Embed Size (px)
Citation preview

Journal of Dermatological Science 57 (2010) 114–122
Effects of macelignan isolated from Myristica fragrans Houtt. on UVB-inducedmatrix metalloproteinase-9 and cyclooxygenase-2 in HaCaT cells
Anggakusuma a, Yanti a,b, Jae-Kwan Hwang a,*a Department of Biotechnology, Yonsei University, 134 Shinchon-dong, Seodaemun-gu, Seoul 120-749, South Koreab Faculty of Biotechnology, Atma Jaya Catholic University, Jakarta, Indonesia
A R T I C L E I N F O
Article history:
Received 18 June 2009
Received in revised form 15 September 2009
Accepted 3 October 2009
Keywords:
Macelignan
UVB irradiation
MMP-9
COX-2
MAPK pathway
PI3K/Akt pathway
HaCaT cells
A B S T R A C T
Background: UVB irradiation (290–320 nm) is the most damaging component of the UV spectrum and
causes both direct and indirect damage to the basal cell layer of the epidermis; this results in the
activation of a number of signaling pathways involved in pathophysiological processes in the skin, such
as photoaging and inflammation. In photoaging UVB irradiation promotes degradation of the
extracellular matrix (ECM) by matrix metalloproteinases (MMPs) and, in inflammation, UVB irradiation
promotes the expression of inducible cyclooxygenase (COX-2), leading to overproduction of
inflammatory mediators.
Objective: We first investigated the protective effects of macelignan from Myristica fragrans Houtt. on
immortalized human keratinocytes (HaCaT) against UVB damage. We then explored the inhibitory
effects of macelignan on UVB-induced MMP-9 and COX-2 and investigated the molecular mechanism
underlying those effects.
Methods: HaCaT cells were treated with macelignan for the indicated times followed by irradiation with
UVB. Secretion of MMP-9 was measured by gelatin zymography. Expression of COX-2, mitogen-activated
protein kinases (MAPKs), phosphatidylinositol 3-kinase/Akt (PI3K/Akt), c-Fos, c-Jun, and CREB were
assayed by western analysis.
Results: Macelignan at a concentration of 0.1–1 mM increased the viability of HaCaT cells following UVB
irradiation and inhibited MMP-9 secretion and COX-2 expression in a concentration-dependent manner.
An inhibitory effect was also seen in the signal transduction network, where macelignan treatment
reduced the activation of UVB-induced MAPKs, PI3K/Akt, and their downstream transcription factors.
Conclusion: These results suggest that macelignan protects skin keratinocytes from UVB-induced
damage and inhibits MMP-9 and COX-2 expression by attenuating the activation of MAPKs and PI3K/Akt.
� 2009 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights
reserved.
Contents lists available at ScienceDirect
Journal of Dermatological Science
journa l homepage: www.e lsev ier .com/ jds
1. Introduction
As the interface between the organism and the environment,the skin is directly and frequently exposed to a pro-oxidantenvironment, including irradiation by ultraviolet B (UVB) light.UVB radiation can cause damage both directly, by forming covalentbonds between adjacent cytosine bases in DNA and producing aclassic cancerous C-T mutation, and indirectly by inducingoxidative stress [1–5]. UVB irradiation acts primarily on the basalcell layer of the epidermis and, therefore, keratinocytes, whichrepresent the major cell population in the basal layer, are theprimary targets of UVB [6].
Chronic exposure to UVB radiation in sunlight results inextracellular matrix (ECM) degradation by matrix metalloprotei-
* Corresponding author. Tel.: +82 2 2123 5881; fax: +82 2 362 7265.
E-mail address: [email protected] (J.-K. Hwang).
0923-1811/$36.00 � 2009 Japanese Society for Investigative Dermatology. Published b
doi:10.1016/j.jdermsci.2009.10.005
nases (MMPs). MMP-9, a 92 kDa gelatinase and type IV collage-nase, is a UVB-inducible MMP. Under normal physiologicalconditions MMP-9 expression is regulated by endogenousinhibitors, the tissue inhibitors of metalloproteinases (TIMP),particularly TIMP-1 [7]. However, UVB treatment disrupts thenormal balance between TIMP-1 and MMP-9 by inducing over-expression of MMP-9 [8]. In skin MMP-9 is thought to play animportant role in the final degradation of fibrillar collagens afterinitial cleavage by collagenases, including MMP-1, MMP-8, andMMP-13. Furthermore, it has been reported that the gelatinaseactivity of MMP-9 cleaves collagens type I, II, and V in the N-terminal nonhelical telopeptide [9]. It is therefore possible thatMMP-9 play a much broader role in the remodeling of thecollagenous ECM than has been previously thought [10].
Excessive exposure to UVB radiation from sunlight also plays amajor role in the development of acute skin inflammation, leadingto the development of nonmelanoma skin cancer. UVB irradiationactivates the inducible form of cyclooxygenase, cylooxygenase-2
y Elsevier Ireland Ltd. All rights reserved.

Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122 115
(COX-2); COX-2 is a rate-limiting factor in the production ofprostaglandins (PGs), including PGE2. PGE2, in turn, causesvasodilatation and induction of other inflammatory mediators inthe skin [11–14]. Under normal physiological conditions, COX-2 isnot expressed in most tissues; but it is highly inducible by multiplestimuli, including UVB irradiation [15–17]. Thus, because COX-2 isspecifically upregulated in the epidermis as a result of chronicexposure to UVB radiation, the level of COX-2 may serve as an earlymarker of acute skin inflammation [16].
UVB irradiation can trigger the activation of a number ofsignaling pathways leading to increased MMP-9 and COX-2expression. Exposure of cells to UV irradiation elicits a complexset of acute cellular responses known as ‘‘UV responses’’ [18,19].One of these responses is mediated by a membrane-associatedcomponent of the Ras pathway and is followed by subsequentactivation of the mitogen-activated protein kinases (MAPKs) [19–21]. The MAPKs are a family of proline-directed Ser/Thr kinasescomposed of extracellular signal-regulated kinase (ERK), c-JunNH2-terminal kinase (JNK), and p38 kinase. Various studies haveshown that activation of ERK, JNK, and p38 are tightly correlatedwith acute inflammation and development of skin photoaging andinflammation. With regard to MMP-9 expression, MAPK signalingcascades play a significant role in activating the transcriptionfactor AP-1, a complex containing c-Fos and c-Jun, and leading tothe activation of AP-1-dependent gene expression [8]; MMP-9 isone of a class of MMPs for which expression is highly dependent onAP-1 activation [22]. With regard to COX-2, the UVB-induced ERKand p38 pathways, but not the JNK pathway, play a major role inpromoting COX-2 expression [23]. UVB treatment induces theformation of the photoproduct 6-formylindolo[2,3-b]carbazole(FICZ), activating the ERK1/2 pathway, and leading to upregulationof COX-2 expression. Activation of p38 kinase results in thephosphorylation of cyclic AMP response element (CRE) bindingprotein (CREB) at Ser-133 and activates transcription factor-1.Transcription factor-1 then binds to the CRE site at the COX-2 genepromoter and activates transcription. UVB exposure also activatesthe phosphatidylinositol 3-kinase (PI3K)/Akt pathway, anothersignaling pathway which is commonly involved in skin inflamma-tion. PI3K activation by reactive oxygen species (ROS)-inducedepidermal growth factor receptor (EGFR) leads to phosphorylation(and activation) of Akt, which in turn phosphorylates glycogensynthase kinase-3b (GSK-3b). GSK-3b normally phosphorylatesCREB at Ser-129, which is an inhibitory phosphorylation site;therefore, inactivation of GSK-3b by Akt leads to dephosphoryla-tion of CREB at Ser-129, relieving CREB inhibition. This sequence ofevents, together with the phosphorylation (and activation) of CREBat Ser-133 via the p38 pathway, allows CREB to bind to the CRE siteat the COX-2 promoter, recruit its coactivator CREB bindingprotein, and interact with the basal transcriptional machinery[16,24].
Macelignan, a natural compound belonging to a group oflignans isolated from Myristica fragrans Houtt., has been reportedto have various biological activities, including antioxidant, anti-inflammatory, anticariogenic, and antihepatotoxic properties[25–30]. Macelignan has also been shown to function as anantidiabetic and endoplasmic reticulum stress-relief agent [31].In skin, macelignan has been shown to have an inhibitory effect onmelanin overproduction and accumulation in murine melano-cytes suggesting that it may have benefits as a skin-whiteningagent [32]. However, no other studies exploring a role formacelignan in treatment of skin conditions and diseases havebeen reported. Therefore, based on macelignan’s demonstratedactivities as a free-radical-scavenging agent and inhibitor ofprostaglandin, we hypothesized that macelignan could alsoreduce UVB-induced ROS damage, such as aging and inflamma-tion, in skin. In this study we investigated the activity of
macelignan in protecting keratinocytes, the primary cell type inthe basal layer of the epidermis, from UVB damage. We used thehuman immortalized keratinocytes (HaCaT cells) and evaluatedthe effect of macelignan on reducing the level of the primarybiomarkers of UVB-induced skin aging and inflammation, MMP-9and COX-2, and investigated the molecular mechanism under-lying this inhibition.
2. Materials and methods
2.1. Plant material
Dried rhizomes of M. fragrans Houtt. were collected in Jakarta,Indonesia and identified by Dr. Nam-In Baek, Department ofOriental Medicinal Materials and Processing, Kyunghee University,Yongin, Korea. A voucher specimen has been deposited in theDepartment of Biotechnology, Yonsei University, Seoul, Korea.
2.2. Isolation of macelignan
Macelignan was isolated from M. fragrans Houtt. according tothe following procedure. One hundred grams of M. fragrans Houtt.was ground and the powder was extracted in 400 ml of 100%ethanol. The extract (7 g) was further fractionated with ethylacetate. The ethyl acetate fraction (4.2 g) was applied to a silica gelcolumn (60, 70–230 mesh, Merck & Co., Whitehouse Station, NJ,USA) and eluted with n-hexane and ethyl acetate solution (10:1, v/v) in six fractions (fractions I–VI). Fraction III was further separatedwith n-hexane and ethyl acetate solution (20:1, v/v), yieldingfraction III-B (0.52 g). Fraction III-B was applied to an Rp-18column (LiChropep, 25–40 mm m, Merck & Co.) and eluted with80% methanol, yielding compound III-B-2 (0.5 g). Comparison ofspectral data from analysis of compound III-B-2 by severalmethods, including 13C NMR, 1H NMR, 13C DEPT, 1H–1H COSY,1H–13C HSQC, 1H–13C HMBC, and FAB-MS with data in theliterature [28] confirmed that the chemical structure was that ofmacelignan (Fig. 1), i.e., (8R,80S)-7-(3,4-methylenedioxyphenyl)-70-(4-hydroxy-3-methoxyphenyl)-8,80-dimethylbutane. NMRspectra were recorded on a Bruker Avance-500 spectrometer(Rheinstein, Germany) at 600 MHz for 1H and 13C NMR in CDCl3
with TMS as an internal standard. Complete proton and carbonassignments were based on 1D (1H, 13C, 13C DEPT) and 2D (1H–1HCOSY, 1H–13C HSQC, 1H–13C HMBC) NMR experiments. Massspectra (FAB-MS) were collected using a JMS-700 (Jeol Ltd., Tokyo,Japan).
2.3. Cell culture
The immortalized human keratinocyte cell line HaCaT wascultured in Dulbecco’s modified Eagle media (DMEM) supple-mented with 10% fetal bovine serum (FBS) (Gibco, Rockville, MD,USA), 100 U/ml penicillin A, and 100 U/ml streptomycin. Cells werecultured at 37 8C in a humidified atmosphere of 5% CO2/95% air.HaCaT cells were maintained until 40–60% confluence and thenpretreated with indicated concentrations of macelignan in 0.1% (v/v) dimethyl sulfoxide (DMSO) or DMSO alone (control) for 24 h inserum-free culture medium. Medium was then removed and thecells were washed with phosphate-buffered saline (PBS). Fresh PBSwas added and the cells were irradiated with UVB at 30 mJ/cm2.PBS was then removed and the cells were further incubated infresh serum-free culture medium in the presence of the indicatedconcentrations of macelignan. For assay of MAPKs c-Jun, c-Fos,PI3K, Akt, and CREB cells were harvested 6 h after UVB irradiation.Assay of COX-2 expression was carried out on cells harvested 24 hafter UVB irradiation. Cells were harvested 72 h after irradiation forcell viability assays and zymography.

Fig. 1. The chemical structure of macelignan.
Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122116
2.4. Determination of cell viability
HaCaT cell viability was measured using the MTT (3-[4,5-diethylthiazol-2-yl]-2,5-dipheniltetrazolium bromide) colori-metric assay (Sigma–Aldrich, St. Louis, MO, USA) according tothe method of Mosmann [33]. Briefly, HaCaT cells were seeded at2 � 104 cells/well in 200 ml DMEM in a 96-well microtiter plateand cultured for 6 h to allow the cells to attach. To assay the effectof macelignan on HaCaT cell viability the cells were furtherincubated in serum-free medium containing various concentra-
Fig. 2. The effects of macelignan and UVB on HaCaT cell viability. (A) The cytotoxicity of
after the cells were treated for 72 h incubation. (B) The cytotoxicity of various concentrat
post-UV irradiation. (C) The protective effect of macelignan on HaCaT cell viability again
with different doses of macelignan for 24 h and then exposed to UVB (30 mJ/cm2) radia
irradiation, percent cell viability was assessed by the MTT assay. Data are expressed as % o
independent four tests. Asterisks indicate a significant difference compared with the cont
tions of macelignan for 72 h. To assay the protective effect ofmacelignan against UVB damage, the cells were pretreated withmacelignan or vehicle alone for 24 h. Cells were then exposed toUVB at 30 mJ/cm2 as described above and cultured in fresh serum-free DMEM containing macelignan or vehicle alone for another72 h. Finally, the culture medium was replaced with 100 ml 1 mg/ml MTT and incubated for 4 h at 37 8C. The MTT solution wasremoved from the wells by aspiration and the insoluble formazandye was solubilized in 200 ml DMSO; spectrophotometric absor-bance was measured at 550 nm with a tunable microplate reader(Versa Max, Sunnyvale, CA, USA).
2.5. Gelatin zymography
MMP-9 secreted into the culture medium was measured bygelatin zymography. HaCaT cell medium was collected after UVBtreatment followed by an additional 72 h incubation and equalvolumes of each sample were mixed with standard SDS-gel loadingbuffer containing 2% SDS without b-mercaptoethanol and loadedon the gel without prior heating at 100 8C. Samples were separatedby SDS-PAGE through a 10% polyacrylamide gel containing 0.2%gelatin at 85 V for 2 h in a Bio-Rad Mini PROTEAN 3 Cellelectrophoretic apparatus (Bio-Rad Laboratories, Hercules, CA,USA). After electrophoresis the gel was washed twice with 25 ml of2.5% Triton X-100 on a gyratory shaker for 1 h at room temperatureto remove the SDS. The gel was then incubated in 50 ml reactionbuffer (50 mM Tris–HCl, pH 7.5, containing 10 mM CaCl2 and0.15 M NaCl) at 37 8C for 20 h, stained with Coomassie brilliant
various concentrations of macelignan on HaCaT cells was determined by MTT assay
ions of UVB on the cells was also determined after the cells were incubated for 72 h
st UVB-mediated cell viability decrease was also assessed. HaCaT cells were treated
tion. After UVB exposure, fresh media plus samples were added. At 72 h after UVB
f control or non-UVB-treated control and each column represents the mean � S.D. of
rol or non-UVB-treated control group, *p < 0.05 and **p < 0.01.

Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122 117
blue R-250, and destained with methanol–acetic acid in water.MMP-9 was detected at 92 kDa as a clear zone against the darkbackground. Relative band densities were analyzed using MultiGauge Ver. 2.0 software (Fuji Film, Tokyo, Japan).
2.6. Western analysis
HaCaT cells were lysed in RIPA buffer containing proteaseinhibitors (Sigma–Aldrich Co., St. Louis, MO, USA). The concentrationof proteins in the lysates was determined using Bradford proteinassay reagents (Bio-Rad Laboratories Inc., Hercules, CA, USA)and equal amounts (60–100 mg) of protein in each sample wereresolved by SDS-PAGE through a 10% gel and transferred tonitrocellulose membranes (Whatman Protran, Whatman GmBH,
Fig. 3. The effects of macelignan on MMP-9 secretion and COX-2 expression in
HaCaT cells. HaCaT cells were treated with different doses of macelignan for 24 h
and then the cells were exposed to UVB (30 mJ/cm2) radiation. After UVB exposure,
fresh media plus samples were added. (A) For MMP-9 assay, at 72 h after UVB
irradiation, equal volume of the media were collected and used in zymography
assay. (B) For COX-2 assay, at 24 h after UVB, total protein was extracted from the
cells and the expression was determined by western blotting. a-Tubulin was used
as an internal control. Relative expression levels of COX-2 expressed as ratios
against a-tubulin. Data are expressed as % of UVB-treated control and each column
represents the mean � S.D. of independent three tests. Asterisks indicate a significant
difference compared with the UVB-treated control group, *p < 0.05 and **p < 0.01.
Dassel, Germany). The blots were then blocked in 5% skim milk for1 h at room temperature and probed with the primary antibodyagainst each target protein at a concentration of 1:500–1:1000 in 5%skim milk in TBS-T overnight at 4 8C. All primary antibodies werepurchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA)with the exception of the total ERK MAPK, total JNK MAPK, phospho-Akt, phospho-CREB, and total CREB antibodies, which werepurchased from Cell Signaling Technology (Beverly, MA, USA).Bound antibodies were detected by horseradish peroxidase-conjugated secondary antibodies and signals were detected usingan enhanced chemoluminescence (ECL) method (Elpis-Biotech,Taejeon, Korea). Finally, the relative band densities were determinedusing Multi Gauge Ver. 2.0 software (Fuji Film, Tokyo, Japan).
2.7. Statistical analysis
Experiments were performed at least in triplicate throughoutthe study and all data are presented as the mean � standarddeviation (SD) of triplicate values. Statistical significance wasdetermined using Student’s t-tests. The differences between treatedand control groups were also analyzed by analysis of variance(ANOVA) and Duncan’s test (SPSS 12.0). *p < 0.05 and **p < 0.01 wereconsidered statistically significant.
3. Results
3.1. Effect of macelignan and UVB on HaCaT cell viability
The cytotoxic effect of macelignan and UVB on HaCaT cells wasdetermined by MTT assay. Treatment with macelignan in aconcentration range of 1–50 mM for 72 h showed that, below5 mM, macelignan did not reduce cell viability (Fig. 2A). However,UVB treatment, even at the lowest dose of 30 mJ/cm2, reducedHaCaT cell viability to approximately 80% of control values and, asthe dose of UVB increased, cell viability was further reduced(Fig. 2B). This result is in agreement with previous findings [34],which showed that HaCaT cell death begins to be induced by UVBirradiation at 30 mJ/cm2.
3.2. Effect of macelignan on UVB-mediated decrease in
viability of HaCaT cells
The protective effect of macelignan against UVB-mediatedreduction in HaCaT cell viability was further examined todetermine if macelignan could prevent UVB-induced cell death.UVB treatment at 30 mJ/cm2 again reduced cell viability toapproximately 80% cell survival, similar to the previous result(Fig. 2C). Macelignan at a concentration of 0.1–1 mM restore theviability of UVB-treated HaCaT cells, with the greatest cell viabilityclosely approaching 100%, i.e., the viability of non-UVB-treatedcontrol cells.
3.3. Effect of macelignan on UVB-induced MMP-9 secretion
MMP-9 is a UVB-inducible matrix metalloproteinase that playsan important role in photoaging of skin. Exposure of HaCaT cells toUVB irradiation at 30 mJ/cm2 followed by a 72 h incubationresulted in an increase MMP-9 secretion (Fig. 3A). MMP-9 activityin the culture medium increased approximately twofold (forproMMP-9) to fivefold (for MMP-9) after UVB treatment. Exposureof HaCaT cells to 0.1–1 mM macelignan suppressed MMP-9secretion from UVB-treated cells in a concentration-dependentmanner; macelignan had no effect on secretion in non-UVB-treated cells. At the highest macelignan concentration (1 mM),secretion of MMP-9 was reduced to the level seen in non-UVB-treated control cells.
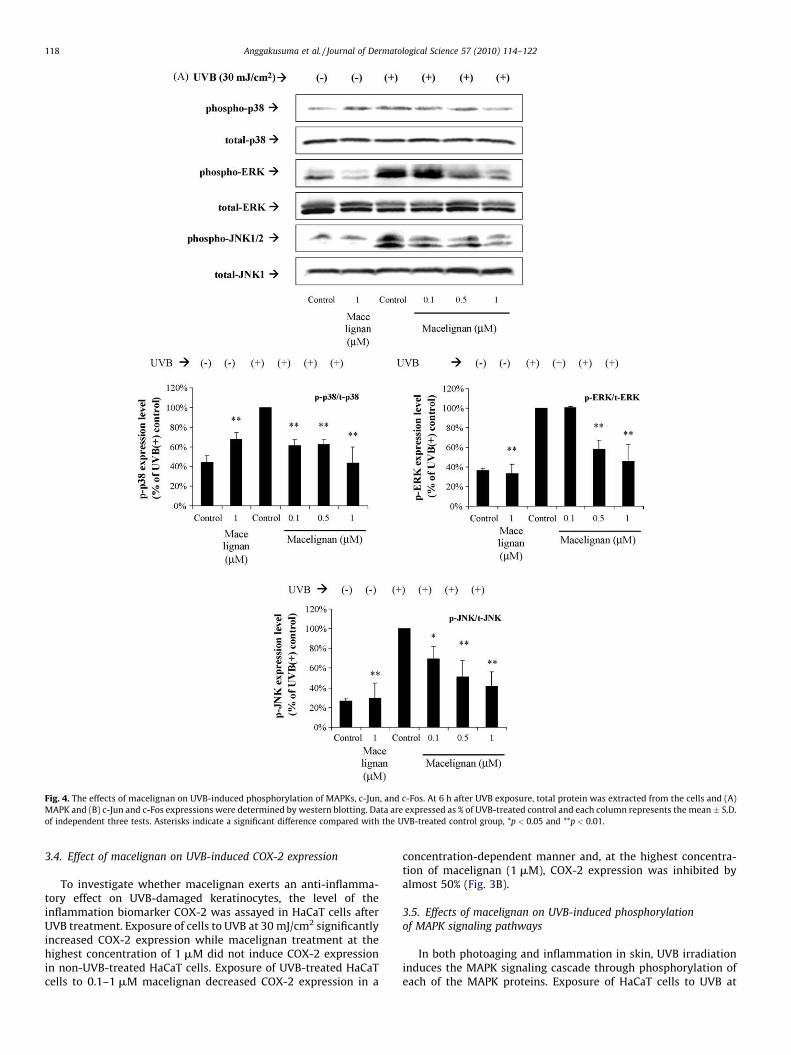
Fig. 4. The effects of macelignan on UVB-induced phosphorylation of MAPKs, c-Jun, and c-Fos. At 6 h after UVB exposure, total protein was extracted from the cells and (A)
MAPK and (B) c-Jun and c-Fos expressions were determined by western blotting. Data are expressed as % of UVB-treated control and each column represents the mean � S.D.
of independent three tests. Asterisks indicate a significant difference compared with the UVB-treated control group, *p < 0.05 and **p < 0.01.
Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122118
3.4. Effect of macelignan on UVB-induced COX-2 expression
To investigate whether macelignan exerts an anti-inflamma-tory effect on UVB-damaged keratinocytes, the level of theinflammation biomarker COX-2 was assayed in HaCaT cells afterUVB treatment. Exposure of cells to UVB at 30 mJ/cm2 significantlyincreased COX-2 expression while macelignan treatment at thehighest concentration of 1 mM did not induce COX-2 expressionin non-UVB-treated HaCaT cells. Exposure of UVB-treated HaCaTcells to 0.1–1 mM macelignan decreased COX-2 expression in a
concentration-dependent manner and, at the highest concentra-tion of macelignan (1 mM), COX-2 expression was inhibited byalmost 50% (Fig. 3B).
3.5. Effects of macelignan on UVB-induced phosphorylation
of MAPK signaling pathways
In both photoaging and inflammation in skin, UVB irradiationinduces the MAPK signaling cascade through phosphorylation ofeach of the MAPK proteins. Exposure of HaCaT cells to UVB at

Fig. 4. (Continued ).
Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122 119
30 mJ/cm2 increased phosphorylation of the MAPK proteins within6 h of irradiation over non-UVB-treated controls (Fig. 4A). Treat-ment with macelignan alone at the highest concentration (1 mM)had no significant effect on the phosphorylation of the MAPKs innon-UVB-treated cells. However, in UVB-treated cells, macelignanat a concentration of 0.1–1 mM attenuated the phosphorylation ofp38, ERK, and JNK in a concentration-dependent manner.
3.6. Effects of macelignan on c-Jun and c-Fos activation in relation to
MMP-9 expression in UVB-treated HaCaT cells
Phosphorylation of MAPKs induces the phosphorylation of c-Jun and the expression of c-Fos, which subsequently form homo- orheterodimers, leading to AP-1 complex formation. Thus, thetranscriptional activity of AP-1 is dependent on the degree ofphosphorylation of c-Jun and the expression level of c-Fos. Ourresults showed that UVB irradiation of HaCaT cells at 30 mJ/cm2
increased the level of c-Fos and phosphorylated c-Jun within 6 hpost-irradiation (Fig. 4B). Treatment of UVB-irradiated cells with0.1–1 mM macelignan inhibited UVB-mediated phosphorylation ofc-Jun and also caused a significant change in UVB-inducedexpression of c-Fos.
3.7. Effects of macelignan on activation of the inflammation-related
PI3K/Akt signaling pathway
The PI3K/Akt pathway is involved in inflammation and isresponsible for COX-2 expression following UVB treatment. ROSformation also plays an important role in the activation of thispathway. Treatment of HaCaT cells with UVB at 30 mJ/cm2
increased PI3K expression as well as Akt phosphorylation.Treatment of cells with macelignan at the highest concentration
(1 mM) had no significant effect on PI3K/Akt activation in non-UVB-treated cells (Fig. 5A). However, treatment of UVB-treatedcells with 0.1–1 mM macelignan inhibited activation of both PI3Kand Akt.
3.8. Effect of macelignan on CREB activation related to COX-2
expression in UVB-treated HaCaT cells
Activation of the PI3K/Akt pathway leads to CREB phosphor-ylation at Ser-133 which, in turn, results in upregulation of COX-2.Since macelignan inhibited PI3K/Akt activation in the previousexperiment, we next evaluated the involvement of CREB in theinhibitory effect of macelignan on UVB-induced COX-2 expression.UVB treatment at 30 mJ/cm2 markedly increased the level of CREBphosphorylation at Ser-133, activating CREB (Fig. 5B). In contrast,macelignan at the highest concentration (1 mM) did not induceCREB phosphorylation. However, treatment of UVB-irradiatedHaCaT cells with macelignan at 0.1–1 mM reduced the phosphor-ylation of CREB at Ser-133 in a concentration-dependent manner.
4. Discussion
The aim of this study was to validate the claim that macelignanacts as antioxidant and anti-inflammatory agent as has been putforth in previous studies. UVB irradiation can induce DNA damagedirectly or indirectly through formation of ROS. Increasing ROSgeneration contributes to several pathological conditions, includ-ing cell death, photoaging, and skin inflammation. Our resultsshowed that the percentage of viable cells decreased in cultures ofHaCaT cells exposed to UVB irradiation compared to control cells.The cell’s antioxidant defenses may be overwhelmed by excessiveROS production induced by the UVB, altering the redox status of

Fig. 5. The effects of macelignan on UVB-induced expression/phosphorylation of PI3K, Akt, and CREB transcription factor. At 6 h after UVB exposure, total protein was
extracted from the cells and the (A) expression of PI3K, phosphorylation of Akt, and (B) phosphorylation of CREB transcription factor at Ser-133 were determined by western
blotting. Data are expressed as % of UVB-treated control and each column represents the mean � S.D. of independent three tests. Asterisks indicate a significant difference
compared with the UVB-treated control group, *p < 0.05 and **p < 0.01.
Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122120

Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122 121
the cell and affecting cell viability. This result is consistent withearlier findings, which show that UVB starts inducing HaCaT celldeath at a dose of 30 mJ/cm2 [34]; therefore, we used 30 mJ/cm2 inthis study because at this dose cell viability remains high, making itpossible to conduct further analyses, such as extraction of proteinand RNA. The protective effect of macelignan on UVB-mediatedHaCaT cell viability decrease was investigated. UVB irradiation isknown to induce apoptosis as a defense against carcinogenesis.However, UVB-induced apoptosis can be suppressed by treatmentwith an antioxidant [34,35]; this suggests that the antioxidantproperties of macelignan may play a critical role in this system. Ourprevious study demonstrated that macelignan exerted antioxidantability against tert-butylhydroperoxide (t-BHP)-induced oxidativedamage in HepG2 cells through reducing cell growth inhibitionand necrosis, ameliorating lipid peroxidation, inhibiting theintracellular ROS formation, and attenuating DNA damage [29].Macelignan protects the cells from UVB-mediated cell death(Fig. 2C) through its antioxidant activity and, therefore, plays asignificant role in ameliorating ROS-related damage.
We also investigated the effect of macelignan on ROS-relatedskin damage, such as photoaging and inflammation. Macelignansignificantly reduced the expression of UVB-inducible MMP-9(Fig. 3A). MMP-9 is a UVB-inducible matrix metalloproteinase thatplays an important role in skin photoaging. The gelatinase activityof MMP-9 plays an important role in UVB-induced skin damage,including skin thickening and wrinkle formation. Recent studiesshow that MMP-9 is produced by HaCaT cells when the cells areexposed to UVB above a certain threshold dose. ROS induced byUVB irradiation play a role in the expression of MMPs through theaction of redox-regulated transcription factors [36–38]. It had beenassumed that macelignan’s antioxidant properties contributed tothis activity because it had been shown in several studies thatantioxidants inhibit the activity of UVB-induced MMP-9 [37–39].To investigate whether macelignan exerts an anti-inflammatoryeffect on UVB-damaged keratinocytes, levels of the inflammationbiomarker COX-2 were measured after exposure of HaCaT cells toUVB. The level of COX-2 is a limiting factor in the production of theinflammatory mediator prostaglandin [40,41]. UVB irradiationalone has been shown to induce COX-2 expression in vitro,including in HaCaT cells where the effect is transmitted throughROS formation [17,42]. As shown in Fig. 3B, COX-2 expression wassignificantly suppressed by macelignan, indicating that mace-lignan also exhibits anti-inflammatory activity in UVB-irradiatedHaCaT cells. As macelignan posseses antioxidant activity byreducing ROS generation and DNA damaging effect caused by t-BHP in HepG2 cells [29], it is conceivable that inhibition of COX-2expression by macelignan may be facilitated through theseparameters. This finding is in agreement with a previous report,which showed that macelignan attenuates COX-2 expression inanother cell type [27].
Additional assays were conducted to investigate the molecularmechanism underlying the inhibitory activity of macelignan onMMP-9 and COX-2 expression. In both skin photoaging andinflammation, UVB induces the MAPK signaling cascade throughphosphorylation of each MAPK protein (Fig. 4A). Accumulatingdata suggest that activation of the MAPKs ERK and p38 are closelyrelated to modulation of levels of both MMP-9 and COX-2 whileactivation of JNK is more closely associated with MMP-9expression [8,23]. With regard to MMP-9 expression, phosphor-ylation of MAPKs induces phosphorylation of c-Jun and expressionof c-Fos (Fig. 4B), which then form homo- or heterodimers, leadingto AP-1 complex formation; AP-1 is a regulator of MMP-9expression. UVB irradiation also can induce the inflammation-related PI3K/Akt pathway, which is responsible for COX-2expression (Fig. 5A). UVB-induced ROS formation also plays animportant role in the activation of this pathway and several studies
have demonstrated the involvement of this pathway in COX-2activity [16]. Activation of PI3K by UVB exposure leads to thephosphorylation of Akt, which plays a critical role in activation ofthe transcription factor CREB by dephosphorylation of Ser-129.This dephosphorylation and phosphorylation at Ser-133 (Fig. 5B)by the p38 pathway enables CREB to induce COX-2 expression.Finally, as can be seen in Figs. 4A, B and 5A, B, we also found thatmacelignan could attenuate the activities of the MAPK and PI3K/Akt pathways and their transcription factors, which are respon-sible for MMP-9 and COX-2 overexpression. Our previous studyalso demonstrated that macelignan exerted anti-inflammatoryeffects through the inhibition of lipopolysaccharide-inducedactivation of nuclear factor-kappa B (NF-kB) by blocking thephosphorylation of MAPKs and the degradation of inhibitory-kappa B (IkB) in microglial cells in vitro [43]. Several evidences hadshown that UVB-induced phosphorylation or activation of theMAPK and PI3K/Akt pathways are inhibited by antioxidants[17,44], suggesting that the antioxidant properties of macelignanmay play a significant role in these events.
In summary, the results of this study show that macelignan canattenuate UVB-induced photoaging and inflammation in skin bymodulating MMP-9 and COX-2 expression through the molecularsignaling pathways that control their expression. The data providestrong evidence for the value of macelignan as an antioxidant andanti-inflammatory agent. Further work is required to determine ifmacelignan has effects on other biomarkers of UVB-induced skininflammation.
Acknowledgements
This work was supported in part by Yonsei BiomoleculeResearch Initiative of the two-step Brain Korea 21 Project.
References
[1] Ley RD, Peak MJ, Lyon LL. Induction of pyrimidine dimers in epidermal DNAof hairless mice by UVB: an action spectrum. J Invest Dermatol 1983;80:188–91.
[2] Peak MJ, Peak JG, Moehring MP, Webb RB. Ultraviolet action spectra for DNAdimer induction, lethality, and mutagenesis in Escherichia coli with emphasison the UVB region. Photochem Photobiol 1984;40:613–20.
[3] Mitchell DL, Jen J, Cleaver JE. Sequence specificity of cyclobutane pyrimidinedimers in DNA treated with solar (ultraviolet B) radiation. Nucleic Acids Res1992;20:225–9.
[4] Amstad PA, Cerutti PA. Ultraviolet-B-light-induced mutagenesis of C-H-rascodons 11 and 12 in human skin fibroblasts. Int J Cancer 1995;63:136–9.
[5] Pfeifer GP, You YH, Besaratinia A. Mutations induced by ultraviolet light. MutatRes 2005;571:19–31.
[6] Fischer TW, Zbytek B, Sayre RM, Apostolov EO, Basnakian AG, Sweatman TW,et al. Melatonin increases survival of HaCaT keratinocytes by suppressingUV-induced apoptosis. J Pineal Res 2006;40:18–26.
[7] Stetler-Stevenson WG. Tissue inhibitors of metalloproteinases in cell signaling:metalloproteinase-independent biological activities. Sci Signal 2008;1:re6.
[8] Rittie L, Fisher GJ. UV-light-induced signal cascades and skin aging. Ageing ResRev 2002;1:705–20.
[9] Okada Y, Naka K, Kawamura K, Matsumoto T, Nakanishi I, Fujimoto N, et al.Localization of matrix metalloproteinase 9 (92 kDa gelatinase/type IV collage-nase = gelatinase B) in osteoclasts: implications for bone resorption. Lab Invest1995;72:311–22.
[10] Kahari VM, Saarialho-Kere U. Matrix metalloproteinases in skin. Exp Dermatol1997;6:199–213.
[11] Ley RD. Photoreactivation in humans. Proc Natl Acad Sci 1993;90:4337.[12] Grewe M, Trefzer U, Ballhorn A, Gyufko K, Henninger H, Krutmann J. Analysis
of the mechanism of ultraviolet (UV) B radiation-induced prostaglandin E2synthesis by human epidermoid carcinoma cells. J Invest Dermatol 1993;101:528–31.
[13] Matsui MS, deLeo VA. Photocarcinogenesis by ultraviolet A. CRC Press Inc;1995. pp. 21-30.
[14] Clydesdale GJ, Dandie GW, Muller HK. Ultraviolet light induced injury: immu-nological and inflammatory effects. Immunol Cell Biol 2001;79:547–68.
[15] Buckman SY, Gresham A, Hale P, Hruza G, Anast J, Masferrer J, et al. COX-2expression is induced by UVB exposure in human skin: implications for thedevelopment of skin cancer. Carcinogenesis 1998;19:723–9.
[16] Rundhaug JE, Fischer SM. Cyclo-oxygenase-2 plays a critical role in UV-induced skin carcinogenesis. Photochem Photobiol 2008;84:322–9.

Anggakusuma et al. / Journal of Dermatological Science 57 (2010) 114–122122
[17] Tsoyi K, Park HB, Kim YM, Chung JI, Shin SC, Lee WS, et al. Anthocyanins fromblack soybean seed coats inhibit UVB-induced inflammatory cylooxygenase-2gene expression and PGE2 production through regulation of the nuclearfactor-kappaB and phosphatidylinositol 3-kinase/Akt pathway. J Agric FoodChem 2008;56:8969–74.
[18] Huang C, Ma WY, Maxiner A, Sun Y, Dong Z. p38 kinase mediates UV-inducedphosphorylation of p53 protein at serine 389. J Biol Chem 1999;274:12229–35.
[19] El-Abaseri TB, Hansen LA. EGFR activation and ultraviolet light-induced skincarcinogenesis. J Biomed Biotechnol 2007;97939.
[20] Davis RJ. The mitogen-activated protein kinase signal transduction pathway. JBiol Chem 1993;268:14553–6.
[21] Karin M, Hunter T. Transcriptional control by protein phosphorylation: signaltransmission from the cell surface to the nucleus. Curr Biol 1995;5:747–57.
[22] Overall CM, Lopez-Otın C. Strategies for MMP inhibition in cancer: innovationsfor the post-trial era. Nat Rev Cancer 2002;2:657–72.
[23] Tsatsanis C, Androulidaki A, Venihaki M, Margioris AN. Signalling networksregulating cyclooxygenase-2. Int J Biochem Cell Biol 2006;38:1654–61.
[24] Bachelor MA, Cooper SJ, Sikorski ET, Bowden GT. Inhibition of p38 mitogen-activated protein kinase and phosphatidylinositol 3-kinase decreases UVB-induced activator protein-1 and cyclooxygenase-2 in a SKH-1 hairless mousemodel. Mol Cancer Res 2005;3:90–9.
[25] Filleur F, Le Bail JC, Duroux JL, Simon A, Chulia AJ. Antiproliferative, anti-aromatase, anti-17beta-HSD and antioxidant activities of lignans isolatedfrom Myristica argentea. Planta Med 2001;67:700–4.
[26] Sadhu SK, Okuyama E, Fujimoto H, Ishibashi M. Separation of Leucas aspera, amedicinal plant of Bangladesh, guided by prostaglandin inhibitory and anti-oxidant activities. Chem Pharm Bull 2003;51:595–8.
[27] Jin DQ, Lim CS, Hwang JK, Ha I, Han JS. Anti-oxidant and anti-inflammatoryactivities of macelignan in murine hippocampal cell line and primary cultureof rat microglial cells. Biochem Biophys Res Commun 2005;331:1264–9.
[28] Chung JY, Choo JH, Lee MH, Hwang JK. Anticariogenic activity of macelignanisolated from Myristica fragrans (nutmeg) against Streptococcus mutans. Phy-tomedicine 2006;13:261–6.
[29] Sohn JH, Han KL, Choo JH, Hwang JK. Macelignan protects HepG2 cells againsttert-butylhydroperoxide-induced oxidative damage. Biofactors 2007;29:1–10.
[30] Cui CA, Jin DQ, Hwang YK, Lee IS, Hwang JK, Ha I, et al. Macelignan attenuatesLPS-induced inflammation and reduces LPS-induced spatial learning impair-ments in rats. Neurosci Lett 2008;448:110–4.
[31] Han KL, Choi JS, Lee JY, Song J, Joe MK, Jung MH, et al. Therapeutic potential ofperoxisome proliferators-activated receptor-alpha/gamma dual agonist withalleviation of endoplasmic reticulum stress for the treatment of diabetes.Diabetes 2008;57:737–45.
[32] Cho Y, Kim KH, Shim JS, Hwang JK. Inhibitory effects of macelignan isolatedfrom Myristica fragrans HOUTT. on melanin biosynthesis. Biol Pharm Bull2008;31:986–9.
[33] Mosmann T. Rapid colorimetric assay for cellular growth and survival: applica-tion to proliferation and cytotoxicity assays. J lmmunol Methods 1983;65:55–63.
[34] Wiswedel I, Keilhoff G, Dorner L, Navarro A, Bockelmann R, Bonnekoh B, et al.UVB irradiation-induced impairment of keratinocytes and adaptive responsesto oxidative stress. Free Radic Res 2007;41:1017–27.
[35] Takahashi H, Hashimoto Y, Aoki N, Kinouchi M, Ishida-Yamamoto A, Iizuka H.Copper, zinc-superoxide dismutase protects from ultraviolet B-induced apop-tosis of SV40-transformed human keratinocytes: the protection is associatedwith the increased levels of antioxidant enzymes. J Dermatol Sci 2000;23:12–21.
[36] Onoue S, Kobayashi T, Takemoto Y, Sasaki I, Shinkai H. Induction of matrixmetalloproteinase-9 secretion from human keratinocytes in culture by ultra-violet B irradiation. J Dermatol Sci 2003;33:105–11.
[37] Moon HI, Chung JH. Meso-dihydroguaiaretic acid from Machilus thunbergiiSIEB et ZUCC, and its effects on the expression of matrix metalloproteinase-2, 9cause by ultraviolet irradiated cultured human keratinocyte cells (HaCaT). BiolPharm Bull 2005;28:2176–9.
[38] Zaid MA, Afaq F, Syed DN, Dreher M, Mukhtar H. Inhibition of UVB-mediatedoxidative stress and markers of photoaging in immortalized HaCaT keratino-cytes by pomegranate polyphenol extract POMx. Photochem Photobiol2007;83:882–8.
[39] Vayalil PK, Mittal A, Hara Y, Elmets CA, Katiyar SK. Green tea polyphenolsprevent ultraviolet light-induced oxidative damage and matrix metallo-proteinases expression in mouse skin. J Invest Dermatol 2004;122:1480–7.
[40] St-Onge M, Flamand N, Biarc J, Picard S, Bouchard L, Dussault AA, et al. Char-acterization of prostaglandin E2 generation through the cyclooxygenase(COX)-2 pathway in human neutrophils. Biochim Biophys Acta 2007;1771:1235–45.
[41] Kim EH, Na HK, Kim DH, Park SA, Kim HN, Song NY, et al. 15-Deoxy-delta12,14-prostaglandin J2 induces COX-2 expression through Akt-driven AP-1 activa-tion in human breast cancer cells: a potential role of ROS. Carcinogenesis2008;29:688–95.
[42] Cho JW, Park K, Kweon GR, Jang BC, Baek WK, Suh MH, et al. Curcumin inhibitsthe expression of COX-2 in UVB-irradiated human keratinocytes (HaCaT) byinhibiting activation of AP-1: p38 MAP kinase and JNK as potential upstreamtargets. Exp Mol Med 2005;37:186–92.
[43] Ma J, Hwang YK, Cho WH, Han SH, Hwang JK, Han JS. Macelignan attenuatesactivations of mitogen-activated protein kinases and nuclear factor kappa B bylipopolysaccharide in microglial cells. Biol Pharm Bull 2009;32:1085–90.
[44] Katiyar SK, Afaq F, Azizuddin K, Mukhtar H. Inhibition of UVB-inducedoxidative stress-mediated phosphorylation of mitogen-activated proteinkinase signaling pathways in cultured human epidermal keratinocytes bygreen tea polyphenol(�)-epigallocatechin-3-gallate. Toxicol Appl Pharmacol2001;176:110–7.








![Chukwuma and Chiamaka iochem Pharmacol (os Angel) · PDF fileChukwuma and Chiamaka iochem Pharmacol ... stimulation of internal antioxidant enzymes [9]. Monodora myristica ... in human](https://img.pdfslide.us/doc/110x75/5aa137fa7f8b9a436d8b51a7/chukwuma-and-chiamaka-iochem-pharmacol-os-angel-and-chiamaka-iochem-pharmacol.jpg)






![Alivrepository.liverpool.ac.uk/3008441/1/polymers-202890... · Web viewLopez-Garcia et al. [26] demonstrated argon plasma treatment improved HaCaT keratinocyte proliferation on collagen](https://img.pdfslide.us/doc/110x75/60e6eab96e238712da3a7f52/web-view-lopez-garcia-et-al-26-demonstrated-argon-plasma-treatment-improved-hacat.jpg)













