Embed Size (px)
Citation preview

(CANCER RESEARCH 45, 5734-5740, November 1985]
Effect of Retinole Acid on Cellular Content and Human Parathyroid HormoneActivation of Cyclic Adenosine 3':5'-Monophosphate-dependent Protein
Kinase Isoenyzmes in Clonal Rat Osteogenic Sarcoma Cells1
Stephen A. Livesey, Kong Wah Ng, Gregory R. Collier, Minoru Kubota, Alton L. Steiner, and T. John Martin2
Department of Medicine, University ot Melbourne. Repatriation General Hospital. Heidelberg. 3081. Australia [S. A. L. K. W. N., G. R. C., M. K.. T. J. M.], and TheUniversity ot Texas, Health Science Center at Houston. Medical School, Houston. Texas [A. L S.I
ABSTRACT
Pretreatment with 10~8 M retinole acid for 4 days caused
changes in three distinct components of the parathyroid hormone(PTH)-stimulated cyclic adenosine 3':5'-monophosphate re
sponse in a clonal rat osteogenic sarcoma cell line, UMR 106-06: (a) the amplitude of the cyclic adenosine 3':5'-monophos-
phate response to PTH was moderately increased after pretreatment with retinoic acid; (b) while the cellular content of the twoisoenzymes of the cyclic adenosine 3':5'-monophosphate-de-
pendent protein kinase was approximately equal in control cells,retinoic acid pretreatment was associated with a marked increase in the ratio of type II to type I holoenzyme activity. Thischange might be due to a decrease in the type I holoenzyme assuggested by immunofluorescence detection of decreased typeI regulatory subunit in fixed cells together with the relativedecrease in type I holoenzyme determined biochemically; (c)there was a marked alteration of the pattern of PTH-stimulated
protein kinase isoenzyme activation from predominantly type Iisoenzyme in control cells to almost exclusively type II isoenzymein retinoic acid-treated cells. Growth inhibition by submaximalamounts of PTH and retinoic acid when added together wasgreater than that for either agent alone.
INTRODUCTION
The ability of retinoids to induce differentiation in murine andhuman stem cell lines has stimulated interest in their use tosuppress carcinogenesis by controlling differentiation in targetcells (1, 2). In embryonal carcinoma cells (3) and human hema-topoietic cells (4, 5) a synergistic interaction between RA3 and
agents that elevate cellular cyclic AMP levels has been demonstrated during the induction of differentiation. The molecularmechanism(s) of this interaction is unknown although it has beenspeculated that RA may either induce a cyclic AMP-dependent
protein kinase or alternatively the production of a substrate whichis phosphorylated in the process of differentiation (6). Otherpossibilities include alteration of the hormone stimulated cyclicAMP response or change in the pattern of selective activation ofthe cyclic AMP protein kinase isoenzymes. This paper describesthe results of a study undertaken to determine the molecular
1This work was supported by grants from the National Health and MedicalResearch Council, the Australian Government Department of Veterans' Affairs, andthe Children's Leukaemia and Cancer Foundation.
1To whom requests for reprints should be addressed.3The abbreviations used are: RA, all-trans-fi-retinoic acid; PCS, fetal calf serum;
A-FCS. vitamin A-deficient PCS; PBS. phosphate-buffered saline (NaCI. 8 g/liter-KCI. 0.2 g/liter-Na2HPO<, 1.15 g/liter-KH2PO., 0.2 g/liter, pH 7.4); »-MEM, «-
modified minimal essential medium; PTH. parathyroid hormone.Received 1/7/85; revised 5/23/85; accepted 7/22/85.
mechanisms involved in the interaction between RA and PTH.The cloned cells were established from cultures of a transplant-
able rat osteogenic sarcoma which has been extensively characterized with respect to its differentiated properties as themalignant counterpart of the osteoblast and has been used as astable PTH-responsive system (7).
The interaction between retinoic acid and the cyclic AMP-
dependent effects of PTH was investigated by studying theirindividual and combined effects on hormone stimulated adenylatecyclase, the basal levels of the cyclic AMP-dependent proteinkinase isoenzymes, the selective activation of the cyclic AMP-
dependent protein kinase isoenzymes, and the growth of cells inmonolayer culture. A previous study has established that thepattern of selective activation of cyclic AMP-dependent proteinkinase isoenzymes differs between normal and malignant oste-
oblasts for the same hormonal stimulus (8). We therefore set outto determine whether treatment with retinoic acid may alter thepatterns of response in malignant osteoblasts to reflect a moredifferentiated phenotype and to relate this effect to their combined effect on cell growth.
MATERIALS AND METHODS
Synthetic human parathyroid hormone (1-34), 5000 units/mg, wasobtained from Beckman Pty., Ltd., Palo Alto, ÇA.«-MEM and fetal calf
serum were purchased from Flow Laboratories. Mt. Waverley, Victoria,Australia. All-frans-/i-retinoic acid was supplied by Sigma Chemical Co.,
St. Louis, MO. Bovine serum albumin was obtained from Armour Pharmaceutical Co., -y-AT32P was obtained from the Radiochemical Centre,
Amersham. Cyclic AMP was purchased from Boehringer Mannheim, Mt.Waverley, Australia. Synthetic heptapeptide protein kinase substrate wassynthesized as described previously (9) and was kindly provided by Dr.B. E. Kemp. Phosphocellulose paper (P81) and microgranular DE52aniónexchange resin were products of Whatman Inc., Maidstone UnitedKingdom. Specific rabbit antisera against R, and R2 regulatory subunitsof cAMP dependent protein kinase were prepared and characterized asdescribed (10).
Cell Culture. UMR 106-06 cells were routinely grown in monolayerculture in 75-cm2 flasks (Lux Scientific Corp., Newbury Park, CA). The
culture medium was «-MEMcontaining 4-(2-hydroxyethyl)-1 -piperazinee-
thane sulfonic acid (20 HIM), gentamicin (80 mg/liter), minocyclin (1 mg/liter), and 10% FCS. They were incubated at 37°C and equilibrated in
5% CO2 in air. Subculture was performed weekly using 0.0125% trypsinand 0.5 HIM disodium EDTA in PBS to harvest the cells.
The UMR 106-06 cell line is one of several subclones derived from
single cells of the clonal line UMR 106 (7) which were allowed to formcolonies in soft agar (11). In the experiments reported here only cellsfrom early passage (up to passage 12) after subcloning were used.
Growth Studies. UMR 106-06 cells were subcultured into 9.6-cm2 6-
place multiwell dishes (Linbro, Flow Laboratories, VA) in «-MEMsupplemented with 10% FCS. Twenty-four h later the cells were washed with
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5734
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

RETiNOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
serum-free medium and incubation was continued for a further 24 h with2 ml/well of «-MEM containing 2% A-FCS (12) before the appropriatehormones were added on day 0. Retinoic acid was dissolved in spectro-
scopic grade absolute ethanol and stored in liquid nitrogen for up to 2weeks. This was added as a 1000-fold concentrated solution to theincubation medium. Control wells received a similar aliquot of hormone-
free carrier so that the final concentration of ethanol was 0.1%. HumanPTH (1-34) was dissolved in 0.01 M acetic acid to make up stocksolutions containing 1 mg/ml. Further dilutions were made up in «-MEM
with 2% A-FCS before it was added to the incubation medium. All
hormone additions were performed in subdued light and the plates werewrapped in aluminum foil to protect RA from light. Fresh hormone wasadded daily when the medium was changed. Cells from replicate multi-wells were harvested with 1 ml/well of 0.0125% trypsin-0.5 mw disodium
EDTA and cell numbers were measured in a Coulter Counter (Model
DN).Cyclic [3H]AMP Formation. Intact cell adenylate cyclase activity was
measured as the amount of cyclic [3H]AMP generated from cells in whichthe ATP was labeled by incorporation of [3H]adenine (13). Replicatecultures of UMR 106-06 cells in 4.5-cm212-plate multiwell dishes (Costar,Flow Laboratories, United Kingdom) were incubated for 2 h with [3H]-
adenine (1-2 ¿iCi/ml).The cells were washed and incubated for 20 minin medium containing 10~3 M isobutyl methylxanthine. Hormone treat
ments were added and incubation was continued for 10 min at 37°C.
Reactions were stopped by removing supernatants, adding 100 /il 1.2 Mtrichloroacetic acid followed by 750 ^l of 5 mw adenine, ATP, AMP, andcyclic AMP together with 100 n\ of cyclic AM32P (5000 cpm) to calculate
recoveries. These samples were then subjected to sequential chroma-tography on Dowex and neutral alumina to purify the cyclic [3H]AMP
formed in the cells.Separation of Cyclic AMP-dependent Protein Kinase Isoenzymes
and Measurement of Selective Isoenzyme Activation by PTH. Themethod for the separation of cyclic AMP-dependent protein kinase
isoenzymes has been described in detail previously (8, 11, 14, 15). Thecells were grown in 75-cm2 tissue culture flasks and prepared as de
scribed in the growth studies. Control cells were incubated in «-MEMwith 2% A-FCS and hormone-free carrier while treated cells were incubated in the same medium containing 10~8 M retinoic acid for 4 days.
Fresh hormone was added when the medium was changed on alternatedays. Human PTH (1-34) dilutions were made up in «-MEMwith 2% A-FCS and added to the cells for a 10-min incubation at 37°C.At the end
of the incubation the cells were washed and scraped from vessel surfacesin an excess (10 ml) of column starting buffer (10 mw 2-(/V-morpho-lino)ethanesulfonic acid-NaOH, pH 6.8-0.2 mw disodium EDTA-0.1 rtiM
dithiothreotol) (8), with the aim of minimizing the possibility of posicelirupture activation (16). Dilute cell suspensions were immediately sonicated and applied to 2 ml DE52 columns (Whatman) equilibrated previously in starting buffer and then washed with 50 ml of the same buffer.When all columns were loaded they were simultaneously eluted with alinear gradient of NaCI 0 to 0.6 M in the starting buffer (total gradient 40ml/column). Eluate was collected as 20-drop fractions.
Reaction mixture for the measurement of cyclic AMP-dependent protein kinase activity contained 20 HIM2-(W-morpholino)ethanesulfonic acid-
NaOH buffer (pH 6.8), 10 mw magnesium acetate, 1 HIMdisodium EDTA,15 HIM bovine serum albumin, 0.125 mM >-AT32P (400-500 cpm/pmol)
and 156 ^M synthetic peptide substrate, total volume 80 ^l. Reactionswere commenced by adding 30-^1 aliquots of column fractions andincubating for 30 min at 37°C. Reactions were terminated by spotting
25 n\ of the mixture onto Whatman P81 paper discs held in a Perspexmanifold. Isoenzyme activity was measured in the presence of 6.25 ¿IMcyclic AMP. Discs were then washed in four changes of 5 HIMphosphoricacid for 10 min each and rinsed in absolute ethanol before counting usinga toluene scintillant in a Packard automatic scintillation counter. Controlexperiments using equivalent numbers of unstimulated cells as an external control (11,14, 17) showed that no postextraction activation by PTHoccurred at the concentrations of PTH used in the experiments. Per
centage of activation of individual isoenzymes of cyclic AMP-dependent
protein kinase was calculated as described previously (8,17).Immunocytochemistry. UMR 106-06 cells were subcultured onto
sterile 18-mm2 glass coverslips in 9.6-cm2 6-plate multiwell dishes (Linbro
Flow Laboratories, McLean, VA) in «-MEM supplemented with 10%FCS. Twenty-four h later cells were washed with serum-free mediumand then incubated for 4 days with 2 ml/well of «-MEM containing 2%A-FCS plus carrier or RA. Carrier or RA was added thereafter on alternatedays after a medium change. PTH (1-34) in «-MEM plus 2% A-FCS asa 50-/jl aliquot was added for 10 min on the day of the experiment. Cells
on coverslips were then washed three times in PBS (3 min) and fixed in2% formaldehyde (electron microscopy grade) in PBS at room temperature for 4 min. Coverslips were then washed an additional three times inPBS (3 min), permeabilized with acetone at -20%C for 2 min, and
washed again in three changes of PBS. The first antibody portion wasadded as a 25-n\ aliquot to the surface of the coverslip in a humidified
chamber and incubated for 45 min at room temperature. Following PBSwashing the second antibody portion was added as a 25-^1 aliquot to
the surface of the coverslip for 30 min at room temperature. After 3further washes in PBS coverslips were mounted in Aquamount (LernerLaboratories, New Haven, CT) and viewed with epifluorescence using aNikon Fluophot microscope and photographed using llford XPI400 filmwith ASA set at 200. Negatives were developed using C41 color processing. For studies of a single series photographs were taken in thememory mode of the Nikon UFX11 photographic equipment to ensureconstant exposure time.
RESULTS
Cyclic AMP Formation. The UMR 106-06 cells provide asensitive model to study the action of PTH. A dose-dependentincrease in cyclic AMP formation was seen when the cells wereacutely stimulated by PTH. Pretreatment with 10~8 M RA for 5
days resulted in an increase in cyclic AMP in the basal state. Inaddition when compared to control cells RA-treated cells gener
ated significantly more cyclic AMP when challenged by 3.3 x1Q-10M PTH and higher doses (Table 1).
Basal Profiles of Cyclic AMP-dependent Protein KinaseIsoenzymes. UMR 106-06 cells contain both types I and IIisoenzymes of the cyclic AMP-dependent protein kinase in ap
proximately equal amounts (11). When cells were exposed to10"8 M RA for 4 days a relative decrease in the amount of type I
isoenzyme occurred (Chart 1, fop). This change was reflected inan alteration in the ratio of the activity of isoenzyme II to I from0.96 in control to 3.3 in RA-treated cells. This could be explained
Table 1EHect of RA on cyclic [3H¡AMPgeneration in response to human PTH (1-34) in
UMR 706-06 cellsUMR 106-06 cells were subcultured in triplicate as described in "Materials and
Methods." After the cells had grown for 5 days in the absence (control) or presenceof 10~8 M RA they were stimulated for 10 min with increasing doses of human PTHand the amount of cyclic [3H]AMP formed was measured. Fresh hormone was
added when the medium was changed on alternate days.
Human PTH (1-34)
(M)03.3
x 10~'23.3 x IO'"3.3 x 10-'°3.3 x 1fr93.3 x 10-"3.3 x 10-7Cyclic
[3H]AMP (cpm/106cells)Control131
±55a
336 ±15494 ±133685 ±78
4,284+ 16616,956 ±43423,338 ±72RA(10~eM)361
44453 78697 52
1,024 305,198 170
22,708 85134,31 5 ±1,229P<0.05
Not significantNot significant
<0.02<0.01<0.005<0.005
" Mean ±SE.
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5735
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
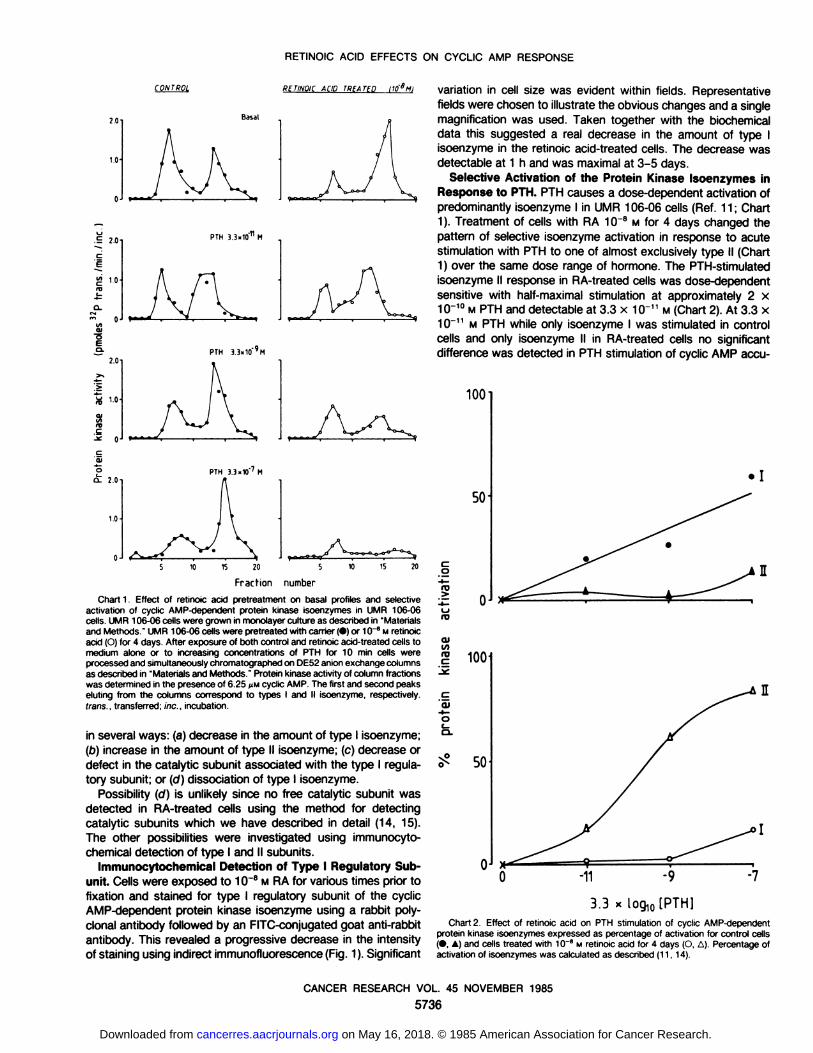
RETINOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
RETIHO/C ACID 7KEATCD 110"Ml
2.0 Basal
.£ 2.0i
y i o
PTH 3.3.10" M
PTH 3.3.10"'M
.fi
et 2.0i
10
PTH 3.3.10'' M
20
Fraction number
Chart 1. Effect of retinoic acid pretreatment on basal profiles and selectiveactivation of cyclic AMP-dependent protein kmase isoenzymes in UMR 106-06cells. UMR 106-06 cells were grown in monolayer culture as described in "Materialsand Methods " UMR 106-06 cells were pretreated with carrier (•)or 10~8 M retinoic
acid (O) for 4 days After exposure of both control and retinoic acid-treated cells tomedium alone or to increasing concentrations of PTH for 10 min cells wereprocessed and simultaneously chromatographed on DE52 aniónexchange columnsas described in "Materials and Methods." Protein kinase activity of column fractions
was determined in the presence of 6.25 MMcyclic AMP. The first and second peakseluting from the columns correspond to types I and II isoenzyme, respectively.trans., transferred; inc.. incubation.
in several ways: (a) decrease in the amount of type I isoenzyme;(b) increase in the amount of type II isoenzyme; (c) decrease ordefect in the catalytic subunit associated with the type I regulatory subunit; or (d) dissociation of type I isoenzyme.
Possibility (d) is unlikely since no free catalytic subunit wasdetected in RA-treated cells using the method for detecting
catalytic subunits which we have described in detail (14, 15).The other possibilities were investigated using immunocyto-
chemical detection of type I and II subunits.Immunocytochemical Detection of Type I Regulatory Sub-
unit. Cells were exposed to 10~8 M RA for various times prior to
fixation and stained for type I regulatory subunit of the cyclicAMP-dependent protein kinase isoenzyme using a rabbit poly-clonal antibody followed by an FITC-conjugated goat anti-rabbit
antibody. This revealed a progressive decrease in the intensityof staining using indirect immunofluorescence(Fig. 1). Significant
variation in cell size was evident within fields. Representativefields were chosen to illustrate the obvious changes and a singlemagnification was used. Taken together with the biochemicaldata this suggested a real decrease in the amount of type Iisoenzyme in the retinoic acid-treated cells. The decrease wasdetectable at 1 h and was maximal at 3-5 days.
Selective Activation of the Protein Kinase Isoenzymes inResponse to PTH. PTH causes a dose-dependent activation ofpredominantly isoenzyme I in UMR 106-06 cells (Ref. 11; Chart1). Treatment of cells with RA 10~8 M for 4 days changed the
pattern of selective isoenzyme activation in response to acutestimulation with PTH to one of almost exclusively type II (Chart1) over the same dose range of hormone. The PTH-stimulatedisoenzyme II response in RA-treated cells was dose-dependentsensitive with half-maximal stimulation at approximately 2 x10~10M PTH and detectable at 3.3 x10~" M (Chart 2). At 3.3 x10~11M PTH while only isoenzyme I was stimulated in control
cells and only isoenzyme II in RA-treated cells no significantdifference was detected in PTH stimulation of cyclic AMP accu-
100
50
o
re>ure
<uI/)re
15
'oÃ
oa.
100
50
3.3Chart 2. Effect of retinoic acid on PTH stimulation of cyclic AMP-dependent
protein kinase isoenzymes expressed as percentage of activation for control cells(•,A) and cells treated with 10~8 M retinoic acid for 4 days (O, A). Percentage of
activation of isoenzymes was calculated as described (11,14).
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5736
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
RETINOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
mulation (Table 1).Immunocytochemical Detection of Type II Regulatory Sub-
unit. In contrast with the data for type I regulatory subunit RA-
treated cells showed no significant change in the intensity ofstaining when an antibody to type II regulatory subunit was used(Fig. 2). The polyclonal antibody used in these experiments hasa slightly higher affinity for the dissociated regulatory subunitthan for the intact holoenzyme (18). When stimulated by 3.3 x10"7 M PTH for 10 min the intensity of the staining pattern was
increased in RA-treated cells but not in control cells treated with
PTH. Differences in the intensity of staining between control andRA-treated cells were more subtle compared to Fig. 1. Thereforea lower magnification was used to include more cells per field.These results taken together with the biochemical data areconsistent with an isoenzyme II response in RA-treated cells
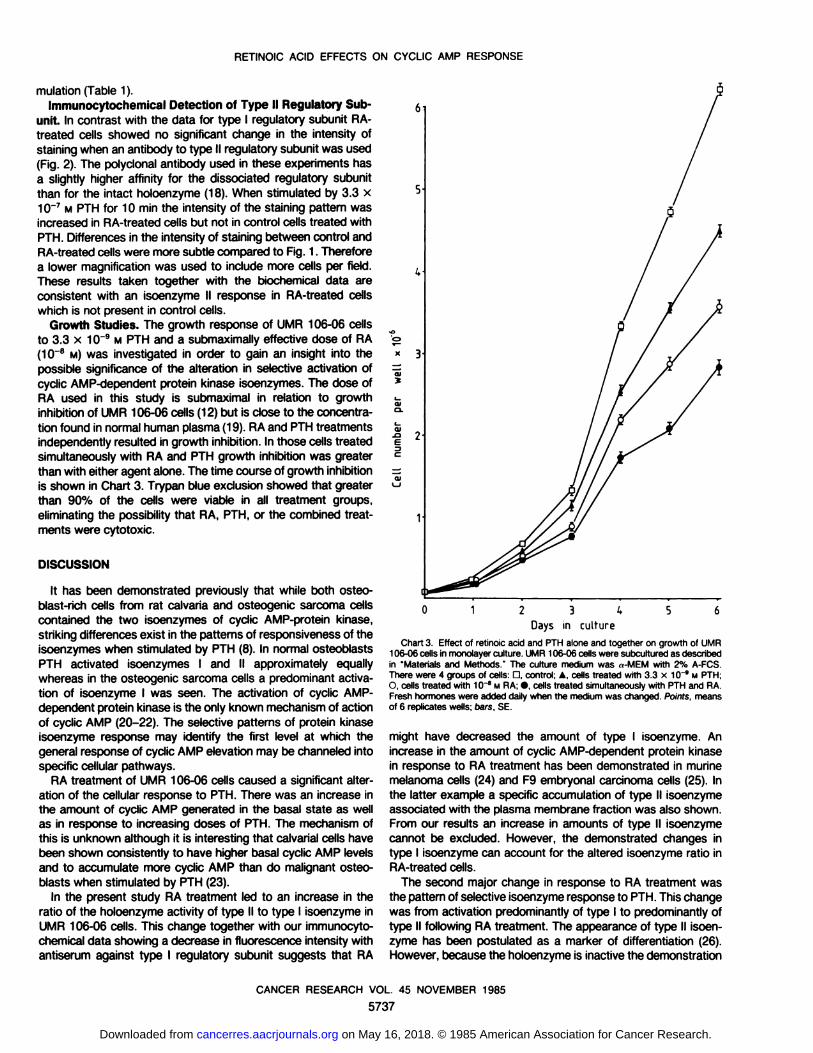
which is not present in control cells.Growth Studies. The growth response of UMR 106-06 cells
to 3.3 x 10"9 M PTH and a submaximally effective dose of RA(10~8 M) was investigated in order to gain an insight into the
possible significance of the alteration in selective activation ofcyclic AMP-dependent protein kinase isoenzymes. The dose ofRA used in this study is submaximal in relation to growthinhibition of UMR 106-06 cells (12) but is close to the concentra
tion found in normal human plasma (19). RA and PTH treatmentsindependently resulted in growth inhibition. In those cells treatedsimultaneously with RA and PTH growth inhibition was greaterthan with either agent alone. The time course of growth inhibitionis shown in Chart 3. Trypan blue exclusion showed that greaterthan 90% of the cells were viable in all treatment groups,eliminating the possibility that RA, PTH, or the combined treatments were cytotoxic.
DISCUSSION
It has been demonstrated previously that while both osteo-blast-rich cells from rat calvarÃaand osteogenic sarcoma cellscontained the two isoenzymes of cyclic AMP-protein kinase,
striking differences exist in the patterns of responsiveness of theisoenzymes when stimulated by PTH (8). In normal osteoblastsPTH activated isoenzymes I and II approximately equallywhereas in the osteogenic sarcoma cells a predominant activation of isoenzyme I was seen. The activation of cyclic AMP-
dependent protein kinase is the only known mechanism of actionof cyclic AMP (20-22). The selective patterns of protein kinase
isoenzyme response may identify the first level at which thegeneral response of cyclic AMP elevation may be channeled intospecific cellular pathways.
RA treatment of UMR 106-06 cells caused a significant alter
ation of the cellular response to PTH. There was an increase inthe amount of cyclic AMP generated in the basal state as wellas in response to increasing doses of PTH. The mechanism ofthis is unknown although it is interesting that calvarial cells havebeen shown consistently to have higher basal cyclic AMP levelsand to accumulate more cyclic AMP than do malignant osteoblasts when stimulated by PTH (23).
In the present study RA treatment led to an increase in theratio of the holoenzyme activity of type II to type I isoenzyme inUMR 106-06 cells. This change together with our immunocyto-
chemical data showing a decrease in fluorescence intensity withantiserum against type I regulatory subunit suggests that RA
0123^56
Days in culture
Chart 3. Effect of retinole acid and PTH alone and together on growth of UMR106-06 cells in monolayer culture. UMR 106-06 cells were subcultured as describedin "Materials and Methods " The culture medium was »-MEM with 2% A-FCS.There were 4 groups of cells: O, control; A. cells treated with 3.3 x 10~9 M PTH;O, cells treated with 10~" M RA; •cells treated simultaneously with PTH and RA.
Fresh hormones were added daily when the medium was changed. Points, meansof 6 replicates wells; oars, SE.
might have decreased the amount of type I isoenzyme. Anincrease in the amount of cyclic AMP-dependent protein kinase
in response to RA treatment has been demonstrated in murinemelanoma cells (24) and F9 embryonal carcinoma cells (25). Inthe latter example a specific accumulation of type II isoenzymeassociated with the plasma membrane fraction was also shown.From our results an increase in amounts of type II isoenzymecannot be excluded. However, the demonstrated changes intype I isoenzyme can account for the altered isoenzyme ratio inRA-treated cells.
The second major change in response to RA treatment wasthe pattern of selective isoenzyme response to PTH. This changewas from activation predominantly of type I to predominantly oftype II following RA treatment. The appearance of type II isoen-
zyme has been postulated as a marker of differentiation (26).However, because the holoenzyme is inactive the demonstration
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5737
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

RETINOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
of an altered pattern of selective activation to a type II responsemay be of fundamental importance to the altered cellular state.This change together with the morphological effects reported ina previous paper (12) is consistent with a more differentiatedphenotype. Whether the changes in the isoenzymes are responsible for or the result of the differentiated state is not yet known.
The mechanism of the above changes have yet to be defined.RA treatment increases the cyclic AMP generation in responseto PTH. However, this effect was not significant at the PTHconcentration where the change in selective activation was firstevident. Furthermore there is no significant difference in the K05for cyclic AMP for the two isoenzymes (8). A decrease in theamount of type I isoenzyme could account for the lack of type Iresponse following retinoic acid treatment. This change howeverdoes not explain why in RA-treated cells isoenzyme II is activated
by PTH at levels of cyclic AMP generation which did not activatethe ¡soenzymein control cells. One possibility is redistribution ofthe type II isoenzyme following RA treatment.
These results demonstrate that RA treatment can change theresponse of malignant cells to a more differentiated pattern.These findings are consistent with our previous report thatselective activation of type II isoenzyme by calcitonin is associated with growth inhibition of breast cancer cell line T47 D (15).A similar association has been found by other investigators (27-
29).In summary the UMR 106-06 cell line provides an excellent
model to study the interactions between RA and hormone effects, in this case the interrelationships between the actions ofRA and the cyclic AMP system. Growth inhibition by RA isclosely related to the induction of differentiation. Apart from thepattern of activation of cyclic AMP-dependent protein kinase
isoenzymes other markers of the differentiated osteoblast phenotype are currently under investigation.
REFERENCES
1. Sporn, M. B., and Newton, D L. Chemoprevention of cancer with retinoids.Fed. Proc.. 38: 2528-2534. 1979.
2. Sporn, M. B., and Roberts. A. B. Role of retinoids in differentiation andcarcinogenesis. Cancer Res., 43: 3034-3040. 1983.
3. Strickland, S., Smith, K. K., and Marotti, K. R. Hormonal induction of differentiation in teratocarcinoma stem cells: Generation of parietal endoderm byretinoic acid and dibutyryl cAMP. Cell, 27: 347-355, 1980.
4. Breitman, T. R., and Keene, B. R. Synergistic induction of differentiation of thehuman promyelocytic leukemia cell line, HL-60, by retinoic acid and prosta-
glandin E. Proc. Am Assoc. Cancer Res., 22. 55. 1981.5 Olsson, l. L., and Breitman. T R. Induction of differentiation of the human
histiocytic lymphoma cell line U-937 by retinoic acid and cyclic adenosine 3' :5'-
monophosphate-inducing agents Cancer Res., 42: 3924-3927, 1982.6 Olsson, I. L., Breitman. T. R., and Gallo. R. C. Priming of human myeloid
leukemic cell lines HL-60 and U-937 with retinoic acid for differentiation effectsof cyclic adenosine 3':5'-monophosphate-inducing agents and a T-lympho-
cyte-derived differentiation factor. Cancer Res.. 42. 3928-3933, 1982.7 Partridge, N. C., Alcorn, O., Michelangeli. V. P., Ryan, G.. and Martin, T. J.
Morphological and biochemical characterization of four clonal osteogeneicsarcoma cell lines of rat origin Cancer Res 43. 4308-4314. 1983.
8. Livesey, S. A., Kemp, B. E., Re, C. A., Partridge, N. C , and Martin T. J.
Selective hormonal activation of cyclic AMP-dependent protein kinase isoen-zymes in normal and malignant osteoblasts. J Biol Chem., 257: 14983-14987,1982.
9. Kemp, B. E. Phosphorylation of acyl and dansyl derivatives of the peptide Leu-Arg-Arg-Ala-Ser-Leu-Gly by the cyclic AMP-dependent protein kinase. J. Biol.Chem.. 255: 2914-2918,1980.
10. Koide, Y., Beavo, J. A., Kapoor, C. L., Spruill, W. A , Huang, H. L., Levine, S.N., Ong, S. L., Bechtel, P. J., Yount, W. J , and Steiner, A. L. Hormonal effectson the immunocytochemical location of 3',5'-cyclic adenosine monophos-phate-dependent protein kinase in rat tissues. Endocrinology, 709: 2226-
2238, 1981.11. Forrest, S. M., Ng, K. W., Findlay. D. M., Michelangeli, V. P., Livesey, S. A.,
Partridge, N. C., Zajac, J. D., and Martin, T. J. Characterization of an osteo-blast-like clonal cell line which responds to both parathyroid hormone andcalcitonin. Calcif. Tissue Int., 37: 51-56, 1985.
12. Ng, K. W., Livesey, S. A., Collier, F., Gummer, P. R., and Martin, T. J. Effectof retinoids on the growth, ultrastructure, and cytoskeletal structures ofmalignant rat osteoblasts. Cancer Res., 45. 5106-5113, 1985.
13. Rodan, S. B., Insogna, K. L., Vignery. AM-L.. Stewart, A. F.. Boradus, A. E.,D'Souza. S. B., Bertolini. D. R., Mundy, G. R., and Rodan. G. A. Factors
associated with humoral hypercalcemia of malignancy stimulate adenylatecyclase in osteoblastic cells. J. Clin. Invest., 72: 1511-1515,1983.
14. Livesey, S. A , Collier. G.. Zajac, J. D.. Kemp, B. E.. and Martin, T. J.Characteristics of selective activation of cyclic AMP-dependent protein kinaseisoenzymes by calcitonin and PGE2 in human breast cancer cells. BiochemJ., 224:361-370, 1984.
15. Ng, K. W., Livesey, S. A , Larkins, R. G., and Martin, T J. Calcitonin effectson growth and on selective activation of type II isoenzymes of cyclic adenosine3':5'-monophosphate-dependent protein kinase in human breast cancer cells
(T47 D). Cancer Res., 43: 794-800. 1983.16. Palmer, W. K., McPherson. J. M., and Walsh, D. A. Critical controls in the
evaluation of cAMP-dependent protein kinase activity ratios as indices ofhormonal action. J. Biol. Chem., 255: 2663-2666, 1980
17. Zajac, J. D., Livesey, S. A., and Martin, T. J. Selective activation of cyclic AMP-dependent protein kinase by calcitonin in a calcitonin secreting lung cancercell line. Biochem. Biophys. Res. Commun., 722: 1040-1046, 1984.
18. Murtaugh, M. P., Steiner, A. L., and Davies, P. J A. Localization of the catalyticsubunit of cyclic AMP dependent protein kinase in culture cells using specificantibody. J. Cell Biol.. 95: 64-72. 1982.
19 De Ruyter. M. G , Lambert, W. E., and De Leenheer, A. P. Retinoic acid: anendogenous compound of human blood. Unequivocal demonstration of endogenous retinoic acid in normal physiologic conditions. Anal. Biochem., 98: 402-409, 1979.
20 Kuo, J. F.. and Greengard, P. Cyclic nucleotide dependent protein kinase. IV.Widespread occurrence of adenosine 3 ' .5 ' -monophosphate dependent protein
kinase in various tissues and phyla of the animal kingdom. Proc. Nati. Acad.Sci. USA, 64: 1349-1355. 1969
21. Walsh, D. A., and Ashby, C. D. Protein kinases: aspects of their regulation anddiversity. Recent Prog. Horm Res . 29: 329-359, 1973.
22. Flockhart, D. A., and Corbin, J. D Regulatory mechanism in the control ofprotein kinases CRC Grit. Rev. Biochem., 72: 133-185. 1982.
23. Partridge, N. C., Kemp. B. E., Veroni, M. C., and Martin, T. J. Activation ofcyclic AMP-dependent protein kinase in normal and malignant bone cells byparathyroid hormone, prostaglandin E2 and prostacyclin. Endocrinology, 708:220-226,1981.
24. Ludwig, K. W., Lowey, B.. and Niles, R. M. Retinoic acid increases cyclic AMP-dependent protein kinase activity in murine melanoma cells J Biol Chem..255:5999-6002.1980.
25. Piet. A., Evain, D., and Anderson. W. B. Effect of retinoic acid treatment of F9embryonal carcinoma cells on the activity and distribution of cyclic AMP-dependent protein kinase. J Biol. Chem., 257: 889-893, 1982.
26. Lee, P. C., Radloff, D., Schweppe, and Jungmann. R. A. Testicular proteinkinases. Characterization of multiple forms of ontogeny. J. Biol. Chem., 257:914-921. 1976.
27. Byus. C. V., Klimpel, G. R.. Lucas. D. 0., and Russell, D. H. Type I and typeII cyclic AMP-dependent protein kinase as opposite effectors of lymphocytemitogenesis. Nature (Lond.), 268: 63-64. 1977.
28. Cho Chung, Y. S. Cyclic AMP and tumor growth in vivo. In: J. A. Kellen, andR. Hilf (eds.), Influences of Hormones in Tumor Development, Vol. 1, pp. 55-94. Boca Raton. FL: CRC Press Inc.. 1980
29. Cho Chung, Y. S On the mechanisms of cyclic AMP mediated growth arrestof solid tumors. Adv. Cyclic Nucleotide Res., 72: 111-121. 1980.
Fig. 1. Immunocytochemical detection of type I regulatory subunit of cyclic AMP-dependent protein kinase in UMR 106-06 cells after treatment with retinoic acid.Cells were plated on glass coverslips for 24 h prior to treatment with 10"8 M retinoic acid for various times. Cells were then fixed and stained for the type I regulatorysubunit as described in "Materials and Methods." a. control cells; £>,retinoic acid, 1 h; c, retinoic acid, 3 days; d. retinoic acid, 5 days; e, control (fluorescein isothiocyanate-
conjugated antibody alone).Fig. 2 Immunocytochemical detection of type II regulatory subunit of cyclic AMP-dependent protein kinase in UMR 106-06 cells following treatment with retinoic acid
for 4 days before treatment for 10 min with PTH. Cells were plated on glass coverslips for 24 h prior to treatment with 10~8 M retinoic acid (b and d) or carrier (a and c)for 4 days. Cells were then exposed to 3.3 x 10~7 M PTH (c and d) or carrier (a and b) for 10 min prior to fixation and staining for the type II regulatory subunit asdescribed in "Materials and Methods." e. control (fluorescein isothiocyanate-conjugated antibody alone)
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5738
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

RETINOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
1
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5739
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

RETINOIC ACID EFFECTS ON CYCLIC AMP RESPONSE
CANCER RESEARCH VOL. 45 NOVEMBER 1985
5740
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1985;45:5734-5740. Cancer Res Stephen A. Livesey, Kong Wah Ng, Gregory R. Collier, et al. Clonal Rat Osteogenic Sarcoma Cells-Monophosphate-dependent Protein Kinase Isoenyzmes in
′:5′Parathyroid Hormone Activation of Cyclic Adenosine 3Effect of Retinoic Acid on Cellular Content and Human
Updated version
http://cancerres.aacrjournals.org/content/45/11_Part_2/5734
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/45/11_Part_2/5734To request permission to re-use all or part of this article, use this link
on May 16, 2018. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























