Embed Size (px)
Citation preview

1
Studies on a blasticidin S deaminase from Streptomyces lividans and its application in 1
engineering a blasticidin S producing strain for easy genetic manipulation 2
Li Li,1, 2† Jun Wu,1† Zixin Deng,1 T. Mark Zabriskie,3 and Xinyi He1* 3
State Key Laboratory of Microbial Metabolism, and School of Life Sciences & Biotechnology , 4
Shanghai Jiao Tong University, 1954 Huashan Road, Shanghai, 200030, China,1 and 5
Engineering Research Center of Industrial Microbiology (Ministry of Education), and College of 6
Life Sciences, Fujian Normal University, Fuzhou, Fujian, 350108, China,2 and Department of 7
Pharmaceutical Sciences, College of Pharmacy, Oregon State University, Corvallis, OR 8
97331-3507 (USA)3 9
10
* Corresponding author. 11
Mailing address: 12
State Key Laboratory of Microbial Metabolism and School of Life Sciences & Biotechnology, 13
Shanghai Jiao Tong University, 1954 Huashan Road, Shanghai, 200030, China. 14
Phone: 86 21 62932943-2131. Fax: 86 21 62932418. 15
E-mail: [email protected]. 16
† L. L. and J. W. contributed equally to the present study. 17
18
Copyright © 2013, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.03254-12 AEM Accepts, published online ahead of print on 1 February 2013
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2
19 Abstract 20 21 Blasticidin S is a peptidyl-nucleoside antibiotic produced by Streptomyces griseochromogenes that 22 exhibits strong fungicidal activity. To circumvent an effective DNA uptake barrier system in the 23 native producer and investigate its biosynthesis in vivo, the blasticidin S biosynthetic gene cluster 24 (bls) was engrafted to the chromosome of Streptomyces lividans. However, the resulting mutant 25 LL2 produced the inactive deaminohydroxyblasticidin S instead of blasticidin S. Subsequently, a 26 blasticidin S deaminase (SLBSD) was identified in S. lividans and shown to govern this in vivo 27 conversion. Purified SLBSD was found to be capable of transforming blasticidin S to 28 deaminohydroxyblasticidin S in vitro. It also catalyzed deamination of the cytosine moiety of 29 cytosylglucuronic acid, an intermediate in blasticidin S biosynthesis. Disruption of SLBSD in S. 30 lividans LL2 led to successful production of active blasticidin S in the resultant mutant, S. lividans 31 WJ2. To demonstrate the easy manipulation of the blasticidin S biosynthetic gene cluster, blsE, F 32 and L, encoding a predicted radical SAM protein, an unknown protein and a guanidino 33 methyltransferase, were individually inactivated to access their role in blasticidin S biosynthesis. 34 35 Introduction 36 37 Blasticidin S (Figure 1) belongs to the peptidyl nucleoside family of antibiotics. This compound is 38 produced by Streptomyces griseochromogenes and was first identified in 1958 (1) and was the 39 first antibiotic used to control rice blast in eastern Asia (2). The cytidine core of blasticidin S can 40
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

3
bind tightly to the corresponding guanine base at the P-site of the 50S subunit of the ribosome 41 through a Watson-Crick base pair(3), which makes it a potent inhibitor of protein synthesis in 42 both prokaryotic and eukaryotic cells (4, 5). There have been three kinds of blasticidin S 43 resistance genes reported to date: BSD from Aspergillus terreus (6), bsr from Bacillus cereus (7) 44 and blasticidin S-acetyltransferase gene from Streptoverticillum sp.(8). Both BSD and bsr encode 45 blasticidin S deaminase which can convert blasticidin S to the non-toxic 46 deaminohydroxyblasticidin S, the base moiety of which is converted to uridine and thus loses the 47 capability of binding to the ribosome. On the other hand, the blasticidin S-acetyltransferase gene 48 encodes an acetyltransferase which, along with acetyl-CoA, can block the inhibition of protein 49 synthesis by blasticidin S in a cell-free system (8). Because of these traits, blasticidin S is widely 50 used to select transformed cells that have been engineered to carry a resistance gene in biological 51 research(9) . 52 53 Puromycin (10), polyoxin (11), nikkomycin (12) and pacidamycin (13) are representative 54 examples of the peptidyl nucleoside family, the biosynthetic pathways of which have been well 55 studied (11, 14). Unlike the above mentioned typical five-membered-ring peptidyl nucleosides, 56 the structure of blasticidin S and mildiomycin (Figure 1) feature the glucuronic acid-derived 57 hexose that is coupled to cytosine or hydroxymethyl cytosine. The biosynthetic gene cluster of 58 blasticidin S was first reported in 1998 and heterologously expressed in Streptomyces lividans 59 (15). However, except for some putative intermediates, blasticidin S could not be identified from 60 the S. lividans heterologous production. Although the biosynthesis gene cluster for blasticidin S 61 was sequenced and analyzed (16), there were no in vivo studies of the biosynthetic pathway due 62
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

4
either to the difficult genetic manipulation system of S. griseochromogenes, or failure of 63 production of blasticidin S in S. lividans. Hence, the biosynthetic pathway of blasticidin S was 64 proposed largely on the basis of labeled precursor incorporation, biochemical studies and 65 bioinformatic analysis (17-20), and several steps in the pathway still remain cryptic. 66 67 In this study, we identified a blasticidin S deaminase in S. lividans that is able to transform the 68 cytotoxic blasticidin S to its inactive form, deaminohydroxyblasticidin S. The activity of SLBSD 69 was measured with several nucleosides in vitro. The blasticidin S biosynthetic gene cluster was 70 then engrafted into a S. lividans strain with SLBSD disrupted to generate mutant WJ2 that 71 produced blasticidin S. Furthermore, to specifically investigate the functions of blsE, F and L in 72 blasticidin S biosynthesisin vivo, these genes were individually disrupted and the fermentation 73 broths of the resultant mutants were analyzed by LC-MS. 74 75 Materials and Methods 76 77 Strains and Culture Conditions 78 Key strains and plasmids are listed in Table S1. S. lividans66 and its derivatives were grown at 79 30 °Con SFM agar plates for sporulation or in TSBY (10.3%) (TSB liquid medium that was 80 supplemented with sucrose (10.3% (w/v)) and yeast extract (1% (w/v)) for growth of mycelia. 81 Exconjugants were grown on SFM agar plates containing appropriate antibiotic (spectinomycin 82 (150µg/ml) or thiostrepton (15µg/ml)) or in TSBY (10.3%) with appropriate antibiotic 83 (spectinomycin (50µg/ml) or thiostrepton (5µg/ml)). For secondary metabolite analysis, seed 84
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

5
cultures were prepared by inoculating a loop of stock culture into 250ml baffled flask containing 85 25ml of seed medium (20g glucose, 30 g soybean flour, 5.0g yeast extraction, 1.0g (NH4)2SO4, 86 0.5g MgSO4·7H2O, 3.0g CaCO3, per 1L), and incubated in a rotary shaker for 40h at 30 °C, 200 rpm. 87 Then 5ml of this seed culture was transferred into a 500ml baffled flask containing 50ml 88 fermentation medium (80g glucose, 40g soybean flour, 1.0g (NH4)2SO4, 0.5g MgSO4·7H2O, 10.0g 89 CaCO3, 0.05g FeSO4, 0.4g K2HPO4 per 1L) and incubated another 6 days under the same 90 conditions. For LC-MS analysis of the production of blasticidin S and its intermediates, strains 91 were cultured in TSBY (10.3%) in baffled flasks at 30 °C and at 220 rpm for 6 days. 92 93 General Molecular Biology Methods 94 Isolation of total DNA, protoplast preparation and transformations were performed according to 95 Kieser et al. (21). Manipulations of E. coli strains were conducted according to Sambrook et al 96 (22). DNA fragments and PCR products were purified from agarose gels (0.8%) using the DNA Gel 97 Extraction Kit (V-gene Biotechnology Limited). PCR product for SLBSD expression was inserted 98 into pMD18-T vector (TaKaRa, Dalian, China) and verified by sequencing. For the Southern 99 hybridization experiments, all the DNA samples were cleaved with BamHI, separated on an 100 agarose gel (0.8%) and transferred onto Hybond-N+ nylon membrane (Amersham Biosciences). 101 BamHI digested-Fosmid 7D11 was labeled with radioactive α-[32P]-dCTP (Beijing Furui 102 Bio-engineering Co., Ltd) using Random Priming Kit (Roche) as the probe. The Southern blot 103 hybridization was carried out overnight at 65 °C followed by high stringency wash (0.1×SSC, 0.1% 104 (w/v) SDS) at the same temperature, and detected with a phosphorimager (Fujifilm). Restriction 105 enzymes, T4 DNA ligase, Taq polymerase and alkaline phosphatase were purchased from MBI 106
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

6
Fermentas. Blasticidin S was purchased from Sigma-Aldrich. Cytosylglucuronic acid was 107 purchased from Shanghai Violetpharm Chem. Tec. Co., Ltd. CMP was purchased from Sangon 108 Biotech (Shanghai) Co., Ltd. 109 110 Bioassay method 111 To measure the resistance level of S. lividans and its bsd knock-out mutant to blasticidin S, 112 serially diluted blasticidin S standard was added to an Oxford cup placed on SFM medium plates 113 that were pre-inoculated with spores of the S. lividans strains. For bioassays, the Streptomyces 114 strains were grown at 30 °C for 5 days on SFM plates for production of secondary metabolite, 115 agar patches were transferred to potato dextrose agar (PDA, diced potatoes 200 g, glucose 20 g 116 and agar 15 g in 1 L tap water) that contained Rhodotorula rubra AS2.166 (Table S1), a yeast that 117 is sensitive to blasticidin S. 118 119 LC-MS Analysis 120 The fermentation broth (25 ml) was harvested and adjusted to pH 5.0 and centrifuged at 12,000 g 121 for 5 min. The supernatant was applied to Supelclean LC-SCX SPE columns (500 mg/3 ml, 122 Supelco), washed with 2 ml water and then with 2 ml 0.5% NH4OH. The fraction eluting with 3% 123 NH4OH was filtered through a 2 μm membrane before injection. The LC-MS analysis was done 124 using an Agilent 1100 LC-MSD with a C18 column (TC-C18, 250 mm × 4.6 mm, Agilent). The 125 isocratic mobile phase was 10 mM trichloroaceticacid:MeCN, 82:18 (v/v) and the flow rate was 126 0.3 ml/min at room temperature. Elution was monitored with a photodiode array detector at 272 127 nm and electro spray ionization (ESI) MS analysis was carried out in the positive mode. 128
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

7
129 Q-TOF MS Analysis 130 The analysis was done using Agilient 6530 Q-TOF LC/MS with a C-18 column (zorbax 3.5 μm 131 SB-C18, 2.1 mm×150 mm, Agilent). The isocratic mobile phase was MeCN : Water with a gradient 132 from 5%-100% and the flow rate was 0.2 ml/min at room temperature. The ion source was ESI 133 and the analysis was carried out in the positive mode. 134 135 Sequencing and Bioinformatic Analysis 136 The genomic library of S. griseochromogeneswas constructed in the pCC1FOS™ vector according 137 to the provided protocol (EPICENTRE Biotechnologies). By chromosomal walking (detail not 138 shown), an 8780 bp DNA fragment immediately downstream of blsN was cloned, sequenced and 139 deposited under accession number of JX244070. DNA sequencing was done at Life Technologies 140 Corporation, Shanghai. Open reading frames were predicted using FramePlot 3.0 beta 141 (http://watson.nih.go.jp/~jun/cgi-bin/frameplot-3.0b.pl) and multiple sequence alignment was 142 performed with BioEdit 7.0. Similarity comparisons of nucleotide or amino acid sequences 143 against public databases were done using the BLAST program on the NCBI website 144 (http://ncbi.nlm.nih.gov/blast) (23). 145 146 Construction of the Recombinant Strains 147 To integrate the entire blasticidin S gene cluster into the genome of S. lividans, fosmid 7D11, 148 containing the published blasticidin S gene cluster as well as its downstream 8780 bp sequence, 149 was selected from the genomic library of the blasticidin S native producer S. griseochromogenes. 150 Fosmid 7D11 was then cut by XbaI and SpeI and precipitated with 70% isopropanol and 10% 3M 151
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

8
sodium acetate. The purified digestion products were ligated at the XbaI-SpeI site of 152 pJTU1289(24) to construct pJTU1780 by rapid screening of the transformants, which contained 153 the complete blasticidin S biosynthesis gene cluster. pJTU1780 was then digested with StuI and 154 ligated with an aadA cassette amplified by primers aadA-T with pIJ779 as template to replace 155 most of the insertion that containing the blasticidin S gene cluster, generating construct 156 pJTU1785. A 4.85 kb dispensable fragment covering the insertion site for genomic island SLG 157 (accession number: EF210454) in S. lividans HXY16 chromosome was amplified with primers 158 HXY16 F&R and inserted at the EcoRV site of pOJ260 to construct pJTU1528. A 0.25kb internal 159 region of the 4.85kb fragment was cut off by StuI, its flanking 1.6kb and 3.0kb fragments were 160 employed as two homologous arms for introduction of foreign DNA sequences. The insertion part 161 of pJTU1785 was excised with EcoRV and ligated with the StuI-cut pJTU1528 to obtain pJTU1786. 162 163 To construct the blsE, blsF and blsL knockout mutants, a 13kb fragment covering blsE-blsN, was 164 excised from pJTU1780 using BglII and ligated with pJTU412 (25) to construct pWJ5. pWJ5 was 165 then modified by module exchanges using ReDirect technology essentially as described 166 previously with the primers Tar-blsE, Tar-blsF and Tar-blsL in Table S2(26). The genes blsE, blsF 167 and blsL were deleted individually by recombinational exchange with an aadA cassette. The 168 unmethylated recombinant plasmids, prepared in E. coli ET12567, were then separately 169 introduced to S. lividans WJ2 by protoplast transformation. The resulting putative double 170 crossover strains were confirmed by PCR with the primers Con-blsE, Con-blsF and Con-blsL listed 171 in Table S2 in the supplemental material. 172 173 Expression and Purification of SLBSD 174 The SLBSD gene was amplified from the genomic DNA of S. lividans HXY16 by PCR with the 175 primers SLBSD F&R (Table S2, NdeI and EcoRI sites are underlined). PCR was carried out with 176
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

9
high fidelity DNA polymerase (KOD-Plus, TOYOBO) and products were gel purified, digested with 177 the appropriate enzymes, and cloned into the corresponding restriction sites of the pET28a(+) 178 vector (Novagen). The resulting plasmid, named pWJ4, was used to transform E. coli DH10B for 179 sequencing. For expression studies, the plasmid was introduced to E. coli BL21 (DE3)/pLysE 180 (Novagen) and grown over night at 37 °C in liquid LB containing chloramphenicol and kanamycin 181 (34 μg/ml and 50 μg/ml, respectively). The seed cultures (10 ml) were used to inoculate 1 L 182 production cultures of LB with the corresponding antibiotics. The cells were grown at 37 °C to an 183 optical density at 600nm (OD600) of 0.6 and then induced with 184 isopropyl-β-D-thiogalactopyranoside (1 mM). The culture was then grown for an additional 5 hrs 185 at 30 °C. After centrifugation, the cells were resuspended in 40 ml binding buffer (20 mM sodium 186 phosphate, 20 mM imidazole, and 0.5 M NaCl, pH 7.4) and lysed by sonication in an ice bath (10 × 187 60 sec at 15 watts with 60 sec pauses). After another round of centrifugation(10000×g, 1 hr), the 188 supernatant was applied to a HisTrap HP column (GE Healthcare), and purified using an AKTA 189 FPLC (GE Healthcare), by eluting with elution buffer (20 mM sodium phosphate, 500 mM 190 imidazole, and 0.5 M NaCl, pH 7.4) in linear gradient. The purified His-tagged SLBSD was desalted 191 using a HiTrap Desalting column (GE healthcare) and stored in 10 mMTris-HCl Buffer (pH 8.0) 192 with 20% glycerol at -80 °C. The purified His-tagged SLBSD was analyzed by 15% SDS-PAGE and 193 protein concentrations were determined using the Bradford Protein Assay Kit (Bio-Rad). 194 195 In vitro Assay of the Purified SLBSD 196 The assays of recombinant SLBSD were carried out at 30 °C overnight in a total volume of 100 μl 197 containing Tris-HCl buffer (10 mM, pH 8.0), blasticidin S/mildiomycin/cytoglucuronic acid (0.5 198
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

10
mM/0.5 mM/0.5 mM), and the corresponding His-tagged SLBSD(10 µM). The reactions were 199 quenched by the addition of chloroform. The products and substrates were analyzed by LC-MS as 200 described above. 201 202 Results 203 204 Engraftment of Blasticidin S biosynthetic gene cluster to S. lividans 205 206 The blasticidin S producer has a very strong barrier system against introduction of foreign DNA, 207 which made prior genetic engineering of the blasticidin S gene cluster unsuccessful. To bypass 208 this tough genetic manipulation system, attachment of the biosynthetic gene cluster onto the 209 genome of S. lividans has been accomplished to generate a genetically stable heterologous host. 210 211 As depicted in Figure 2A, the complete blasticidin S gene cluster and flanking sequence was 212 excised from 7D11 (Figure S1A) and inserted into pJTU1289 to generate pJTU1780 (Table S1 and 213 Figure S1A&B). All internal StuI-fragments of the insertion were removed and replaced with a 214 blunt aadA cassette to form pJTU1785 (Figure S1C), leaving a 2.0 kb and a 7.6 kb DNA fragment at 215 5’ end and 3’ end of the insertion, respectively. A 4.85 kb dispensable DNA fragment from S. 216 lividans HXY16 on pJTU1528(Figure S1D) was separated into 1.6kb and 3.0kb by replacing its 217 internal 0.25 kb StuI fragment with the 2.0 kb-aadA-7.6 kb EcoRV fragment from pJTU1785, 218 generating pJTU1786 (Figure S1E). The 1.6 kb and 3.0 kb arms were then employed as the target 219 sites for integration of the sandwiched 2.0kb-aadA-7.6kb fragment into S. lividans HXY16 220 chromosome via double crossover to give mutant S. lividans LL1 (aadA). Finally, pJTU1780 was 221 transferred to S. lividans LL1 to replace the 2.0kb-addA-7.6kb fragment with the blasticidin S 222 biosynthetic gene cluster and the flanking sequences. Thus, S. lividans LL2, a mutant strain 223
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

11
containing the blasticidin S gene cluster, was constructed and confirmed by Southern blot (Figure 224 2B). 225 226 When assayed for antifungal activity, S. lividans LL2 showed a very slight inhibitory effect versus 227 the indicator strain Rhodotorula rubra (Figure S2). To more accurately measure the blasticidin S 228 production in S. lividians LL2, the fermentation broth was analyzed by LC-MS but no peak 229 corresponding to a blasticidin S standard was observed in the LC profile. However, a new 230 compound was detected in the fermentation broth with a molecular weight one mass unit greater 231 than that of blasticidin S ([M+H]+ 424, Figure 3A). The MS/MS analysis of this compound was 232 identified it as the non-toxic deaminohydroxyblasticidin S (Figure 3B), which is the known 233 product of blasticidin S deaminase (BSD, Figure 3C) (28). To confirm this result, this compound 234 was further purified and analyzed by Q-TOF MS (Figure S3). 235 236 Identification of a BSD gene in S. lividansHXY16 237 The existence of deaminohydroxyblasticidin S in the fermentation broth of LL2 prompted us to 238 search for a homolog of the Aspergillus terreus BSD in the genome of S. lividans. Hence, we 239 identified a cytidinedeaminase with accession number EFD66414 in S. lividans TK24 that showed 240 51% identity and 67% similarity to the A. terreus BSD. The S. lividans HXY16 strain was also 241 found to contain a BSD homolog, named SLBSD that is almost identical to the TK24 enzyme 242 except for a single mutation (E112D, Figure S4). 243 244 To provide means to measure the ability of SLBSD to convert blasticidin S to 245 deaminohydroxyblasticidin S, SLBSD was heterologously expressed in E. coli. A purified soluble 246
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

12
protein 16.9 KD in size, consistent with the calculated molecular weight of the His-tagged SLBSD, 247 was obtained (Figure 4A). LC-MS analysis of an overnight reaction incubating SLBSD with 248 blasticidin S showed that blasticidin S ([M+H]+ 423) was completely transformed to 249 deaminohydroxyblasticidin S ([M+H]+ 424) (Figure 4B). Moreover, the substrate specificity of 250 SLBSD was surveyed by examining its activity on mildiomycin, a nucleoside with similar structure 251 to blasticidin S, cytoglucuronic acid, an intermediate in blasticidin S biosynthesis and CMP. The 252 deaminase activity was observed with mildiomycin and cytoglucuronic acid as substrates but 253 conversion was comparatively low (Figure 4C&4D). However, SLBSD did not catalyze the 254 deamination of CMP (data not shown), suggesting the hexose moiety may be an essential 255 component for recognition by SLBSD. 256 257 Inactivation of SLBSD in S. lividans LL2 leads to the production of Blasticidin S 258 To stop the conversion of blasticidin S to deaminohydroxyblasticidin S by SLBSD in the blasticidin 259 S producing S. lividans LL2, SLBSD was disrupted via double crossover. The resulting strain was 260 termed S. lividans WJ2 and the correct genotype was confirmed by PCR (Figure 5A). LC-MS 261 analysis of the purified fermentation broth of mutant WJ2 grown in production medium revealed 262 a peak with same retention time as the blasticidin S standard. MS analysis of this peak further 263 confirmed the production of blasticidin S in the mutant (Figure 5B). 264 265 To compare the effect of SLBSD disruption in S. lividans strains with different genotypes, SLBSD 266 was also inactivated in S. lividans HXY16 via double crossover to generate mutant S. lividans WJ1 267 (Figure S5A). WJ1, S. lividans HXY16 and WJ2 were then assayed for their resistance level against 268
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

13
blasticidin S at different concentrations (20μg – 200μg/ml). The growth of HXY16 was not greatly 269 inhibited by blasticidin S at the concentration of 200μg/ml, while the SLBSD mutant WJ1 was 270 completely inhibited by blasticidin S at a concentration of 100μg/ml, as indicated by a clear zone 271 on an agar test plate. Moreover, WJ2, containing the blasticidin S self-resistance gene blsJ, was 272 also not affected by blasticidin S (Figure S5B). This observation suggested that the heterologous 273 production of blasticidin S in WJ2 should not significantly affect growth of the host strain. 274 275 We previously found that the mildiomycin producer, Streptoverticillium rimofaciens ZJU5119, 276 could produce three additional mildiomycin derivatives in the seed/fermentation broth (29). To 277 check the secondary metabolite production by S. lividans WJ2, the strain was grown in the 278 standard seed/fermentation medium, and analysis revealed a new peak exhibiting a dominant 279 ion at [M+H]+ 409 in the LC profile of the fermentation broth (Figure 6A). This product was 280 identified as demethylblasticidin S by MS/MS analysis (Figure 6B) and Q-TOF MS (Figure S6). 281 However, only blasticidin S was detected when S. griseochromogenes was grown in the same 282 seed/fermentation medium (data not shown). 283 284 Generation of blsE, blsF and blsL disruption mutants 285 To demonstrate the ability to manipulate the blasticidin S gene cluster in a heterologous host, 286 three genes, blsE, blsF and blsL, were disrupted individually and the fermentation broths from the 287 resulting mutant strains were analyzed by LC-MS. 288 289 Because demethylblasticidin S accumulated in the fermentation broth of WJ2, and was 290 considered an intermediate in the blasticidin S biosynthetic pathway (15), the formation of 291 demethylblasticidin S in WJ2 was investigated. An N-methyltransferase is the predicted product 292
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14
of blsL, thus this gene was selectively inactivated and the genotype of resulting mutant WJ3 was 293 confirmed by PCR (Figure 6C). 294 295 There remain several steps in the biosynthetic pathway to blasticidin S that are unclear. To 296 narrow down the genes involved in blasticidin S assembly, blsF a gene homologous to milL in the 297 mildiomycin gene cluster that is predicted to encode a protein of unknown function (30), was 298 disrupted. Disruption of blsF (Figure 7A) did not abolish the production of blasticidin S. On the 299 contrary, the production of blasticidin S was increased in the mutant, S. lividans WJ4 (Figure 7B). 300 Moreover, in the crude extract of the fermentation broth of WJ4, a known intermediate of the 301 blasticidin S biosynthetic pathway, cytosylglucuronic acid, accumulated (Figure S7). 302 303 The accumulation of cytosylglucuronic acid in the WJ4 mutant implied that the enzyme acting on 304 this intermediate was very important in increasing the production of blasticidin S. BLAST analysis 305 showed that BlsE belonged to the radical SAM superfamily, members of which are capable of 306 catalyzing multiple reaction types, such as decarboxylation, methylation and dehydration (31). In 307 our earlier work, the disruption of milG, a homologue to blsE in the mildiomycin biosynthetic 308 pathway, led to the accumulation of hydroxymethyl cytosylglucuronic acid (30). To analyze the 309 function of blsE in blasticidin S formation, the gene was inactivated and the resulting mutant, S. 310 lividans WJ5, was confirmed by PCR (Figure 7C). As expected, production of blasticidin S was 311 abolished in WJ5 (Figure S8), and the accumulation of cytosylglucuronic acid was observed in the 312 fermentation broth of this mutant (Figure 7D). 313 314
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

15
Discussion 315 316 As introduction of alien DNA into the chromosome of the blasticidin S native producer S. 317 griseochromogenes was never successful, in vivo functional studies access the role of each gene 318 involved in blasticidin S biosynthesis was hindered. To bypass the strong DNA uptake barrier 319 system of S. griseochromogenes, we set forth to engineer a S. lividans-derived BS-producing host 320 that was genetically stable and manipulable. However, the resulting S. lividans LL2 mutant that 321 carried the intact blasticidin S gene cluster was found to only produce a blasticidin S derivative, 322 deaminohydroxyblasticidin S, this observation led to the discovery of a blasticidin S deaminase in 323 S. lividans. 324 325 BSD was first discovered in Aspergillus terreus in 1975(32). It was shown to be a zinc-dependent 326 enzyme (28) and the blasticidin S-bound crystal structure of BSD revealed the position of the zinc 327 ion was very close to C2 of the cytosine base of blasticidin S (33). This is the site occupied by a 328 hydroxymethyl group in mildiomycin and may thus hinder its coordination to the active zinc ion 329 center. This assumption may in part explain the low in vitro transformation efficiency (Figure 4C) 330 of mildiomycin to deaminhydroxymildiomycin by SLBSD. Consistent with this assumption, 331 mildiomycin was successfully produced in S. lividans (30) whereas all of the blasticidin S was 332 converted into deaminohydroxyblasticidin S. As for cytosylglucuronic acid, another substrate for 333 SLBSD, this compound was nearly completely transformed into 334 deaminohydroxycytosylglucuronic acid. The detection of cytosylglucuronic acid in the 335 fermentation broth of the earlier heterologous expression experiments (15) might be the result of 336
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

16
the high yield of this compound. Finally, the finding that CMP was not recognized by SLBSD might 337 be due to the lack of carboxyl group on its sugar ring or the negative charge carried by the 338 phosphate group, both of which were essential for the recognition of blasticidin S to BSD as 339 proposed by Kumasaka et al (33). 340 341 The disruption of SLBSD led to the successful heterologous expression of blasticidin S for the first 342 time. This engineered blasticidin S-producing strain provided necessary means to investigate the 343 blasticidin S biosynthesis pathway in vivo. The disruption of blsF, a homologue of milL that had 344 been confirmed as a gene unnecessary for mildiomycin biosynthesis (30), raised the yields of 345 blasticidin S and other intermediates instead of abolishing their production. This result suggested 346 that blsF might have a negative effect on blasticidin S biosynthesis. Moreover, accumulation of 347 cytosylglucuronic acid and demethylblasticidin S in WJ4 suggested that the production of the 348 cytotoxic blasticidin S repressed or slowed down the metabolic flow toward the final product. 349 However, in the native producer, the co-evolution of blasticidin S and a self-resistant mechanism 350 can efficiently counter the cytotoxic activity. A good case for this is that there exist two 351 chromosomal fragments containing putative resistance genes to blasticidin S (15). One fragment 352 contains a transporter-encoding gene blsJ (16) while the other resistant element is located 353 outside of blasticidin S biosynthetic gene cluster. 354 355 The accumulation of cytosylglucuronic acid in the WJ4 mutant may also result from the inefficient 356 expression, or the deficient performance, of some gene(s) or gene products in the heterologous 357 host. Identification of the proteins responsible for directly processing cytosylglucuronic acid to 358
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

17
advanced intermediates is therefore very important for fully utilizing the substrates and 359 increasing the yield of blasticidin S in a heterologous expression host. In mildiomycin 360 biosynthesis, disruption of milG, a blsE homolog, led to the accumulation of hydroxymethyl 361 cytosylglucuronic acid (30). This result provided a hint that blsE was a key gene for the 362 accumulation of cytoglucuronic acid. As expected, disruption of blsE abolished the production of 363 blasticidin S while the accumulation of cytosylglucuronic acid remained. 364 365 The wild-type S. griseochromogenes can produce demethylblasticidin S when inoculated in YEME 366 medium (15) and demethylblasticidin S can be transformed to blasticidin S by a cell-free extract 367 of this strain (34). It is very likely that BlsL alone is in charge of this transformation. In a previous 368 report, the non-toxic leucylblasticidin S was detected in the heterologous expression strain and a 369 route involving leucyldemethylblasticidin S and leucylblasticidin S as precursors to blasticidin S 370 pathway was proposed (16). However, in present study, leucylblasticidin S was not regularly 371 detected in the fermentation broth. A blasticidin S acetyltransferase was discovered in 372 Streptoverticillum sp in 1990 (8) and a homolog exists in many Streptomyces including S. lividans 373 (accession number: ZP_06530511). The formation of leucylblasticidin S in the original study thus 374 may have resulted from a host resistance or detoxification mechanism rather than from an 375 enzyme in the blasticidin S biosynthetic pathway. Furthermore, the inactivation of blsL led solely 376 to production of demethylblasticidin S, which suggests the direct conversion of 377 demethylblasticidin S to blasticidin S is more likely. 378 379 In conclusion, blasticidin S is a very important nucleoside antibiotic both in agriculture and in 380
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

18
laboratory research. The successful heterologous expression of this antibiotic in S. lividans HXY16 381 bypassed the difficult genetic manipulation of its native producer and led to the discovery of a 382 blasticidin S deaminase from the S. lividans HXY16. Moreover, the easy genetic manipulation of S. 383 lividans unveiled the function of several genes in the biosynthetic gene cluster. This is very 384 important for the demonstration of the whole biosynthetic pathway of blasticidin S and for 385 increasing its production in both native and heterologous host. 386 387 Acknowledgement 388 This study was supported in part by grants from the National Natural Science Foundation of 389 China; the Ministry of Science and Technology of China (973 and 863 Programs); Ministry of 390 Education of China; the Chen Xing Young Scholars Program, Shanghai Jiao Tong University. 391 392 Reference 393 1. Takeuchi, S., K. Hirayama, K. Ueda, H. Sakai, and H. Yonehara. 1958. Blasticidin S, a 394
new antibiotic. J Antibiot (Tokyo) 11:1-5. 395 2. Huang, K. T., T. Misato, and H. Asuyama. 1964. Selective Toxicity of Blasticidin S to 396
Piricularia oryzae and Pellicularia sasakii. J Antibiot (Tokyo) 17:71-74. 397 3. Hansen, J. L., P. B. Moore, and T. A. Steitz. 2003. Structures of five antibiotics bound at 398
the peptidyl transferase center of the large ribosomal subunit. J Mol Biol 330:1061-1075. 399 4. Ichikawa, Y., K. Hirata, M. Ohbayashi, and M. Isobe. 2004. Total synthesis of 400
(+)-blasticidin s. Chemistry 10:3241-3251. 401 5. Yamaguchi, H., C. Yamamoto, and N. Tanaka. 1965. Inhibition of protein synthesis by 402
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

19
blasticidin S. I. Studies with cell-free systems from bacterial and mammalian cells. J 403 Biochem 57:667-677. 404
6. Tamura, K., M. Kimura, and I. Yamaguchi. 1995. Blasticidin S deaminase gene (BSD): a 405 new selection marker gene for transformation of Arabidopsis thaliana and Nicotiana 406 tabacum. Biosci Biotechnol Biochem 59:2336-2338. 407
7. Karreman, C. 1998. New positive/negative selectable markers for mammalian cells on 408 the basis of Blasticidin deaminase-thymidine kinase fusions. Nucleic Acids Res 409 26:2508-2510. 410
8. Pérez-González, J. A., D. Ruiz, J. A. Esteban, and A. Jiménez. 1990. Cloning and 411 characterization of the gene encoding a blasticidin S acetyltransferase from 412 Streptoverticillum sp. Gene 86:129-134. 413
9. Mamoun, C. B., I. Y. Gluzman, S. Goyard, S. M. Beverley, and D. E. Goldberg. 1999. A 414 set of independent selectable markers for transfection of the human malaria parasite 415 Plasmodium falciparum. Proc Natl Acad Sci U S A 96:8716-8720. 416
10. Tercero, J. A., J. C. Espinosa, R. A. Lacalle, and A. Jimenez. 1996. The biosynthetic 417 pathway of the aminonucleoside antibiotic puromycin, as deduced from the molecular 418 analysis of the pur cluster of Streptomyces alboniger. J Biol Chem 271:1579-1590. 419
11. Chen, W., T. Huang, X. He, Q. Meng, D. You, L. Bai, J. Li, M. Wu, R. Li, Z. Xie, H. Zhou, X. 420 Zhou, H. Tan, and Z. Deng. 2009. Characterization of the polyoxin biosynthetic gene 421 cluster from Streptomyces cacaoi and engineered production of polyoxin H. J Biol Chem 422 284:10627-10638. 423
12. Bormann, C., W. Huhn, H. Zahner, R. Rathmann, H. Hahn, and W. A. Konig. 1985. 424
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

20
Metabolic products of microorganisms. 228. New nikkomycins produced by mutants of 425 Streptomyces tendae. J Antibiot (Tokyo) 38:9-16. 426
13. Rackham, E. J., S. Gruschow, A. E. Ragab, S. Dickens, and R. J. Goss. 2010. Pacidamycin 427 biosynthesis: identification and heterologous expression of the first uridyl peptide 428 antibiotic gene cluster. Chembiochem 11:1700-1709. 429
14. Bormann, C., V. Mohrle, and C. Bruntner. 1996. Cloning and heterologous expression of 430 the entire set of structural genes for nikkomycin synthesis from Streptomyces tendae 431 Tu901 in Streptomyces lividans. J Bacteriol 178:1216-1218. 432
15. Cone, M. C., A. K. Petrich, S. J. Gould, and T. M. Zabriskie. 1998. Cloning and 433 heterologous expression of blasticidin S biosynthetic genes from Streptomyces 434 griseochromogenes. J Antibiot (Tokyo) 51:570-578. 435
16. Cone, M. C., X. Yin, L. L. Grochowski, M. R. Parker, and T. M. Zabriskie. 2003. The 436 blasticidin S biosynthesis gene cluster from Streptomyces griseochromogenes: sequence 437 analysis, organization, and initial characterization. Chembiochem 4:821-828. 438
17. Grochowski, L. L., and T. M. Zabriskie. 2006. Characterization of BlsM, a nucleotide 439 hydrolase involved in cytosine production for the biosynthesis of blasticidin S. 440 Chembiochem 7:957-964. 441
18. Gould, S. J., and J. Guo. 1994. Cytosylglucuronic acid synthase (cytosine: 442 UDP-glucuronosyltransferase) from Streptomyces griseochromogenes, the first 443 prokaryotic UDP-glucuronosyltransferase. J Bacteriol 176:1282-1286. 444
19. Seto, H., K. Furihata, and H. Yonehara. 1976. Studies on the biosynthesis of blasticidin 445 S. V. Isolation and structure of pentopyranic acid. J Antibiot (Tokyo) 29:595-596. 446
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

21
20. Liu, H. W., and J. S. Thorson. 1994. Pathways and mechanisms in the biogenesis of novel 447 deoxysugars by bacteria. Annu Rev Microbiol 48:223-256. 448
21. Kieser, T., M. J. Bibb, K. F. Chater, M. J. Butter, and D. A. Hopwood (ed.). 2000. Practical 449 Streptomyces genetics. The John Innes Foundation, United Kingdom, Norwich. 450
22. Sambrook, J., E. F. Fritsch, and T. Maniatis (ed.). 1989. Molecular cloning: a laboratory 451 manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. 452
23. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, and D. J. 453 Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database 454 search programs. Nucleic Acids Res 25:3389-3402. 455
24. He, Y., Z. Wang, L. Bai, J. Liang, X. Zhou, and Z. Deng. 2010. Two pHZ1358-derivative 456 vectors for efficient gene knockout in streptomyces. J Microbiol Biotechnol 20:678-682. 457
25. Sun, Y., X. He, J. Liang, X. Zhou, and Z. Deng. 2009. Analysis of functions in plasmid 458 pHZ1358 influencing its genetic and structural stability in Streptomyces lividans 1326. 459 Appl Microbiol Biotechnol 82:303-310. 460
26. Gust, B., G. L. Challis, K. Fowler, T. Kieser, and K. F. Chater. 2003. PCR-targeted 461 Streptomyces gene replacement identifies a protein domain needed for biosynthesis of 462 the sesquiterpene soil odor geosmin. P Natl Acad Sci USA 100:1541-1546. 463
27. He, X., H. Y. Ou, Q. Yu, X. Zhou, J. Wu, J. Liang, W. Zhang, K. Rajakumar, and Z. Deng. 464 2007. Analysis of a genomic island housing genes for DNA S-modification system in 465 Streptomyces lividans 66 and its counterparts in other distantly related bacteria. Mol 466 Microbiol 65:1034-1048. 467
28. Kimura, M., S. Sekido, Y. Isogai, and I. Yamaguchi. 2000. Expression, purification, and 468
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

22
characterization of blasticidin S deaminase (BSD) from Aspergillus terreus: the role of 469 catalytic zinc in enzyme structure. J Biochem 127:955-963. 470
29. LI, L., X. HE, and Z. Deng. 2009. Analysis of Mildiomycin and Its Derivatives from the 471 Culture Filtrate of Streptoverticillium rimofaciens ZJU5119. JOURNAL OF SHANGHAI 472 JIAOTONG UNIVERSITY 43:1-4. 473
30. Wu, J., L. Li, Z. Deng, T. M. Zabriskie, and X. He. 2012. Analysis of the Mildiomycin 474 Biosynthesis Gene Cluster in Streptoverticillum remofaciens ZJU5119 and 475 Characterization of MilC, a Hydroxymethyl cytosyl-glucuronic Acid Synthase. 476 Chembiochem 13:1613-1621. 477
31. Layer, G., D. W. Heinz, D. Jahn, and W. D. Schubert. 2004. Structure and function of 478 radical SAM enzymes. Curr Opin Chem Biol 8:468-476. 479
32. Yamaguchi, I., H. Shibata, H. Seto, and T. Misato. 1975. Isolation and purification of 480 blasticidin S deaminase from Aspergillus terreus. J Antibiot (Tokyo) 28:7-14. 481
33. Kumasaka, T., M. Yamamoto, M. Furuichi, M. Nakasako, A. H. Teh, M. Kimura, I. 482 Yamaguchi, and T. Ueki. 2007. Crystal structures of blasticidin S deaminase (BSD): 483 implications for dynamic properties of catalytic zinc. J Biol Chem 282:37103-37111. 484
34. Guo, J., and S. Gould. 1991. Biosynthesis of Blasticidin S. Cell-free Demonstration of 485 N-methylation as the last step. Bioorganic and Medicinal Chem. Lett I:497-500. 486 487
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

23
488 Figure Legend 489 490 Figure 1 Chemical structures of blasticidin S, mildiomycin and cytosylglucuronic acid(CGA). 491 492 Figure 2 Construction of the S. lividans-derived strain LL2 that was engrafted with blasticidin 493 S biosynthetic gene cluster. A) Schematic representation of the construction of S. lividans LL2. 494 B) Confirmation of the engraftment of the bls gene cluster to S. lividans HXY16 by Southern 495 blot. Genomic DNA preparations from: Lane 1, S. lividans HXY16; 2 and 3, two individual 496 strains of S. lividans LL2; 4, S. griseochromogenes, were digested with BamHI, separated by 497 agarose gel electrophoresis and probed with BamHI-cut pJTU1780. 498 499 Figure 3 LC-MS analysis of the metabolite production of S. lividans LL2. A) HPLC comparison 500 of the blasticidin S standard to the purified fermentation broth of S. lividansLL2 and S. 501 lividans HXY16, LC-MS analysis revealed the peak presented in the fermentation broth of LL2 502 having a dominant peak at m/z of 424; B) MS/MS analysis of the m/z 424 parent ion. Each 503 daughter peak labeled could be derived from deaminohydroxyblasticidin S. C) Blasticidin S 504 was transformed to deaminohydroxyblasticidin S by BSD. 505 506 Figure 4 In vitro and in vivo analysis of SLBSD. A) SDS-PAGE analysis of purified SLBSD. First 507 two lanes, purified SLBSD; right lane, protein marker. The calculated MW of His-tagged 508 SLBSD was 16.9KDa. B) LC-MS analysis of products after overnight incubation with SLBSD 509
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

24
and boiled SLBSD with blasticidin S as substrate. C) LC-MS analysis of products after 510 overnight incubation with SLBSD and boiled SLBSD with mildiomycin was as substrate. D) 511 LC-MS analysis of products after overnight incubation with SLBSD and boiled SLBSD with 512 cytosylglucuronic acid as substrate. 513 514 Figure 5 HPLC analysis of the metabolite production of S. lividans WJ2. A) Confirmation of the 515 genotype of ΔSLBSD mutant WJ2 by PCR with primer Con-BKO (Table S2). M, DNA marker; 516 HXY16, PCR product targeting SLBSD from S. lividans HXY16; WJ2, PCR product targeting 517 SLBSD from S. lividans WJ2. B) HPLC comparison of the blasticidin S standard to the purified 518 fermentation broth of S. lividans WJ2 and S. lividans HXY16. 519 520 Figure 6 Assay of the blasticidin S production of the ΔblsL mutant strain S. lividans WJ3. A) 521 HPLC comparison of the blasticidin S standard to the purified fermentation broth of WJ3, 522 WJ2 and HXY16. LC-MS analysis revealed the peak presented in the fermentation broth of 523 LL2 and WJ2 having a dominant peak at m/z of 409. B) MS/MS analysis of the m/z 409 524 parent ion. Each labeled daughter peak could be derived from demethylblasticidin S. C) PCR 525 confirmation of the genotype of ΔblsL mutant WJ3 with primer Con-blsL (Table S2). M, DNA 526 marker; WJ2, PCR product targeting blsL from S. lividans WJ2; WJ3, PCR product targeting 527 blsL from S. lividans WJ3. 528 529 Figure 7 Assay of the blasticidin S production of the ΔblsF and ΔblsE mutant strain, S. lividans 530 WJ4 and S. lividans WJ5. A) Confirmation of the ΔblsF mutant WJ4 by PCR with primer 531
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

25
Con-blsF (Table S2). M, DNA marker; WJ2, PCR product targeting blsF from S. lividans WJ2; 532 WJ4, PCR product targeting blsF from S. lividans WJ4. B) HPLC comparison of the purified 533 fermentation broth of WJ4 to that of WJ2. C) Confirmation of the ΔblsE mutant WJ5 by PCR 534 with primer Con-blsE (Table S2). M, DNA marker; WJ2, PCR product targeting blsE from S. 535 lividans WJ2; WJ5, PCR product targeting blsE from S. lividans WJ5. D) HPLC comparison of 536 the untreated fermentation broth of WJ5 to that of WJ2 and HXY16. 537
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Blasticidin S Mildiomycin Cytosylglucuronic acid (CGA)
Fig. 1
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from
[M+H]+ =42460
UV272/mAUA) C)
[M+H]+ =4231000
LL2
600
BS
HXY16 Blasticidin S[M+H]+ 423
Deaminhydroxyblasticidin S[M+H]+ 424
0min
34323028262422201816
B)
[ ] [M H] 4 4
B)
a
d-c407.0
+MS2(424.5), 27.8min
OH-BS
b
c
e
-a
-ac
b-bc -ce
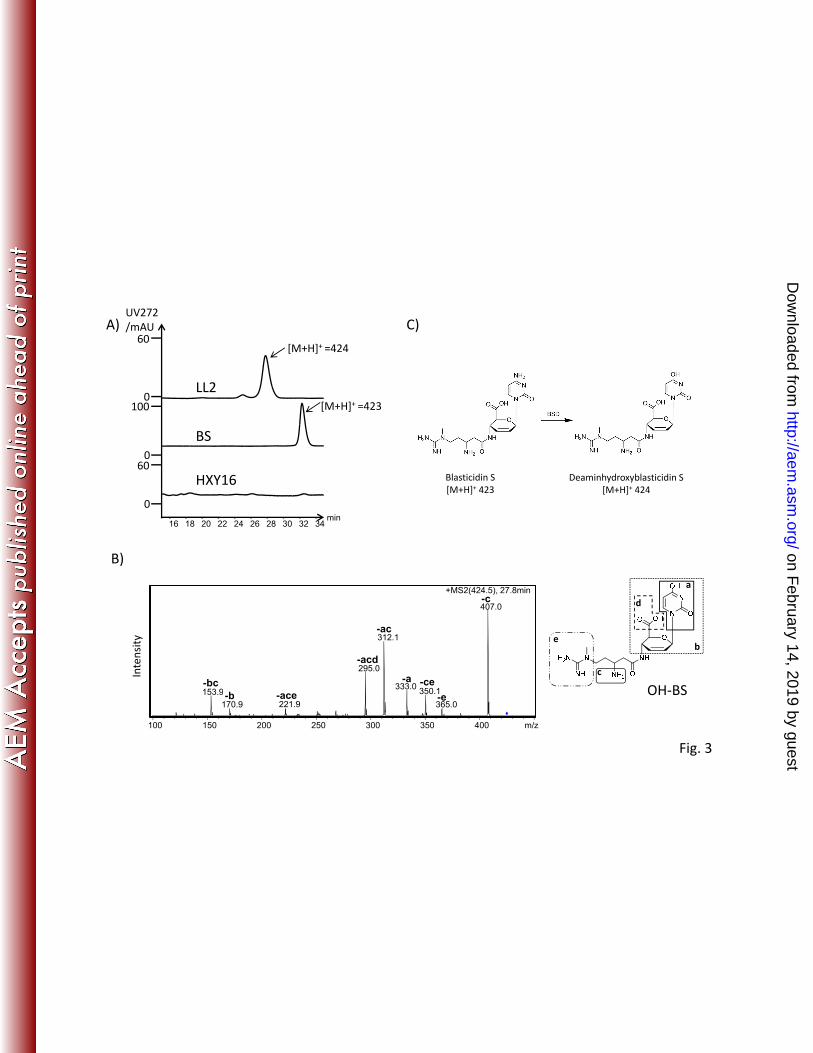
-acd
153 9
295.0
312.1
333.0 350 1
Inte
nsity
Fig. 3
OH BS-b -e-ace153.9170.9 221.9
350.1365.0
100 150 200 250 300 350 400 m/z
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

[M H]+ 424
UV272/mAUA) B)
BS with SLBSD
[M+H]+ =424
[M+H]+ =4235000
200/
BS with boiled SLBSD
[M+H] 423
302520151050min0
500
20
UV272/mAU [M+H]+ =516
80
UV272/mAU
CGA i h SLBSD
[M+H]+ =289C) D)
[M+H]+ =288
060
MIL with SLBSD
MIL with boiled SLBSD
[M+H]+ =5150
100
CGA with SLBSD
CGA with boiled SLBSD
[M+H]+ =288
[M+H]+ =288
3025201510500 min min1614121086420
0
Fig. 4
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

M WJ2 HXY16UV272/
A) B)M WJ2 HXY16
100
0
/mAU
BS
0 5kb
1.8kb
) )
0
0
60
60
WJ2(ΔSLBSD)
0.5kb
0 min16 18 20 22 24 26 28 30 32 34
HXY16
Fig. 5
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

100
UV272/mAU
A) C)M WJ3 WJ2
0120 [M+H]+ =409
WJ3(ΔblsL)
BS1.0kb
1.7kb
[M+H]+ =409
0
060
WJ3(ΔblsL)
WJ2
min343230282624222018160
060
HXY16
-a298.0
sity
B) +MS2(409.1), 30.2mina
-ac-acb221.9
280.9
Inten
b
c
100 150 200 250 300 350 400
Fig. 6
m/z
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from

M WJ5 WJ2WJ4 WJ2 M A) C)
1.2kb1.4kb1.3kb
1.4kb
UV272UV272
350
/mAU
WJ5(ΔblsE)
[M+H]+=288140
UV272/mAU
WJ2
B) D)
0
0350
WJ2
WJ5(ΔblsE)0
140
WJ2
WJ4(ΔblsF)
min0 5 10 15 20 25 30 35 40
0
0750
HXY16
min3530252015
0
0 5 10 15 20 25 30 35 40
Fig. 7
on February 14, 2019 by guest
http://aem.asm
.org/D
ownloaded from








![ilOTO}KEHUE - admpolazna.ru filenorlyJrrrpn3arJr4r4 n rrpo.4rBr{xeHr{to 3AopoBofo o6pa3a xu3Hr{ B MOJIOAeII(HOII CpeAe, B TOM rrrrcJre [oBbrrrreHr4ro ypoBHfl npoBeAeH]aq](https://img.pdfslide.us/doc/110x75/5e0b1fc808103b01350a2ee9/ilotokehue-n-rrpo4rbrxehrto-3aopobofo-o6pa3a-xu3hr-b-mojioaeiihoii-cpeae.jpg)





















