Embed Size (px)
DESCRIPTION
1%. : state variables : reaction kinetics : reaction constants : external stimulus. The steady state concentration of proteins in the network must satisfy: The steady state concentration of CheYp must satisfy: At the same time, the reaction rate constants must be independent of stimulus:. - PowerPoint PPT Presentation
Citation preview

Use Newton-Raphson (root finding algorithm with back-tracking), to solve for the steady state of augmented system,
Use Dsode (stiff ODE solver), to verify time- dependent behavior for different ranges of external stimulus by solving:
Robust Perfect Adaptation in Bacterial Chemotaxis Yang Yang & Sima Setayeshgar Department of Physics, Indiana University, Bloomington
MotivationThe biochemical basis of robustness of perfect adaptation is not as yet fully understood. In this work, we develop a novel method for elucidating regions in parameter space of which the E. coli chemotaxis network adapts perfectly:
Broader impactThis method should have applicability to other cellular signal transduction networks and engineered systems that exhibit robust homeostasis.
The shapes of resulting manifolds determine relationships between reaction parameters (for example, methylation and phosphorylation rates) that must be satisfied in order for the network to exhibit perfect adaptation, thereby shedding light on biochemical steps and feedback mechanisms underlying robustness.
Given lack of complete data on values of in vivo reaction rates, the numerical ranges of the resulting manifolds will shed light on values of unknown or partially known parameters.
ConclusionsI.Successful implementation of a novel method for elucidating regions in parameter space allowing precise adaptation II.Numerical results for (near-) perfect adaptation manifolds in parameter space for the E. coli chemotaxis network, allowing determination of
i. conditions required for perfect adaptation, consistent with and extending previous works [1-3]
ii. numerical ranges for unknown or partially known kinetic parametersIII.Extension to modified chemotaxis networks, for example with no CheZ homologue and multiple CheYs
Work in progress Extension to other signaling networks:
• vertebrate phototransduction • mammalian circadian clock
allowing determination of• parameter dependences underlying robustness• plausible numerical values for unknown network parameters
0|2
|
|2
|
0);;(
)1(
11
11
s
kk
s
uu
skuFdt
ud
siss
iN
iN
i
iN
iN
iii
ilowi
AlgorithmSTART with a fine-tuned model of chemotaxis network that:
reproduces key features of experiments (adaptation times to small and large ramps, perfect adaptation of the steady state value of CheYp)
is NOT robustAUGMENT the model explicitly with the requirements that: steady state value of CheYp
values of reaction rate constants, are independent of the external stimulus, s, thereby achieving robustness of perfect adaptation.
s
k
F
u
skuFdt
ud
0);;(
: state variables
: reaction kinetics
: reaction constants
: external stimulus
The steady state concentration of proteins in the network must satisfy:
The steady state concentration of CheYp must satisfy:
At the same time, the reaction rate constants must be independent of stimulus: 0
ds
kd
0);;( skuFdt
ud
N
N
u
ds
du
||
: allows for near-perfect adaptation
= CheYp
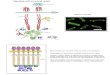
E. coli exhibits an important behavioral response known as chemotaxis - motion toward desirable chemicals (usually nutrients) and away from harmful ones - which is also shared by various other prokaryotic and eukaryotic cells. The cell’s motion consists of series of “runs” punctuated by “ tumbles”.
E .coli
It is considered to be an ideal model organism for understanding the behavior of cells at the molecular level from the perspectives of several scientific disciplines-anatomy, genetics, chemistry and physics since :• Ease of experimentation, through microscopy and genetic analysis• Small genome (4288 genes), most of which encode proteins
2
Importantance of perfect adaptation
Chemotaxis signal transduction network in E. coli
Importance of robustnessThe E. coli chemotaxis signal transduction network exhibits robust perfect adaptation, where the concentration of CheYp returns to its prestimulus value despite large changes in the values of many of the biochemical reaction rate constants. These rate constants depend on concentrations of enzymes, which are often present in small copy numbers, making fluctuations in their numbers significant.
Chemotaxis in E. coli involves temporal measurement of the change in concentration of an external stimulus. This is achieved through the existence of fast and slow reaction time scales, in the chemotaxis signal transduction network: fast measurement of the current external concentration is compared with the cell’s “memory” of the concentration some time ago to determine whether to extend a run in a given direction or to tumble, thereby randomly selecting a new direction.
E. coli is a single-celled organism that lives primary in our intestines. It is approximately 1-2 microns long and 1 micron in diameter, and weights 1 picogram. Each cell has 4-6 flagella, approximately 10-20 microns long, driven by an intracellular rotary motor operated by the protonmotive force.
The chemotaxis signal transduction pathway in E. coli – a network of ~50 interacting proteins – converts an external stimulus (change in concentration of chemoattractant / repellent) into an internal stimulus (change in concentration of intracellular response regulator, CheYp) which in turn interacts with the flagella motor to bias the cell’s motion.
It is used as a well-characterized model system for the study of properties of (two-component) cellular signaling networks in general.
Perfect adaptation is an important and generic property of signaling systems, where the response (e.g. running bias in chemotaxis) returns precisely to the pre-stimulus level while the stimulus persists. This property allows the system to compensate for the presence of continued stimulation and to be ready to respond to further stimuli.Thus, E. coli is able to respond to changes in chemoattractant concentrations spanning 5 orders of magnitude! Similarly, the vertebrate visual system responds to changes in light intensity spanning 10 orders of magnitude during the night-day cycle.
Ref: H. C. Berg, “Motile behavior of bacterial”, Physics Today, January 2000
Ref: P. A. Spiro, J. S. Parkinson, and H. G. Othmer, “A model of excitation and adaptation in bacterial chemotaxis”, Proc. Natl. Acad. Sci. USA 94, 7263(1997)
Ref: N.Barkai & S. Leibler, “Robustness in simple biochemical network”, Nature 387, 913(1997)
2
Chemical reactions: Ligand binding
Methylation
Phosphorylation
)()( )(7/7~5/5
)( CheRLTCheRTL pnkmkkmk
pn
ppnmkmk
ppn
pnckck
pn
CheBTLCheBTL
CheRTLCheRTL
)(14~1
)(
)(14~1
)(
)()(
)()(
PCheBCheB
PCheYCheZCheZCheY
BCheYCheRTBCheYCheRTL
ADPCheRTLATPCheRTL
kmbp
kmyp
pnbky
np
npkk
n
)()()()()(
)()()()()(
9~7
There are n system variables, m system parameters and 1 small variable to allow near perfect adaptation, giving a total of (n+m+1)H equations and (n+m+1)H variables.
Discretizing s
into H points
Measurement of c = [CheY-P] by flagellar motor constrained by diffusive noise Relative accuracy*,
Signaling pathway required to adapt “nearly” perfectly, to within this lower bound
(*) Berg & Purcell, Biophys. J. (1977).
%101
~
cDac
c
: diffusion constant (~ 3 µM): linear dimension of motor C-ring (~ 45 nm): CheY-P concentration (at steady state ~ 3 µM): measurement time (run duration ~ 1 second)
c
a
D
0
||
0);(
ds
kdds
dysyF
N
},,{ kuy
);;( skuFdt
ud
T4
au
top
hosp
hory
lati
on
rate
(k
10)
LT4 a
uto
ph
osp
hory
lati
on
rate
(k
10)
● 3%<<5% ● 1%<<3% ● 0%<<1%
Parameter Surfaces
Time (s)
Con
cen
trati
on
(µ
M)
Verify steady state NR solutions dynamically using DSODE for different stimulus ramps:
•{k3c= 5 s-1, k10 = 36 s-1, km2 = 3e+4 M-1s-1}
•{k3c= 5 s-1, k10 = 101 s-1, km2 = 6.3e+4 M-1s-1}
Validation
(1,12.7)
Violating and Restoring Perfect Adaptation
1%
k1c : 0.17 s-1 1 s-1
k8 : 15 s-1 12.7 s-1
Step stimulus from 0 to 1e-6M at t=250s
(1,15)
(1,12.7)
T2 Methylation rate (k1c)
T2 a
uto
ph
osp
hory
lati
on
rate
(k
8)
Conditions for Perfect Adaptation
T3 autophosphorylation rate
T3 d
em
eth
yla
tion r
ate
/ T2
meth
yla
tion
rate
T4 autophosphorylation rate
T4 d
em
eth
yla
tion r
ate
/ T3
meth
yla
tion
rate
LT3 autophosphorylation rate
T3 d
em
eth
yla
tion r
ate
/ T2
meth
yla
tion
rate
LT4 autophosphorylation rate
LT4 d
em
eth
yla
tion r
ate
/ LT
3
meth
yla
tion
rate
CheB
phosp
hory
lati
on
rate
(k
b)
/ lit
era
ture
valu
e
CheY p
hosp
hory
lati
on
rate
(k
y)
/ lit
era
ture
valu
e
(L)Tn autophosphorylation rate / literature value
(L)Tn autophosphorylation rate / literature value
● T2● T3● T4● LT3● LT4
● T2● T3● T4● LT3● LT4
Ch
eB
ph
osp
hory
lati
on
rate
LT2 autophosphorylation rate
Ch
eY p
hosp
hory
lati
on
ra
te
LT2 autophosphorylation rate
T3
dem
eth
yla
tion r
ate
(k
m1)
T3 autophosphorylation rate (k9)
T4 autophosphorylation rate (k10)
T4
dem
eth
yla
tion r
ate
(k
m2)
LT3 autophosphorylation rate (k12)
LT3
dem
eth
yla
tion r
ate
(k
m3)
LT4 autophosphorylation rate (k13)
LT4
dem
eth
yla
tion r
ate
(k
m4)
Demethylation Rate is proportional to Autophosphorylation Rate2
T2 autophosphorylation rate (k8)
T2
Meth
yla
tion r
ate
(k
1c)
T3 autophosphorylation rate (k9)
T3
Meth
yla
tion r
ate
(k 2
c)Methylation Rate is proportional to Autophosphorylation Rate
LT2 autophosphorylation rate (k12)
LT2
Meth
yla
tion r
ate
(k
3c)
LT3 autophosphorylation rate (k13)
LT3
Meth
yla
tion r
ate
(k
4c)
Demethylation Rate/Methylation Rate is proportional to Autophosphorylation Rate
CheB, CheY Phosphorylation Rate is proportional to Autophosphorylation Rate
Condition for Robust Perfect Adaptation
By varying 3 parameters(Ttot, k11, k3c) in the code to find a region where Ttot can vary a lot while the
others remain constant.
L=0(solid)L=1µM(dashed)L=1mM(dashed dot)
Spiro model.
Barkai–Leibler model
L=0(solid)L=1µM(dashed)L=1mM(dashed dot)
Ref: P. A. Spiro, J. S. Parkinson, and H. G. Othmer, “A model of excitation and adaptation in bacterial chemotaxis”, Proc. Natl. Acad. Sci. USA 94, 7263(1997)T.M.Yi, Y. Huang, M. I. Simon, and J. Doyle, “Robust perfect adaptation in bacterial chemotaxisthrough integral feedback control”,PNAS, 97, 4649 (2000)
Verifying the conditions for perfect adaptation of two-state model
Diversity of Chemotaxis Systems
Eg., Rhodobacter sphaeroides, Caulobacter crescentus and several rhizobacteria possess multiple CheYs while lacking of CheZ homologue.
In different bacteria, additional protein components as well as multiple copies of certain chemotaxis proteins are present.
Response regulatorCheY1
CheY2 Phosphate “sink”
Requiring: Faster phosphorylation/autodephosphorylation rates of CheY than CheY1
Faster phosphorylation rate of CheB
Ch
eY
1p
(µM
)
Time(s)
Exact adaptation in modified chemotaxis network with CheY1, CheY2 and no CheZ:
Inactive active
1. Fast ligand (un-)binding reaction
2. Only acitive receptor can bind to CheB
3. Only inactive receptor can bind to CheR
4. Autophosphorylation rates of receptors
are proportional to the activity
Slope=0.18
Slope=0.15
T2
dem
eth
yla
tion c
ata
lyti
c ra
te
T1 methylation catalytic rate
T1
dem
eth
yla
tion c
ata
lyti
c ra
te
T0 methylation catalytic rate
T3
dem
eth
yla
tion c
ata
lyti
c ra
te
T2 methylation catalytic rate
T4
dem
eth
yla
tion c
ata
lyti
c ra
te
T3 methylation catalytic rate
5. Phosphorylation transferrate form CheA to CheY and CheB are porportional to
the activity
6. The ratios between the CheR catalytic rate and CheB-p catalytic rate of the next
methylation level are the same for all methylation states.Verifying condition 6: reference value kb/kr=0.155/0.819=0.19
Ref: B. Mello, Y. Tu,”Perfect and near-perfect adaptation in a model of bacterial chemotaxis”, Biophysical Journal, 2003
Slope=0.15 Slope=0.18 Slope=0.15 Slope=0.19