Embed Size (px)
Citation preview

310
Effects of Hypoxia-Ischemia and MK-801Treatment on the Binding of a Phencyclidine
Analogue in the Developing Rat BrainFaye S. Silverstein, MD, John W. McDonald III, BS,
Michael Bommarito, BS, and Michael V. Johnston, MD
The phencyclidine analogue [3H](l-[2-thienyl]cyclohexyl)piperidine (3H-TCP) binds to the ionchannel associated with the JV-methyl-D-aspartate receptor channel complex. In vitro autora-diography indicates that the distribution of 3H-TCP binding in brain closely parallels that of[3H]glutamate binding to the iV-methyl-D-aspartate receptor. In nine 7-day-old rats, an acutefocal hypoxic-ischemic insult produced by unilateral carotid artery ligation and subsequentexposure to 8% oxygen acutely reduced 3H-TCP binding ipsilateral to the ligation by 30% in theCA1, by 27% in the CA3, by 26% in the dentate gyms, and by 17% in the striatum compared withvalues from the contralateral hemisphere. In 10 littermates that received 1 mg/kg of theneuroprotective noncompetitive A'-methyl-D-aspartate antagonist MK-801 immediately beforehypoxic exposure, the regional distribution of 3H-TCP binding in hypoxic-ischemic brain wasrelatively preserved and there were no interhemispheric asymmetries in 3H-TCP bindingdensities. In addition, in three unoperated rats decapitated 24 hours after MK-801 treatment,3H-TCP binding was reduced by 15-35%; similar bilateral suppression of 3H-TCP binding wasdetected in MK-801-treated ligates. Our data indicate that 3H-TCP autoradiography can beused to assay the efficacy of neuroprotective agents in this experimental model of perinatalstroke. (Stroke 1990;21:310-315)
Alarge body of experimental evidence indicatesthat overactivation of the iV-methyl-D-
^ aspartate (NMDA)-type glutamate receptorcontributes to the pathogenesis of hypoxic-ischemicneuronal injury.1-2 The NMDA receptor channelcomplex includes a glutamate recognition site, anassociated cation channel, and additional regulatorysites.3 The dissociative anesthetic ketamine, phency-clidine, and the closely related drug MK-801 allselectively antagonize NMDA's actions in a noncom-petitive fashion.4-7 Administration of both competi-tive and noncompetitive antagonists of NMDA canattenuate hypoxia-ischemia-induced neuronal injuryin vivo.8-9
From the Departments of Pediatrics and Neurology (F.S.S.,M.B.) and the Medical Science Training Program (J.W.M.),University of Michigan, Ann Arbor, Michigan, and the Depart-ments of Neurology and Pediatrics and The Kennedy Institute(M.VJ.), Johns Hopkins University, Baltimore, Maryland.
Supported by United States Public Health Service grants NS01171 and 26142 (to F.S.S.), NS 19613 (to M.VJ.), and MedicalScience Training Program grant 5T32 6 MO7863-07 (to J.W.M.).MK-801 was provided by Merck Sharp & Dohme, West Point,Pennsylvania.
Address for correspondence: Faye S. Silverstein, MD, TheUniversity of Michigan, 1120C Neuroscience Laboratory Building,1103 East Huron, Ann Arbor, MI 48104.
Received July 14, 1989; accepted September 19, 1989.
Recent data indicate that these pathophysiologicmechanisms are also relevant in immature brain. In7-day-old rat pups, unilateral carotid artery ligationfollowed by 2-3 hours of exposure to 8% oxygen (thebalance nitrogen) results in ipsilateral forebraininjury10-11; the striatum, hippocampus, and overlyingcortex are damaged. Early after injury, the distribu-tion of [3H]glutamate binding is disrupted; 24 hoursafter the ligation, [3H]glutamate binding is selectivelyreduced in brain regions destined for irreversibleinjury.12 In this experimental model, pretreatmentwith the noncompetitive NMDA antagonist MK-8015-6 reduces the severity of tissue damage13; theexcitatory amino acid antagonist kynurenic acid isalso neuroprotective in neonatal rats.14 A recentstudy in a closely related animal model confirmedthat MK-801 reduces ischemia-induced histologicdamage in immature brain.15
[3H](l-[2-thienyl]-cyclohexyl)piperidine (3H-TCP), a phencyclidine analogue, is a ligand for theion channel associated with the NMDA re-ceptor.316 We used in vitro autoradiography toquantify 3H-TCP binding in rats decapitated 24hours after right carotid artery ligation and hypoxicexposure following pretreatment with 1 mg/kg MK-801 or an equal volume of saline. Based on previous
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from
Silverstein et al Hypoxia-Ischemia Alters 'H-TCP Binding 311
[Cd
H
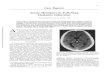
F I G U R E 1. Autoradiogramsdemonstrating distribution of[3H](l-[2-thienyl]cyclohexyl)piperidine (3H-TCP) binding in(left) untreated postnatal day 8rat brain and brain from (right)rat that received 1 mg/kg MK-801 2 hours before decapitation.Twenty-micron horizontal brainsections were incubated with 30nM 3H-TCP. Lighter regionsindicate higher 3H-TCP bindingdensity; brightness in right half ofleft figure is artifactuaL Arrow-heads outline right hippocampus.C, cortex; Cd, caudate; H, hippo-campus. Scale bar, 3.2 mm.
observations of focal reductions in [3H]glutamatebinding, we anticipated that 3H-TCP binding wouldbe selectively reduced ipsilateral to the ligation.The major goals of our study were to determine iftreatment with neuroprotective doses of MK-801attenuated the acute loss of specific 3H-TCP bind-ing and if 3H-TCP binding could be used to quan-titatively assess neuroprotection.
Materials and MethodsAll experiments were done in postnatal day (PND)
7 or 8 Sprague-Dawley rats. We included results fromfour groups: three PND 8 untreated unoperated ratpups, six rat pups that received 1 mg/kg MK-801 2hours (n=3) or 24 hours (n=3) before sacrifice butno ligation, nine saline-pretreated pups that under-went right carotid artery ligation followed by expo-sure to 8% oxygen for 2.5 hours, and 10 MK-801-pretreated pups that underwent right carotidartery ligation followed by exposure to 8% oxygen for2.5 hours.
Pups that underwent surgery were anesthetizedwith ether; the right carotid artery was then exposedand ligated in <5 minutes. After recovery fromanesthesia, the pups were returned to their dams tofeed for 1.5-2 hours. Within 10 minutes beforehypoxic exposure, the pups received a 50-yu.lintraperitoneal injection of saline or 1 mg/kg MK-801 dissolved in saline. The pups were then placed incovered plastic chambers, warmed in a water bath,and exposed to 8% oxygen for 2.5 hours as previouslydescribed.10-11 The pups were returned to their damsand decapitated 24 hours later. The surgical protocolwas approved by the University of Michigan Commit-tee on Care and Use of Animals.
To visualize the regional distribution of 3H-TCPbinding, in vitro autoradiography was done.16 Hori-zontal 20 -ji.m frozen brain sections were incubated
with 30 nM 3H-TCP (55 Ci/mmol, New EnglandNuclear, Boston, Massachusetts) in 50 mM Trisacetate with 1 mM Mg2+, pH 7.4. Nonspecific bindingwas assayed in the presence of 20 /JM TCP.
Adjacent sections from selected brains wereassayed for [3H]glutamate binding to the NMDAsite.17 Twenty-micron sections were incubated with40 nM [3H]glutamate (59 Ci/mmol, Amersham,Arlington Heights, Illinois) in 50 mM Tris acetate,pH 7.2, with 2.5 /xM quisqualic acid. Nonspecificbinding was determined in the presence of 1 mML-glutamate.
Sections and carbon-14 standards (AmericanRadiolabeled Chemicals, Inc., St. Louis, Missouri)were apposed to tritium-sensitive film (LKB-Ultrafilm, Gaithersburg, Maryland) for up to 6 weeksto generate the autoradiograms. Quantitative analy-sis of regional binding was done using a video-basedimage analysis system (MCID, St. Catharines, Cana-da). Optical density was measured bilaterally in threeregions of the hippocampus (CA1, CA3, and thedentate gyms), in the striatum, in the cingulum, andin the fornix; the latter two are typically spared fromhypoxic-ischemic injury (five readings per region persection, four sections per brain).
Specific binding values were derived from polyno-mial regression analysis of optical density values inthe standards calibrated against brain paste withknown amounts of tritium and expressed asmean±SEM femtomoles per milligram protein. Side-to-side differences in hypoxic-ischemic pups werecompared using paired t tests. The effects of MK-801on binding in unoperated controls were assessed byanalysis of variance using a microcomputer-basedstatistical package (SYSTAT, Evanston, Illinois).
ResultsRegionally selective specific 3H-TCP binding was
consistently detected in autoradiograms prepared
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from
312 Stroke Vol 21, No 2, February 1990
NORMALMK801 2hMK801 24h
CA 1 CA 3 DG STRIATUM FORNIX CINGULUM
REGION
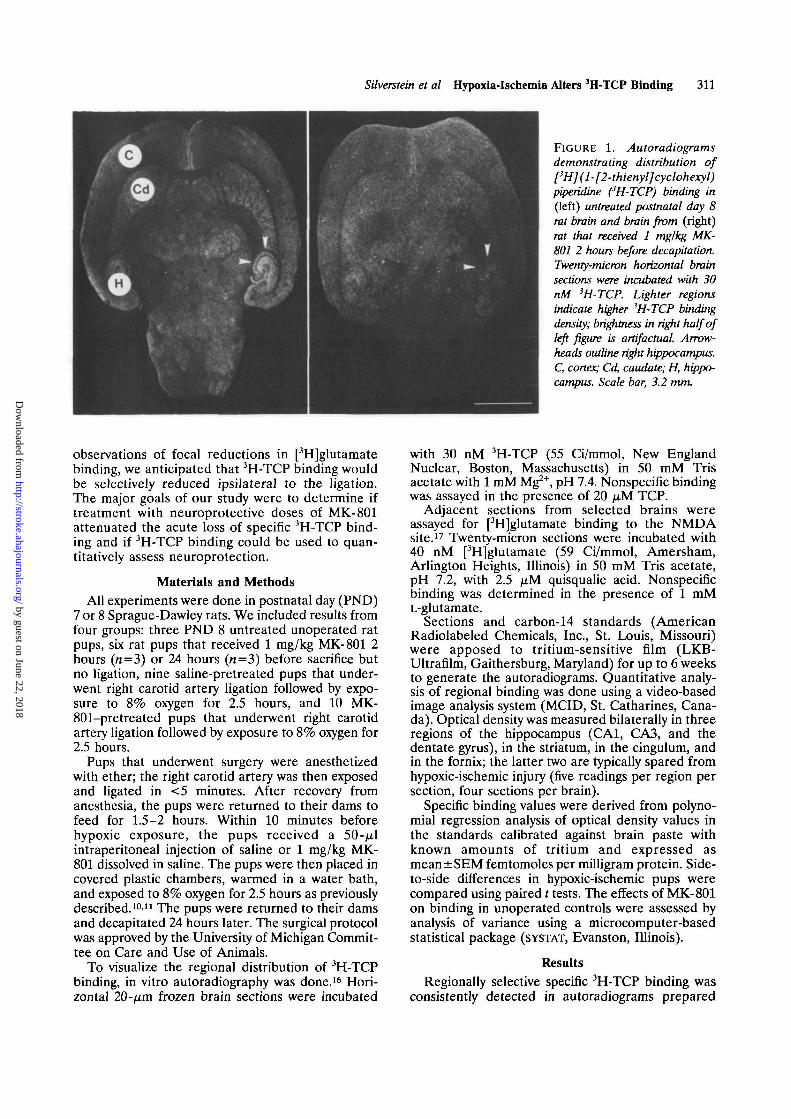
FIGURE 2. Bar graph. [3H](1-[2-thienyl]cyclohexyl)piperidine(3H-TCP) binding in six brainregions in three groups of post-natal day 8 rats. Normal, unop-erated untreated rats (n=3);MK-801 2h, unoperated ratsthat received 1 mg/kg i.p. MK-801 2 hours before sacrifice (n-3); MK-801 24h, unoperatedrats that received 1 mg/kg i.p.MK-801 24 hours before sacri-fice (n=3). Analysis of varianceindicated that MK-801 altered3H-TCP binding in each brainregion, p<0.001.
from untreated PND 8 rat brain (Figure 1, left);binding was highest in the hippocampus. In contrast,virtually no specific binding was present if the pupswere treated with 1 mg/kg MK-801 2 hours beforedecapitation (Figure 1, right; Figure 2).
Figure 2 compares 3H-TCP binding in six brainregions of PND 8 unoperated untreated controls and inunoperated rats treated with 1 mg/kg MK-801 2 or 24hours before decapitation. Binding in all brain regionswas suppressed by 75-82% 2 hours and by 15-35% 24hours after MK-801 administration (/?<0.001).
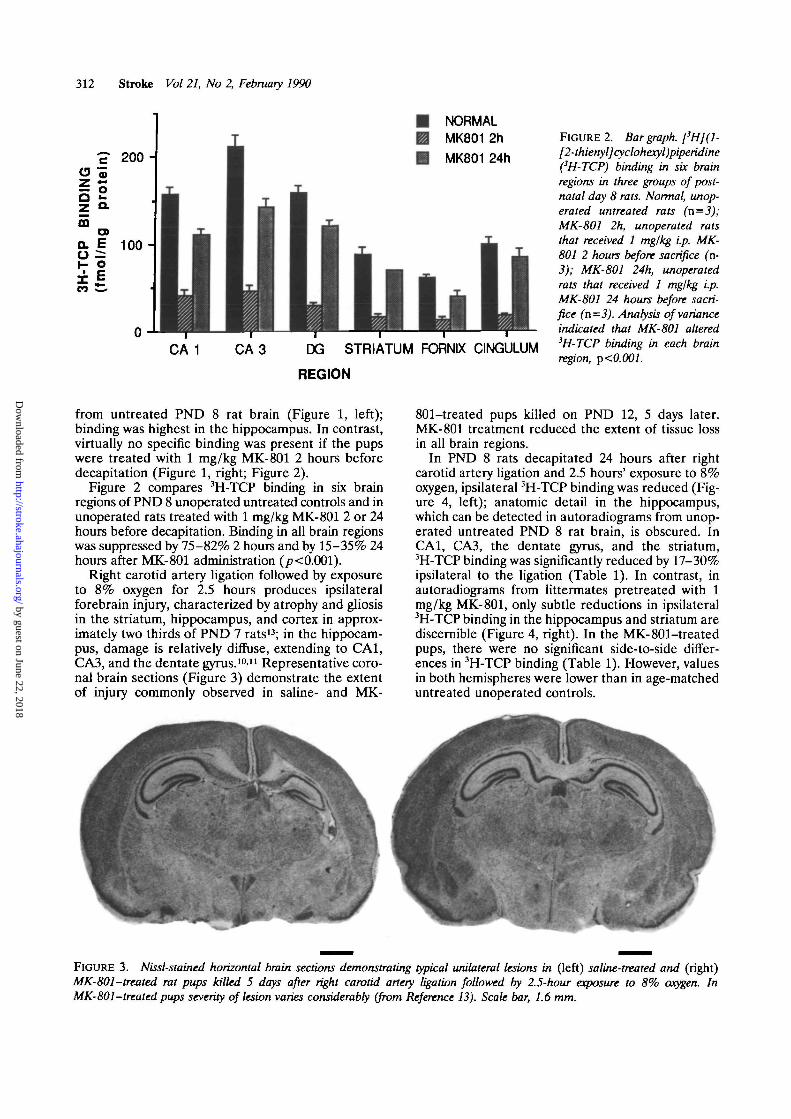
Right carotid artery ligation followed by exposureto 8% oxygen for 2.5 hours produces ipsilateralforebrain injury, characterized by atrophy and gliosisin the striatum, hippocampus, and cortex in approx-imately two thirds of PND 7 rats13; in the hippocam-pus, damage is relatively diffuse, extending to CA1,CA3, and the dentate gyrus.10'11 Representative coro-nal brain sections (Figure 3) demonstrate the extentof injury commonly observed in saline- and MK-
801-treated pups killed on PND 12, 5 days later.MK-801 treatment reduced the extent of tissue lossin all brain regions.
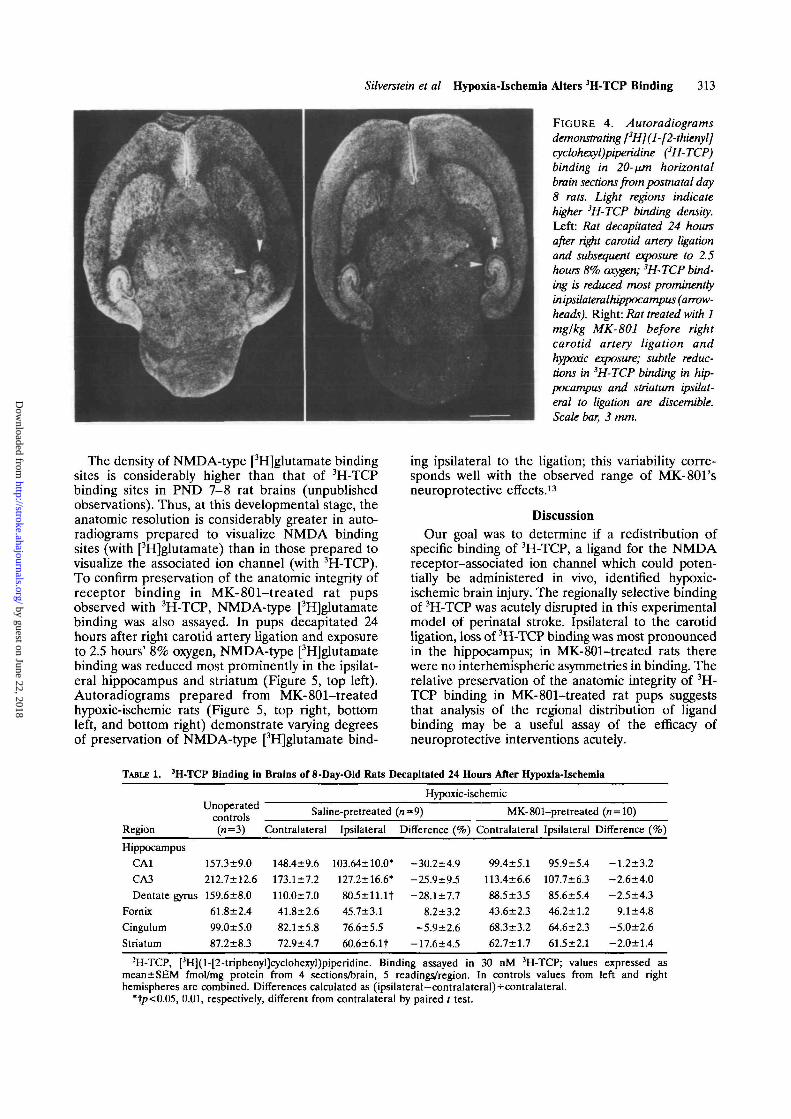
In PND 8 rats decapitated 24 hours after rightcarotid artery ligation and 2.5 hours' exposure to 8%oxygen, ipsilateral 3H-TCP binding was reduced (Fig-ure 4, left); anatomic detail in the hippocampus,which can be detected in autoradiograms from unop-erated untreated PND 8 rat brain, is obscured. InCA1, CA3, the dentate gyrus, and the striatum,3H-TCP binding was significantly reduced by 17-30%ipsilateral to the ligation (Table 1). In contrast, inautoradiograms from littermates pretreated with 1mg/kg MK-801, only subtle reductions in ipsilateral3H-TCP binding in the hippocampus and striatum arediscernible (Figure 4, right). In the MK-801-treatedpups, there were no significant side-to-side differ-ences in 3H-TCP binding (Table 1). However, valuesin both hemispheres were lower than in age-matcheduntreated unoperated controls.
FIGURE 3. Nissl-stained horizontal brain sections demonstrating typical unilateral lesions in (left) saline-treated and (right)MK-801-treated rat pups killed 5 days after right carotid artery ligation followed by 2.5-hour exposure to 8% oxygen. InMK-801-treated pups severity of lesion varies considerably (from Reference 13). Scale bar, 1.6 mm.
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from
Sitverstein et al Hypoxia-Ischemia Alters 3H-TCP Binding 313
FIGURE 4. Autoradiograms
demonstrating [3H](l-[2-thienyl]cyclohexyl)piperidine (3H-TCP)binding in 20-nm horizontalbrain sections from postnatal day8 rats. Light regions indicatehigher 3H-TCP binding density.Left: Rat decapitated 24 hoursafter right carotid artery ligationand subsequent exposure to 2.5hours 8% oxygen; 3H-TCP bind-ing is reduced most prominentlyinipsilateralhippocampus (arrow-heads). Right: Rat treated with 1mglkg MK-801 before rightcarotid artery ligation andhypoxk. exposure; subtle reduc-tions in 3H-TCP binding in hip-pocampus and striatum ipsilat-eral to ligation are discernible.Scale bar, 3 mm.
The density of NMDA-type [3H]glutamate bindingsites is considerably higher than that of 3H-TCPbinding sites in PND 7-8 rat brains (unpublishedobservations). Thus, at this developmental stage, theanatomic resolution is considerably greater in auto-radiograms prepared to visualize NMDA bindingsites (with [3H]glutamate) than in those prepared tovisualize the associated ion channel (with 3H-TCP).To confirm preservation of the anatomic integrity ofreceptor binding in MK-801-treated rat pupsobserved with 3H-TCP, NMDA-type [3H]glutamatebinding was also assayed. In pups decapitated 24hours after right carotid artery ligation and exposureto 2.5 hours' 8% oxygen, NMDA-type [3H]glutamatebinding was reduced most prominently in the ipsilat-eral hippocampus and striatum (Figure 5, top left).Autoradiograms prepared from MK-801-treatedhypoxic-ischemic rats (Figure 5, top right, bottomleft, and bottom right) demonstrate varying degreesof preservation of NMDA-type [3H]glutamate bind-
ing ipsilateral to the ligation; this variability corre-sponds well with the observed range of MK-801'sneuroprotective effects.13
DiscussionOur goal was to determine if a redistribution of
specific binding of 3H-TCP, a ligand for the NMDAreceptor-associated ion channel which could poten-tially be administered in vivo, identified hypoxic-ischemic brain injury. The regionally selective bindingof 3H-TCP was acutely disrupted in this experimentalmodel of perinatal stroke. Ipsilateral to the carotidligation, loss of 3H-TCP binding was most pronouncedin the hippocampus; in MK-801-treated rats therewere no interhemispheric asymmetries in binding. Therelative preservation of the anatomic integrity of 3H-TCP binding in MK-801-treated rat pups suggeststhat analysis of the regional distribution of ligandbinding may be a useful assay of the efficacy ofneuroprotective interventions acutely.
TABLE 1. 3H-TCP Binding in Brains of 8-Day-Old Rats DecapiUted 24 Hours After Hypoxia-Ischemia
Region
HippocampusCA1
CA3
Dentate gymsFornixCingulumStriatum
Unoperatedcontrols(»=3)
157.3±9.0212.7±12.6159.6±8.061.8±2.499.0±5.087.2±8.3
Saline-pretreated
Contralateral
148.4±9.6173.1±7.2110.0±7.041.8±2.682.1 ±5.872.9+4.7
Ipsilateral
103.64±10.0*127.2±16.6*
8O5±ll.lt45.7±3.176.6±5.5
60.6±6.1t
Hypoxic-ischemic
(n-9)
Difference (%)
-30.2±4.9-25.9±9.5-28.1±7.7
8.2±3.2-5.9±2.6
-17.6+4.5
MK-801-pretreated (rc = 10)
Contralateral
99.4±5.1113.4±6.688.5±3.543.6±2368.3+3.262.7±1.7
1 Ipsilateral
95.9±5.4107.7±6.385.6+5.446.2±1.264.6±2.361.5±2.1
Difference (%)
-1.2±3.2-2.6±4.0-2.5±4.3
9.1±4.8-5.0±2.6-2.0±1.4
[3H](l-[2-triphenyl]cyclohexyl)piperidine. Binding assayed in 30 nM 3H-TCP; values expressed asmean±SEM fmol/mg protein from 4 sections/brain, 5 readings/region. In controls values from left and righthemispheres are combined. Differences calculated as (ipsilateral-contralateral)+contralateral.
*t/?<0.05, 0.01, respectively, different from contralateral by paired t test.
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from

314 Stroke Vol 21, No 2, February 1990
F I G U R E 5. Autoradiograms demonstrating N-methyl-D-aspartate (NMDA)-type [3H]glutamate binding in postnatalday 8 rats that underwent right carotid artery ligation andexposure to 2.5 hours of 8% oxygen the day before. Horizontal20-pm frozen brain sections were incubated with 40 nM[3H]glutamate with 2.5 fiM quisqualic acid. Top left: Insaline-treated rat, [3H]glutamate binding is markedly reducedin hippocampus and to lesser degree in striatum and cortexipsilateral to ligation (right side of image); anatomic details ofhippocampus are obscured. Top right, bottom left, andbottom right: In MK-801-treated rats [3H]glutamate bindingipsilateral to ligation is considerably preserved. Scale bar,3 mm.
The reductions in 3H-TCP binding were similar tothe pattern of disruption in total [3H]glutamate bind-ing previously reported in this model.9 These find-ings, together with the results from the autoradio-grams prepared under conditions favoring[3H]glutamate binding to the NMDA site, indicatethat there is no preferential loss of this receptorsubtype acutely with hypoxic-ischemic injury. Thecorresponding preservation of NMDA binding in theMK-801-treated group suggests that the pattern of3H-TCP binding does indeed reflect neuroprotection.
In these experiments, a single concentration ofligand was used and we could not determine ifreductions in binding reflected loss of binding sites orchange in receptor affinity. Based on examination ofcorresponding histologic sections at this time intervalthat reveal loss of Nissl staining in lesioned regionsand minimal edema, it appears likely that the reduc-tions reflect a loss of binding sites. A decrease in the
number of binding sites could reflect reversible mem-brane dysfunction, suppression of protein synthesis,hypoxia-ischemia-induced receptor downregulation,or irreversible membrane damage. Although wecould not address the issue of one-to-one correspon-dence of reductions in 3H-TCP binding with irrevers-ible neuronal injury, available information about theevolution of neuropathology in this model suggeststhat the focal reductions in binding ipsilateral to theligation reflect, in large part, irreversible damage.
Although pretreatment with the noncompetitiveNMDA antagonist MK-801 preserved both tissueintegrity and the normal regional distribution of3H-TCP binding, the binding density was suppressedbilaterally. Similar reductions in 3H-TCP bindingwere observed in unoperated controls decapitated 2or 24 hours after MK-801 administration. The sup-pression of 3H-TCP binding could reflect persistentreceptor occupation by MK-801 or receptordownregulation5'6'18; our experiments did not addressthis issue directly. Of note, the behavioral effects ofMK-801 in vivo (e.g., sedation) are long-lasting (upto 24 hours). Jarvis et al19 reported decreases in3H-TCP binding in gerbil forebrain 4 days after anepisode of transient ischemia in animals treated withthe competitive NMDA antagonist CGS 19755. Theyinterpreted their data as possibly reflecting ischemia-induced receptor desensitization; however, theirobservation could also reflect a direct effect of theNMDA antagonist on 3H-TCP binding.
Acute and chronic changes in binding of ligandsfor glutamate receptors have been reported in mod-els of ischemic brain injury in mature animals. Leachet al20 found that 1 week after bilateral carotid arteryocclusion in adult gerbils 3H-TCP binding assayed intissue homogenates was significantly decreased. Wes-terberg et al21 reported that after an ischemic insult adecrease in the binding of 3H-AMPA, a ligand for asubset of quisqualate-type glutamate receptors, pre-ceded cell death in CA1. In a subsequent study, thesesame authors22 suggested that postischemic receptorchanges, especially decreased NMDA bindingacutely, did not necessarily predict neuronal necrosis.
It is uncertain to what extent results obtained inperinatal animals can be extrapolated to the adultbrain. The cellular mechanisms that lead to irrevers-ible neuronal injury in the immature brain may differin part from pathophysiologic mechanisms in adultanimals, as may the pharmacologic response to drugssuch as NMDA antagonists. The susceptibility toNMDA-induced neurotoxicity peaks in the early post-natal period.23 As well, there is a transient earlydevelopmental peak in glutamate receptor ex-pression.24 Although overactivation of NMDA recep-tors contributes to irreversible ischemia-induced neu-ronal injury at both developmental stages, somecomponents of the response to injury may, in fact, beenhanced during the perinatal period.
In this experimental model, acute disruption of3H-TCP binding occurs in brain regions susceptibleto hypoxic-ischemic injury.10-12 Despite the phar-
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from

Silverstein et al Hypoxia-Ischemia Alters 3H-TCP Binding 315
macologic interactions that complicated our interpre-tation of these data, it appeared feasible to use thisapproach to assay the efficacy of neuroprotectiveagents. In particular, in vitro autoradiography (andpossibly in vivo imaging) with appropriate ligands mayprovide useful information about the temporal evolu-tion of irreversible injury and regional differences inthe efficacy of neuroprotective agents. Furthermore,analysis of alterations in neurotransmitter ligand bind-ing may delineate subtle functional changes in recep-tor organization that evolve after injury.
References1. Rothman SM, Olney JW: Glutamate and the pathophysiology
of hypoxic-ischemic brain damage. Ann Neurol 1986;19:105-114
2. Jorgensen MD, Diemer NA: Selective neuronal loss aftercerebral ischemia in the rat. Possible role of the transmitterglutamate. Ada Neurol Scand 1982;66:536-546
3. Reynolds IJ, Miller RJ: Multiple sites for the regulation of theA'-methyl-D-aspartate receptor. Mol Pharmacol 1988;33:581-584
4. Anis NA, Berry SC, Burton NR, Lodge D: The dissociativeanesthetics, ketamine and phencyclidine selectively reduceexcitation of central mammalian neurones by A'-methyl-aspartate. Br J Pharmacol 1983;79:565-573
5. Wong EHF, Kemp JA, Priestley T, Knight AR, Woodruff G:The anticonvulsant MK-801 is a potent Af-methyl-D-aspartateantagonist. Proc Nad Acad Sci USA 1986;83:7104-7108
6. Sircar R, Rappaport M, Nichtenhauser R, Zukin SR: Thenovel anticonvulsant MK-801: A potent and specific ligand ofthe brain phencyclidine/sigma receptor. Brain Res 1987;435:235-240
7. Huettner JE, Bean BP: Block of NMDA-activated current bythe anticonvulsant MK-801: Selective binding to open chan-nels. Proc Nad Acad Sci USA 1988;85:1307-1311
8. Simon RP, Swan JH, Griffiths T, Meldrum BS: Blockade ofN-methyl-D-aspartate receptors may protect against ischemicdamage in the brain. Science 1984;226:850-852
9. Gill R, Foster LCA, Woodruff GN: Systemic administration ofMK-801 protects against ischemia induced hippocampal neu-rodegeneration in the gerbil. J Neurosci 1987;7:3343-3349
10. Rice JE, Vannucci RC, Brierley JB: The influence of imma-turity on hypoxic-ischemic brain damage in the rat. Ann Neurol1981;9:131-141
11. Johnston MV: Neurotransmitter alterations in a model ofperinatal hypoxic-ischemic brain injury. Ann Neurol 1983;13:511-518
12. Silverstein FS, Torke L, Barks J, Johnston MV: Hypoxia-ischemia produces focal disruption of glutamate receptors indeveloping brain. Dev Brain Res 1987;34:33-39
13. McDonald JW, Silverstein FS, Johnston MV: MK-801 pro-tects the neonatal brain from hypoxic-ischemic damage. EurJPharmacol 1987; 140:359
14. Andine P, Lehmann A, Ellren K, Wennberg E, Kjellmer I,Nielsen T, Hagberg H: The excitatory amino acid antagonistkynurenic acid administered after hypoxic-ischemia in neona-tal rats offers neuroprotection. Neurosci Lett 1988;90:209-212
15. Hattori H, Morin AM, Schwartz PH, Fujikawa DG, Waster-lain CG: Post-hypoxic treatment with MK-801 reduceshypoxic-ischemic damage in the neonatal rat. Neurology 1989;39:713-718
16. Maragos WF, Penney JB, Young AB: Anatomic correlation ofNMDA and 3H-TCP labeled receptors in rat brain. J Neurosci1988;8:493-501
17. Greenamyre JT, Olson JMM, Penney JB, Young AB: Auto-radiographic characterization of NMDA, quisqualate andkainate-sensitive glutamate binding sites. / Pharmacol ExpTher 1985;233:254-263
18. Johnston MV, Silverstein FS, McDonald JW: MK-801 pre-treatment enhances NMDA mediated brain injury and altersNMDA receptor and PCP receptor binding characteristics inperinatal rats (abstract). Soc Neurosci Abs 1988;299:10
19. Jarvis MF, Murphy DE, Williams M, Gerhardt SC, Boast CA:The novel A'-methyl-D-aspartate (NMDA) antagonist CGS19755 prevents ischemia induced reductions of adenosine Al,NMDA and PCP receptors in gerbil brain. Synapse 1988;2:577-584
20. Leach MJ, Hollox KJ, O'Donnell RA, Miller AA: Hippocam-pal NMDA/phencyclidine receptor binding sites are reducedfollowing forebrain ischemia in the gerbil. Eur J Pharmacol1988;152:189-192
21. Westerberg E, Monaghan DT, Cotman CW, Wieloch T:Excitatory amino acid receptors and ischemic brain damage inthe rat. Neurosci Lett 1987;73:119-124
22. Westerberg E, Monaghan DT, Kalions H, Cotman CW, Wie-loch TW: Dynamic changes of excitatory amino acid receptorsin the rat hippocampus following transient cerebral ischemia.J Neurosci 1989;9:798-805
23. McDonald JW, Silverstein FS, Johnston MV: Neurotoxicity ofAT-methyl-D-aspartate is markedly enhanced in developing ratcentral nervous system. Brain Res 1988;459:200-203
24. Greenamyre JT, Penney JB, Young AB, Hudson C, SilversteinFS, Johnston MV: Evidence for transient perinatal gluta-matergic innervation of globus pallidus. J Neurosci 1987;7:1022-1030
KEY WORDSrats
anoxia • MK-801 • A'-methyl-D-aspartate
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from

F S Silverstein, J W McDonald, 3rd, M Bommarito and M V Johnstonanalogue in the developing rat brain.
Effects of hypoxia-ischemia and MK-801 treatment on the binding of a phencyclidine
Print ISSN: 0039-2499. Online ISSN: 1524-4628 Copyright © 1990 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Stroke doi: 10.1161/01.STR.21.2.310
1990;21:310-315Stroke.
http://stroke.ahajournals.org/content/21/2/310World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://stroke.ahajournals.org//subscriptions/
is online at: Stroke Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer available in the
Permissions in the middle column of the Web page under Services. Further information about this process isOnce the online version of the published article for which permission is being requested is located, click Request
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office.Stroke Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on June 22, 2018http://stroke.ahajournals.org/
Dow
nloaded from