Embed Size (px)
Citation preview

Comp. Biochem. Physiol. Vol. 71B, pp. 145 to 148, 1982 0305-0491/82/010145-04503.00/0 Printed in Great Britain. All rights reserved Copyright © 1982 Pergamon Press Ltd
EFFECTS OF CHROMATIN PROTEIN FRACTIONS ON TRANSCRIPTIONAL ACTIVITY OF CHICKEN
THROMBOCYTE AND ERYTHROCYTE CHROMATIN
W. M. KRAJEWSKA and L K L Y S Z E J K O - S T E F A N O W I C Z
Department of Biochemistry, Institute of Biochemistry and Biophysics, University of L6d2, Banacha 12/16, 90-237 L6d2, Poland
(Received 26May 1981)
Abstract--1. Chromatin proteins of chicken thrombocytes and erythrocytes were separated into three fractions by successive extraction with 5 M urea containing various salt concentrations and pH values. Molecular composition of protein fractions was determined by SDS-polyacrylamide gel electrophoresis.
2. The efficiences of the chromatin residues after sequential protein extractions as well as those of reconstituted DNA-protein fraction complexes, in serving as a template for the in vitro RNA synthesis were measured in order to identify the effect of each fraction.
3. The different involvement of chromatin protein fractions on template properties of thrombocyte and erythrocyte chromatin was stated.
INTRODUCTION
Chromatin transcription in vivo is highly regulated in eukaryotes. There is a lot of evidence that chromatin proteins are involved in the differential regulation of gene expression through their structure and compo- sition. Histones have been suggested to be nonspecific repressors of transcription of chromatin D N A as well as structural components of nucleosomes (Olins & Olins, 1974; Van Holde et al., 1974). Non-histone pro- teins contain fractions that activate (Kostraba & Wang, 1972; James et al., 1977; Legravered & Glazer, 1980) and inhibit RNA synthesis in vitro (Kostraba & Wang, 1975; Hirasawa et al., 1978).
Thrombocytes and erythrocytes seem to offer par- ticular advantages for the study of the possible mech- anisms controlling gene expression. Functional specialization of thrombocytes and erythrocytes regarded as genealogically closely related cells is probably conditioned by the specific factors regulat- ing transcription. Our previous paper on the charac- terization of the chromatin, nuclear phosphoproteins, histones and chromatin-bound protease activity from chicken thrombocytes and erythrocytes seems to con- firm the concept of a close relationship of the cell studied (Krajewska & Ktyszejko-Stefanowicz, 1980). In this paper the involvement of chromatin protein in template activity of chicken thrombocyte and erythro- cyte chromatin was compared.
MATERIALS AND METHODS
Thrombocytes and erythrocytes were separated from chicken blood. Isolation of cells, cell nuclei and chromatin was carried out as described previously (Krajewska & Klyszej ko-Stefanowicz, 1980).
Fractionation of chromatin proteins Chromatin proteins were fractionated into non-histone
proteins soluble in urea at low ionic strength (UP fraction), histones (HP fraction) and DNA-associated non-histone chromatin proteins (NP fraction) according to the method
C.B.P. 71/In J
of Chiu et al. (1975). Briefly, chromatin was gently hom- ogenized in 5 M urea--50mM sodium phosphate buffer (pH 7.6)--0.1 mM PMSF. After adjusting the DNA con- centration to approx. 4 0 D U/ml at 260 nm and stirring for 1 hr, the mixture was centrifuged at 17,000 O for 30 rain. The supernatant containing the bulk of chromatin non- histone proteins was designated as UP fraction. Histones were dissociated from the remaining pellet by 5M urea--2.5M NaCI--10mM sodium phosphate buffer (pH 6)--0.1 mM PMSF (final concentration of chromatin DNA was 2 0 D U/ml at 260 nm). After centrifugation at 110,000O for 36 hr DNA and associated non-histone pro- teins were pelleted (NP fraction) while histones (HP frac- tion) remained in the supernatant.
The supernatants containing UP and HP fractions were concentrated by ultrafiltration (Amicon PM-10 mem- brane).
Isolation of DNA
DNA was isolated from chicken liver by a modification of the method of Marmur (1961).
Polyacrylamide gel electrophoresis Samples were dialyzed against 0.1~ SDS--0.1~o 2-mer-
captoethanol--8 M urea--10 mM sodium phosphate buffer (pH 7). The electrophoretic separation of proteins was per- formed by the method of Weber & Osborn (1969) with the addition of 3~ stacking gel (Briggs et al., 1976). The gels were stained with Coomassie Brilliant Blue R in isopropyl alcohol--acetic acid solution and scanned at 560 nm using an ERJ-65 Carl Zeiss densitograph. Approximate molecu- lar weights were determined against the relative migrations of ~-phosphorylase (94,000), bovine serum albumin (67,000), catalase monomer (60,000), ovalbumin (43,000), deoxyribonuclease I (31,000), chymotrypsinogen (25,000), myoglobin (17,800) and cytochrome c (12,000).
Assay of in vitro R N A synthesis The reaction mixture (0.25ml final vol) contained
40mM Tris-HCl buffer (pH 7.9), 120mM KC1, 0.1 mM EDTA, 1 mM MnC12, 4 mM MgC12, i mM dithiothreitol, 0.08 mM each ATP, GTP and CTP, 0.05 mM aH-UTP (2/tCi), 4.5/zg DNA or chromatin equivalent to 4.5,ug DNA and 1 U of E. coli RNA polymerase (specific activity
145
146 W.M. KRAJEWSKA and L. KLYSZEJKO-STEFANOWICZ
A >100 H I H5
54K I>100 f/ 73K I I i 16K
K
H3+H2B+ H2A+ H4
IOK ~ Total
J I H ~ UP
~ 6 5 K 17K
~ 1 < aP ® - ®
>lOOK 190K
HI+H5
~K
H3+H2B+H2A+H4
Total
K
H3+H2B 2A ,H4
® . ®
U P
H P
NP
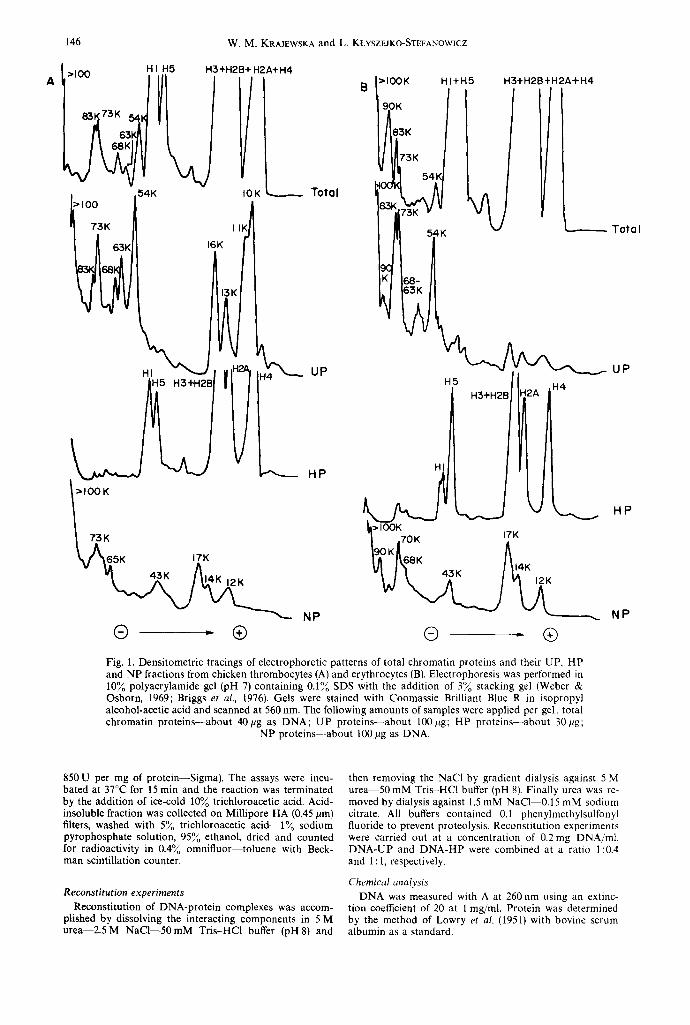
Fig. 1. Densitometric tracings of electrophoretic patterns of total chromatin proteins and their UP, HP and NP fractions from chicken thrombocytes (A) and erythrocytes (B). Electrophoresis was performed in 10~o polyacrylamide gel (pH 7) containing 0.1~ SDS with the addition of 3~o stacking gel (Weber & Osborn, 1969; Briggs et al., 1976). Gels were stained with Coomassie Brilliant Blue R in isopropyl alcohol-acetic acid and scanned at 560 rim. The following amounts of samples were applied per gel: total chromatin proteins--about 40pg as DNA; UP proteins--about 100#g; HP proteins--about 30F~g;
NP proteins--about 100 pg as DNA.
850 U per mg of protein--Sigma). The assays were incu- bated at 37°C for 15 rain and the reaction was terminated by the addition of ice-cold 10~o trichloroacetic acid. Acid- insoluble fraction was collected on Millipore HA (0.45 #m) filters, washed with 5~o trichloroacetic acid--l~o sodium pyrophosphate solution, 95~ ethanol, dried and counted for radioactivity in 0.4~o omnifluor toluene with Beck- man scintillation counter.
Reconstitution experiments
Reconstitution of DNA-protein complexes was accom- plished by dissolving the interacting components in 5 M urea--2.5 M NaCI--50mM Tris-HC1 buffer (pH 8) and
then removing the NaC1 by gradient dialysis against 5 M u r e a 5 0 mM Tris-HC1 buffer (pH 8). Finally urea was re- moved by dialysis against 1.5 mM NaCI 0.15 mM sodium citrate. All buffers contained 0.1 phenylmethylsulfonyl fluoride to prevent proteolysis. Reconstitution experiments were carried out at a concentration of 0.2 mg DNA/ml. DNA-UP and DNA-HP were combined at a ratio 1:0.4 and 1 : 1, respectively.
Chemical analysis DNA was measured with A at 260 nm using an extinc-
tion coefficient of 20 at 1 mg/ml. Protein was determined by the method of Lowry et al. (1951) with bovine serum albumin as a standard.

Chromatin transcription in vitro 147
RESULTS AND DISCUSSION
Characterization of chromatin protein fractions
Total chromatin proteins were separated into three main groups of proteins designated as UP, HP and NP fractions by Chiu et al. (1975) method. The frac- tionation scheme was based on the solubility proper- ties of chromatin proteins in 5 M urea at different pH values and salt concentrations. The UP (non-histone chromatin proteins soluble in urea at low ionic strength), HP (histones) and NP (DNA-associated non-histone chromatin proteins) fractions comprising about 33-41%, 58-66% and 1-1.57oo of total chromatin proteins, respectively, were characterized by SDS- polyacrylamide gel electrophoresis (Fig. 1).
The UP proteins isolated from thrombocyte chromatin revealed a remarkable similarity in the electrophoretic patterns to that of UP proteins from erythrocytes. A number of common bands was ob- served. However, the relative content of the particular components of the UP fraction was very different in cells studied. In UP proteins of thrombocytes there were higher relative amounts of 10,000, 11,000, 13,000, 16,000, 54,000 and 63,000 daltons fractions and lower content of components with mol. wt 73,000 and 83,000. Only one additional fraction of about 90,000 daltons was found in UP proteins of erythrocytes when compared with thrombocytes by using one- dimensional SDS-polyacrylamide gel electrophoresis.
The second extraction at high ionic strength (2.5 M NaC1) and relatively low pH (6.0) removed essentially all histones-HP fraction. The different content of H1 and H5 histones in both cells can be observed.
Non-histone chromatin proteins that remained bound to DNA in the conditions of fractionation method--NP fraction--separated into several com- ponents with mol. wt from 12,000 to above 100,000. The striking differences concerning high mol. wt com- ponents (65,000-100,000) can be noticed. The 90,000 daltons fraction absent in thrombocytes but present in erythrocytes at a high relative amounts seems to be especially interesting. This very fraction was found to be immunologically cell and species specific for chicken erythroid chromatin (Krajewska et al., 1979; 1980).
Effects of chromatin protein fractions on R N A synthesis
Two independent methods were used to evaluate
the role of chromatin proteins in template activity of chicken thrombocyte and erythrocyte chromatins. First, the efficiences of the chromatin in serving as a template for the in vitro RNA synthesis were deter- mined after sequential removal of protein fractions. Each extraction was repeated twice to insure total removal of particular protein fractions from the remaining complex. The extractions effectiveness was checked by polyacrylamide gel electrophoresis of chromatin residue (data not shown). Secondly, protein fractions except NP which remained complexed with DNA were reconstituted with DNA and such com- plexes measured for transcriptional activities. Our experiments demonstrated that studied chromatin protein fractions were differently involved in tran- scriptional activity of chromatin from thrombocytes and erythrocytes. The initial removal of UP proteins approximately doubled the templating efficiency of the chromatin from thrombocytes whereas had almost no effect on RNA synthesis in vitro in erythrocytes (Table 1). The sequential extraction of HP fraction depressed the chromatin DNA, bringing its templat- ing efficiency to about 57~o of the control DNA in thrombocytes while to about 86% in erythrocytes. Simultaneously such results indicated that NP frac- tion remained in the analyzed complex restricted the templating activity of chromatin DNA to a higher degree in thrombocytes than in erythrocytes. These observations were furthermore confirmed by reconsti- tution experiments (Table 2). In erythrocytes HP pro- teins had the highest capability to repress template activity of DNA. On the contrary in thrombocytes similar level of inhibition was observed by NP pro- teins. NP fraction from chicken reticulocyte was found to be essential for the in vitro transcription of globin mRNA by chromatin preparations reconsti- tuted from DNA and chromatin protein components (Chiu et al., 1975).
The results presented suggest that gene expression in chicken thrombocytes and erythrocytes might be highly regulated at the transcriptional level. It was stated by Lasky & Tobin (1979) that transcription in avian erythroid cells is more selective than in other cell types examined. The absence of the vast majority of liver mRNA sequences in reticulocyte hnRNA resulted probably from the transcriptional control.
Since it is now evident that an eukaryotic template transcribed with bacterial polymerase cannot yield
Table 1. Templating activity of chicken thrombocyte and erythrocyte chromatin resi- due after sequential removal of proteins
Thrombocyte Erythrocyte
Template cpm ~ free DNA cpm ~o free DNA
Chromatin 1080 4.0 567 2.1 Chromatin devoid 3161 11.7 973 3.6 of UP proteins Chromatin devoid of 15,400 57.0 23,316 86.3 UP and HP proteins DNA 27,018 100 27,018 100
Chromatin protein fractions were selectively extracted from chromatin with 5 M urea containing various salt concentrations and pH values as described in Materials and Methods. Chromatin residues (4.5 #g as DNA) were assayed for transcriptional activity in the presence of 1 U of E. coil polymerase. Results are expressed as percent of free DNA activity.

148 W.M. KRAJEWSKA and L. KLYSZEJKO-STEFANOWICZ
Table 2. Effects of various chromatin protein fractions on RNA synthesis in vitro
Thrombocyte Erythrocyte
Template cpm % free DNA cpm ~% free DNA
DNA 23,011 100 23,011 100 DNA-UP proteins 18,800 81.7 23,862 103.7 DNA-HP proteins 17,166 74.6 13,369 58.1 DNA-NP proteins 13,438 58.4 20,917 90.9
DNA-UP and DNA-HP protein complexes were reconstituted by the gradient dialy- sis as described in Materials and Methods. DNA-NP protein complex was pelleted after extraction of UP and HP fractions from chromatin. Complexes containing equal amounts of DNA (4.5/~g) were assayed for transcriptional activity in the presence of 1 U of E. coil polymerase. Results are expressed as percent of free DNA activity.
relevant information on transcript ional regulation, interest will certainly shift to more homologous assays. However, it was indicated that specific syn- thesis of globin RNA sequence from chromat in can be performed by nonhomologous prokaryotic RNA polymerase as well (Reff & Davidson, 1979).
Acknowledyement This work was supported by the Polish Academy of Sciences within the project 09.7.1.
REFERENCES
BRIGGS R. C., ROTHSTE1N H. & WAINWRIGHT N. (1976) Cell cycle variations in chromosomal protein of the lens. Expl. Cell Res. 99, 95 105.
CHIU J.-F., WANG S., FUJITAN1 H. & HNILICA L. S. (1975) DNA-binding chromosomal nonhistone protein isola- tion, characterization and tissue specificity. Biochemistry 14, 4552-4558.
CHIU J.-F., TSAI Y. H., SAKUMA K. & HN1LICA L. S. (1975) Regulation of in ~itro mRNA transcription by a fraction of chromosomal proteins. J. biol. Chem. 250, 9431-9433.
HIRASAWA E., TAKAHASHI E. & MATSUMOTO H. (1978) A transcription inhibitor in non-histone proteins of germinated pea cotyledon. PI. Cell Physiol. 19, 599 608.
JAMES G. T., YEOMAN L. C., MATSUI S., GOLDBERG A. H. & BUSCH H. (1977) Isolation and characterization of non- histone chromosomal protein C-14 which stimulates RNA synthesis. Biochemistry 16, 2384~2389.
KOSTRABA N. C. & WANG T. Y. (1972) Differential acti- vation of transcription of chromatin by non-histone frac- tion. Biochim. biophys. Acta 262, 169 181.
KOSTRABA N. C. & WANG T. Y. (1975) Inhibition of tran- scription in vitro by a non-histone protein isolated from
Ehrlich ascites tumor chromatin. J. biol. Chem. 250, 8938 8942.
KRAJEWSKA W. M., BRIGGS R. C. & HNILICA L. S. (1979) Cell-specific antigens in chicken erythroid nuclei: species specificity. Biochemistry 18, 5720-5725.
KRAJEWSKA W. M. • KLYSZEJKO-STEFANOWICZ L. (1980) Comparative studies on chromatin proteins from chicken thrombocytes and erythrocytes. Biochim. biD- phys. Acta 624, 522 530.
KRAJEWSKA W. M., BRIGGS R. C., CHIU J.-F. & HNILICA L. S. (1980) Immunologically specific complexes of chromo- somal nonhistone proteins with deoxyribonucleic acid in chicken erythroid nuclei. Biochemistry 19, 4667 4673.
LASKY L. & TOBIN A. J. (1979) Transcriptional regulation in avian erythroid cells. Biochemistry 18, 1594-1598.
LEGRAVEREND M. & GLAZER R. I. (1980) Characterization of non-histone chromosomal protein which stimulates RNA polymerase II. Biochim. biophys. Acta 607, 92-101.
LOWRY O. H., ROSENBROUGH N. J., FARR A. L. & RANDALL R. J. (1951) Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 26%275.
MARMUR J. (1961) A procedure for the isolation of deoxy- ribonucleic acid from microorganisms. J. molec. Biol. 3, 208-218.
OLINS A. L. & OLINS D. E. {1974) Spheroid chromatin units (v bodies). Science 183, 330-332.
REFF M. E. & DAVlDSON R. L. (1979) In vitro DNA depen- dent synthesis of globin RNA sequences from erythro- leukemic cell chromatin. Nucleic Acid Res. 6, 275-287.
VAN HOLDE K. E., SAHASRABUDDHE C. G. & SHAW B. R. (1974) A model for particulate structure of chromatin. Nucleic Acid Res. I, 1579-1575.
WEBER K. & OSBORN M. (1969) The reliability of molecular weight determination by dodecyl sulphate polyacryl- amide gel electrophoresis. J. biol. Chem. 244, 440(~4411.