Embed Size (px)
Citation preview

Journal of Theoretical Biology 229 (2004) 197–208
ARTICLE IN PRESS
*Correspond
386-231.
E-mail addr
University of L
0022-5193/$ - se
doi:10.1016/j.jtb
Dynamic polyhedral models of globular proteins
B. Tom Burnleya,1, Jonathan P.L. Coxb,*aDepartment of Biology and Biochemistry, University of Bath, Bath BA2 7AY, UK
bDepartment of Chemistry, University of Bath, Bath BA2 7AY, UK
Received 21 January 2004; received in revised form 25 March 2004; accepted 26 March 2004
Abstract
We have devised several mechanical models of globular proteins by approximating them to various polyhedra (dodecahedron,
truncated octahedron, icosahedron, truncated icosahedron). The models comprise hollow blocks linked together in a flexible chain.
Between blocks there is a set of several reversible, weak magnetic interactions such that when the chain is agitated, it will fold into a
stable polyhedral structure about the size of a hand. Folding may be followed in real time with a video camera. Key to the success of
the folding process is the lightness of the chain. Several side chains may also be added to the blocks such that they come together to
create a polyhedral core when the chain folds. The models have a number of similarities to globular proteins: each chain folds into a
unique, but dynamic, three-dimensional structure; the instructions that determine this structure are built into the configuration of
blocks; and it is difficult to predict this structure given the unfolded block configuration. Furthermore, the chains fold quickly,
generally in less than a minute, several pathways are involved, and these pathways progress through elements of ‘‘native’’ structure.
In particular, the models emphasize the importance of restricted conformational mobility in assisting the chain to fold, and also in
eliminating undesirable interactions. Because of these similarities to globular proteins, we believe that the polyhedral models will,
with continued development, be helpful in understanding the protein folding process, while at the same time acting as valuable
educational visual aids. They might also inspire the construction of new types of microscopic, self-assembling devices.
r 2004 Elsevier Ltd. All rights reserved.
Keywords: Globular protein; Protein folding; Dynamic mechanical model; Polyhedron; Magnetic interaction
1. Introduction
Although the folding of globular proteins has beenextensively modelled using computers (see, for example,Shea and Brooks, 2001), no one has yet produced amechanical model of this process (‘‘model’’ in the senseof a molecular model kit, an object that can be held inthe hand). Such a model would be useful both from ascientific point of view, and from an educational one.Scientifically, it would offer a new and radically differentapproach to the protein-folding problem (Dobson,2003), and could give fresh insight into this process.Educationally, the model would help convey the
ing author. Tel.: +44-1225-386-548; fax: +44-1225-
esses: [email protected] (B.T. Burnley),
ac.uk (J.P.L. Cox).
ress: School of Biochemistry and Molecular Biology,
eeds, Leeds, LS2 9JT, UK.
e front matter r 2004 Elsevier Ltd. All rights reserved.
i.2004.03.020
remarkable phenomenon of protein folding to school-children and students.
Here we describe several abstract, mechanical modelsof a globular protein. The models are based on twofacts. The first is that a globular protein, as its namesuggests, can be thought of as a sphere, and a sphere canbe approximated to a convex polyhedron. The second isthat many different continuous paths can be drawnthrough the faces of a polyhedron, such that each pathpasses through every face once and once only.
Each of our models comprises a series of blocksconnected to each other by flexible linkages to reproduceone of these continuous paths. The blocks in the chainmay be thought of as amino acids in a polypeptidechain. The lower surface of each block corresponds toone face of a polyhedron. Thus the chain of blocks canbe folded into a polyhedron, or more precisely apolyhedral shell. The type of polyhedron created bythis folding process is dictated by the shape of the

ARTICLE IN PRESS
Fig. 1. Component block of a polyhedral model. The pentagonal
block shown is part of either a dodecahedral or truncated icosahedral
model. C: center of polyhedron; Inter-radius: inter-radius of poly-
hedron. Upper, lower and side edges of the block are indicated. The
circle on the front side face shows where a barium ferrite magnetic disk
might be attached to the block. (Circles on other side faces are not
shown.)
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208198
blocks, and the number of blocks in the chain. Thus achain of 12 pentagonal blocks (‘‘pentagonal’’ refers tothe shape of the blocks’ lower surfaces) in an appro-priate configuration would give rise to a dodecahedron.
The polyhedral shell may be stabilized by introducingseveral reversible magnetic interactions between theblocks. These magnetic linkages are strong enough tohold the polyhedron together under gentle agitation, butweak enough to allow the chain to be unfolded by hand,or under more vigorous agitation. Most importantly, theunfolded chain can be prompted, guided by themagnetic interactions, to fold into the polyhedron whenagitated. Furthermore, side chains may be added to theblocks such that the side chains come together duringthe folding process to form a second, smaller poly-hedron inside the first, and one that stabilizes the outerpolyhedral shell. This internal polyhedron can bethought of as the hydrophobic core of a globularprotein.
We describe the construction of several such poly-hedral models. The models contain 11–26 blocks and arebased on two regular Platonic polyhedra—a dodecahe-dron and an icosahedron—and two semi-regular (Ar-chimedean) polyhedra—a truncated octahedron and atruncated icosahedron (see Cundy and Rollett, 1961, fora detailed description of polyhedra). We show that themodels do in fact act and fold like globular proteins; wealso highlight another possible application they mayhave as the forerunners of new types of self-assemblingmicroscopic devices.
2. Materials and methods
Nets for the main chain and side chain blocks weredrawn with the Adobe Illustrator software package(version 5.5) and printed on A4 Copyprint Plus paper(80 g/m2) or thin card (176 g/m2). Nets for constructingthe models may be obtained from JPLC. The nets werecut out, scored, and then glued together (Pritt Stick,Henkel Consumer Adhesives) to produce the three-dimensional main chain block or a side chain subunit.
Blocks and side chain subunits are referred tothroughout the text according to the shape of theirlower surfaces (Fig. 1). Thus the blocks making up themain chain of the dodecahedral model are pentagonalblocks, and the subunits making up this model’s sidechains are pentagonal subunits.
The angle y between the side and lower edges of ablock (Fig. 1) was calculated knowing the (lower)edge:inter-radius ratio ðRÞ of the polyhedron of whichthe block is part (the inter-radius is the distance from thecenter of the polyhedron to the mid-point of one of itsedges), since, given the relationships shown in Fig. 1, itcan be shown that tan y ¼ 2=R: R values were obtainedfrom Cundy and Rollett (1961). Values for R and y are
given in Table 1, together with the other blockdimensions.
Side chain-bearing blocks (Fig. 2) were assembled bygluing together the sides of four pentagonal subunitsand then gluing the composite side chain to the topsurface of a main chain block either directly (as in thecase of the dodecahedral model) or indirectly, via afoam spacer (as in the case of the truncated icosahedralmodel). Foam spacers were cut from cylindrical earplugs(Safir) with a razor blade.
Main chain blocks were linked to each other by stripsof Scotch Magic tape (70 g/m2). The tape was stuck tothe lower surface of each block to leave the lower edgesof the connected blocks parallel and separated by a gapof about 1mm (Fig. 3a). The magnetic linkage betweentwo blocks (Figs. 3b and 4) was constructed as follows.Two circular magnetic disks (diameter 6mm) werepressed from adhesive-backed barium ferrite magnetictape (Farnell, UK, order code 723-9166) with a hole-punch. The disks were predominantly stuck, using thecircles as guides (Fig. 1), to the inside surface of theinteracting blocks, one disk on each surface. In onlyone model (dodecahedral model Ib, Fig. 5) were disksstuck to the outer surface of a block. A cylindricalneodymium–iron–boron (NdFeB) magnet (width 6mm,height 2mm, Farnell, UK, order code 723-0370) wasattached to one of these disks. The permanent magneticdipole of this magnet is coincident with the long axis ofthe cylinder. In every magnetic linkage, the NdFeB

ARTICLE IN PRESS
Table 1
Dimensions of models’ component blocks
Model Lower edge (cm) Side edge (cm) Ra y (�)
Dodecahedron (pentagonal block)b 3 2 0.764 69.1
Truncated octahedron (square and hexagonal blocks)b 2 1.6 0.667 71.6
Icosahedron (triangular block)b 4 2 1.236 58.3
Truncated icosahedron (pentagonal and hexagonal blocks)b 1.5 1 0.412 78.4
Side chain subunit (pentagonal)b,c 1.6 1 0.764 69.1
aR=Inter-radius of polyhedron (Cundy and Rollett, 1961).bGeometrical descriptors in brackets refer to lower face of block.cHeight of cylindrical foam spacer for truncated icosahedron=0.8 cm.
Fig. 3. Permanent and reversible linkages in the models. (a) Permanent
tape connection between lower faces of two pentagonal main chain
blocks; (b) Cut-away of two permanently connected blocks showing
reversible magnetic linkage between them.
Fig. 4. A magnetic linkage (circled) connecting two blocks in the
icosahedral model. Both halves of the magnetic linkage can be seen.
Upper half: NdFeB magnet mounted on magnetic disk; lower half:
magnetic disk (black).
Fig. 2. The side chain-bearing block. This block, which is part of a
dodecahedral model, comprises four small pentagonal subunits
(shaded) which make up the side chain itself, and a main chain block
(shown in white). In the truncated icosahedral model, a foam spacer
links the side chain to the main chain block, and the main chain block
is rotated by 36�, such that one of its side edges would be pointing
directly at the viewer, with the side chain remaining where it is.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208 199
magnet was always placed in the same orientation, withthe north-seeking pole exposed and the magnet on theinner surface of the block. Positioning of the magneticinteractions in the models is shown in Fig. 5 (boxes).The polyhedral models themselves are shown in Fig. 6.
Skeletal dodecahedral models containing only sidechain-bearing blocks were also made. In these models,card pentagons replaced blocks linking the side chain-bearing blocks, and the termini beyond the outermostside chain-bearing blocks were omitted (Fig. 7).
The stability and folding of the polyhedral modelswere investigated in a rotating plastic vessel, whichimparts conformational mobility to the polyhedralchain. (In the case of a polypeptide chain this mobilitywould come from energy acquired from its environ-ment—e.g. collisions with other molecules.) The vesselwas lined with bubble wrap to prevent the modelsbecoming unduly damaged during agitation. The roughsurface of the bubble wrap also helps to agitate themodel—without it the model slides over the plasticsurface of the vessel and is not properly agitated.

ARTICLE IN PRESS
Fig. 5. Schematic showing the block configurations of the models, viewed from the blocks’ upper surfaces. Filled rectangles: magnetic disks; clear
rectangles: NdFeB magnets mounted on magnetic disks. Four configurations (I–IV) are shown for the dodecahedral models. Configuration Ib lacks
block 1 of configuration Ia, and possesses three side chains, but is otherwise the same as Ia. The truncated octahedral model lacks the first block
(indicated by dashed line). Shaded pentagons represent either main chain blocks bearing side chains (dodecahedral model Ib and the truncated
icosahedral model), or main chain blocks to which side chains were added for a manual folding experiment (dodecahedral model II), or main chain
blocks which bore side chains in a skeletal version of that particular model (dodecahedral models Ib and III). Arrowheads within or near the shaded
pentagons indicate the orientation of the side chain: looking along the arrowhead (point facing away) will give the same view of the side chain as seen
in Fig. 2. Boxes show the magnetic interactions (short-, medium- and long-range) present in the model. Dodecahedral model Ib and the truncated
icosahedral model contain 6 and 17 other magnetic disks, respectively, that are not involved in any magnetic linkages. These models also contain
three side chain/side chain magnetic interactions.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208200
Ferrous materials were not used in constructing thevessel as they would have attracted the model’s magneticcomponents. The vessel was rotated by means of amotor connected to a brass axle fixed to the base of thevessel. Rotation rates varied between 0.75 and 1revolution(s) per second. At faster rates, the chainstended not to fold into stable polyhedra, as anyemerging structure was quickly destroyed by the rapidagitation. The vessel was held at a slight angle, such thatits lowest side was approximately 5� to the horizontal.This shallow angle prevented the models from fallingout of the vessel as it rotated, whilst at the same timeencouraging the chain to compact (rotation of the vesselalso encouraged compaction), an effect which tended toassist the folding process. At greater angles, however,the unfolded chain tended to become flattened againstthe base of the bucket, preventing folding.
The stability of the folded polyhedral models wasmonitored over 30 s by rolling them on to the lowersurface of the rotating vessel and then beginning timing.To initiate folding, the unfolded chain (all magneticinteractions broken) was held in the middle of the chainand then dropped on to the rotating lower surface of thevessel.
Stability/folding experiments were recorded on VHSvideotape with a Philips Explorer VKR 6850 videocamera with shutter speed set to 1
500s; and white
balance set to ‘‘Auto’’. The fast shutter speed consider-ably improved picture quality, allowing frame-by-frameanalysis of the video footage. The camera was mountedon a tripod and pointed at the mouth of the plasticvessel. For each model, one stability experimentand five folding experiments were recorded onvideotape.

ARTICLE IN PRESS
Fig. 6. The folded polyhedral models: (a) Dodecahedral model with three side chains (model Ib). The model lacks a twelfth block, and consequently
one pentagonal face of the internal dodecahedron (made up of three side chains) can be seen; (b) Truncated octahedral model. Block 1 was omitted in
the stability/folding experiments; (c) Icosahedral model; (d) Truncated icosahedral model with three side chains. One of the orange foam spacers
connecting a side chain to a main chain block can be clearly seen in the upper part of the model. Scale bars: 1 cm.
Fig. 7. Skeletal model. The model shown is a skeletal model of
dodecahedral model Ib, comprising side chain-bearing block 4 (to the
right) linked to side chain-bearing blocks 6 and 7 by a card pentagon.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208 201
The integrity of the magnetic interactions was checkedboth before and after each stability/folding experiment,as occasionally the NdFeB magnets were dislodged fromtheir disk mounting.
Blocks were numbered on their outer lower surfaces,and sometimes on their inner lower surfaces, so that thebehavior of individual blocks could be monitored duringthe stability/folding experiments.
For convenience, a model’s correctly folded three-dimensional structure is referred to in the text as itsnative structure, in line with protein nomenclature.
3. Results
3.1. Model design
3.1.1. The blocks
The folding polyhedral models comprise blocks linkedtogether in a chain. The upper and lower surfaces of theblocks are the same shape as the faces of the polyhedronbeing formed. For example, these surfaces are pentago-nal for a dodecahedral model, square or hexagonal forthe truncated octahedral model, and so on. The sides ofthe blocks are bevelled such that, when they cometogether, the angle between their adjacent lower surfacescorresponds to the dihedral angle of the polyhedronbeing formed. Bevelling the sides to achieve this state fora given block can be done by setting the angle y (Fig. 1)according to Table 1. The blocks are also hollow, andunless capped by a side chain, they lack an uppersurface. This facilitated the addition and removal ofNdFeB magnets to the magnetic disks, and repair of themodels when occasionally a NdFeB magnet wasdislodged from its disk mounting.
3.1.2. Material used to construct the blocks
Paper was chosen as the material for constructing theblocks because it is very light, yet the resulting blocksare stiff and robust. The magnetic interactions were notstrong enough to hold together blocks made of a heaviermaterial such as card. For instance, a card dodecahedralmodel (configuration Ia), weighing 1.5 times as much asthe paper version of this model (Table 2), was unstable:

ARTICLE IN PRESS
Table 2
Masses of models and their component blocks
Modela Mass of glued blockb (g) Number of magnetic linkages Mass of modelc (g)
Dodecahedron (configurations Ia, II–IV) 0.34 8 8.6–9.0
Dodecahedron (configuration Ia, card) 0.75 8 13.7
Dodecahedron (configuration Ib) 0.35d, 0.90e 6f 9.5g
Truncated octahedron 0.13h, 0.23i 7 6.6
Icosahedron 0.20 7 8.5
Truncated icosahedron 0.11j, 0.13i, 0.69e 8f 10.8k
aAll models are made of paper unless otherwise stated.bMasses represent averages of all the blocks in the chain, or, in the case of the dodecahedral models Ia and II–IV, all the blocks in these models.c Including tape and magnetic linkages. Mass of tape linkage: 0.04 g (dodecahedral and icosahedral models), 0.02 g (truncated octahedral model),
0.01 g (truncated icosahedral model). Mass of magnetic linkage (comprising two barium ferrite magnetic disks and a NdFeB magnet)=0.56 g.dMain chain block.eSide chain-bearing block.fThis number includes three side chain/side chain magnetic linkages.gThis figure includes six barium ferrite magnetic disks not involved in any magnetic linkages.hSquare block.iHexagonal block.jPentagonal block.kThis figure includes 17 barium ferrite magnetic disks not involved in any magnetic linkages.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208202
it collapsed when rolled into the rotating vessel.Furthermore, its unfolded chain did not form anylasting elements of native structure when agitated. Acard dodecahedral core comprising three isolated sidechains was also extremely unstable (it could not even bemanually assembled into a stable structure). In short,the folding and stability of the models are criticallydependent on the balance between the strength of themagnetic interactions and the weight of the materialused to construct the blocks.
All the models made in this work weighed between 5and 15 g (Table 2).
3.1.3. Permanent and reversible linkages in the models
The blocks were connected to each other by a tapelinkage (Fig. 3a) that remained securely attached to theblocks throughout the folding experiments, and did nottear during these experiments. It was therefore apermanent linkage. The tape was attached to the blockssuch that there was a small gap of about 1mm betweenthem. This gap allows the blocks to be bent towardseach other; the sides meet such that they are flush(Fig. 3b).
The tape linkage restricts the movement of adjacentblocks to two dimensions (relative to each other). Thustogether the tape linkages greatly reduce the conforma-tional space available to the chain. The stiffness of thetape favors a situation in which the lower faces ofadjacent blocks are held at 180� (although videotapeanalysis of the folding chains showed that the tapebetween adjacent blocks could be bent to its extreme,such that the lower faces of these two blocks touch).
The other type of linkage included in the models is amagnetic one (Fig. 3b), reversibly connecting the side of
one block to another block. The magnetic linkagecomprises two barium ferrite magnetic disks, one oneach interacting surface, and a NdFeB magnet mountedon one of these disks. The components of the magneticlinkage were usually placed on the inside of eachinteracting surface, thus affording better contact be-tween the surfaces. In dodecahedral model Ib, however,the disks of a magnetic linkage were placed on the outersurfaces of the interacting blocks, and the NdFeBmagnet was placed on the inner surface (Fig. 5). In thisinstance, there was a small gap between the twointeracting surfaces in the folded model.
NdFeB magnets were chosen because they are lightand strong. Other magnets, such as the ceramic ferritedisks used in fridge magnets, were not suitable becausethey are too large and too heavy; they are also not asstrong as the NdFeB magnets.
The magnetic linkage is only formed when the twointeracting surfaces are in close proximity (B0.5 cm).(The force between the two components of the magneticlinkage is inversely proportional to the square of thedistance between them (Duckworth, 1961)). Onceformed, the linkage is easily broken, either by gentlypulling the blocks apart by hand, or as the result of acollision, during the course of a folding experiment forexample. Thus the magnetic linkage is reversible. In theabsence of the NdFeB magnet, the interacting surfacesfall apart, as the attraction between the two bariumferrite magnetic disks on their own is extremely weak.On the other hand, a magnetic linkage between twoNdFeB magnets is too strong, and is effectivelyirreversible.
Magnetic linkages were placed either between blocksinvolved in sharp turns in the chain (short-range

ARTICLE IN PRESS
Fig. 8. Complementary surfaces of side chains making up internal core
of a dodecahedral model. One side chain (to the right) is about to make
contact with a complementary cavity in the partial core formed by the
other two side chains (to the left of the zigzag cleft).
Fig. 9. Extended dodecahedral chain with side chains (model Ib). Just
visible in the side chain on the left-hand side is a NdFeB magnet
involved in a side chain/side chain magnetic interaction.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208 203
interactions) or between two blocks separated by severalintervening blocks (medium- and long-range interac-tions) (Fig. 5).
The positioning of magnetic linkages within the chainwas somewhat arbitrary; however, an attempt was madeto distribute the NdFeB magnets uniformly throughoutthe chain, and also to have about three long-range mainchain/main chain magnetic interactions per model(Fig. 5). Each model contains 6–8 magnetic interactions(including side chain/side chain magnetic interactions),giving a reasonably stable structure. Fewer interactionsare possible, however. For example, the dodecahedralmodel in configuration III was able to fold into amarginally stable three-dimensional structure with justthree magnetic interactions present (of a possible 19).
3.1.4. The number of blocks in a chain
The type of polyhedron being formed determines themaximum number of blocks in a chain. For example, adodecahedral model allows a maximum number of 12blocks in the chain, a truncated octahedral model 14blocks, and so on. In other words, a large chain requiresa polyhedron with a large number of faces. Of course,smaller chains can be made by simply omitting blocks,giving a partial polyhedron.
We made 11- and 12-block dodecahedral models, a13-block truncated octahedral model, a 20-block icosa-hedral model and a 26-block truncated icosahedralmodel (Fig. 6). The 11-block dodecahedral model(model Ib) and the 26-block truncated icosahedralmodel contain three side chains each.
3.1.5. The blocks in a chain can be arranged in many
configurations
Although there are a large number of differentconfigurations for the chain of blocks making up aparticular polyhedral model (calculating precisely howmany is a difficult, and quite possibly intractable,problem (G.C. Smith, personal communication)), welooked at only four dodecahedral configurations andone configuration each for the other polyhedral models(Fig. 5). It should be noted that there are a large numberof block configurations that do not fold into the correctpolyhedral structure.
3.1.6. Inclusion of side chains to form an internal
polyhedral core
Some blocks in the main chain can also be equippedwith side chains, the complementary surfaces of whichmeet to form a second polyhedron inside the first. Forour experiments, we chose a dodecahedron to be theinternal polyhedron.
The side chains were derived from this dodecahedronby dividing it into three identical segments (one of thesesegments is shown shaded in Fig. 2). Each of thesesegments is a composite of four hollow pentagonal
subunits. Each side chain has a surface complementaryto portions of the other two side chains (Fig. 8). Twointeracting side chains are held together by a single,reversible magnetic linkage (one part of which can beseen in Fig. 9). Therefore, there are three magneticlinkages in total holding the core dodecahedrontogether. There are no magnetic linkages between sidechains and main chain blocks.
In the dodecahedral model, the core dodecahedronpacks directly against the folded main chain; this is notpossible in the case of the truncated icosahedral model,and for this reason a spacer had to be included betweenthe side chain and main chain block to which the sidechain was being attached (Fig. 6d). Because thepentagonal faces of a truncated icosahedron are ar-ranged in a dodecahedral fashion (i.e., if imploded they

ARTICLE IN PRESSB.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208204
would form a dodecahedron), the side chains can beattached to certain pentagonal main chain blocks suchthat when these side chains come together, they form aregular dodecahedron.
An unfolded dodecahedral chain with side chain-bearing blocks is shown in Fig. 9.
3.1.7. Special measures taken to permit folding of the
truncated octahedral model
Several additional measures were taken to enable thetruncated octahedron model to fold. First, the blockswere given relatively deep sides. Second, whereverpossible, magnets were placed on the steeper sidedsquare blocks. Both measures prevented a magnet onone block being detached from its disk mounting by amagnet on another block. The third measure was toremove block 1 from the chain (Fig. 5). The steep sidesof this square block made it awkward for it to fit into theotherwise complete truncated octahedron, thus inhibit-ing the folding process. Although block 14 behaved in asimilar manner, it was only necessary to remove block 1to allow the chain to fold properly.
3.1.8. Skeletal dodecahedral models
Skeletal models (Fig. 7) are very easy to assemble anddisassemble, and only one set of side chain-bearingblocks is required to make a variety of different models.This allowed different side chain arrangements, indifferent chain configurations, to be explored usingone set of side chain-bearing blocks, rather thanlaboriously constructing separate models for eachparticular arrangement/configuration.
3.1.9. Errors in the construction of the models
Small errors that arose when constructing the blocks(cutting out the nets, gluing the blocks) and linking themtogether meant that the blocks did not fit togetherperfectly in the folded models. These errors influencedthe folding and stability of the models. For example, wemade three dodecahedral models in configuration III(Fig. 5), but they were not all as stable as each other,and the range of folding times differed considerablyfrom model to model. Although these errors did notseriously interfere with folding of the models’ mainchains, it did result in some sets of side chains not fittingtogether properly. Fortunately, the use of skeletalmodels (above) meant that only one set of side chainswas needed.
3.2. Stability and folding experiments
All the models apart from the truncated icosahedronwere tested in the rotating vessel for stability andfolding. The truncated icosahedral model was not testedin the rotating vessel for either type of behavior becauseof its fragility.
3.2.1. Stability experiments
The folded polyhedral models tested showed variousdegrees of stability (Table 3). A card version ofdodecahedral model Ia unfolded on contact with therotating vessel, and remained unfolded for the rest of theexperiment, although elements of native structure wereoccasionally observed. The other models tested gener-ally maintained their three-dimensional polyhedralstructure over the course of the experiment. They diddisplay some structural instability, including the appear-ance of cracks between two sections of the folded chainconnected by long-range interactions, and loose termini(1 to 3 consecutive blocks). These features weretemporary, however. The truncated octahedral modelalso partially unfolded and then refolded.
The largest model made in this work, in terms of thenumber of blocks in the main chain, was a partialtruncated icosahedron, comprising 26 blocks. Althoughthe stability of the folded version of this model was nottested in the rotating vessel, it was investigated by gentlyrolling it around. The folded version of this modellacking side chains was quite stable, partially unfoldingand then refolding. However, the stability of thetruncated icosahedral model was further improved byadding side chains able to form a dodecahedral core.Only three side chain/side chain and five main chain/main chain interactions were required to form theimproved truncated icosahedral model. The structure ofthis model is quite irregular, however (Fig. 6d).
3.2.2. Folding experiments
All the models tested folded; folding times variedfrom 3 s to just over a minute; for a particular model, thefolding time also varied (Table 3). Only in the skeletalversion of dodecahedral model III was misfoldingobserved. Otherwise the folding chains did not gettrapped in a non-native state.
Detailed analysis of the video footage of the modelsfolding indicated that folding proceeded via elements ofnative structure formed by short- and medium-rangeinteractions (Fig. 5). These elements were often short-lived, and several sets of elements could be formed anddestroyed before the chain eventually folded. Thefolding times quoted in Table 3 include these initial,unproductive folding events.
A given model did not fold via one clearly definedpathway; rather, it folded via several different pathways.For instance, dodecahedral model IV folded viaelements of native structure formed at either end ofthe chain, or in the middle. Similarly dodecahedralmodel II folded via 4- or 6-block elements of nativestructure at either end of the chain, with the structuregradually spreading from the nucleation site; likewise,the folding of the icosahedral model could be initiatedby a pentameric cluster of blocks at either the beginning(blocks 1–5) or towards the middle (blocks 11–15) of the

ARTICLE IN PRESS
Table 3
Stability and folding times of models
Modela Stability Dynamic behavior Folding time (s)b
Dodecahedron (configuration Ia) Quite stable Cracks, loose termini 15–23
Dodecahedron (configuration Ia, card) Very unstable Unfolds —
Dodecahedron (configuration Ib) Quite stable Cracks, loose termini 7–24
Dodecahedron (configuration II) Stable Cracks 14–50
Dodecahedron (configuration III)c Stable Cracks 3–11
Dodecahedron (configuration IV) Quite stable Cracks, loose termini 5–19
Truncated octahedron Marginally stable Cracks, loose termini, unfolds/refolds 3–29
Icosahedron Stable Cracks 7–68
aAll models are made of paper unless otherwise stated.bRanges quoted are for five experiments.cThree models were made in this configuration. Data are only given for the most stable model, which was also the model with the fastest folding
times.
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208 205
chain. However, these pathways were not clearlydefined—there was considerable variety in the waystructured units/blocks interacted in a given pathway.
One interesting structural feature seen in the foldingexperiments with the dodecahedral models—and whichwas also apparent in the stability experiments—was aloose, slightly squashed, globular form. Usually thisform appeared during the series of productive foldingevents, but it could also appear during the initial non-productive folding stages. It was also apparent in a‘‘folding’’ experiment in which the NdFeB magnets wereremoved from dodecahedral model Ia, i.e. in the absenceof any significant magnetic interactions.
Folding of a dodecahedral main chain with sidechains (model Ib, Fig. 5) occurred on the same time-scale as the dodecahedral models without side chains(model Ib was slightly heavier than these other models,but had two fewer magnetic interactions (Table 2)).However, formation of the internal dodecahedral coreby the three side chains was rapid (4–9 s). (Formation ofthe dodecahedral core in the skeletal version of thismodel (Fig. 7) was even faster (2–4 s)). In particular, thetwo side chains on the adjacent main chain blocks 6 and7 formed a persistent interaction almost immediately(1-4 s). Closely spaced side chains therefore appear toassist folding. Widely spaced side chains, on the otherhand, are unlikely to assist folding. For example, twowidely spaced side chains in a skeletal version ofdodecahedral model III (Fig. 5) became locked togetherin an undesirable conformation during folding. Also,manually folding a side chain-bearing dodecahedralchain in configuration II (Fig. 5) was an awkwardprocess.
Even when its side chains lacked magnetic interac-tions (leaving three long-range main chain/main chaininteractions), dodecahedral model Ib could still fold, onthe same time-scale as the other dodecahedral models,into a marginally stable dodecahedral structure. Incontrast, the folded structure of dodecahedral model Iawith only its three long-range main chain/main chain
interactions was unstable. This suggests that thecomplementary, steric interactions between side chainslacking NdFeB magnets also contribute favorably to thefolding process and the stability of the final structure.
In one experiment, three isolated side chains wereagitated in the rotating vessel on their own, to see if theywould still associate correctly (they could be puttogether by hand into a stable core dodecahedron).Almost immediately, a NdFeB magnet from one sidechain was pulled away from its disk mounting by aNdFeB magnet in another side chain, locking the twoside chains together in a misaligned fashion. Since thistype of event does not occur when the side chains areattached to the main chain in model Ib, the configura-tion in which they are held by this chain must protect theside chains from such inappropriate interactions.
Although the truncated icosahedral model with sidechains was too fragile to use in a proper foldingexperiment, the unfolded chain could be successfullyfolded by gently rolling it from a flat, extended state inwhich all three side chains were directed upwards. It wasnoticeable that, as with dodecahedral model Ib, the twoside chains lying closest to each other (on blocks 16 and20) formed a stable interaction about which the rest ofthe structure developed. Also noticeable was theformation of a short-range magnetic interaction withoutthe model being agitated. This interaction occurs at asharp turn in the chain, between blocks 15 and 17(Fig. 5).
4. Discussion
We have developed several dynamic mechanicalmodels that will, when agitated, fold into uniquepolyhedral shapes. The models display a number ofstructural similarities to globular proteins (Branden andTooze, 1999):
(1) They are made up of a chain of blocks, just as apolypeptide chain is made up of amino acids. Like

ARTICLE IN PRESS
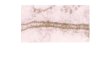
Fig. 10. Model of truncated icosahedron showing a path tracing out a
b-hairpin-like feature (the diagonal loop).
B.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208206
amino acids, these blocks can possess side chains.Complementary side chains come together to form aninner core, in much the same way that hydrophobicamino acid side chains cluster together to form acompact hydrophobic core in a globular protein.
(2) The blocks are held together by permanent (tape)linkages and reversible (magnetic) interactions, similarto covalent and non-covalent bonds, respectively, inproteins. The flexible tape linkage restricts the move-ment of adjacent blocks thereby greatly reducing theconformational space available to the chain, in the sameway that amino acid side chains and peptide bondsrestrict the conformational space of a polypeptide chain.The magnetic interactions are effective only over shortranges, as are non-covalent interactions. Specifically, inboth types of interaction the force between the inter-acting components is inversely proportional to theirseparation (r) raised to the power n, where n is a smallnumber: the force between the magnetic components isproportional to 1/r2 (Duckworth, 1961), whereas, forexample, London interactions are proportional to 1/r6
(Atkins, 1998). Several weak magnetic interactions actcooperatively within a model to lend it stability.
(3) The models fold into a unique three-dimensionalshape, and the instructions for producing this shape areinherently built into the sequence (configuration) ofblocks making up the main chain.
(4) It is difficult to predict the structure of the modelbeing formed from a two-dimensional plan of it (Fig. 5),just as it is difficult to predict the tertiary structure of aprotein from its primary structure. Rather interestingly,many different block configurations give rise to the samethree-dimensional structure, i.e. the same polyhedron, inmuch the same way that different amino acid sequencesgive rise to the same protein fold.
(5) Structural elements similar to reverse turns andb-sheets may be discerned in the folded models (Fig. 10).
(6) A variety of cavities can be made in the polyhedralsurface of a model by omitting one or more blocks fromthe main chain, and such a cavity may be thought of asthe active site of an enzyme, or the site for a transactionwith another molecule.
(7) The folded models are conformationally mobile,with termini becoming loose and temporary cracksappearing in the structure. In a similar manner, proteinscan have flexible termini and also exhibit breathingmotions.
(8) Like proteins, the models show varying degrees ofstability, from the very unstable to the stable (Table 3).Even the stable models, however, will unfold if agitatedvigorously enough.
As well as being structurally similar to proteins, themodels fold in a manner very reminiscent of the proteinfolding process (Branden and Tooze, 1999; Daggett andFersht, 2003a,b). Thus the models fold quickly, in amatter of seconds, which is within the time-scale for
protein folding, although it is certainly not as fast as themicro- to millisecond folding times of small, singledomain proteins; folding proceeds via elements of nativestructure, including side chain clusters, and sometimesthese elements persist in the unfolded state; folding mayinitially involve one or more non-productive phases,where elements of native structure are formed and thendestroyed, consistent with the process having anactivation energy; folding of a given model can alsoproceed via several different pathways, although thesepathways are somewhat indistinct; finally, folding mayinvolve a loose oblate form, not dissimilar from amolten globule (this form probably arises from theinclination and rotation of the vessel in which thefolding is done).
Choosing a light but durable material such as paper toconstruct the models was critical to the success of theproject. On the scale we were building, a heaviermaterial like card gives rise to models that are unstableand therefore do not fold. This is not to say that card, orany other relatively light material, could not be used toconstruct folding polyhedral models, providing themagnetic interactions were strong enough to holdthe blocks together, but not so strong as to make theinteractions irreversible. Indeed, plastic models, madefrom a mold, are likely to be more accurate andtherefore better fitting than the paper models madehere, and would be better suited to comparative foldingexperiments (e.g. comparing different side chains, ordifferent side chain arrangements). Furthermore, plasticmodels could be mass-produced in kit form.
There may also be some benefit in miniaturizing themodels, to length scales of millimetres or less, where

ARTICLE IN PRESSB.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208 207
they might act as some form of container. Rather morefancifully, one might imagine a pentagonal cavity on thesurface of a miniature dodecahedron (say) acting as atemplate for the assembly of two halves of a pentagonalmain chain block. This would enable the dodecahedronto replicate itself, given an appropriate template, whichcould even be the dodecahedron chain itself. In otherwords, the miniature dodecahedron could act as a self-replicating, enzyme-like entity. In connection with thesepossibilities, Love et al. (2003) have recently madesub-micron diameter metallic rods that self-assemblethrough magnetic interactions. The models describedhere, if miniaturized, would offer this type of system anadded degree of sophistication, assuming that theblocks, and in particular their links, could be fabricatedat this scale.
It should be noted that polyhedra have been usedbefore as the basis for three-dimensional models ofbiological molecules: Caspar and Klug (1962) describedpolyhedral models for the structure of spherical viruses,and Murzin and Finkelstein (1988) modelled a-helixpacking in globular proteins by placing the helices onthe edges of a polyhedron. It is important, however, todistinguish the models presented here from these twoearlier models. In the latter models, the faces (Caspar/Klug) or edges (Murzin/Finkelstein) of the polyhedronrepresent protein subunits and a-helices, respectively. Inthe models presented here, on the other hand, each faceof the polyhedron represents an amino acid in apolypeptide chain. Another important difference be-tween the two earlier models and the models presentedhere is that the former are static, whereas the currentmodels are dynamic.
It should also be noted that Boncheva et al. (2002)have produced a self-assembling chain like the onesdescribed here. However, the Boncheva et al. chainassembles through formation of permanent interactions,leading to two static, prism-like polyhedra; fabricationof the chain’s components relies on specialist (andexpensive) techniques, and the assembly process requiresa hot, aqueous solution. In contrast, the chainsdescribed here assemble through simple, reversibleinteractions, giving dynamic, regular or semi-regularpolyhedra; their component blocks are simple toconstruct, and the chains do not need to be immersedin a liquid for assembly to occur. Despite thesedifferences, certain features of each system are com-plementary. For example, side chain-like elements couldbe incorporated into the Boncheva et al. chain, andlight-emitting diodes that become illuminated whensingle or multiple interactions are formed (to helpmonitor the folding process, for example) could beincorporated into the chains described here.
Another model of relevance to our own models is awooden toy consisting of 64 cubes (side B1 cm)connected by a rubber string that runs through each
cube once. The chain of cubes can be manuallyassembled into a large cube comprising 4 cubes perside. This toy, whose existence was pointed out to us byan anonymous reviewer, is a macroscopic representationof the lattice models of protein folding ( &Sali et al., 1994;Yue et al., 1995). However, the chain of cubes does notfold up by itself, and manually assembling the unfoldedchain into the large cube is difficult.
As protein analogues, the models presented here havesome obvious limitations:
(1) The main chain blocks are abstract representationsof amino acids, having no chirality or direction.
(2) Although b-sheet and reverse turn-like features areapparent in the models, helical features are not.
(3) The models do not take the hydrophobic effectinto account—in other words, the medium in which themodels are in (air) does not drive the side chains (ifpresent) together, although the vessel in which thefolding is done is inclined at a slight angle so as toencourage the chain to cluster; rotating this vessel alsoencourages compaction.
(4) The tape linkage between blocks only allowsadjacent blocks to move in two dimensions relative toone another, placing severe conformational restraints onthe chain. This means that distant parts of the chain mayonly contact each other after the formation of theappropriate short- and medium-range interactions.Hence the elements of native structure seen in thefolding models are confined to continuous sections ofthe chain. On the other hand, one beneficial conse-quence of the conformational restraints placed on thechain by the tape linkages is that only thedesired interactions (Fig. 5) are formed; the chainconformation prevents undesirable interactions.Furthermore, it is quite clear that the conformationalrestraints can predispose the chain towards a native-likestructure. But in any case, the models could be made tobe more protein-like by having rotating links betweenblocks.
(5) All the magnetic interactions are roughly the samestrength, unlike non-covalent interactions in proteins.With models made to a high degree of precision, it maybe possible to vary the strength of the magneticinteractions, for example by using several small NdFeBmagnets of different strength.
(6) The chains constructed in this work contain 11–26blocks, and are therefore shorter than the polypeptidechains of even the smallest globular proteins, which areabout 50 amino acids long. Making larger models oughtto be possible, although longer chains will also give riseto larger internal cavities, and these may be harder topack with side chains than the cavity of a smallerpolyhedron.
(7) Although the positioning of magnetic linkages inthe model chains was not completely random—weendeavoured to distribute the magnets uniformly along

ARTICLE IN PRESSB.T. Burnley, J.P.L. Cox / Journal of Theoretical Biology 229 (2004) 197–208208
the chains and to have approximately three long-rangemain chain/main chain interactions per chain—it wasrather arbitrary. In future, the magnetic linkages couldbe positioned to give specific types of network, forexample small-world networks (Watts and Strogatz,1998). Proteins in fact exhibit small-world behavior(Greene and Higman, 2003).
Nevertheless, these are the first dynamic models of afolding polymeric chain, and they should be useful aidsfor investigating the protein-folding process, as well asperforming an educational role. With regard to the formerpoint, the easy-to-construct skeletal models describedhere, comprising side chain-bearing blocks linked by flatpolygons, could, because they are quick to put togetherand take apart, be very useful in identifying the patternsof key residues that determine the fold a particularpolypeptide chain adopts, a process likely to be importantin predicting the three-dimensional structure ofa globular protein from its amino acid sequence(Dobson, 2003).
Acknowledgements
We would like to thank Alan Cox and Matthew Pack forphotography, Steve Routley for helping to build therotating vessel, Dek Woolfson, Ian Williams, Geoff Smithand Alexey Murzin for valuable discussions, and theUniversity of Bath for financial support. We wouldalso like to thank an anonymous reviewer for helpfulcomments.
References
Atkins, P.W., 1998. Physical Chemistry, Oxford University Press,
Oxford, p. 665.
Boncheva, M., Gracias, D.H., Jacobs, H.O., Whitesides, G.M., 2002.
Biomimetic self-assembly of a functional asymmetrical electronic
device. Proc. Natl. Acad. Sci. USA 99, 4937–4940.
Branden, C., Tooze, J., 1999. Introduction to Protein Structure.
Garland, New York.
Caspar, D.L.D., Klug, A., 1962. Physical principles in the construction
of regular viruses. Cold Spring Harbor Symp. Quant. Biol. 27, 1–24.
Cundy, H.M., Rollett, A.P., 1961. Mathematical Models, Oxford
University Press, Oxford, pp. 76–160.
Daggett, V., Fersht, A.R., 2003a. Is there a unifying mechanism for
protein folding? Trends Biochem. Sci. 28, 18–25.
Daggett, V., Fersht, A., 2003b. The present view of the mechanism of
protein folding. Nat. Rev. Mol. Cell Biol. 4, 497–502.
Dobson, C.M., 2003. Protein folding and misfolding. Nature 426,
884–890.
Duckworth, H.E., 1961. Electricity and Magnetism, Holt, Rinehart
and Winston, New York, pp. 255–257.
Greene, L.H., Higman, V.A., 2003. Uncovering network systems
within protein structures. J. Mol. Biol. 334, 781–791.
Love, J.C., Urbach, A.R., Prentiss, M.G., Whitesides, G.M., 2003.
Three-dimensional self-assembly of metallic rods with submicron
diameters using magnetic interactions. J. Am. Chem. Soc. 125,
12696–12697.
Murzin, A.G., Finkelstein, A.V., 1988. General architecture of the a-helical globule. J. Mol. Biol. 204, 749–769.
&Sali , A., Shakhnovich, E., Karplus, M., 1994. How does a protein
fold? Nature 369, 248–251.
Shea, J.-E., Brooks, C.L., 2001. From folding theories to folding
proteins: a review and assessment of simulation studies of protein
folding and unfolding. Ann. Rev. Phys. Chem. 52, 499–535.
Watts, D.J., Strogatz, S.H., 1998. Collective dynamics of ‘small-world’
networks. Nature 393, 440–442.
Yue, K., Fiebig, K.M., Thomas, P.D., Chan, H.S., Shakhnovich, E.I.,
Dill, K.A., 1995. A test of lattice protein folding algorithms. Proc.
Natl. Acad. Sci. USA 92, 325–329.