Embed Size (px)
Citation preview

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
Annu. Rev. Neurosci. 2001. 24:869–96Copyright c© 2001 by Annual Reviews. All rights reserved
EARLY ANTERIOR/POSTERIOR PATTERNING
OF THE MIDBRAIN AND CEREBELLUM
Aimin Liu1 and Alexandra L Joyner1,2
Howard Hughes Medical Institute and Developmental Genetics Program, SkirballInstitute of Biomolecular Medicine, Departments of1Cell Biology and2Physiology &Neuroscience, New York University School of Medicine, New York, NY10016;e-mail: [email protected], [email protected]
Key Words midbrain/hindbrain organizer, Fibroblast growth factor,Otx2, Gbx2,Engrailed
■ Abstract Transplantation studies performed in chicken embryos indicated thatearly anterior/posterior patterning of the vertebrate midbrain and cerebellum might beregulated by an organizing center at the junction between the midbrain and hindbrain.More than a decade of molecular and genetic studies have shown that such an organizeris indeed central to development of the midbrain and anterior hindbrain. Furthermore, acomplicated molecular network that includes multiple positive and negative feedbackloops underlies the establishment and refinement of a mid/hindbrain organizer, aswell as the subsequent function of the organizer. In this review, we first introducethe expression patterns of the genes known to be involved in this patterning processand the quail-chick transplantation experiments that have provided the foundation forunderstanding the genetic pathways regulating mid/hindbrain patterning. Subsequently,we discuss the molecular genetic studies that have revealed the roles for many genesin normal early patterning of this region. Finally, some of the remaining questions andfuture directions are discussed.
INTRODUCTION
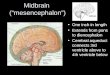
Regionalization of the central nervous system (CNS) is a critical early event invertebrate neural development. By embryonic day 9.5 (E9.5), the main regionsof mouse CNS can be clearly distinguished morphologically along the ante-rior/posterior (A/P) axis (Figure 1). The forebrain consists of the telencephalonsituated at the most rostral end of the neural tube and the more posterior dien-cephalon, which abuts the midbrain (mesencephalon). The forebrain is dividedinto several prosomeres, with prosomere 1 being the most caudal. The hindbrainis separated from the midbrain by a constriction that is called the mid/hindbrainjunction or isthmus and is divided into the rostral metencephalon and the caudalmyelencephalon. The hindbrain, also called rhombencephalon, is further divided
0147-006X/01/0621-0869$14.00 869

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
870 LIU ¥ JOYNER
into eight rhombomeres. The metencephalon consists of rhombomere 1 (r1), whichgives rise to the cerebellum and pons (Millet et al 1996, Wingate & Hatten 1999),and r2. In this review we refer to the region that gives rise to the midbrain andcerebellum and that includes the mesencephalon and anterior metencephalon (r1)as the mes/met region.
Quail-chick transplantation experiments showed that development of themes/met region could be regulated by a group of organizer cells located at themid/hindbrain junction (for review, see Alvarado-Mallart 1993). Subsequently,several secreted growth factor genes were found to be expressed in the isthmus,and a number of genes encoding transcription factors were found to be expressedmore broadly in the mes/met (Wassef & Joyner 1997, Joyner et al 2000) (Figure 1).Gain- and loss-of-function studies over the past 10 years have demonstrated thatthese genes are indeed critical regulators of vertebrate mes/met development. Sig-nificantly, Fibroblast growth factor-8 (FGF8), which is expressed in the isthmus,is both necessary for mes/met development and can induce ectopic midbrain orcerebellum development. Furthermore, the junction between the expression do-mains of two transcription factors,Otx2 and Gbx2, defines the position of thecells with organizer function. Finally, a complicated set of genetic interactionsinvolving these and other growth factors and transcription factors regulate bothestablishment of the mes/met and subsequent A/P patterning of the midbrain andcerebellum.
EXPRESSION PATTERNS OF MANY MES/MET GENESARE CONSISTENT WITH THE MID/HINDBRAINJUNCTION CONTAINING AN ORGANIZER
An organizer, such as the well-known Spemann’s organizer, is a group of cellsthat produce signals that can “organize” neighboring cells into well-patterned andfunctional structures (for a review, see Nieto 1999). One important characteristicshared by all known organizers is that they can direct competent tissues to takeon new fates that are normally taken only by their native neighbors. Meinhardt(1983) studied the formation of several well-known organizers and found that theywere all formed at points of apposition of two or more differentially specifiedtissues. Therefore, he proposed a model in which determinants from two (or more)adjoining compartments acting on the same border cells lead to the induction ofexpression in the border cells of a patterning signal(s), which in turn, function(s)to sharpen the border and guide the patterning of the adjacent compartments.
The expression patterns of a number of mes/met genes during early developmentare consistent with patterning of the region being regulated in a manner similarto that predicted by the model proposed by Meinhardt (1983). Two differentiallyspecified regions of the neural plate, anteriorly locatedOtx2-expressing tissue andposteriorly locatedGbx2-expressing tissue, are found to confront each other in

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 871
mice as early as E7.5 (Wassarman et al 1997). Expression of another transcriptionfactor genePax2is then initiated at the presomite stage (E7.5) surrounding the siteof confrontation, followed by similar expression of several transcription factors(Figure 1, see color insert) (for a review, see Wassef & Joyner 1997, Joyner et al2000). Most interesting, the expression patterns of the two secreted molecules,FGF8 and WNT1, become highly restricted to adjacent narrow transverse bands atthe Otx2/Gbx2border (Wilkinson et al 1987, Crossley et al 1995). A relativelyfuzzy boundary betweenGbx2andOtx2expression domains also becomes sharpat the 4- to 6-somite stage, and this boundary colocalizes with the mid/hindbrainjunction that in mice is morphologically clear by E9.5. Genes such asEn1,En2, andPax5continue to be expressed in both the mesencephalon and the metencephalonafter E9.5 (Figure 1). This scenario of gene expression correlates well with the“organizer model” proposed by Meinhardt (1983). Indeed, numerous transplanta-tion experiments with chicks have shown that the mid/hindbrain junction has anorganizing activity (for a review, see Alvarado-Mallart 1993, Joyner et al 2000).We review in more detail the expression patterns of various mes/met genes mainlyin mice in this section and then discuss the experiments that revealed the organizingfunction of the isthmus.
Otx2 and Gbx2
The transcription factors OTX1 and -2 are homologues of the Drosophila orth-odenticle protein (Simeone et al 1992). Prior to gastrulation,Otx2 is expressedthroughout the epiblast and in the anterior visceral endoderm (Simeone et al 1992,1993). As the primitive streak forms,Otx2expression becomes progressively re-stricted to an anterior region of the mouse embryo in all three germ layers andin the visceral endoderm.Otx1 expression begins at the 1- to 3-somite stage inthe anterior neuroectoderm. Subsequently, the caudal boundaries ofOtx1and -2expression are located in the isthmic constriction in mouse embryos. Fate map-ping experiments with chicks have provided evidence that the caudal limit ofOtx2expression as early as Hamburger & Hamilton (HH) stage 10 (10 somites)(Hamburger & Hamilton 1992) marks the boundary between the midbrain andhindbrain (Millet et al 1996).
At E7.5, the homeobox geneGbx2is expressed in all three germ layers in theposterior region of a mouse embryo (Bouillet et al 1995, Wassarman et al 1997).Subsequently, its expression in the CNS becomes restricted to the anterior hind-brain by E8.5. At E9.5,Gbx2expression is restricted to the anterior metencephalonand four longitudinal lateral stripes through the entire length of the hindbrain andspinal cord. Starting at E10.0,Gbx2expression in the CNS is also found in pre-cursors of the thalamus in the dorsal diencephalon.
Fgf8, Fgf17, and Fgf18
Fgf8 is expressed in many locations of a developing mouse embryo at differentstages, which suggests that it has multiple roles in regulating cell proliferation and

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
872 LIU ¥ JOYNER
differentiation in diverse processes (Heikinheimo et al 1994, Crossley & Martin1995). In mouse mes/met development,Fgf8 is initially expressed in a broadregion in the prospective anterior hindbrain starting at the 3-somite stage. Thisexpression quickly becomes more intense and more restricted to a narrow ring justcaudal to the mid/hindbrain junction and persists until E12.5.
More recently, it has been reported that two additional mouseFgf genes,Fgf17and Fgf18, which share very high sequence homology withFgf8 (63.7% and56.8%, respectively, at the amino acid level), are also expressed in the anteriorhindbrain region at E8.5 (Maruoka et al 1998, Xu et al 1999). However, it seemsthat their expression levels at early somite stages are weaker than that ofFgf8.Furthermore,Fgf17expression is broader than that ofFgf8, it occurs in both themesencephalon and the metencephalon, and it persists until at least E14.5, a stagewhenFgf8 is no longer expressed. The fact thatFgf8 expression seems strongerat early somite stages and the fact of the severe mes/met phenotype ofFgf8 loss-of-function mutants (see below) suggest that FGF8 is the major FGF molecule thatis essential for early patterning of this region.
Wnt1, Lmx1b, En1/2, and Pax2/5
Wnt1is a homologue of the Drosophila segmentation genewinglessand encodesa secreted signaling molecule (for a review, see Nusse & Varmus 1992).En1andEn2are homologues of the Drosophila segmentation geneengrailedand encodehomeodomain-containing transcription factors (Joyner & Martin 1987).Pax2andPax5 are the homologues of the Drosophila pair-rule genepaired and encodepaired-domain-containing transcription factors (Gruss & Walther 1992).Lmx1bisa gene that encodes a LIM homeodomain-containing transcription factor.Lmx1bexpression has been described in detail only from chick brain (Yuan & Schoenwolf1999, Adams et al 2000). It is first expressed broadly in the caudal forebrain,midbrain, and hindbrain. By HH stage 10, the expression is restricted to the dorsaland ventral midline of the midbrain, overlapping withWnt1expression.Lmx1bisalso expressed in the isthmus at this stage, and its expression overlaps with boththeWnt1andFgf8domains.
Pax2expression in mice begins at the presomite stage in cells surrounding theOtx2/Gbx2boundary (Rowitch & McMahon 1995).Wnt1andEn1expression theninitiates within thePax2domain at the 1-somite stage, withWnt1expression re-stricted toOtx2-positive midbrain cells, andEn1expression in the entire mes/met(Davis & Joyner 1988, Rowitch & McMahon 1995, Liu & Joyner 2001).En2andPax5expression initiates around the 5-somite stage across theOtx2/Gbx2bound-ary (Davis & Joyner 1988, Adams et al 1992, Song et al 1996). The expressionpatterns of all these genes are dynamic in the mes/met region. For example,Wnt1is expressed in the entire mesencephalon at early somite stages and soon becomesrestricted to a narrow ring rostral to the mid/hindbrain junction, as well as beingexpressed along the dorsal midline of the midbrain and caudal diencephalon, cau-dal hindbrain, and spinal cord. Transient expression ofWnt1in the ventral midline

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 873
of the midbrain and diencephalon is also seen at E9.5–E10.5.Pax2expressionalso becomes restricted to the isthmus by E9, just caudal to theWnt1expressiondomain. In contrast, the expression ofEn1, En2, andPax5remains relatively broadon both sides of the mid/hindbrain junction (Figure 1)
TRANSPLANTATION STUDIES IN CHICKS REVEALTHAT THE ISTHMUS HAS ORGANIZER ACTIVITY
The Isthmus Serves as an Organizing Center
Many transplantation experiments between quail and chicken embryos at the 11-to 14-somite stage have shown that a mid/hindbrain organizer is present at themid/hindbrain junction (for a review, see Alvarado-Mallart 1993, Wassef & Joyner1997). These experiments can be summarized into two groups. The first group isa series of inversions. When quail rostral mesencephalic tissue is inverted andtransplanted into the corresponding region of a chick host, the transplant adjustsits polarity according to the new environment. However, if the transplanted tissueincludes tissue from the caudal part of the mesencephalic vesicle [precursors ofisthmic nuclei and cerebellum (Millet et al 1996)], it not only maintains its ownpolarity in most cases, it also can induce the adjacent, more-anterior host dien-cephalic tissue to form caudal mesencephalic structures (Figure 2A). In the secondgroup of transplantation experiments, isthmus tissue is transplanted into either thediencephalic or myelencephalic region. In the diencephalic region, the transplantsinduce caudal mesencephalic development, whereas in the hindbrain they can in-duce ectopic cerebellar structures (Figure 2B). The reverse has also been done, andin these cases diencephalic or hindbrain tissue transplanted into the region of theisthmus is transformed into mes/met tissue.
In all the experiments where ectopic mes/met structures were induced by isthmictissue, expression ofEn2 was shown to be induced before the ectopic mes/metstructures developed. The expression ofWnt1was also altered in the diencephalon,such that it extended from the dorsal midline where endogenousWnt1expressionresides to the vicinity of the graft. Based on these results, it was suggested thatEn2andWnt1could be involved in the ectopic development of the midbrain andcerebellar tissues (for a review, see Wassef & Joyner 1997).
The Isthmic Organizer Can be Generated by the Appositionof Midbrain and Anterior Hindbrain Tissue
Consistent with the general proposal made by Meinhardt (1983), Irving & Mason(1999) have shown both in vitro and in vivo that when chick midbrain tissue isplaced adjacent to posterior r1 tissue, expression ofFgf8 is induced at the newboundary between the tissues (Figure 2C). EctopicWnt1expression and alteredtectal polarity was also observed after grafting posterior r1 tissue into the midbrain.

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
Figure 2 The isthmic region can act like a midbrain/cerebellum organizer. (A) When theentire mesencephalic vesicle is inverted in the chick at HH stage 10, a bicaudal midbrainis formed, and the adjacent caudal forebrain is transformed into an ectopic midbrain. (B)Transplantation of isthmic tissue can induce ectopic expression of mes/met genes, suchasEn2 and ectopic mes/met development. The strongest endogenousEn2 expression isin the mid/hindbrain junction, and it decreased on both sides of the junction. (C) Theapposition of caudal rhombomere 1 (r1) tissue and midbrain or caudal forebrain tissue leadsto induction ofFgf8expression. T, telencephalon; D, diencephalon; Mes, mesencephalon;Met, metencephalon; My, myelencephalon.

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 875
These results suggest that formation of an isthmic organizer might result from aninteraction between midbrain and anterior hindbrain tissues. Furthermore,Otx2andGbx2might be involved in this process, given the correlation between theisthmic organizer and theOtx2/Gbx2border. Nevertheless, Irving & Mason (1999)suggested that there must be other genes involved inFgf8 induction because whenr2 tissue, which also expressesGbx2, is placed adjacent to midbrain tissue, noFgf8 is induced at the border. However, compared with expression in r1,Gbx2expression in r2 appears weaker and restricted dorsally and, thus, may not be strongenough to induceFgf8expression.
Hidalgo-Sanchez et al (1999) also addressed whether juxtaposition ofOtx2-positive andGbx2-positive cells could induce an organizer using transplants. Theyshowed that whenOtx2-positive diencephalic tissue is transplanted intoGbx2-positive r1,Gbx2is induced andOtx2 is repressed in the grafted cells that contacttheGbx2-positive host r1 tissue.Fgf8, En2, Wnt1, andPax2are also induced inthe grafts at the newGbx2/Otx2border. The only complication with these studies,as mentioned by the authors, is that transient contact between the graft andFgf8-expressing host cells in the isthmus is hard to avoid. Therefore, it is difficult todistinguish whether the mes/met gene expression that is induced in grafted cellsis solely due to the initial confrontation betweenGbx2- andOtx2-expressing cellsand/or to the inductive function ofFgf8 in the host.
FIBROBLAST GROWTH FACTORS ARE CRITICALFOR MIDBRAIN AND CEREBELLAR DEVELOPMENT
General Properties of Fibroblast Growth Factorsand Their Receptors
The mouse fibroblast growth factor (FGF) family currently consists of at least19 members, named FGF1–19 (for a review, see Basilico & Moscatelli 1992,Coulier et al 1997). Most FGFs are secreted proteins and are expressed not only intransformed tumor cells but also in many signaling centers during normal vertebrateembryonic development. It has been shown in Xenopus that FGFs are involved ininducing mesoderm formation and posterior neural fate (for a review, see Slacket al 1996). The importance of theFgf genes in mouse development has beendemonstrated by the analysis of null mutants for severalFgf genes (Mansour et al1993, Hebert et al 1994, Feldman et al 1995, Min et al 1998, Sekine et al 1999,Sun et al 1999).
The best-characterized FGF receptors are high-affinity, signal-transducing tyro-sine kinase coupled receptors, which include at least four members, called FGFR1–4. Many FGFR isoforms are generated by alternative splicing of the mRNAs. Thedimerization of the FGFRs on ligand binding triggers the activation of the recep-tors and downstream signaling events. Each kind of FGF molecule is a preferentialligand for a specific FGFR, and each kind of FGFR can bind several different FGF

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
876 LIU ¥ JOYNER
molecules (Basilico & Moscatelli 1992). It has been suggested that heparan-sulfateproteoglycans may help to stabilize the FGF-FGFR complex, and structural anal-ysis has provided evidence for such a function (Plotnikov et al 1999).
Comparative expression studies have been done in medaka fish and in chicks toinvestigate the expression patterns of differentFgfr genes as a means of gaininginformation as to which gene(s) could be involved in mes/met development. Inmedaka fish,Fgfr2, 3, and4 are expressed in the midbrain and anterior hindbrain;however,Fgfr2 is the only gene that is expressed in cells adjacent to, and on bothsides of, the mid/hindbrain junction at early somite stages (Carl & Wittbrodt 1999).In contrast, chickFgfr1, 2, and3 are all expressed throughout the presumptiveneural plate during early neural induction (∼HH stage 5) and elevated in theanterior neural plate before neuromeres form (∼HH stage 8) (Walshe & Mason2000). However, when neuromeres form (HH stage 9–11), onlyFgfr1 is expressedin the mes/met region. Detailed comparative study of the expression patterns ofFgfrs in mouse brain at early stages has not been reported, although it has beenshown thatFgfr1 and -2, but not -4, are expressed in embryonic brain (Stark et al1991; Yamaguchi et al 1992; Orr-Urtreger et al 1993; Peters et al 1992, 1993).Our preliminary studies, however, show that in miceFgfr1 is weakly expressedthroughout the CNS andFgfr2 and -3 are largely excluded from the posteriormidbrain and r1 at E8.5–E9.5 (A Liu, AL Joyner, unpublished data). Taken toge-ther, the expression patterns of the FGF receptors are dynamic, and at least FGFR1and 2 could be involved in mes/met development. All fourFgfr genes have beenmutated in mice by gene targeting, but mes/met patterning defects have not beenreported for anyFgfr mutant, either in mutant embryos or in chimeras containingmutant cells in the mes/met. It is possible that the FGFRs have overlapping func-tions (Deng et al 1994, 1996; Yamaguchi et al 1994; Colvin et al 1996; Cirunaet al 1997; Arman et al 1998, 1999; Weinstein et al 1998; Xu et al 1998).
Fgf8 was first identified as an oncogene responsible for androgen-dependentgrowth of mammary gland carcinoma cells and was initially called androgen-induced growth factor (Tanaka et al 1992). FGF8 has seven isoforms (Crossley &Martin 1995, MacArthur et al 1995b), and biochemical and cell transformationassays have demonstrated that most of the FGF8 isoforms activate the c isoformsof FGFR2, -3, and -4 and can transform NIH3T3 cells in culture. Among theFGF8 isoforms, FGF8b has the strongest affinity for the three receptors and hasthe strongest ability to transform NIH3T3 cells, whereas FGF8a shows little affinityto the receptors and very weak transforming activity (MacArthur et al 1995a,b;Blunt et al 1997).
Fgf8 is Required for Normal Developmentof the Vertebrate Mes/Met Region
A series of mouseFgf8 mutant alleles have been generated by gene targeting inES cells (Meyers et al 1998). A null mutation in which the second and third exonsare deleted [Fgf812, 3/12, 3] leads to a gastrulation defect (Meyers et al 1998, Sunet al 1999). Mice homozygous for a hypomorphic allele,Fgf8neo, which contains

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 877
an insertion of aneocassette into the first intron, or trans-heterozygous for thehypomorphic and null alleles, lack most of the midbrain and cerebellum, indicatingFgf8 is required for mes/met development (Meyers et al 1998). However, the lossof mes/met tissue either could be due to theFgf8expression in the metencephalonor could be secondary to an early mild gastrulation defect. This will be addressed byanalyzing conditional mutants in which theFgf8gene is mutated only in mes/metprecursor cells.
Zebrafishace(acerebellar) mutants lack the isthmus and cerebellum and havea point mutation in a zebrafish homologue ofFgf8 that leads to production of atruncated form of protein (Brand et al 1996, Reifers et al 1998). The midbrain inacemutants is expanded caudally and has impaired polarity along both the A/P anddorsal/ventral axes (Picker et al 1999). Analysis of marker gene expression showedthat mes/met expression ofeng, wnt1, andpax2.1was initiated properly at the endof gastrulation but was then progressively lost. The apparent milder phenotypein acemutants compared with that ofFgf8 hypomorphic mouse mutants couldbe because either theacemutation is a hypomorphicFgf8 allele or anotherFgffamily member can partially compensate for the loss ofFgf8 in fish.
FGF8b-Soaked Beads Can Induce Mes/Met Genes and EctopicMidbrain and Cerebellar Structures in Regions of Chick Brain
A striking finding is that FGF8b-soaked beads inserted into either the posteriordiencephalon (prosomere 1 and 2) or anterior mesencephalon of stage 9–12 chickenembryos induceEn1, En2, Wnt1, andPax2expression within 24–48 h (Crossleyet al 1996, Shamim et al 1999). An interesting recent finding is thatSprouty2,which encodes a putative FGF antagonist, is also induced by FGF8-soaked beads,but more rapidly and within an hour (Minowada et al 1999, Chambers et al 2000).Normally,Sprouty1and -2 are expressed around the isthmus, and thus they couldnormally play a role in modulating FGF signaling from r1 (Minowada et al 1999,Chambers et al 2000, Chambers & Mason 2000).
The long-term consequence of insertion of FGF8-soaked beads into certainbrain regions is induction of ectopic mes/met-derived structures. Both the rostralmesencephalon and caudal diencephalon can be transformed into caudal mesen-cephalic structures, with an ectopic isthmus close to the FGF8 bead (Figure 3A).FGF8b-soaked beads placed in prosomere 1 or the anterior midbrain were foundto induceGbx2expression and repressOtx2 in cells around the beads (Irving &Mason 1999, Martinez et al 1999). Furthermore, in some cases theOtx2-negativecells in contact with the beads can form an outgrowth that protrudes from theneural tube (Martinez et al 1999). The outgrowth later develops proximally intoisthmic nuclei and distally into cerebellum-like structures, which suggests thatone of the normal functions for FGF8 might be to repressOtx2expression in theanterior hindbrain and allow, or induce, cerebellum development (Figure 3A). Re-cently, it was shown that although FGF8-soaked beads can induceEn2andPax2in r2, it does not lead to formation of ectopic structures (Irving & Mason 2000)(Figure 3A).

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
878 LIU ¥ JOYNER
Figure 3 FGF8 can induce ectopic mid/hindbrain development in chicks and mice. Inthe diencephalon and midbrain of both chicken embryos and mouse brain explants, FGF8-soaked beads can induce mid/hindbrain gene expression and, in chicks, ectopic developmentof midbrain and cerebellar structures. FGF8-soaked beads inserted into the anterior forebraindo not induce mes/met gene expression or mes/met structures. In the anterior hindbrain,FGF8 can induce the expression ofEn2, Pax2, andWnt1 but no induction of mes/metstructures. p, prosomere; r, rhombomere.
It is interesting that Shamim et al (1999) found that both FGF4- and FGF8-soaked beads when implanted into the midbrain can induceFgf8expression, withFGF4 inducing expression in a broader region. In contrast to what was describedby Crossley et al (1996), endogenousFgf8was not found to be induced by FGF4or -8 in the posterior forebrain. However, weak expression ofFgf8 was inducedin the anterior midbrain when FGF4-, but not FGF8-, soaked beads were insertedinto the caudal forebrain. Taken together, these chick studies using FGF-soakedbeads show that FGF8 is sufficient to induce midbrain, isthmic, and cerebellarstructures. However, not all brain regions are equally competent to respond toFGF8.
Wnt1-Fgf8b Transgenic Mouse Embryos Show an EarlyTransformation of the Midbrain and Posterior Forebraininto an Anterior Hindbrain Fate
To study the role ofFgf8in mouse mes/met patterning, transgenic embryos (Wnt1-Fgf8) have been produced in whichFgf8 is expressed under the control of aWnt1regulatory element such thatFgf8 expression is extended into the entire mesen-cephalon at early somite stages and along the dorsal midline into the caudal dien-cephalon after E9.0 (Lee et al 1997, Liu et al 1999).Fgf8b-expressing embryosshow severe exencephaly and die shortly after E15.5 (Liu et al 1999). Detailed

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 879
marker gene expression studies at E9.5 indicate that the midbrain and posteriorforebrain do not form in such embryos, and the hindbrain is found adjacent to theremaining anterior forebrain. This phenotype appears to result from an early trans-formation of the midbrain and posterior forebrain into an anterior hindbrain fate,as indicated by an expansion ofGbx2expression, repression ofOtx2expression,and a rostral shift ofFgf8 expression by the 5- to 7-somite stage, soon after thetransgene is first expressed. In contrast, ectopic expression ofFgf8amainly causesoverproliferation of the midbrain and caudal diencephalon and up-regulation ofEn2, as well asEphrinA2, a prospective downstream target ofEn2 (Logan et al1996) in the anterior dorsal midbrain (Lee et al 1997, Liu et al 1999). This differ-ence between the phenotypes is consistent with results obtained in tissue culturesystems demonstrating that FGF8b is a stronger isoform than FGF8a.
The patterning function of FGF8 in mouse mid/hindbrain development hasalso been investigated in brain explant culture systems (Shimamura & Rubenstein1997, Liu et al 1999, Liu & Joyner 2001). Similar to the in vivo experiments inmice, FGF8 can induceEn1, En2, Pax5, andGbx2, but notFgf8, in both midbrainand forebrain explants.Pax6, a diencephalon gene (Walther & Gruss 1991), isrepressed by FGF8 in caudal forebrain explants butOtx2 is repressed only inmidbrain explants. It is interesting that bothLmx1bandWnt1are initially inducedbroadly by FGF8 in midbrain explants, but subsequentlyWnt1 is excluded fromcells adjacent to the FGF8 source. These explant assays provide a simple in vitromodel system to study the early responses of mes/met genes to FGF8 in normaland mutant brain tissue.
Fgf17 Collaborates with Fgf8 in Patterning theAnterior Cerebellum
As discussed above, following initiation ofFgf8 expression,Fgf17 andFgf18are also expressed in the mid/hindbrain region. Mice lackingFgf17have only amild cerebellar defect, with a decrease in precursor cell proliferation in the medialpart of the cerebellum after E11.5 (Xu et al 2000).Fgf812, 3/+; Fgf17−/− mutantembryos have a more severe phenotype than doFgf17−/− mutants, indicatingthat FGF17 and FGF8 have partially overlapping functions in development of theanterior hindbrain.
OTX1, OTX2, AND GBX2 ARE REQUIRED TO POSITIONTHE ISTHMUS ORGANIZER AND FOR LATER MIDBRAINOR CEREBELLUM DEVELOPMENT, RESPECTIVELY
Multiple Essential Roles for Otx2 in Anterior Patterning
Otx2null mutant embryos have severe gastrulation defects resulting in deletion ofthe rostral part of the neural tube anterior to r3 (Acampora et al 1995, Matsuo et al

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
880 LIU ¥ JOYNER
1995, Ang et al 1996, Rhinn et al 1998) (Figure 4B). The expression patterns ofmarker genes inOtx2mutants revealed that the forebrain and midbrain are absentas early as E7.75 and the metencephalon is deleted by E8.5. Chimeric embryoswere made to distinguish the functions of OTX2 in the visceral endoderm andepiblast (Rhinn et al 1998). Chimeras composed mostly of wild-type cells in theepiblast andOtx2−/− cells in the visceral endoderm have a phenotype similar to
Figure 4 Otx2 is required in the visceral endoderm for normal gastrulation and initiation ofanterior brain development and in the epiblast for the maintenance of anterior brain structures.(A) Otx2 is expressed in both the epiblast and visceral endoderm of wild-type embryos, and itguides normal development of anterior structures. (B) Otx2−/− embryos that lackOtx2expressionin both the epiblast and visceral endoderm fail to gastrulate normally and lose structures anteriorto rhombomere (r)3 at an early stage. (C) In chimeric embryos in whichOtx2 is only expressedin the visceral endoderm, or in embryos in which hOTX1 replacesOtx2and is present only in thevisceral endoderm, forebrain and midbrain structures initially form at early stages, but are lostsubsequently.

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 881
that of Otx2 null mutants. In contrast, chimeras in which the epiblast is mostlycomposed ofOtx2−/− cells and the visceral endoderm wild-type cells undergonearly normal gastrulation but fail to maintain the forebrain and midbrain afterE8 (Figure 4C). Taken together, these studies show that the first requirement forOtx2is in the extraembryonic endoderm and thatOtx2plays a cell nonautonomousrole in the anterior visceral endoderm in inducing the formation of anterior headstructures. This role ofOtx2 likely reflects a function in generating an organizingcenter in the anterior visceral endoderm (for a review, see Beddington & Robertson1999).Otx2also has a second role in the epiblast, where it is required to maintainthe survival and/or identity of rostral brain structures.
Distinct functions forOtx2 in the visceral endoderm and the epiblast were fur-ther demonstrated by analyzing a mouse mutant in which a humanOTX1cDNAwas inserted in place of the endogenousOtx2 gene, such thatOTX1mRNA isonly translated in the visceral endoderm and not the epiblast. Embryos homozy-gous mutant for thisOtx2 allele undergo nearly normal gastrulation and initialanterior brain development but subsequently lose anterior brain structures, includ-ing the forebrain and midbrain by E8.5 (Acampora et al 1998; Y Li, AL Joyner,unpublished data) (Figure 4C). Nevertheless,Fgf8 and other mes/met genes areexpressed even at late stages, but in overlapping domains at the rostral tip of themutant embryo.
Otx1+/−; Otx2+/− (Suda et al 1997) orOtx1−/−; Otx2+/− (Acampora et al 1997)mouse embryos have the striking phenotype that the midbrain and caudal fore-brain are replaced with cerebellar tissue. Gene expression analysis in such mutantsindicates that at early somite stages, expression of all the mes/met genes is initi-ated normally. However,Fgf8 expression soon fails to be restricted to the rostralhindbrain region and expands into more rostral brain regions (Figure 5B). Thisexpansion is followed by a retraction ofOtx2expression and a rostral shift in theEn1/2, Wnt1, andGbx2domains. Therefore, the midbrain and caudal forebrain aretransformed into an anterior hindbrain fate during early neural patterning, whenthe dose of OTX proteins is not sufficient to maintain their normal fate. Thesestudies indicate that the caudal limit ofOtx2could be critical for determining theposition of the isthmic organizer, or theFgf8expression domain, and a high levelof Otx2 is required for midbrain development.
The aboveOtx1/2compound mutant phenotype showed that bothOtx geneshave overlapping functions late in development of the midbrain and caudal fore-brain. In contrast, when mouseOtx1 is inserted into theOtx2 locus such that itis expressed likeOtx2 in the embryo,Otx1can rescue the gastrulation defect ofOtx2 mutant embryos but not the deletion of most of the rostral brain region atearly somite stages (Suda et al 1999). Thus, the two OTX proteins appear to havedifferent functions during early specification of forebrain and midbrain precursors.
Finally, recent gain-of-function studies in whichOtx2was ectopically expressedin the dorsal metencephalon from early somite stages provided further evidence thatOtx2is not only required to position the organizer and direct midbrain development,it also is sufficient to direct both these processes. WhenOtx2was expressed in the

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
882 LIU ¥ JOYNER
Figure 5 Otx2andGbx2regulate mes/met development differently. (A) Schematic repre-sentation of a wild-type E9.5 mouse brain. (B) In Otx1−/−; Otx2+/− embryos, the midbrainand posterior forebrain are transformed into an anterior hindbrain. (C) Ectopic expressionof Otx2 in the dorsal metencephalon from theEn1 locus results in a caudal expansion ofthe midbrain and partial deletion of the metencephalon. (D) In the absence ofGbx2, rhom-bomere (r)1–r3 do not form and the midbrain expands caudally, such that the midbraindirectly abuts r4. (E) Ectopic expression ofGbx2in the posterior midbrain of transgenicsfrom E8.5 leads to a smaller midbrain and an enlarged metencephalon at E9.5.

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 883
dorsal anterior hindbrain of mouse embryos, the caudal midbrain was extendedand the cerebellum partially deleted (Broccoli et al 1999) (Figure 5C). Thesemorphological changes were accompanied by a repression ofFgf8 andGbx2 inthe dorsal anterior most hindbrain from early stages. Similar results were obtainedin chicken embryos by electroporating anterior hindbrain tissue with anOtx2expression construct (Katahira et al 2000).
Gbx2 is Required for Normal Isthmic Organizer Functionand Position as well as Anterior Hindbrain Development
MouseGbx2null mutants die at birth with loss of anterior hindbrain (r1-r3) deriva-tives, which suggests thatGbx2might be involved in maintaining the identity and/orsurvival of the anterior hindbrain (Wassarman et al 1997). Gene expression anal-ysis between E9.5 and E12.5 showed that the caudalOtx2expression border wasadjacent to that of the r4 markerHoxb1, showing an early deletion of the anteriorhindbrain. Furthermore, unlike in wild-type embryos, whereFgf8expression doesnot overlap with that ofOtx2andWnt1in the isthmus, the expression domains ofthe three genes overlap inGbx2mutant embryos. More interesting, the morphologyof the posterior midbrain is not normal, which suggests thatGbx2also contributesto midbrain patterning, possibly indirectly through a function in regulating theorganizer.
More recently, it has been shown that inGbx2−/− embryos, theOtx2domainis greatly expanded caudally by the 4- to 6-somite stage, and its caudal boundaryis never sharp (Millet et al 1999) (Figure 5D). The Wnt1andFgf8 expressiondomains also shift caudally andFgf8 overlapsWnt1andOtx2. These expressionstudies demonstrated that an early phenotype inGbx2mutants is a transformationof presumptive r1–r3 cells into a midbrain fate. In a complementary manner, mis-expression ofGbx2in the midbrain from aWnt1promoter (Wnt1-Gbx2transgenics)at early somite stages results in a repression ofOtx2expression and a rostral shiftof isthmicWnt1andFgf8expression (Millet et al 1999) (Figure 5E). EndogenousGbx2expression is also induced by the transgene. The consequence of the earlychanges in gene expression is a reduction of the midbrain and expansion of thehindbrain at E9.5. Taken together, these two types of mutant studies suggest GBX2functions to antagonizeOtx2expression and midbrain development, in addition tobeing involved in positioning the isthmic organizer.
It is striking that midbrain development begins to recover soon after the trans-gene expression is lost at E9.5, and by E12.5 the brains ofWnt1-Gbx2embryoslook indistinguishable from those of wild types. Thus, the midbrain is still plasticat E9.5 in mice and can recover from early transientGbx2misexpression. In con-trast, transient ectopic expression ofGbx2 in the chick midbrain at HH stage 10leads to repression ofOtx2expression only transiently in rostral midbrain regionsbut permanently in caudal regions. Furthermore, caudal midbrain misexpression ofGbx2results in a rostral shift of the mid/hindbrain junction and a smaller midbrainat late stages (Katahira et al 2000).

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
884 LIU ¥ JOYNER
Taken together, the gain-of-function and loss-of-function studies of bothGbx2andOtx2 mutants provide strong evidence that these genes act antagonistically,and that they are also required for anterior hindbrain and midbrain development,respectively. Furthermore, the two genes are involved in positioning and normalfunctioning of the isthmic organizer. However, the two genes are not required toinitiate expression ofFgf8 and other mes/met genes because in embryos lackingboth Otx2 andGbx2, all the genes are expressed in the rostral CNS (Y Li, ALJoyner, manuscript in preparation).
WNT1, EN1/2, AND PAX2/5 ARE ESSENTIALFOR NORMAL MES/MET DEVELOPMENT
Wnt1 is Required for Mes/Met Developmentand Positioning of the Caudal Otx2 Border
Loss ofWnt1 function leads to death at birth with an early large-scale deletionfirst of the mesencephalon and then of r1 (McMahon & Bradley 1990, Thomas &Capecchi 1990, Mastick et al 1996). Furthermore,En1expression is initiated nor-mally in Wnt1mutants but is lost by the 27-somite stage, before the loss of ther1 tissue (McMahon et al 1992). One possible interpretation of these studies isthat one function ofWnt1 is to maintainEn1 expression. Consistent with this,a transgene with aWnt1enhancer drivingEn1 expression was found to rescuemost of theWnt1mutant phenotype (Danielian & McMahon 1996). However, inWnt1 mutants,Fgf8 expression in the metencephalon is lost by the 14-somitestage, which suggests that WNT1 more directly regulatesFgf8expression in thisregion and that loss ofEn1 could be secondary to the loss ofFgf8 (Lee et al1997). BecauseEn1 misexpression in chicks leads to induction ofFgf8 in bothmidbrain and diencephalon (see below), transgene drivenEn1expression inWnt1mutants might lead to maintenance ofFgf8 expression and, thus, rescue of thephenotype.
Analysis of aWnt1mutant allele,Swaying, that appears to be a hypomorphhas revealed a function forWnt1 in maintaining a stable mid/hindbrain junc-tion (Thomas et al 1991, Bally-Cuif et al 1995).Swayingcontains a frame-shiftmutation that causes premature termination of WNT1 protein, such that a largeportion of the C terminus of WNT1 is deleted.Wnt1sw/sw mutant mice have aless-severe brain phenotype compared with targetedWnt1null mutants, with themidbrain and hindbrain regions being only partially reduced. It is striking thatanalysis of the relatively late (E9.0 and onward) midbrain and hindbrain pheno-type of Swayingmutants showed that formation of the straightOtx2 andWnt1caudal expression borders was perturbed, and small ectopic islands ofOtx2-positive (mes) orOtx2-negative (met) cells were located inappropriately withinthe hindbrain or midbrain, respectively. Moreover,Wnt1expression was inducedat the border of the ectopic mes and met islands, consistent with the idea that

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 885
interactions between mes (Otx2 positive) and met (Otx2 negative) cells posi-tively regulate organizer gene expression, as reflected byWnt1expression at theboundary.
Wnt1gain-of-function studies have failed to demonstrate thatWnt1is sufficientto alter mes/met patterning. In chicks,Wnt1 misexpression in the midbrain atHH stage 9–12 does not result in any phenotype, although in the telencephalonit causes overproliferation (Adams et al 2000). Furthermore, ectopic expressionof Wnt1in the ventricular zone of the neural tube posterior to the r6/r7 boundarybetween E8.5 and E10.5 results only in expansion of the ventricular zone and hasno obvious changes in patterning (Dickinson et al 1994).
A recent study showed that ectopicLmx1bexpression leads to up-regulationof Wnt1expression in chick midbrain (Adams et al 2000). Furthermore,Lmx1bis induced in chick caudal forebrain by FGF8-soaked beads. This study suggeststhatLmx1bmight be in the same pathway asWnt1downstream ofFgf8and couldplay a role in early mes/met patterning. The requirement forLmx1bin mes/metdevelopment has not been reported, althoughLmx1bmouse mutants have beengenerated (Chen et al 1998).
En Genes are Required for Mes/Met Developmentand Likely Repress Forebrain Development
En1mutant mice have a deletion including most of the midbrain and cerebellum,a phenotype similar to, but milder than, that ofWnt1null mutants (Wurst et al1994). UnlikeEn1mutants,En2mutants are viable and show only subtle defectsin cerebellar development, which include an early reduction in size of the cerebellaranlage and later abnormal foliation (Joyner et al 1991; Millen et al 1994, 1995). Thissuggests at least one role forEn2is in patterning the cerebellum. Two experimentshave demonstrated that the two EN proteins can carry out similar functions in thebrain, given correct temporal and spatial expression of the genes. First, when theEn1 coding sequences are replaced with those of anEn2 cDNA using a gene-targeting approach, the mes/met phenotype ofEn1mutants is rescued (Hanks et al1995). It is interesting thatEn2cannot fully rescue all the limb defects seen inEn1mutants, and it is significant that Drosophilaencan rescue theEn1brain defectsbut none of the limb defects (Hanks et al 1998). Second, double mutants of the twoEngenes have a more severe early phenotype than do either of the single mutants(Liu & Joyner 2001). In such double mutants,Wnt1, Fgf8, andPax5expressionis initiated by the 5-somite stage but fails to be maintained in the mes/met by the11-somite stage. Taken together, these studies show that an EN protein must bepresent during early somite stages whenEn1, but notEn2, is normally expressed.Without early expression of an EN protein, further mes/met development is greatlycompromised.
Based on a correlation between the decreasing EN protein gradient from pos-terior to anterior midbrain and the A/P polarity of the midbrain before and after180◦ rotation of the anterior midbrain in chicks, it was suggested that theEngenes

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
886 LIU ¥ JOYNER
could be involved in setting up the topographic axon projection map of the mid-brain (Itasaki et al 1991). This suggestion was addressed by expressingEn genesectopically in the anterior midbrain in both chicken and mouse embryos. In chicks,ectopic expression ofEn1 or En2 in the rostral midbrain inducesEphrinA2andEphrinA5, two genes involved in repulsing retinal temporal axons in the poste-rior midbrain and ectopic projections of retinal nasal axons in the rostral tectaltissue (Friedman & O’Leary 1996, Itasaki & Nakamura 1996, Logan et al 1996).In mice, ectopic expression ofEn1 driven by aWnt1 enhancer causes ectopicEphrinA2expression in the dorsal midline of the rostral midbrain (Lee et al 1997).Thus, the expression level ofEn genes can directly, or indirectly, determine theA/P positional cues of midbrain cells.
In chicks, ectopic expression ofEn2in the diencephalic region results in expan-sion of tectal tissue at the expense of caudal diencephalic tissue (Araki & Nakamura1999). It is striking that this phenotype is accompanied by repression of the di-encephalon genePax6prior to induction of the mes/met genesPax5, Wnt1, andFgf8. Because it has been shown that the EN family of proteins are transcriptionalrepressors (Jaynes & O’Farrell 1991, Smith & Jaynes 1996, Hanks et al 1998), itis possible that EN is normally involved in setting up the mid/forebrain boundaryby repressing forebrain genes. Consistent with this hypothesis, a mutant form ofEN2 in which a single amino acid substitution disrupts its interaction with thecorepressor, GROUCHO, cannot transform the forebrain or induce mid/hindbraingene expression. Furthermore, when a chimeric EN protein in which the repressiondomain of EN2 was replaced by the transactivation domain of VP16 was expressedin chick brain, the midbrain was greatly reduced andPax6expression expandedinto the midbrain region (Araki & Nakamura 1999).
It has also been shown in medaka fish and frog embryos that ubiquitous expres-sion of the fishEn2gene,Ol-eng2, can induce ectopic midbrain development andrepress formation of diencephalic tissue, whereas expression of anOl-eng2genewith a mutation in the homeodomain does not produce such an effect (Ristoratoreet al 1999). It is interesting to note that despite widespread expression of injectedOl-eng2, ectopic expression offgf8, pax2, and endogenouseng2is restricted onlyto regions of the posterior forebrain. Furthermore, the ectopic midbrain that is in-duced always has a reversed A/P polarity. Induction of an ectopic isthmic organizerby Ol-eng2therefore seems to require cofactors that are expressed locally in theforebrain.
The Pax2/5 Genes have Overlapping Functionsin Development of the Mes/Met
The Pax2andPax5genes have been shown to be required for mouse mes/metpatterning using loss-of-function mutants. Several mousePax2 mutant alleleshave been generated and surprisingly they have different phenotypes. One tar-geted deletion allele that removes most of the coding sequences, referred to asPax2ko/ko, appears to show no mes/met A/P patterning defects, just a varied degree

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 887
of exencephaly, depending on the genetic background (Torres et al 1996, Schwarzet al 1997). In contrast, homozygotes for the spontaneousPax21Neuallele, whichcontains a frame-shift mutation that truncates the protein, have a brain deletionthat includes most of the mes/met region (Favor et al 1996). More recently, an-other mousePax2allele was generated by gene targeting, which is thought to be anull allele, and homozygous embryos for this allele have an early deletion of themes/met, similar toPax21Neumutants (Bouchard et al 2000).Pax5−/−mutant mice,in contrast, have only a partial deletion of the inferior colliculi (posterior midbrain)and a slightly enlarged third lobe of the cerebellum (Urbanek et al 1994).
Overlapping functions ofPax2andPax5have been demonstrated using doublemutants. For example,Pax2ko/+; Pax5−/− andPax2ko/ko; Pax5−/−mutants lack mostof the midbrain and cerebellum (Schwarz et al 1997). Although heterozygousKrd mutants, which contain a 7-cM deletion that includes thePax2gene (Kelleret al 1994), andPax5+/− mice are phenotypically normal, compound mutantsKrd+/−; Pax5+/− andKrd+/−; Pax5−/− have deletions of the midbrain and cere-bellum, similar toPax2null mutants (Urbanek et al 1997). Furthermore,Pax5isable to rescue the brain phenotype of thePax2null allele by expressing it from thePax2locus. Therefore, like the EN proteins, a PAX2/5 protein must be expressed atearly somite stages to sustain mes/met development. In addition,Pax6expression,as well as the posterior commisure, a morphological landmark of the posteriordiencephalon, are expanded posteriorly inPax2/5double mutants, which suggeststhatPax2/5might be involved in maintaining mes/met development by repressingforebrain development (Schwarz et al 1999).
The function of the PAX2/5/8 subfamily ofPaxgenes in brain patterning has alsobeen studied in zebrafish. There are four genes in thePax2/5/8family in zebrafish,pax2.1, pax2.2, pax5, andpax8(Lun & Brand 1998). Injection of a neutralizingantibody against PAX2.1 into zebrafish early embryos resulted in a deletion ofmost parts of the midbrain and cerebellum (Krauss et al 1992).Noi (no isthmus)mutants in which the zebrafishpax2.1gene is mutated have a similar deletion of thecaudal midbrain and cerebellum (Brand et al 1996). Expression ofpax2.2, pax5,andpax8 is never detected innoi mutants, indicating that PAX2.1 regulates theotherpaxgenes and that thenoi mutant is actually equivalent to apax2.1/2.2/5/8quadruple mutant (Lun & Brand 1998). Similarly, PAX2 in mice has been shownto be involved in regulatingPax5mes/met expression (Pfeffer et al 2000).
In noi mutants,eng2andeng3expression is not initiated normally, whereasfgf8 andwnt1expression initiates normally but is lost before the mes/met tissueundergoes apoptosis (Pfeffer et al 1998). These changes in mes/met gene expres-sion indicate thatpax2.1 is necessary for the normal pattern of mes/met geneexpression, as well as for the survival of mes/met tissue. ThePaxgenes also havebeen implicated in regulating expression ofEn2 in mice. PAX2/5/8 proteins wereshown to bind to two DNA sequences in anEn2 DNA enhancer fragment thatis sufficient to direct expression of alacZ reporter gene in the isthmus (Loganet al 1993, Song et al 1996). Furthermore, mutation of the PAX2/5/8 binding sitesabolishes the reporter gene expression. Deletion of the two PAX2/5/8-binding sites

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
888 LIU ¥ JOYNER
in the endogenousEn2gene by gene targeting, however, showed that the bindingsites are required only for normal initiation, not maintenance, of endogenousEn2expression (Song & Joyner 2000).
Gain-of-function studies in chicks have shown that thePaxgenes, likeEn1/2andFgf8, can induce midbrain development. Ectopic expression of eitherPax2or Pax5in the diencephalon is sufficient to induce ectopic expression of mes/metgenes, such asFgf8 andEn, and to transform diencephalic tissue into midbrainstructures (Funahashi et al 1999, Okafuji et al 1999). Furthermore,Pax5, but notPax2, misexpression in the anterior midbrain can induceFgf8andEn2expression,indicating that the two PAX proteins have developed some different functionsduring evolution.
CROSS-REGULATION BETWEEN THE MES/METGENES UNDERLIES ORGANIZER FUNCTIONAND MES/MET DEVELOPMENT
The expression studies and functional analyses of mes/met genes have begunto elucidate a set of complex cross-regulatory interactions between these genesduring midbrain and cerebellum development. Some of these same gene familiesare also involved in regulatory networks in Drosophila. For example, thePaxhomologue,prd, is required for initiation ofen andwg expression in ectodermsegments; thenenexpression andwgexpression become dependent on each other,and finallyen is autoregulated (for a review, see Hooper & Scott 1992, Perrimon1994). Some similar, but not identical, regulatory interactions are found duringvertebrate mes/met development. As mentioned above,Pax2expression precedesand overlaps withEn1 andWnt1 in mice, and genetic studies implicatePax2/5in regulatingEn1/2expression in zebrafish and mice. In contrast to the situationin Drosophila, aspects ofWnt1expression do not seem to require PAX2 function(Lun & Brand 1998, Schwarz et al 1999). There is also no evidence for autoregula-tion of theEngenes (Logan 1993, Liu & Joyner 2001).
Several lines of evidence indicate thatEn andWnt1are involved in regulatingeach other’s expression, although it is not clear whether this is direct. As describedabove, inWnt1null mutant mice,En1expression is initiated normally but lost, andin En1/2double mutant embryos,Wnt1expression is rapidly lost. Furthermore,whenEn1expression is maintained inWnt1mutants, a nearly complete rescue oftheWnt1brain phenotype is observed. In addition, ectopic expression ofEn caninduce ectopicWnt1expression (Araki & Nakamura 1999, Ristoratore et al 1999),although the reverse does not seem to be true (Adams et al 2000).
Many gain- and loss-of-function studies have shown thatOtx2 andGbx2areinvolved in a negative feedback loop (see above). We recently explored whetherGbx2acts directly or indirectly to repressOtx2using a midbrain explant system andFGF8-soaked beads (Liu & Joyner 2001). It is surprising that althoughGbx2wasrequired for repressingWnt1expression, it was not required for repressingOtx2.

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 889
Therefore, FGF8 can regulateOtx2expression through aGbx2-independent path-way, and it is possible thatGbx2can repressOtx2through up-regulation ofFgf8.
Finally, as discussed above, FGF8 has been shown to be able to inducePax2,Pax5, En1, En2, Wnt1, Lmx1b, andGbx2in midbrain and caudal forebrain tissue,as well as to repressOtx2. Nevertheless, the normal timing of gene expressionshows thatFgf8 expressed in the metencephalon is unlikely to be responsiblefor inducing initial expression of these mes/met genes. Whether another FGF,possibly one expressed outside the neural tube, initiates mes/met gene expressionis not clear.Fgf8 is, however, likely to be critical for modulating and maintainingexpression of many mes/met genes after the 5-somite stage in mice.
PROSPECTS AND FUTURE DIRECTION
The many genetic studies we have described have revealed that multiple geneticpathways regulate early patterning of the vertebrate midbrain and cerebellum. Anearly step in patterning the mes/met seems to be a genetic interaction at the borderof Otx2- andGbx2-expressing cells, which determines where the midbrain andcerebellum will form. It is also clear that by the 10-somite stage, a self-sustaininggenetic network is set up within the mes/met that is controlled centrally by anisthmic organizer. Furthermore, FGF8-like molecules alone can appropriately reg-ulate the expression of most the other mes/met genes and induce midbrain and cere-bellum development. Thus, FGF8 and related factors are likely central to thisself-sustaining network. It must, however, be more complicated because loss ofEn, Wnt1, or Pax2/5function alone leads to a collapse of mes/met development.
A number of key questions nevertheless remain to be addressed. For example,none of the loss-of-function mutants described seems to lack initial expression ofall other genes in the mes/met region, raising the question of what signal triggersthe mes/met molecular pathway before the early somite stages. Also, the epistaticrelationships among the known genes need to be established using new approachesbecause they have been hard to explore in null mutants owing to simultaneousearly loss of expression of multiple genes. Another important and long-standingquestion is how the signals from the mid/hindbrain organizer are transmitted acrossthe entire length of the midbrain and r1. Does a relay mechanism exist or are signalstransmitted from the isthmus directly to cells at a distance? Finally, it has beenshown that isthmic tissue cannot induce ectopic mes/met development in eitherthe anterior forebrain (Martinez et al 1991) or the spinal cord (Grapin-Botton et al1999), raising the question of what regulates the competence of cells to respondto isthmic organizing signals.
ACKNOWLEDGMENTS
We thank Sandrine Millet and Yuanhao Li for insightful discussions and help-ful comments on the manuscript. The work described from our laboratory was

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
890 LIU ¥ JOYNER
supported by grants from the NINDS to ALJ. ALJ is an investigator of the HowardHughes Medical Institute.
Visit the Annual Reviews home page at www.AnnualReviews.org
LITERATURE CITED
Acampora D, Avantaggiato V, Tuorto F, Bri-ata P, Corte G, Simeone A. 1998. Visceralendoderm-restricted translation of Otx1 me-diates recovery of Otx2 requirements forspecification of anterior neural plate and nor-mal gastrulation.Development125:5091–104
Acampora D, Avantaggiato V, Tuorto F, Sime-one A. 1997. Genetic control of brain mor-phogenesis through Otx gene dosage require-ment.Development124:3639–50
Acampora D, Mazan S, Lallemand Y, Avan-taggiato V, Maury M, et al. 1995. Forebrainand midbrain regions are deleted in Otx2−/−mutants due to a defective anterior neuroec-toderm specification during gastrulation.De-velopment121:3279–90
Adams B, Dorfler P, Aguzzi A, Kozmik Z, Ur-banek P, et al. 1992. Pax-5 encodes the tran-scription factor BSAP and is expressed in Blymphocytes, the developing CNS, and adulttestis.Genes Dev.6:1589–607
Adams KA, Maida JM, Golden JA, Riddle RD.2000. The transcription factor Lmx1b main-tains Wnt1 expression within the isthmic or-ganizer.Development127:1857–67
Alvarado-Mallart RM. 1993. Fate and po-tentialities of the avian mesencephalic/metencephalic neuroepithelium.J. Neuro-biol. 24:1341–55
Ang SL, Jin O, Rhinn M, Daigle N, Steven-son L, Rossant J. 1996. A targeted mouseOtx2 mutation leads to severe defects in gas-trulation and formation of axial mesodermand to deletion of rostral brain.Development122:243–52
Araki I, Nakamura H. 1999. Engrailed de-fines the position of dorsal di-mesencephalicboundary by repressing diencephalic fate.Development126:5127–35
Arman E, Haffner-Krausz R, Chen Y, Heath JK,Lonai P. 1998. Targeted disruption of fibro-blast growth factor (FGF) receptor 2 suggestsa role for FGF signaling in pregastrulationmammalian development.Proc. Natl. Acad.Sci. USA95:5082–87
Arman E, Haffner-Krausz R, Gorivodsky M,Lonai P. 1999. Fgfr2 is required for limb out-growth and lung-branching morphogenesis.Proc. Natl. Acad. Sci. USA96:11895–99
Bally-Cuif L, Cholley B, Wassef M. 1995.Involvement of Wnt-1 in the formation ofthe mes/metencephalic boundary.Mech. Dev.53:23–34
Basilico C, Moscatelli D. 1992. The FGF familyof growth factors and oncogenes.Adv. Can-cer Res.59:115–65
Beddington SP, Robertson EJ. 1999. Axis de-velopment and early asymmetry in mam-mals.Cell 96:195–209
Blunt AG, Lawshe A, Cunningham ML, SetoML, Ornitz DM, MacArthur CA. 1997. Over-lapping expression and redundant activa-tion of mesenchymal fibroblast growth fac-tor (FGF) receptors by alternatively splicedFGF-8 ligands.J. Biol. Chem.272:3733–38
Bouchard M, Pfeffer P, Busslinger M. 2000.Functional equivalence of the transcriptionfactors Pax2 and Pax5 in mouse develop-ment.Development127:3703–13
Bouillet P, Chazaud C, Oulad-AbdelghaniM, Dolle P, Chambon P. 1995. Sequenceand expression pattern of the Stra7 (Gbx-2) homeobox-containing gene induced byretinoic acid in P19 embryonal carcinomacells.Dev. Dyn.204:372–82
Brand M, Heisenberg CP, Jiang YJ, BeuchleD, Lun K, et al. 1996. Mutations in zebrafishgenes affecting the formation of the boundary

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 891
between midbrain and hindbrain.Develop-ment123:179–90
Broccoli V, Boncinelli E, Wurst W. 1999. Thecaudal limit of Otx2 expression positions theisthmic organizer.Nature401:164–68
Carl M, Wittbrodt J. 1999. Graded interferencewith FGF signalling reveals its dorsoventralasymmetry at the mid-hindbrain boundary.Development126:5659–67
Chambers D, Mason I. 2000. Expression ofsprouty2 during early development of thechick embryo is coincident with known sitesof FGF signalling.Mech. Dev.91:361–64
Chambers D, Medhurst AD, Walsh FS, Price J,Mason I. 2000. Differential display of genesexpressed at the midbrain-hindbrain junc-tion identifies sprouty2: an FGF8-induciblemember of a family of intracellular FGFantagonists.Mol. Cell. Neurosci. 15:22–35
Chen H, Lun Y, Ovchinnikov D, Kokubo H,Oberg KC, et al. 1998. Limb and kidney de-fects in Lmx1b mutant mice suggest an in-volvement of LMX1B in human nail patellasyndrome.Nat. Genet.19:51–55
Ciruna BG, Schwartz L, Harpal K, YamaguchiTP, Rossant J. 1997. Chimeric analysis offibroblast growth factor receptor-1 (Fgfr1)function: a role for FGFR1 in morphogeneticmovement through the primitive streak.De-velopment124:2829–41
Colvin JS, Bohne BA, Harding GW, McEwenDG, Ornitz DM. 1996. Skeletal overgrowthand deafness in mice lacking fibroblastgrowth factor receptor 3.Nat. Genet.12:390–97
Coulier F, Pontarotti P, Roubin R, Hartung H,Goldfarb M, Birnbaum D. 1997. Of wormsand men: an evolutionary perspective on thefibroblast growth factor (FGF) and FGF re-ceptor families.J. Mol. Evol.44:43–56
Crossley PH, Martin GR. 1995. The mouse Fgf8gene encodes a family of polypeptides andis expressed in regions that direct outgrowthand patterning in the developing embryo.De-velopment121:439–51
Crossley PH, Martinez S, Martin GR. 1996.
Midbrain development induced by FGF8 inthe chick embryo.Nature380:66–68
Danielian PS, McMahon AP. 1996. Engrailed-1as a target of the Wnt-1 signalling pathwayin vertebrate midbrain development.Nature383:332–34
Davis CA, Joyner AL. 1988. Expression pat-terns of the homeo box-containing genes En-1 and En-2 and the proto-oncogene int-1diverge during mouse development.GenesDev.2:1736–44
Deng CX, Wynshaw-Boris A, Shen MM,Daugherty C, Ornitz DM, Leder P. 1994.Murine FGFR-1 is required for early postim-plantation growth and axial organization.Genes Dev.8:3045–57
Deng CX, Wynshaw-Boris A, Zhou F, KuoA, Leder P. 1996. Fibroblast growth factorreceptor 3 is a negative regulator of bonegrowth.Cell 84:911–21
Dickinson ME, Krumlauf R, McMahon AP.1994. Evidence for a mitogenic effect ofWnt-1 in the developing mammalian centralnervous system.Development120:1453–71
Favor J, Sandulache R, Neuhauser-Klaus A,Pretsch W, Chatterjee B, et al. 1996. Themouse Pax2(1Neu) mutation is identical to ahuman PAX2 mutation in a family with renal-coloboma syndrome and results in develop-mental defects of the brain, ear, eye, and kid-ney.Proc. Natl. Acad. Sci. USA93:13870–75
Feldman B, Poueymirou W, Papaioannou VE,DeChiara TM, Goldfarb M. 1995. Require-ment of FGF-4 for postimplantation mousedevelopment.Science267:246–49
Friedman GC, O’Leary DD. 1996. Retroviralmisexpression of engrailed genes in the chickoptic tectum perturbs the topographic target-ing of retinal axons.J. Neurosci.16:5498–509
Funahashi J, Okafuji T, Ohuchi H, Noji S,Tanaka H, Nakamura H. 1999. Role of Pax-5 in the regulation of a mid-hindbrain orga-nizer’s activity.Dev. Growth Differ.41:59–72
Grapin-Botton A, Cambronero F, Weiner HL,Bonnin MA, Puelles L, Le Douarin NM.1999. Patterning signals acting in the spinal

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
892 LIU ¥ JOYNER
cord override the organizing activity of theisthmus.Mech. Dev.84:41–53
Gruss P, Walther C. 1992. Pax in development.Cell 69:719–22
Hamburger V, Hamilton HL. 1992. A series ofnormal stages in the development of the chickembryo.Dev. Dyn.195:231–72
Hanks M, Wurst W, Anson-Cartwright L, Auer-bach AB, Joyner AL. 1995. Rescue of the En-1 mutant phenotype by replacement of En-1with En-2.Science269:679–82
Hanks MC, Loomis CA, Harris E, Tong CX,Anson-Cartwright L, et al. 1998. Drosophilaengrailed can substitute for mouse En-grailed1 function in mid-hindbrain, but notlimb development.Development125:4521–30
Hebert JM, Rosenquist T, Gotz J, Martin GR.1994. FGF5 as a regulator of the hair growthcycle: evidence from targeted and sponta-neous mutations.Cell 78:1017–25
Heikinheimo M, Lawshe A, Shackleford GM,Wilson DB, MacArthur CA. 1994. Fgf-8 ex-pression in the post-gastrulation mouse sug-gests roles in the development of the face,limbs and central nervous system.Mech. Dev.48:129–38
Hidalgo-Sanchez M, Simeone A, Alvarado-Mallart RM. 1999. Fgf8 and Gbx2 inductionconcomitant with Otx2 repression is corre-lated with midbrain-hindbrain fate of caudalprosencephalon.Development 126:3191–203
Hooper JE, Scott MP. 1992. The molecular ge-netic basis of positional information in insectsegments.Results Probl. Cell Differ.18:1–48
Irving C, Mason I. 1999. Regeneration of isth-mic tissue is the result of a specific and directinteraction between rhombomere 1 and mid-brain.Development126:3981–89
Irving C, Mason I. 2000. Signalling by FGF8from the isthmus patterns anterior hindbrainand establishes the anterior limit of Hox geneexpression.Development127:177–86
Itasaki N, Ichijo H, Hama C, Matsuno T, Naka-mura H. 1991. Establishment of rostrocaudalpolarity in tectal primordium: engrailed ex-
pression and subsequent tectal polarity.De-velopment113:1133–44
Itasaki N, Nakamura H. 1996. A role for gradi-ent en expression in positional specificationon the optic tectum.Neuron16:55–62
Jaynes JB, O’Farrell PH. 1991. Active repres-sion of transcription by the engrailed home-odomain protein.EMBO J.10:1427–33
Joyner AL, Herrup K, Auerbach BA, Davis CA,Rossant J. 1991. Subtle cerebellar phenotypein mice homozygous for a targeted deletionof the En-2 homeobox.Science251:1239–43
Joyner AL, Liu A, Millet S. 2000. Otx2, Gbx2and Fgf8 interact to position and maintaina mid-hindbrain organizer.Curr. Opin. Cell.Biol. 12:736–41
Joyner AL, Martin GR. 1987. En-1 and En-2,two mouse genes with sequence homologyto the Drosophila engrailed gene: expressionduring embryogenesis.Genes Dev.1:29–38
Katahira T, Sato T, Sugiyama S, Okafuji T,Araki I, et al. 2000. Interaction between Otx2and Gbx2 defines the organizing center forthe optic tectum.Mech. Dev.91:43–52
Keller SA, Jones JM, Boyle A, Barrow LL,Killen PD, et al. 1994. Kidney and retinaldefects (Krd), a transgene-induced mutationwith a deletion of mouse chromosome 19 thatincludes the Pax2 locus.Genomics23:309–20
Krauss S, Maden M, Holder N, Wilson SW.1992. Zebrafish pax[b] is involved in the for-mation of the midbrain-hindbrain boundary.Nature360:87–89
Lee SM, Danielian PS, Fritzsch B, McMahonAP. 1997. Evidence that FGF8 signallingfrom the midbrain-hindbrain junction regu-lates growth and polarity in the developingmidbrain.Development124:959–69
Liu A, Joyner AL. 2001. EN and GBX2 playessential roles downstream of FGF8 in pat-terning the mouse mid/hindbrain region.De-velopment128:181–91
Liu A, Losos K, Joyner AL. 1999. FGF8 canactivate Gbx2 and transform regions of therostral mouse brain into a hindbrain fate.De-velopment126:4827–38

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 893
Logan C, Khoo WK, Cado D, Joyner AL. 1993.Two enhancer regions in the mouse En-2 lo-cus direct expression to the mid/hindbrainregion and mandibular myoblasts.Develop-ment117:905–16
Logan C, Wizenmann A, Drescher U, Mon-schau B, Bonhoeffer F, Lumsden A. 1996.Rostral optic tectum acquires caudal charac-teristics following ectopic engrailed expres-sion.Curr. Biol. 6:1006–14
Lun K, Brand M. 1998. A series of no isth-mus (noi) alleles of the zebrafish pax2.1 genereveals multiple signaling events in devel-opment of the midbrain-hindbrain boundary.Development125:3049–62
MacArthur CA, Lawshe A, Shankar DB, Heik-inheimo M, Shackleford GM. 1995a. FGF-8isoforms differ in NIH3T3 cell transformingpotential.Cell Growth Differ.6:817–25
MacArthur CA, Lawshe A, Xu J, Santos-Ocampo S, Heikinheimo M, et al. 1995b.FGF-8 isoforms activate receptor spliceforms that are expressed in mesenchymal re-gions of mouse development.Development121:3603–13
Mansour SL, Goddard JM, Capecchi MR. 1993.Mice homozygous for a targeted disruptionof the proto-oncogene int-2 have develop-mental defects in the tail and inner ear.De-velopment117:13–28
Martinez S, Crossley PH, Cobos I, RubensteinJL, Martin GR. 1999. FGF8 induces for-mation of an ectopic isthmic organizer andisthmocerebellar development via a repres-sive effect on Otx2 expression.Development126:1189–200
Martinez S, Wassef M, Alvarado-Mallart RM.1991. Induction of a mesencephalic pheno-type in the 2-day-old chick prosencephalonis preceded by the early expression of thehomeobox gene en.Neuron6:971–81
Maruoka Y, Ohbayashi N, Hoshikawa M, ItohN, Hogan BLM, Furuta Y. 1998. Compari-son of the expression of three highly relatedgenes, Fgf8, Fgf17 and Fgf18, in the mouseembryo.Mech. Dev.74:175–77
Mastick GS, Fan CM, Tessier-Lavigne M,
Serbedzija GN, McMahon AP, Easter SSJr. 1996. Early deletion of neuromeres inWnt-1−/− mutant mice: evaluation by mor-phological and molecular markers.J. Comp.Neurol.374:246–58
Matsuo I, Kuratani S, Kimura C, Takeda N,Aizawa S. 1995. Mouse Otx2 functions inthe formation and patterning of rostral head.Genes Dev.9:2646–58
McMahon AP, Bradley A. 1990. The Wnt-1(int-1) proto-oncogene is required for devel-opment of a large region of the mouse brain.Cell 62:1073–85
McMahon AP, Joyner AL, Bradley A, McMa-hon JA. 1992. The midbrain-hindbrain phe-notype of Wnt-1-/Wnt-1-mice results fromstepwise deletion of engrailed-expressingcells by 9.5 days postcoitum.Cell 69:581–95
Meinhardt H. 1983. Cell determination bound-aries as organizing regions for secondary em-bryonic fields.Dev. Biol.96:375–85
Meyers EN, Lewandoski M, Martin GR. 1998.An Fgf8 mutant allelic series generated byCre- and Flp-mediated recombination.Nat.Genet.18:136–41
Millen KJ, Hui CC, Joyner AL. 1995. A rolefor En-2 and other murine homologues ofDrosophila segment polarity genes in regu-lating positional information in the develop-ing cerebellum.Development121:3935–45
Millen KJ, Wurst W, Herrup K, Joyner AL.1994. Abnormal embryonic cerebellar devel-opment and patterning of postnatal foliationin two mouse Engrailed-2 mutants.Develop-ment120:695–706
Millet S, Bloch-Gallego E, Simeone A,Alvarado-Mallart RM. 1996. The caudallimit of Otx2 gene expression as a marker ofthe midbrain/hindbrain boundary: a study us-ing in situ hybridisation and chick/quail ho-motopic grafts.Development122:3785–97
Millet S, Campbell K, Epstein DJ, Losos K,Harris E, Joyner AL. 1999. A role for Gbx2in repression of Otx2 and positioning themid/hindbrain organizer.Nature401:161–64
Min H, Danilenko DM, Scully SA, Bolon B,

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
894 LIU ¥ JOYNER
Ring BD, et al. 1998. Fgf-10 is required forboth limb and lung development and exhibitsstriking functional similarity to Drosophilabranchless.Genes Dev.12:3156–61
Minowada G, Jarvis LA, Chi CL, Neubuser A,Sun X, et al. 1999. Vertebrate Sprouty genesare induced by FGF signaling and can causechondrodysplasia when overexpressed.De-velopment126:4465–75
Nieto MA. 1999. Reorganizing the organizer 75years on.Cell 98:417–25
Nusse R, Varmus HE. 1992. Wnt genes.Cell69:1073–87
Okafuji T, Funahashi J, Nakamura H. 1999.Roles of Pax-2 in initiation of the chick tec-tal development.Brain Res. Dev. Brain Res.116:41–49
Orr-Urtreger A, Bedford MT, Burakova T, Ar-man E, Zimmer Y, et al. 1993. Developmentallocalization of the splicing alternatives of fi-broblast growth factor receptor-2 (FGFR2).Dev. Biol.158:475–86
Perrimon N. 1994. The genetic basis of pat-terned baldness in Drosophila.Cell 76:781–84
Peters K, Ornitz D, Werner S, Williams L. 1993.Unique expression pattern of the FGF recep-tor 3 gene during mouse organogenesis.Dev.Biol. 155:423–30
Peters KG, Werner S, Chen G, Williams LT.1992. Two FGF receptor genes are differ-entially expressed in epithelial and mes-enchymal tissues during limb formation andorganogenesis in the mouse.Development114:233–43
Pfeffer PL, Bouchard M, Busslinger M. 2000.Pax2 and homeodomain proteins coopera-tively regulate a 435 bp enhancer of themouse Pax5 gene at the midbrain-hindbrainboundary.Development127:1017–28
Pfeffer PL, Gerster T, Lun K, Brand M, Bus-slinger M. 1998. Characterization of threenovel members of the zebrafish Pax2/5/8family: dependency of Pax5 and Pax8expression on the Pax2.1 (noi) function.De-velopment125:3063–74
Picker A, Brennan C, Reifers F, Clarke JD,
Holder N, Brand M. 1999. Requirementfor the zebrafish mid-hindbrain boundary inmidbrain polarisation, mapping and confine-ment of the retinotectal projection.Develop-ment126:2967–78
Plotnikov AN, Schlessinger J, Hubbard SR,Mohammadi M. 1999. Structural basis forFGF receptor dimerization and activation.Cell 98:641–50
Reifers F, Bohli H, Walsh EC, Crossley PH,Stainier DY, Brand M. 1998. Fgf8 is mu-tated in zebrafish acerebellar (ace) mutantsand is required for maintenance of midbrain-hindbrain boundary development and somi-togenesis.Development125:2381–95
Rhinn M, Dierich A, Shawlot W, Behringer RR,Le Meur M, Ang SL. 1998. Sequential rolesfor Otx2 in visceral endoderm and neuroec-toderm for forebrain and midbrain inductionand specification.Development125:845–56
Ristoratore F, Carl M, Deschet K, Richard-Parpaillon L, Boujard D, et al. 1999.The midbrain-hindbrain boundary geneticcascade is activated ectopically in the di-encephalon in response to the widespreadexpression of one of its components, themedaka gene Ol-eng2Development126:3769–79
Rowitch DH, McMahon AP. 1995. Pax-2 ex-pression in the murine neural plate precedesand encompasses the expression domains ofWnt-1 and En-1.Mech. Dev.52:3–8
Schwarz M, Alvarez-Bolado G, Dressler G, Ur-banek P, Busslinger M, Gruss P. 1999. Pax2/5and Pax6 subdivide the early neural tube intothree domains.Mech. Dev.82:29–39
Schwarz M, Alvarez-Bolado G, Urbanek P,Busslinger M, Gruss P. 1997. Conserved bio-logical function between Pax-2 and Pax-5 inmidbrain and cerebellum development: evi-dence from targeted mutations.Proc. Natl.Acad. Sci. USA94:14518–23
Sekine K, Ohuchi H, Fujiwara M, YamasakiM, Yoshizawa T, et al. 1999. Fgf10 is essen-tial for limb and lung formation.Nat. Genet.21:138–41
Shamim H, Mahmood R, Logan C, Doherty P,

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
MID/HINDBRAIN EARLY PATTERNING 895
Lumsden A, Mason I. 1999. Sequential rolesfor Fgf4, En1 and Fgf8 in specification andregionalisation of the midbrain.Development126:945–59
Shimamura K, Rubenstein JL. 1997. Inductiveinteractions direct early regionalization of themouse forebrain.Development124:2709–18
Simeone A, Acampora D, Gulisano M, Stor-naiuolo A, Boncinelli E. 1992. Nested ex-pression domains of four homeobox genes indeveloping rostral brain.Nature358:687–90
Simeone A, Acampora D, Mallamaci A, Stor-naiuolo A, D’Apice MR, et al. 1993. A verte-brate gene related to orthodenticle contains ahomeodomain of the bicoid class and demar-cates anterior neuroectoderm in the gastru-lating mouse embryo.EMBO J.12:2735–47
Slack JM, Isaacs HV, Song J, Durbin L, PownallME. 1996. The role of fibroblast growth fac-tors in early Xenopus development.Biochem.Soc. Symp.62:1–12
Smith ST, Jaynes JB. 1996. A conserved re-gion of engrailed, shared among all en-, gsc-,Nk1-, Nk2- and msh-class homeoproteins,mediates active transcriptional repression invivo. Development122:3141–50
Song DL, Chalepakis G, Gruss P, Joyner AL.1996. Two Pax-binding sites are requiredfor early embryonic brain expression of anEngrailed-2 transgene.Development122:627–35
Song DL, Joyner AL. 2000. Two Pax2/5/8-binding sites in Engrailed-2 are requiredfor proper initiation of endogenous mid-hindbrain expression.Mech. Dev.90:155–65
Stark KL, McMahon JA, McMahon AP. 1991.FGFR-4, a new member of the fibroblastgrowth factor receptor family, expressed inthe definitive endoderm and skeletal mus-cle lineages of the mouse.Development113:641–51
Suda Y, Matsuo I, Aizawa S. 1997. Coop-eration between Otx1 and Otx2 genes indevelopmental patterning of rostral brain.Mech. Dev.69:125–41
Suda Y, Nakabayashi J, Matsuo I, Aizawa S.1999. Functional equivalency between Otx2
and Otx1 in development of the rostral head.Development126:743–57
Sun X, Meyers EN, Lewandoski M, MartinGR. 1999. Targeted disruption of Fgf8 causesfailure of cell migration in the gastrulatingmouse embryo.Genes Dev.13:1834–46
Tanaka A, Miyamoto K, Minamino N, TakedaM, Sato B, et al. 1992. Cloning and char-acterization of an androgen-induced growthfactor essential for the androgen-dependentgrowth of mouse mammary carcinoma cells.Proc. Natl. Acad. Sci. USA89:8928–32
Thomas KR, Capecchi MR. 1990. Targeted dis-ruption of the murine int-1 proto-oncogeneresulting in severe abnormalities in mid-brain and cerebellar development.Nature346:847–50
Thomas KR, Musci TS, Neumann PE, Capec-chi MR. 1991. Swaying is a mutant allele ofthe proto-oncogene Wnt-1.Cell 67:969–76
Torres M, Gomez-Pardo E, Gruss P. 1996. Pax2contributes to inner ear patterning and opticnerve trajectory.Development122:3381–91
Urbanek P, Fetka I, Meisler MH, BusslingerM. 1997. Cooperation of Pax2 and Pax5 inmidbrain and cerebellum development.Proc.Natl. Acad. Sci. USA94:5703–8
Urbanek P, Wang ZQ, Fetka I, Wagner EF, Bus-slinger M. 1994. Complete block of earlyB cell differentiation and altered patterningof the posterior midbrain in mice lackingPax5/BSAP.Cell 79:901–12
Walshe J, Mason I. 2000. Expression of FGFR1,FGFR2 and FGFR3 during early neural de-velopment in the chick embryo.Mech. Dev.90:103–110
Walther C, Gruss P. 1991. Pax-6, a murinepaired box gene, is expressed in the devel-oping CNS.Development113:1435–49
Wassarman KM, Lewandoski M, Campbell K,Joyner AL, Rubenstein JL, et al. 1997. Spec-ification of the anterior hindbrain and estab-lishment of a normal mid/hindbrain organizeris dependent on Gbx2 gene function.Devel-opment124:2923–34
Wassef M, Joyner AL. 1997. Early mes-encephalon/metencephalon patterning and

P1: GDL
May 1, 2001 10:35 Annual Reviews AR121-28
896 LIU ¥ JOYNER
development of the cerebellum.Perspect.Dev. Neurobiol.5:3–16
Weinstein M, Xu X, Ohyama K, Deng CX.1998. FGFR-3 and FGFR-4 function coop-eratively to direct alveogenesis in the murinelung.Development125:3615–23
Wilkinson DG, Bailes JA, McMahon AP. 1987.Expression of the proto-oncogene int-1 is re-stricted to specific neural cells in the devel-oping mouse embryo.Cell 50:79–88
Wingate RJ, Hatten ME. 1999. The role ofthe rhombic lip in avian cerebellum devel-opment.Development126:4395–404
Wurst W, Auerbach AB, Joyner AL. 1994. Mul-tiple developmental defects in Engrailed-1mutant mice: an early mid-hindbrain dele-tion and patterning defects in forelimbs andsternum.Development120:2065–75
Xu J, Lawshe A, MacArthur CA, Ornitz DM.1999. Genomic structure, mapping, activityand expression of fibroblast growth factor 17.Mech. Dev.83:165–78
Xu J, Liu Z, Ornitz DM. 2000. Temporal
and spatial gradients of Fgf8 and Fgf17regulate proliferation and differentiation ofmidline cerebellar structures.Development127:1833–43
Xu X, Weinstein M, Li C, Naski M, Cohen RI,et al. 1998. Fibroblast growth factor receptor2 (FGFR2)-mediated reciprocal regulationloop between FGF8 and FGF10 is essentialfor limb induction.Development125:753–65
Yamaguchi TP, Conlon RA, Rossant J. 1992.Expression of the fibroblast growth factor re-ceptor FGFR-1/flg during gastrulation andsegmentation in the mouse embryo.Dev.Biol. 152:75–88
Yamaguchi TP, Harpal K, Henkemeyer M,Rossant J. 1994. fgfr-1 is required for embry-onic growth and mesodermal patterning dur-ing mouse gastrulation.Genes Dev.8:3032–44
Yuan S, Schoenwolf GC. 1999. The spatial andtemporal pattern of C-Lmx1 expression inthe neuroectoderm during chick neurulation.Mech. Dev.88:243–47

P1: FQP
April 28, 2001 17:49 Annual Reviews AR121-28-COLOR
Figure 1 Expression patterns of mes/met genes in the mouse at E8.5 and E9.5. (A) Atthe 3-5 somite stage (E8.5),Otx2 andWnt1are expressed broadly in the midbrain withOtx2also in the forebrain.Fgf8 andGbx2are expressed broadly in the anterior hindbrain,andEn1, En2, Pax2andPax5are expressed in the entire mes/met region. (B) At E9.5,Wnt1expression is restricted to a narrow ring anterior to the mid/hindbrain junction.Pax2,Fgf8 andGbx2are expressed in narrow rings caudal to the mid/hindbrain junction.En1,En2andPax5are expressed in regions of the midbrain and anterior hindbrain. The thicklines indicate that expression is along the entire D-V axis and the thin lines indicate thatexpression is only in the dorsal or ventral midline of the neural tube.