Embed Size (px)
Citation preview

E. coli exhibits an important behavioral response known as chemotaxis - motion toward desirable chemicals (usually nutrients) and away from harmful ones - which is also shared by various other prokaryotic and eukaryotic cells. The cell’s motion consists of series of “runs” puntuated punctuated by “ tumbles”.
Robust Perfect Adaptation in Bacterial Chemotaxis Yang Yang & Sima Setayeshgar Department of Physics, Indiana University, Bloomington
My research: I am a physics graduate student working in theoretical biophysics.
Conclusions Successful implementation of the augmented model of the chemotaxis signal transduction network in E. coli that explicitly takes into account robust perfect adaption Preliminary results on projections of robustness manifolds in parameter space
What is E coli ? Why study it?
It is considered to be an ideal model organism for understanding the behavior of cells at the molecular level from the perspectives of several scientific disciplines-anatomy, genetics, chemistry and physics- because of :
• Ease of experimentation, through microscopy and genetic analysis
• Small genome (4288 genes), most of which encode proteins
1
2
What is prefect adaptation? Why is it important?
This property allows the system to compensate for the presence of continued stimulation and to be ready to respond to further stimuli.
Perfect adaptation is an important and generic property of signaling systems, where the response (e.g. running bias in chemotaxis) returns precisely to the pre-stimulus level while the stimulus persists.
What is the chemotaxis signal transduction network in E. coli?
Increasing attractants or Decreasing repellents
What is robustness? Why is it important?The E. coli chemotaxis signal transduction network exhibits robust perfect adaptation, where the concentration of CheYp returns to its prestimulus value despite large changes in the values of many of the biochemical reaction rate constants. These rate constants depend on concentrations of enzymes, which are often present in small copy numbers, making fluctuations in their numbers significant.
5
8
Motivation
Chemotaxis in E. coli involves temporal measurement of the change in concentration of an external stimulus. This is achieved through the existence of fast and slow reaction time scales, in the chemotaxis signal transduction network: fast measurement of the current external concentration is compared with the cell’s “memory” of the concentration some time ago to determine whether to extend a run in a given direction or to tumble, thereby randomly selecting a new direction.
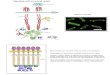
E. coli is a single-celled organism that lives primary in our intestines. It is approximately 1-2 microns long and 1 micron in diameter, and weights 1 picogram. Each cell has 4-6 flagella, approximately 10-20 microns long, driven by an intracellular rotary motor operated by the protonmotive force.
The chemotaxis signal transduction pathway in E. coli – a network of ~50 interacting proteins – converts an external stimulus (change in concentration of chemoattractant / repellent) into an internal stimulus (change in concentration of intracellular response regulator, CheYp) which in turn interacts with the flagella motor to bias the cell’s motion.
It is used as a well-characterized model system for the study of properties of (two-component) cellular signaling networks in general.
Thus, E. coli is able to respond to changes in chemoattractant concentrations spanning 5 orders of magnitude! Similarly, the vertebrate visual system responds to changes in light intensity spanning 10 orders of magnitude during the night-day cycle.
The biochemical basis of robustness of perfect adaptation is not as yet fully understood. In this work, we develop a novel method for elucidating regions in parameter space of which the E. coli chemotaxis network adapts perfectly:
Ref: H. C. Berg, “Motile behavior of bacterial”, Physics Today, January 2000
Ref: P. A. Spiro, J. S. Parkinson, and H. G. Othmer, “A model of excitation and adaptation in bacterial chemotaxis”, Proc. Natl. Acad. Sci. USA 94, 7263(1997)
Ref: N.Barkai & S. Leibler, “Robustness in simple biochemical network”, Nature 387, 913(1997)
2
3
4
Chemical reactions: Ligand binding
Methylation
Phosphorylation
)()( )(7/7~5/5
)( CheRLTCheRTL pnkmkkmk
pn
ppnmkmk
ppn
pnckck
pn
CheBTLCheBTL
CheRTLCheRTL
)(14~1
)(
)(14~1
)(
)()(
)()(
PCheBCheB
PCheYCheZCheZCheY
BCheYCheRTBCheYCheRTL
ADPCheRTLATPCheRTL
kmbp
kmyp
pnbky
np
npkk
n
)()()()()(
)()()()()(
9~7
Broader impactThis method should have applicability to other cellular signal transduction networks and engineered systems that exhibit robust homeostasis.
Algorithm Computational results
The shapes of resulting manifolds determine relationships between reaction parameters (for example, methylation and phosphorylation rates) that must be satisfied in order for the network to exhibit perfect adaptation, thereby shedding light on biochemical steps and feedback mechanisms underlying robustness.
Given lack of complete data on values of in vivo reaction rates, the numerical ranges of the resulting manifolds will shed light on values of unknown or partially known parameters.
The main idea is to:
START with a fine-tuned model of chemotaxis network
that: reproduces key features of experiments (adaptation times to small and large ramps, perfect adaptation of the steady state value of CheYp)
is NOT robustAUGMENT the model explicitly with the requirements that: steady state value of CheYp
values of reaction rate constants, are independent of the external stimulus, s,
thereby achieving robustness of perfect adaptation.
s
k
F
u
skuFdt
ud
0);;(
: state variables
: reaction kinetics
: reaction constants
: external stimulus
There are n system variables, m system parameters and 1 small variable to allow near perfect adaptation, giving a total of (n+m+1)H equations and (n+m+1)H variables.
Discretizing sinto H points
0
||
0);;(
ds
kdds
du
skuFdt
ud
N
02
|2
|
0);;(
)1(
11
11
s
kks
uu
skuFdt
ud
sjss
jm
jm
j
jN
jN
jjj
jlowj
Augmented system
The steady state concentration of proteins in the network must satisfy:
The steady state concentration of CheYp must satisfy:
At the same time, the reaction rate constants must be independent of stimulus:0
ds
kd
0);;( skuFdt
ud
N
N
u
ds
du
||
: allows for near-perfect adaptation
= CheYp
Some examples of the parameter space giving perfect adaptation
Obtained for sslow< s < shigh
Verification for different stimuli s > shigh
05.0
Work in progress Complete construction of manifolds in parameter space, allowing insight into parameter dependence giving rise to robustness. Extend to other signaling systems, such as phototransduction.
0);;( skuFdt
ud
Implementation• Newton-Raphson, to solve for the steady state of augmented system
Simplest multidimensional root finding method Efficient way of converging to a root with a sufficiently good initial guess.
f(x) 1
x2
works well unfortunate case unfortunate case
f(x)1
32
x
f(x) 12
x
• Dsode (stiff ODE solver), to verify Newton-Raphson result for different ranges of external stimulus
Single result from fine-tuned model
Ref: P. A. Spiro, J. S. Parkinson, and H. G. Othmer, “A model of excitation and adaptation in bacterial chemotaxis”, Proc. Natl. Acad. Sci. USA 94, 7263(1997)
6 7
Demethylation rate (km1,km2) and methylation rate (k1c,k2c) versus phosphorylation rate(k9)
Demethylation rate (km1,km2) and methylation rate (k1c,k2c) versus dephosphorylation rate(kmy)
Demethylation rate (km1,km2) and methylation rate (k1c,k2c) versus dephosphorylation rate(kmb)