immersion depths. A fitted curve is shown,where R2 ¼ 0.79 and 0.58 for the root immersion (RI) and canopy immersion (CI)respectively, both p < 0.01.
W. Lu et al. / Acta Oecologica 49 (2013) 83e9188
inundation treatments. In the CI-treated seedlings, biomassdeclined progressively with increasing inundation periods. In theRI-treated seedlings, there was no difference in total biomassamong the five inundation periods (Table 2). The seedling shoot toroot ratio did not show any significant response to inundationperiod or inundation depth (Fig. 4C; Table 2). However, the CI-treated seedlings had significantly higher specific leaf area (SLA)than those under the RI treatments (Fig. 4C; Table 2). Although SLAshowed significant response to either inundation period or inun-dation depth, there was no interactive effect of inundation periodand inundation depth on this parameter (Table 2).
3.2.2. Leaf gas exchangeThe net photosynthetic rate (Pn) for the mature leaves in the CI-
treated seedlings increased fromthe0e4-hour treatments, and then
Fig. 5. Assimilation rate Pn (A) and transpiration rate Trmmol (B) of mature leaves ofA. marina seedlings under different immersion periods and depths. Mean and standarderrors of three replicates are shown. RI: root immersion, CI: canopy immersion.
declined thereafter, with the maximum Pn for the seedlings underthe 4-hour inundation treatment (Fig. 5B). For the RI-treated seed-lings, Pn declined progressively with increasing inundation period,and the maximum Pn occurred in the 0- and 2-hour inundationtreatments (Fig. 5B). Transpiration rate (Tr) followed the same trendas Pn (Fig. 5C). Both of these parameters showed significant inter-active effects for inundation depth and period (Fig. 5C; Table 1).
Compared to the RI-treated seedlings, the CI-treated seedlingstended to have higher and more variable WUEi (Fig. 6). For bothinundation depths, significant relationships were found between Lsand WUEi (both p < 0.01). However, the slopes were significantlydifferent between the two inundation depths, with a steeper slopefor the CI treatments, indicating the CI-treated seedlings had alarger response in WUEi as Ls increased.
3.2.3. Chlorophyll fluorescenceThe ETRmax in the CI-treated seedlings was lower than that in
the RI-treatment (Table 3). For the CI-treated seedlings, the ETRmaxdeclined from the 0- to 12-hour inundation treatments, indicatingthat longer inundation periods resulted in lower ETRmax values(p ¼ 0.045). For the RI-treated seedlings, inundation period did nothave a significant effect on ETRmax (p > 0.05) (Table 3). Quantumuse efficiency (a) was significantly affected by inundation period,but photo-inhibition (b) and half-saturation light intensity (Ik) wereaffected only by inundation depth (Table 3). There were no signif-icant interactive effects of inundation depth and period on thesefluorescence parameters.
4. Discussion
4.1. Effects of sea level on mangrove colonization
Our field survey results supported the hypothesis that acceler-ated sea level rise will have strong negative impacts on forestgrowth, colonization and establishment. The population densityand tree growth of A. marina were negatively correlated with tidalelevation. Previous studies pointed out that sufficient availablepropagules and fruit dispersal ranges influence the initial stage ofcolonization (Clarke and Allaway, 1993; Blanchard and Prado, 1995;Kamali and Hashim, 2011). Kumara et al. (2010) also showed thathigh seedling density benefits the speed of sedimentation, whichalso benefits mangrove colonization and establishment.
For mangrove populations, zonation patterns typically coincidewith gradients in physico-chemical factors, such as tidal inunda-tion, soil redox potential, salinity, pH, and sediment nutrients

Table 3Results of relative electron transport rate curve regression by ETR ¼ ETRmax$ð1� e�a$PAR=ETRmax Þ$e�b$PAR=ETRmax for A. marina seedlings under different inundation periods anddepths (RI: root immersion; CI: Canopy immersion).
Parameter 0 h 2 h 4 h 6 h 12 h
ETRmax RI 57.1 � 8.89 58.0 � 5.35 62.7 � 8.85 61.3 � 3.10 50.3 � 1.84CI 52.6 � 1.92 50.0 � 2.15 47.0 � 1.79 43.7 � 2.61 47.3 � 3.72
a RI 0.33 � 0.01 0.33 � 0.01 0.33 � 0.02 0.34 � 0.01 0.32 � 0.01CI 0.32 � 0.00 0.37 � 0.01 0.35 � 0.01 0.35 � 0.01 0.32 � 0.01
b RI 0.00 � 0.01 0.00 � 0.00 0.01 � 0.01 0.00 � 0.00 0.00 � 0.00CI 0.01 � 0.00 0.01 � 0.00 0.01 � 0.00 0.01 � 0.00 0.01 � 0.00
Ik RI 174 � 23.8 179 � 17.9 194 � 35.7 184 � 16.8 158 � 8.21CI 167 � 7.88 137 � 8.00 135 � 7.35 126 � 6.66 150 � 12.3
ETRmax: maximum relative electron transport rate; a: the curve’s initial slope of rapid light curves; b: photo-inhibition parameter; Ik: half-saturation light intensity.
Fig. 7. Correlation between plant productivity and tidal elevation for A. marina seed-lings in the greenhouse manipulation experiment and in the field. Note that manip-ulation elevations were inferred according to their respective immersion periods.Numbers above the curve represent the immersion period in a tidal cycle for seedlingsin the greenhouse experiment.
W. Lu et al. / Acta Oecologica 49 (2013) 83e91 89
(McKee, 1995). Inundation depth and duration depend on seedlingposition in the tidal zones (Ye et al., 2003; Krauss et al., 2008; Yeet al., 2010). Present research suggested that the soil environmentincluding pH, salinity and SOC content at high elevation was infavor of the seedling colonization and growth. Our results suggestthat different elevations in the tidal zones result in different foreststructure patterns, even though there were sufficient propagules inall plots. As shown in Fig. 2, seedlings density and average agegradually increased with increasing elevation, implying that higherelevation benefits to seedling colonization and growth.
As sea level rise accelerates, the elevation of mangroves maydescend to a lower level (e.g. the elevation of Plot E1 will shift tothat of Plot E3), which would severely affect the mangrove com-munity at the lower elevation (Plot E5), as seedlings would die as aresult of serious waterlogged stress (Ross et al., 2000; Sanders et al.,2008; Rivera-Monroy et al., 2011). At the same time, some propa-gules in lower plots will dispersed to higher elevation with shortertidal immersion periods (e.g. the shift from Plot E4 to Plot E2).However, dams and levees built landward of many mangrove for-ests would prevent the retreat of mangrove forests (Fan and Li,1997; Colonnello and Medina, 1998). This provides further evi-dence that accelerated sea level rise would limit seedling’s colo-nization, establishment, and growth of mangroves.
4.2. Impact of canopy immersion and prolonged inundation onmangrove seedling growth
The present study also demonstrated that, when compared withprolonged inundation, canopy immersion exerted greater negativeimpacts on biomass accumulation and almost all of the leaf phys-iological parameters measured for the A. marina seedlings. Thissupports the hypothesis that complete canopy immersion byseawater will have greater negative effects on the growth and leafphysiology of mangrove seedlings than will prolonged inundation.If inundation occurs only in the root systems, A. marina seedlingssurvived and maintained a reasonable growth, even in non-stopinundation (i.e. the 12-hour inundation in a semi-diurnal tidal cy-cle). However, when the canopy was immersed for 2 h or longer,seedlings of the same species either could not survive the 12-hourinundation treatment (after 10 weeks) or maintained significantlylower growth. Ashford and Allaway (1995) reported that the innerparts of A. marina seedlings contain many gas spaces connected toeach other that form a continuous exchange with the atmosphere.When the roots are immersed, the gas space continuum will haveaccess to the atmosphere through the stomata, and leaves can serveas a source of photosynthetically-produced oxygen for the seed-lings (Laan and Blom, 1990). However, in complete canopy im-mersion, the gas exchange is blocked which seriously inhibitsphotosynthetic assimilation and respiration.
Our results indicated that A. marina seedlings could maintainnormal or increased growth and physiological processes under the
2- through 6-hour tidal inundation periods. This suggested that, asa pioneer species, A. marina adapts well to changing inter-tidalhabitats and develops tolerance to various tidal inundations. Heet al. (2007) also reported that A. marina seedlings on the lowerwater tidal flats grew quicker in terms of height, diameter, leafproduction and biomass. They also accumulated more neonatalbiomass than those on the mean water mudflats. Similar resultswere found for other mangrove species, such as Rhizophora mangle,Kandelia obovata and Bruguiera gymnorrhiza (Ellison andFarnsworth, 1993, 1997; McKee, 1996; Kitaya et al., 2002; Ye et al.,2003, 2004; Chen et al., 2004; Cardona-Olarte et al., 2006). Theseresults indicated that the duration of inundation affected thegrowth and development of mangrove seedlings. Hovenden et al.(1995) demonstrated that, in younger mangrove plants, there wassufficient aerenchyma to supply the oxygen requirements of theroot system for a period of about 1.5e3.5 h per tide. When theinundation periods exceed this time, the plants become anaerobicand consume more energy in order to maintain their metabolisms.
The A. marina seedling biomass accumulation, as well as manyfoliar photosynthetic properties such as net photosynthetic rate,transpiration rate and stomatal conductance showed significantinteractive effects between inundation period and depth. Thissuggested that future accelerated sea level rise is likely to reducethe photosynthesis and growth of A. marina seedlings due to thesynergistic effects of tidal inundation period and depth. Such im-pacts were less profound than, or different from, previous studieswhen only either inundation period or inundation depth wasconsidered. In the present study, there were no significant differ-ences in seedling growth among different inundation periods;however, there were significant differences between the twoinundation depths.

W. Lu et al. / Acta Oecologica 49 (2013) 83e9190
4.3. Effects of accelerated sea level rise on mangrove productivity
Both the field survey and manipulation experiment demon-strated that mangrove productivity was sensitive to predicted as-pects of accelerated sea level rise. We found that the tidal elevationin the field ranged from 45 to 110 cm, corresponding to 4e6 h oftidal immersion in the greenhouse.
A linear relationship between plant productivity and tidal eleva-tion was found in the greenhouse experiment (Fig. 7). For bothinundation depth treatments (RI and CI), there was an oppositetendency in this relationship (see part 4.2). However, we found thatproductivity increasedwith elevation, and then decreasedwhen theelevation exceeded some degree in the field survey (Fig. 7). Withincreasingelevation, the range of variation inproductivity in thefieldwas larger than in the manipulated experiment. This was attributedto nutrient (mainly nitrogen) limitation. Generally, mangroves areconsidered to be limited by nitrogen (McKee et al., 2002; Lovelocket al., 2006). For the seawater used in our manipulation experi-ment, the inorganic nitrogen (IN) concentration was 54.2 mmol/L(unpublished data), and the total IN input during the 100-day cultureperiod for seedlings was 3.04 g m�2. In the field survey, the soil INpool was 3.22 g m�2 (to the depth of 1 m, unpublished data).Considering tidal water IN input, in the total IN input in the fieldwasabout twice that of the inmanipulation experiment. This may be thereason why field seedlings have higher growth rates at comparableelevations than greenhouse growth seedlings. Thus, nutrient inputsshould be considered in further research.
We also found it interesting that the suitable range of mangroveelevation in the field was very narrow when compared to themanipulation experiment results. At elevations lower than 45 cm inthe field, low seedling productivity implied extreme conditions offuture accelerated sea level rise, as seen in the most extrememanipulated condition. However, therewere fewA.marina seedlingsdistributed at elevation higher than 110 cm at our field site. There-fore, the future researches onmangrove and sea level change shouldintegrate the immersion period with the tidal regime together.
5. Conclusions
The present study indicated that the colonization and establish-mentofA.marinawouldbe seriouslyaffectedbyaccelerated sea levelrise. We concluded that the growth and physiological characteristicsof A. marina seedlings were determined by the interactive effects ofinundation depth and inundation period, and that inundation depthwas a more important limiting factor than inundation period. Aspredicted by IPCC (2007), global sea level may rise by at least 30 cmby 2100, and A. marina seedlings may suffer negative impacts fromaccelerated sea level rise in the near future. Accelerated sea level risewill occur graduallybut there is little information so far regarding thepossible acclimation potential of mangroves to gradual changes insea levels. In order to better predict the dynamics of mangrovecommunities, future studies are needed to provide more specificinformation on the responses of mangroves to accelerated sea levelrises, especially focusing on the increase in inundation depths.
Acknowledgments
The authors would like to thank Yihui Zhang, Jin Wang, HuiChen, Jiemin Guo, Canmou Wang, Xia Yuan and Ronghao Peng fortheir assistances and advice throughout the various stages of thestudy. We thank Abby Lunstrum, Julie Doumbia and Yvonne M.Jeary for improving English language. This research was supportedfinancially, in part, by grants from the Chinese National Science
Foundation (30930017 and 30700092) and the National BasicResearch Program of China (2009CB426306).
References
Abel, N.A.N., Gorddard, R., Harman, B., Leitch, A., Langridge, J., Ryan, A., Heyenga, S.,2011. Sea level rise, coastal development and planned retreat: analyticalframework, governance principles and an Australian case study. Environ. Sci.Policy 14, 279e288.
Ashford, A.E., Allaway, W.G., 1995. There is a continuum of gas space in young plantsof Avicennia marina. Hydrobiologia 295, 5e11.
Blake, G.R., 1965. Methods of Soil Analysis e Physical and Mineralogical PropertiesIncluding Statistics of Measurements and Sampling (Part 1). American Societyof Agronomy, Madison, pp. 374e390.
Blakemore, L., 1987. Extractable iron, aluminium and silicon. In methods forchemical analysis of soils. NZ Soil Bur. Scientific Rep. 80, 71e76.
Blanchard, J., Prado, G., 1995. Natural regeneration of Rhizophora mangle in stripclearcuts in northwest Ecuador. Biotropica 27, 160e167.
Buckley, D.S., Sharik, T.L., Isebrands, J.G., 1998. Regeneration of northern red oak:positive and negative effects of competitor removal. Ecology 79, 65e78.
Cardona-Olarte, P., Twilley, R., Krauss, K., Rivera-Monroy, V., 2006. Responses ofneotropical mangrove seedlings grown in monoculture and mixed cultureunder treatments of hydroperiod and salinity. Hydrobiologia 569, 325e341.
Chen, L., 2005. Studies on the Mechanisms of Mangrove Seedlings in Response toDuration of Tidal Immersion. Doctoral dissertation, Xiamen University, Xiamen.
Chen, L., Wang, W., Lin, P., 2004. Influence of water logging time on the growth ofKandelia candel seedlings. Acta Oceanol. Sin. 23, 149e150.
Chen, L., Wang, W., Lin, P., 2005. Photosynthetic and physiological responses ofKandelia candel L. Druce seedlings to duration of tidal immersion in artificialseawater. Environ. Exp. Bot. 54, 256e266.
Chin, A., Kyne, P.M., Walker, T.I., McAuley, R.B., 2010. An integrated risk assessmentfor climate change: analysing the vulnerability of sharks and rays on Australia’sGreat Barrier Reef. Glob. Change Biol. 16, 1936e1953.
Clarke, P., Allaway, W., 1993. The regeneration niche of the grey mangrove (Avi-cennia marina): effects of salinity, light and sediment factors on establishment,growth and survival in the field. Oecologia 93, 548e556.
Colonnello, G., Medina, E., 1998. Vegetation changes induced by dam constructionin a tropical estuary: the case of the Mánamo river, Orinoco Delta (Venezuela).Plant Ecol. 139, 145e154.
Day, J.W., Christian, R.R., Boesch, D.M., Yanez-Arancibia, A., Morris, J., Twilley, R.R.,Naylor, L., Schaffner, L., Stevenson, C., 2008. Consequences of climate change onthe ecogeomorphology of coastal wetlands. Estuar. Coast. 31, 477e491.
Di Nitto, D., Dahdouh-Guebas, F., Kairo, J.G., Decleir, H., Koedam, N., 2008. Digitalterrain modelling to investigate the effects of sea level rise on mangrovepropagule establishment. Mar. Ecol.-Prog. Ser. 356, 175e188.
Duke, N., 1991. A systematic revision of the mangrove genus Avicennia (Avicennia-ceae) in Australasia. Aust. Syst. Bot. 4, 299e324.
Duke, N.C., Ball, M.C., Ellison, J.C., 1998. Factors influencing biodiversity and distri-butional gradients in mangroves. Glob. Ecol. Biogeogr. Lett. 7, 27e47.
Ellison, A.M., Farnsworth, E.J., 1993. Seedling survivorship, growth, and response todisturbance in Belizean mangal. Am. J. Bot. 80, 1137e1145.
Ellison, A.M., Farnsworth, E.J., 1997. Simulated sea level change alters anatomy,physiology, growth, and reproduction of red mangrove (Rhizophora mangle L.).Oecologia 112, 435e446.
Erlandson, J.M., 2012. As the world warms: rising seas, coastal archaeology, and theerosion of maritime history. J. Coast. Conserv. 16, 137e142.
Fan, H., Li, G., 1997. Effect of sea dike on the quantity, community characteristics andrestoration of mangrove forest along Guangxi coast. Chin. J. Appl. Ecol. 8, 240e244 (in Chinese with English abstract).
Fisher, J.B., Honda, H., 1977. Computer simulation of branching pattern and geom-etry in Terminalia (Combretaceae), a tropical tree. Bot. Gaz. 138, 377e384.
Gilman, E.L., Ellison, J., Duke, N.C., Field, C., 2008. Threats to mangroves from climatechange and adaptation options: a review. Aquat. Bot. 89, 237e250.
He, B., Lai, T., Fan, H., Wang, W., Zheng, H., 2007. Comparison of flooding-tolerancein four mangrove species in a diurnal tidal zone in the Beibu Gulf. Estuar. Coast.Shelf Sci. 74, 254e262.
Hickey, D., Bruce, E., 2010. Examining tidal inundation and salt marsh vegetationdistribution patterns using spatial analysis (Botany Bay, Australia). J. Coast. Res.26, 94e102.
Hoppe-Speer, S.C.L., Adams, J.B., Rajkaran, A., Bailey, D., 2011. The response of thered mangrove Rhizophora mucronata Lam. to salinity and inundation in SouthAfrica. Aquat. Bot. 95, 71e76.
Hovenden, M.J., Curran, M., Cole, M.A., Goulter, P.F.E., Skelton, N.J., Allaway, W.G.,1995. Ventilation and respiration in roots of one-year-old seedlings of greymangrove Avicennia marina (Forsk.) Vierh. Hydrobiologia 295, 23e29.
Hutton, J., 1955. A method of particle size analysis of soils. CSIRO Division of Soils,.Divisional Rep. 11, 55.
IPCC, 2007. Climate Change 2007 Synthesis Report. The Intergovernmental Panel onClimate Change IPCC. Switzerland, Geneva.
Kamali, B., Hashim, R., 2011. Mangrove restoration without planting. Ecol. Eng. 37,387e391.

W. Lu et al. / Acta Oecologica 49 (2013) 83e91 91
Kemp, A.C., Horton, B.P., Donnelly, J.P., Mann, M.E., Vermeer, M., Rahmstorf, S., 2011.Climate related sea-level variations over the past two millennia. Proc. Natl.Acad. Sci. 108, 11017e11022.
Kitaya, Y., Jintana, V., Piriyayotha, S., Jaijing, D., Yabuki, K., Izutani, S., Nishimiya, A.,Iwasaki, M., 2002. Early growth of seven mangrove species planted at differentelevations in a Thai estuary. Trees-Struct. Funct. 16, 150e154.
Krauss, K.W., Lovelock, C.E., McKee, K.L., Lopez-Hoffman, L., Ewe, S.M.L., Sousa, W.P.,2008. Environmental drivers in mangrove establishment and early develop-ment: a review. Aquat. Bot. 89, 105e127.
Kumara, M., Jayatissa, L., Krauss, K., Phillips, D., Huxham, M., 2010. High mangrovedensity enhances surface accretion, surface elevation change, and tree survivalin coastal areas susceptible to sea-level rise. Oecologia 164, 545e553.
Laan, P., Blom, C.W.P.M., 1990. Growth and survival responses of Rumex species toflooded and submerged conditions: the importance of shoot elongation, un-derwater photosynthesis and reserve carbohydrates. J. Exp. Bot. 41, 775e783.
Lin, P., 1999. Mangrove Ecosystem in China. Science Press, Beijing.Lovelock, C.E., Feller, I.C., Ball, M.C., Engelbrecht, B.M.J., Ewe, M.L., 2006. Differences
in plant function in phosphorus- and nitrogen-limited mangrove ecosystems.New Phytol. 172, 514e522.
Martinkova, J., Klimes, L., Klimesova, J., 2011. Multiple regenerative strategies ofshort-lived species: an effect on geographical distribution, preference ofhuman-made habitats and invasive status. Folia Geobot. 46, 181e189.
McKee, K.L., 1995. Seedling recruitment patterns in a Belizean mangrove forest:effects of establishment ability and physico-chemical factors. Oecologia 101,448e460.
McKee, K.L., 1996. Growth and physiological responses of neotropical mangroveseedlings to root zone hypoxia. Tree Physiol. 16, 883e889.
McKee, K.L., Feller, I.C., Popp, M., Wanek, W., 2002. Mangrove isotopic (d15N andd13C) fractionation across a nitrogen VS phosphorus limitation gradient. Ecology83, 1065e1075.
McKee, K.L., Rooth, J.E., Feller, I.C., 2007. Mangrove recruitment after forest distur-bance is facilitated by herbaceous species in the Caribbean. Ecol. Appl. 17, 1678e1693.
Naidoo, G., Naidoo, Y., Achar, P., 2010. Responses of the mangroves Avicennia marinaand Bruguiera gymnorrhiza to oil contamination. Flora 205, 357e362.
Naidoo, G., Rogalla, H., von Willert, D., 1997. Gas exchange responses of a mangrovespecies, Avicennia marina, to waterlogged and drained conditions. Hydro-biologia 352, 39e47.
Panapitukkul, N., Duarte, C.M., Thampanya, U., Kheowvongsri, P., Srichai, N., Geertz-Hansen, O., Terrados, J., Boromthanarath, S., 1998. Mangrove colonization:
mangrove progression over the growing Pak Phanang (SE Thailand) mud flat.Estuar. Coast. Shelf Sci. 47, 51e61.
Platt, T., Gallegos, C.L., Harrison, W.G., 1980. Photoinhibition of photosynthesis innatural assemblages of marine phytoplankton. J. Mar. Res. 38, 687e701.
Rivera-Monroy, V.H., Twilley, R.R., Davis, S.E., Childers, D.L., Simard, M.,Chambers, R., Jaffe, R., Boyer, J.N., Rudnick, D.T., Zhang, K., Castañeda-Moya, E., Ewe, S.M.L., Price, R.M., Coronado-Molina, C., Ross, M., Smith, T.J.,Michot, B., Meselhe, E., Nuttle, W., Troxler, T.G., Noe, G.B., 2011. The role ofthe everglades mangrove ecotone region (EMER) in regulating nutrientcycling and wetland productivity in South Florida. Crit. Rev. Environ. Sci.Technol. 41, 633e669.
Ross, M.S., Meeder, J.F., Sah, J.P., Ruiz, P.L., Telesnicki, G.J., 2000. The southeast salineeverglades revisited: 50 years of coastal vegetation change. J. Veg. Sci. 11, 101e112.
Sanders, C.J., Smoak, J.M., Naidu, A.S., Patchineelamt, S.R., 2008. Recent sedimentaccumulation in a mangrove forest and its relevance to local sea-level rise (IlhaGrande, Brazil). J. Coast. Res. 24, 533e536.
Tomlinson, P.B., 1995. The Botany of Mangroves. Cambridge University Press,Cambridge.
White, A.J., Critchley, C., 1999. Rapid light curves: a new fluorescence methodto assess the state of the photosynthetic apparatus. Photosynth. Res. 59,63e72.
Xiao, Y., Jie, Z., Wang, M., Lin, G., Wang, W., 2009. Leaf and stem anatomical re-sponses to periodical waterlogging in simulated tidal floods in mangrove Avi-cennia marina seedlings. Aquat. Bot. 91, 231e237.
Ye, Y., Gu, Y.T., Gao, H.Y., Lu, C.Y., 2010. Combined effects of simulated tidal sea-levelrise and salinity on seedlings of a mangrove species, Kandelia candel (L.) Druce.Hydrobiologia 641, 287e300.
Ye, Y., Tam, F., Wong, Y., Lu, C., 2003. Growth and physiological responses of twomangrove species (Bruguiera gymnorrhiza and Kandelia candel) to waterlogging.Environ. Exp. Bot. 49, 209e221.
Ye, Y., Tam, N.F.Y., Wong, Y.S., Lu, C.Y., 2004. Does sea level rise influence propaguleestablishment, early growth and physiology of Kandelia candel and Bruguieragymnorrhiza? J. Exp. Mar. Biol. Ecol. 306, 197e215.
Yin, J., Yin, Z., Wang, J., Xu, S.Y., 2012. National assessment of coastal vulnerability tosea-level rise for the Chinese coast. J. Coast. Conserv. 16, 123e133.
Yu, K.F., Kamber, B.S., Lawrence, M.G., Greig, A., Zhao, J.X., 2007. High-precisionanalysis on annual variations of heavy metals, lead isotopes and rare earth el-ements in mangrove tree rings by inductively coupled plasma mass spec-trometry. Nucl. Instr. Meth. B. 255, 399e408.





























