Embed Size (px)
Citation preview

Effects of Dynorphin1 _ 13 on Opiate Binding and Dopamine and GABA Uptake in a Cat Model of Stroke Y. Hosonucm, H. KuRODA, and T. MATSUI1
Opiates are best known for their analgesic properties, but they have many other effects on central nervous system (CNS) functions, including respiration, temperature control, and behavior. We have reported that certain opiates also may produce recovery from cerebral ischemia [1, 2, 4, 9]. Subsequent studies performed by others to confirm this finding in some [4, 9-11], although not all [6, 8], cases indicated that, in both experimental animals and in humans, opiate agonists exacerbate the symptoms of stroke, whereas opiate antagonists may prolong survival and, in some cases, ameliorate neurological deficits. Dynorphin1_ 13 , an endogenous opioid peptide with both agonist and antagonist properties [12, 13], also has been shown to improve survival in animals [3].
Although these findings have great clinical potential, the mechanisms of these actions are unknown. As several studies have indicated that opiates do not alter regional cerebral blood flow or systemic variables, such as blood pressure and heart rate [4], a direct action on the CNS seems a likely basis for the observed effects. In view of accruing evidence that endogenous opioids may act as a neurotransmitter or as modulators of a neurotransmitter [12], it is reasonable to hypothesize that opioid antagonists act on specific nervous pathways critical to survival or recovery from stroke. This conclusion is consistent with the findings in several studies demonstrating altered levels of endogenous opioids and of neurotransmitter uptake associated with stroke. For example, Brandt et al. [5] reported that {3-endorphin was elevated in the cerebrospinal fluid (CSF) of a comatose patient with acute necrotizing encephalomyelopathy; similarly, we found a twofold increase in immunoreactive {3-endorphin-like material (lr-{JE) and leucine-
1 Department of Neurological Surgery, School of Medicine, University of California, San Francisco, California, USA
enkephalin in the CSF of one patient who had had a stroke. Focal elevations oflr-{JE have also been observed in the ischemic hemispheres of gerbils subjected to experimental stroke [9].
With regard to neurotransmitters, Weinberger and Cohen [14] reported differences in the sensitivity of dopamine (DA), gamma-aminobutyric acid (GABA), and glutamate uptake to stroke in gerbils, DA being the most sensitive.
In the present study, we explored these relationships further. We determined levels of both opiate-receptor binding and uptake of DA and GABA in various regions of brain in control, ischemic, and dynorphin 1_ 13-treated ischemic cats.
Materials and Methods
Cat Model of Stroke
Transorbital occlusion of the middle cerebral artery (MCA) in cats was performed. Briefly, adult male cats (3-5 kg) were anesthetized; the trachea was intubated, and each cat was placed in a stereotaxic device. The contents of the right orbit (except for the uptake studies, in which the left orbit was used) were removed, and a drill was used to enlarge the optic foramen. The MCA was accessible through an arachnoid dissection. The segment proximal to the lenticulostriate arteries was coagulated with bipolar forceps and transected with microscissors. The orbit was then filled with dental cement and the wound was closed with sutures.
The cats were fully awake within 1-2 h after the operation. We allowed an additional4 h (for cats in uptake studies) or 6 h (for cats in binding studies) during which to assess their neurological status. If analgesia was necessary postoperatively, we administered 0.25% marcaine as a supraorbital and infraorbital nerve block. In specified experiments, dynorphin 1_ 13 (2 mg/kg) was injected intraperitoneally 1 h before the cat
J. Suzuki (ed.), Advances in Surgery for Cerebral Stroke© Springer Japan 1988

242
was killed. This model of cerebral ischemia produced a
highly consistent extent of infarction in studies of more than 80 cats. There was no significant difference in the size of the infarct produced among all control and treatment groups [3].
Preparation of Brain Tissue Fractions
The cat brain, less cerebellum and brain stem, was removed and divided into right and left halves. Each half was further divided into cortical and subcortical regions. Thereby, four regions were obtained from each cat brain. Each brain region was washed in ice-cold 0.32 M sucrose 25 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES), pH 7.7, then homogenized in a TeflonR glass homogenizer. The homogenate was centrifuged at I 000 gat 4 ac for 10 min in a Sorvall RC2B centrifuge; the pellet was discarded and the supernatant processed to obtain either brain membranes (for binding experiments) or crude synaptosomes (for uptake studies). For brain membranes, the supernatant was centrifuged at 20 000 x g at 4oC for 20 min and the resulting pellet was washed once by resuspension in HEPES-sucrose and was centrifuged at 20 000 x g at 4 oc for 20 min. The final pellet (p2) was resuspended in HEPESsucrose and was stored at - 20°C. For synaptosomes, the supernatant was centrifuged at 27 000 x g at 4oC for 30 min and the pellet was retained. The pellet was then resuspended in Krebs-Ringer phosphate buffer (KRB), pH 7.4, containing 0.05 mM pargyline and 1. 7 mM ascorbic acid at a concentration of 150 mg wet weight of tissue/mi. This preparation was used immediately for uptake studies.
Binding Experiments
In saturation experiments, 15 020 concentrations of [3H] ethylketocyclazocine (EKC) ranging from 0.05 to 20 nM were used. The mixture also contained HEPES buffer (25 mM, pH 7.7) and 1 mg brain membrane protein. The mixtures were incubated in a shaking water bath at 37°C for 30 min and then in ice for 60 min, followed by filtration through glass fiber filters. Each filter was washed twice with 5 ml ice-cold HEPES buffer (5 mM), then placed into a polyethylene counting vial with 9 ml scintillation cocktail (Scintiverse II, Fisher Scientific). Radioactivity was counted in a Beckman LS-lOOC scintillation counter. Specific binding was determined as the
Y. Hosobuchi, et a!.
difference between the total brain membrane protein bound and that bound in the presence of I J.-LM EKC. The binding data were analyzed by assuming two independent binding sites (high and low). All samples were analyzed in triplicate.
Uptake of Dopamine and GABA
Aliquots of crude synaptosomes were preincubated with KRB at 37°C for 10 min, then radioactive neurotransmitter-in 100 J.-Ll was added to start the uptake. For DA uptake, the mixture contained 20 J.-Ll (100 J.-Lg) synaptosomes, 880 f.-ll KRB and 50-600 nM DA. For GABA uptake, the mixture contained 40 J.-Ll (200 J.-Lg synaptosomes) in 760 J.-Ll KRB, 100 f.-lllO mM B-alanine, and 4-64 J.-LM GABA.
After 10 min at 37°C, the mixtures were passed through 0.65 f.-till DA Millipore filters. The filters were rapidly washed twice with 5 ml saline, placed into polyethylene counting vials in 9 ml scintillation cocktail and, 72 h later, were counted in a Beckman LS-1 OOC scintillation counter. Specific uptake was determined as the difference between uptake determined at 37oc and that in samples incubated at ooc throughout.
The KRB contained 118 mM NaCl, 4.7 mM KCl, 32.0 mM sodium phosphate (pH 7.4), 1.8 mM CaC12 , 1.2 mM MgS04 , 5.6 mM glucose, 1.3 mM ethylenedaminetetraacetate (EDT A), 1.7 mM ascorbic acid, and 0.05 mM pargyline. Radiolabeled neurotransmitters were obtained from New England Nuclear (Boston, Massachusetts): [3H] DA (23.8 Ci/mmol) and [3H] GABA (30.8 Ci/mmol). The [3H]DA or [3 H] GABA was mixed with unlabeled DA or GABA. (The unlabeled DA and GABA were obtained from Sigma Chemical Company.)
Results
(3H) EKC Binding
In each experiment, the Scatchard plot of [3H] EKC was curvilinear, suggesting the existence of high-affinity and low-affinity binding sites. Figure I shows representative Scatchard plots for control and stroked cat brains. For subcortex, tissue from two identically treated cats was sometimes pooled in order to obtain enough material for assay. The plots for right and left cortices were virtually identical for control cats,
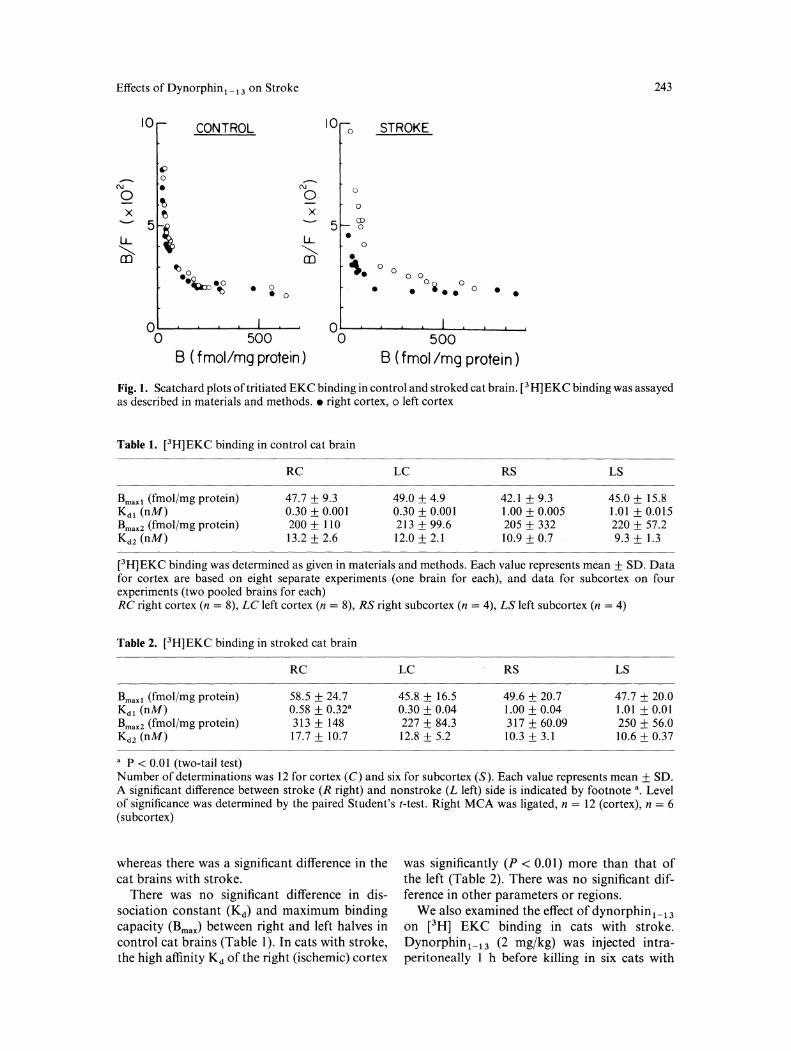
Effects of Dynorphin 1 _ 13 on Stroke 243
10 10 CONTROL 0 STROKE
" - 0 ,....... N • N 0 0 t
0 0
X t> X
5\ 5 CD 0
LL LL • ~ "'-..._
0
CD • ~0 "· 0
0 0 0 •eo eo oi o 4Q:o~ • 0 • • • • 0 • • 0 •
oo~~~~~~~~ 500
00~~~~~~~~~-J 500
B ( fmol/mg protein) B ( fmol /mg protein)
Fig. 1. Scatchard plots of tritiated EKC binding in control and stroked cat brain. [3 H]EKC binding was assayed as described in materials and methods. • right cortex, o left cortex
Table 1. [3H]EKC binding in control cat brain
Bmax 1 (fmol/mg protein) Kd 1 (nM) Bmaxz (fmol/mg protein) Kd2 (nM)
RC
47.7 ± 9.3 0.30 ± 0.001 200 ± 110 13.2 ± 2.6
LC
49.0 ± 4.9 0.30 ± 0.001 213 ± 99.6 12.0 ± 2.1
RS
42.1 ± 9.3 1.00 ± 0.005 205 ± 332 10.9 ± 0.7
LS
45.0 ± 15.8 1.01 ± 0.015 220 ± 57.2 9.3 ± 1.3
[ 3H]EKC binding was determined as given in materials and methods. Each value represents mean± SD. Data for cortex are based on eight separate experiments (one brain for each), and data for subcortex on four experiments (two pooled brains for each) RC right cortex (n = 8), LC left cortex (n = 8), RS right subcortex (n = 4), LS left subcortex (n = 4)
Table 2. [3H]EKC binding in stroked cat brain
Bmax 1 (fmol/mg protein) Kd 1 (nM) Bmaxz (fmoljmg protein) Kd 2 (nM)
" P < 0.0 I (two-tail test)
RC
58.5 ± 24.7 0.58 ± 0.32" 313 ± 148 17.7 ± 10.7
LC
45.8 ± 16.5 0.30 ± 0.04 227 ± 84.3 12.8 ± 5.2
RS
49.6 ± 20.7 1.00 ± 0.04 317 ± 60.09 10.3±3.1
LS
47.7 ± 20.0 1.01 ± 0.01 250 ± 56.0 10.6 ± 0.37
Number of determinations was 12 for cortex (C) and six for subcortex (S). Each value represents mean± SD. A significant difference between stroke (R right) and nonstroke (L left) side is indicated by footnote •. Level of significance was determined by the paired Student's t-test. Right MCA was ligated, n = 12 (cortex), n = 6 (subcortex)
whereas there was a significant difference in the cat brains with stroke.
There was no significant difference in dissociation constant (Kd) and maximum binding capacity (Bmax) between right and left halves in control cat brains (Table 1 ). In cats with stroke, the high affinity Kd of the right (ischemic) cortex
was significantly (P < 0.01) more than that of the left (Table 2). There was no significant difference in other parameters or regions.
We also examined the effect of dynorphin 1_ 13
on [3H] EKC binding in cats with stroke. Dynorphin1_ 13 (2 mgjkg) was injected intraperitoneally 1 h before killing in six cats with

244 Y. Hosobuchi, et al.
Table 3. [3H]EKC binding in stroked cat brain treated with dynorphin 1 _ 13
RC LC RS LS
Bmaxl (fmoljmg protein) 48.0 ± 9.1 50.2 ± 8.5 45.7 ± 16.6 53.2 ± 14.7 Kd 1 (nM) 0.31 ± 0.008 0.31 ± 0.008 0.90 ± 0.08 0.93 ± 0.05 Bmaxz (fmoljmg protein) 2.36 ± 41.5 236 ± 39.1 296 ± 87.7 326 ± 76.8 Kd 2 (nM) 12.0 ± 2.6 12.5 ± 3.6 11.6 ± 0.9 11.4 ± 2.5
Dynorphin 1 _ 13 (2 mgjkg) was injected intraperitoneally 1 h before killing. Number of determinations for cortices (C) and subcortices (S) were six and four, respectively. Each value represents mean± SD., n = 6 (cortex), n = 4 (subcortex)
stroke. This treatment abolished the difference in high affinity Kd between right and left cortex; the binding affinity in right cortex was now increased to the previous level in control (Table 3).
Dopamine and GABA Uptake in Control Cats
In the control cat brain, DA uptake was saturated with 600 nM DA in both cortex and subcortex. In either cortex or subcortex values, there was no significant difference in Km or V max between right and left halves (Table 4). GABA uptake was saturated with 64 11M GABA in both cortex and subcortex. Again, there was no significant difference between right and left halves of either cortex or subcortex (Table 4).
Dopamine and GABA Uptake in Cats with Stroke
All eight cats showed typical hemiplegia 4 h after left MCA occlusion. In the DA uptake study, the Km was significantly reduced (P < 0.001) and the Vmax was increased (P < 0.05) in the right cortex (nonoccluded side) as compared with that in the left cortex (Table 5). The latter values were not significantly different from right or left cortex values in control cats (Table 4). There was no significant difference in Km or V max between right and left subcortex. In the GABA uptake study, there was no significant difference in Km or V max between right and left sides either in cortex or subcortex, or between them and corresponding control values (Table 5).
In cats with stroke treated with dynorphin1_ 13,
the Km of DA uptake in the right cortex was increased so that it no longer differed significantly from the corresponding value of the control cats (Table 4). However, the Km of the left cortex in the dynorphin animals was also raised so that it was significantly higher than the control left cortex value (Table 6). The V max value of the right cortex DA uptake was decreased
under these conditions so that it did not differ significantly from either control values or from the values for the left cortex for the same animal. Finally, the Km ofDA uptake of right subcortex was reduced in dynorphin1_ 13-treated animals, relative to left subcortex or controls. There were no dynorphin-associated changes in GABA uptake parameters.
Discussion
In these studies, we used [3H] EKC to assess opiate binding in control cats and cats with stroke. This opioid agonist binds to at least four major subtypes of opiate receptors found in the brain: mu, delta, kappa, and sigma [7]. We felt this lack of selectivity would be an advantage in our study, as we did not know which, if any, of these subtypes is affected by stroke.
When opiate binding was determined in this manner, the only significant effect of stroke was to increase the high-affinity Kd about twofold in ischemic cortex (Table 2). This effect was reversed by dynorphin 1_ 13 treatment, which otherwise had no effect (Table 3). Because EKC is nonselective under these conditions, it is conceivable that a single opiate receptor subtype is being affected by stroke to a much greater degree than is apparent from our data. As treatment of animals with irreversible alkylating agents selective for particular subtypes does not cause death, it seems unlikely that any magnitude of change in an opiate receptor by itself could be responsible for the debilitating effects of stroke. Nevertheless, the fact that dynorphin1_ 13 was able to improve survival and reverse the change in EKC binding due to stroke suggests that this peptide plays a role in the process. Since the Kd change occurred in the high-affinity site of ischemic cortex only, and not in nonischemic cortex, it appears to be a direct reflection of ischemia, and
Effects of Dynorphin1 _ 13 on Stroke 245
Table 4. Dopamine and GABA uptake in control cat brain
RC LC RS LS
DA uptake Km (nM) 254 ± 39.5 237 ± 36.4 317 ± 64.0 329 ± 48.4 vmax (pmoljmg prot/10 min) 21.4 ± 3.2 21.5 ± 3.7 38.1 ± 7.0 34.6 ± 7.5
GABA uptake Km (J1M) 13.2 ± 3.5 12.9 ± 3.5 12.0 ± 2.7 11.5 ± 3.4 vmax (pmol/mg prot/10 min) 1091 ± 312 993 ± 217 1967 ± 512 2100 ± 446
Dopamine and GABA uptake were determined as given in materials and methods. Number of determinations for cortices (C) and subcortices (S) was nine. Each value represents mean± SD, n = 9
Table 5. Dopamine and GABA uptake in stroked cat brain
DA uptake Km (nM) vmax (pmoljmg prot/10 min)
GABA uptake Km (J1M) vmax (pmol/mg prot/10 min)
a P < 0.001 b p < 0.05
RC
181 ± 28.2• 26.1 ± 4.lb
11.5 ± 2.0 1059 ± 268
LC RS LS
286 ± 64.4 337 ± 107 293 ± 75.3 21.0 ± 3.8 33.4 ± 6.4 33.3 ± 14.3
11.4 ± 2.1 9.9 ± 3.3 9.3 ± 2.9 1043 ± 310 2344 ± 61 1932 ± 671
Number of determinations for cortices (C) and subcortices (S) was eight. Each value represents mean± SD. A significant difference between nonstroke (R right) and stroke (L left) side is indicated by footnotes a and b. Significance was determined by the paired Student's t-test. Left MCA was ligated, n = 8
Table 6. Dopamine and GABA uptake in dynorphin 1 _ 13-treated stroked cat brain
RC LC RS LS
DA uptake Km (nM) 246 ± 62.4 333 ± 70.6 227 ± 74.2 309 ± 116 vmax (pmol/mg prot/10 min) 21.9±4.1 19.6 ± 4.3 37 ± 12.0 32.3 ± 9.3
GABA uptake Km (J1M) 11.4 ± 4.0 14.2 ± 5.8 10.6 ± 3.5 11.5 ± 4.5 vmax (pmol/mg prot/10 min) 1260 ± 269 1106 ± 231 2334± 480 2215 ± 614
Dynorphin1 _ 13 (2 mgjkg) was injected intraperitoneally 1 hour before killing. Number of determinations for cortices (C) and subcortices (S) was eight. Each value represents mean ± SD. A significant difference between nonstroke (R right) and stroke (L left) side is indicated by the asterisk. Significance was determined by the paired Student's t-test. Left MCA was ligated, n = 8
not a compensatory change. We also found stroke-associated changes in
DA uptake. In this case, it was the nonischemic cortex affected, with Km significantly reduced while V max was increased (Table 5). Thus, this could be a compensatory change in which healthy cortex adapts its functions in order to take over for injured tissue. Again, dynorphin1_13 treatment reversed the effects, although the peptide raised the Km in both cortices
so that the ischemic cortex Km was significantly higher than control (Table 6). As with opiate binding, we observed no significant changes in DA uptake in the subcortex, except for a decrease in Km, or right subcortex of stroked cats treated with dynorphin1_13, and no changes in either cortex or subcortex of GABA uptake. In the latter respect, our findings thus agree with those of Weinberger and Cohen [14], who reported that DA uptake was most sensitive to

246 Y. Hosobuchi, et al./Effects of Dynorphin 1_ 13 on Stroke
stroke. The possible relationship to stroke of changes
in DA uptake, like that of opiate binding, is difficult to gauge. Because of the vital role that transmitter uptake plays in· transmitter action, such changes could conceivably have significant effects on brain function. Be that as it may, the selectivity of the effect, with GABA uptake being unaffected, argues against its simply reflecting decreased energy availability. Furthermore, as with opiates, overall changes we observed may mask a larger change in a select group of neurons.
In summary, occlusion of the MCA induced changes in the opiate receptor and DA uptake systems in cats. Treatment with dynorphin1_ 13
reversed these changes. We suggest that the action of dynorphin 1_ 13 must be specific because not all systems were affected. We previously reported that dynorphin prolonged survival in cats subjected to experimental stroke; 60% of treated cats survived for at least 7 days, whereas no untreated cat with stroke survived for more than two days. This rapid mortality for cats in the control condition prohibits controlled, longterm biochemical studies of dynorphin's effects. However, it could be that the biochemical changes we observed after 1 hour are merely initial changes, and prolonged study would show a greater potency of dynorphin in altering binding or uptake. Thus, we may be observing only a fraction of dynorphin's potency in altering binding or uptake.
References
1. Baskin DS, Hosobuchi Y (1981) Naloxone reversal of ischemic neurological deficits in man. Lancet 2:272-275
2. Baskin DS, Hosobuchi Y, Greve! JC (1986) Treatment of experimental stroke with opiate antagonists: Effects on neurological function, infarct size, and survival. J Neurosurg 64: 99-103
3. Baskin DS, Hosobuchi Y, Loh HH, Lee NM (1984) Dynorphin (1-13) improves survival in cats with focal cerebralischemia. Nature 312:551-552
4. Baskin DS, Kieck CF, Hosobuchi Y (1984) Naloxone reversal and morphine exacerbation of neurologic deficits secondary to focal cerebral ischemia in baboons. Brain Res 290: 289-296
5. Brandt NJ, Terenius L, Jacobsen BB, Klinken L, Nordius A, Brandt S, Blegvad K, Yssing M (1980) Hyperendorphin syndrome in a child with necrotizing encephalomyelopathy. N Eng! J Med 303:914-916
6. Cutler JR, Bredesen DE, Edwards R, Simon RP (1983) Failure of naloxone to reverse vascular neurological deficits. Neurology 33: 1517-1518
7. Garzon J, Sanchez-Blazquez P, Lee NM (1984) [ 3H]-ethylketocyclazocine binding to mouse brain membranes: Evidence for a kappa opioid receptor type. J Pharmacol Exp Ther 231:33-37
8. Holoday JN, D'Amato RJ (1982) Naloxone or the TRH fail to improve neurologic deficits in gerbil models of stroke. Life Sci 31 : 385-392
9. Hosobuchi Y, Baskin DS, Woo SK (1982) Reversal of induced ischemic neurologic deficit in gerbils by the opiate antagonist naloxone. Science 215:69-71
10. Iselin HA, Weiss P (1981) Naloxone reversal of ischemic neurologic deficit. Lancet 2: 642-643
11. Jovaily J, Davis JB (1982) Naloxone partially reverses neurologic deficits in some but not all stroke patients. Neurology 32: A 194
12. Lee NM, Smith AP (1984) Possible regulatory function of dynorphin and its clinical implications. Trends Pharmacol Sci 5: 108-110
13. Talunay FC, Jen MF, Chang JK, Loh HH, Lee NM (1981) Possible regulatory role of dynorphin on morphine and P-endorphin-induced analgesia. J Pharmacol Exp Ther 219: 296-298
14. Weinberger J, Cohen G (1982) The differential effect of ischemia on the active uptake of dopamine and aminobutyric acid, and glutamate by brain synaptosomes. J Neurochem 38: 963-968





























