Embed Size (px)
Citation preview

LETTERS
Doushantuo embryos preserved inside diapause eggcystsLeiming Yin1, Maoyan Zhu1, Andrew H. Knoll2, Xunlai Yuan1, Junming Zhang1 & Jie Hu1
Phosphatized microfossils in the Ediacaran (635–542 Myr ago)Doushantuo Formation, south China, have been interpreted asthe embryos of early animals1–4. Despite experimental demonstra-tion that embryos can be preserved5, microstructural evidence thatthe Doushantuo remains are embryonic6 and an unambiguousrecord of fossil embryos in Lower Cambrian rocks7, questionsabout the phylogenetic relationships of these fossils remain. Mostrecently, some researchers have proposed8 that Doushantuo micro-fossils may be giant sulphur-oxidizing bacteria comparable toextant Thiomargarita sp. Here we report new observations thatprovide a test of the bacterial hypothesis. The discovery ofembryo-like Doushantuo fossils inside large, highly ornamentedorganic vesicles (acritarchs) indicates that these organisms wereeukaryotic, and most probably early cleavage stage embryos pre-served within diapause egg cysts. Large acanthomorphic micro-fossils of the type observed to contain fossil embryos first appearin rocks just above a 632.5 6 0.5-Myr-old ash bed9, suggesting thatat least stem-group animals6 inhabited shallow seas in the imme-diate aftermath of global Neoproterozoic glaciation.
Simple embryos and Thiomargarita sp. are both spheroidal, andboth divide repeatedly without intervening growth to form multicel-lular structures with overall dimensions comparable to those of thesingle cells from which they derive. Although members of the genusThiomargarita lack a number of features observed in Doushantuofossils10,11, it is later developmental stages that would provide definitivesupport for the embryo hypothesis; however, these have been recog-nized only tentatively11. There is another test, though. Bacteria may besurrounded by polysaccharide envelopes of simple morphology, butthey do not form large, preservable cysts with complex ornamentation.In contrast, the life cycles of many eukaryotes, including some animals,include resting stages characterized by ornate and preservable walls12.Doushantuo beds containing fossils interpreted as embryos also pre-serve abundant and diverse acritarchs characterized by large size (100to ,700mm diameter) and conspicuous ornamentation that is distri-buted symmetrically over the vesicle surface (Fig. 1)13,14. Among theextant organisms, by far the closest morphological matches to thesefossils are diapause egg cysts, or hulls, produced by a phylogeneticallydisparate array of aquatic invertebrates15,16.
1State Key Laboratory of Paleobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China. 2Botanical Museum,Harvard University, Cambridge, Massachusetts 02138, USA.
Figure 1 | Large acanthomorphic acritarchs, some containing preservedembryos, in the Ediacaran Doushantuo Formation, Xiaofenghe section,western Hubei Province, China. a, Ericiasphaera magna, sample numberXFHB-35. b, Appendisphaera tenuis, sample number XFHB-33.c–l, Tianzhushania spinosa; c–h, k, l sample number XFHB-2; i, j samplenumber XFHB-29. Panels c–h illustrate one (c), two (d, e), eight (f) and 16(g, with greater detail in h) cell stages of preserved embryos within
encompassing structures interpreted as diapause egg cysts; k, l, show detailsof wall structure in the specimens illustrated in c and g. Panel j, magnified inl, shows a Tianzhushania fossil from , 632-Myr-old beds of the lowerDoushantuo Formation. Scale bar (in h) represents 32 mm for a; 30 mm forb; 185mm for c; 182mm for d; 185mm for e; 163mm for f; 195mm forg; 130mm for h; 50 mm for i; 98 mm for j; 38mm for k; and 98 mm for l.
Vol 446 | 5 April 2007 | doi:10.1038/nature05682
661Nature ©2007 Publishing Group
It has been proposed that the acritarch genus Tianzhushania (vesi-cles of 300–700 mm diameter, each with a wall ornamented by numer-ous cylindrical processes and surrounded by an outer membrane)represents the external hull of fossil eggs and embryos17. This con-clusion is based on petrological and scanning electron micrographobservations that link Tianzhushania to spheroidal fossils with cere-broid to tuberuclate surfaces that are interpreted as egg envelopes2.One Tianzhushania vesicle with two cells inside are known, but untilnow no contents with higher cell numbers have been discovered17.
New collections from the Xiaofenghe section of the DoushantuoFormation (Fig. 2), along the eastern limb of the Huangling anticlinenear Yichang, China, confirm and extend previous interpretations17,showing that cleavage, at least through the 16-cell stage, occurredwithin Tianzhushania hulls (Fig. 1). The 1, 2, 4, 8 and 16-cell stagespreserved inside Tianzhushania vesicles are identical to other popu-lations interpreted as early cleavage embryos2. Thus, the life cycle ofthe organisms that produced the embryos also included formation ofa complex cyst or hull. This is inconsistent with the interpretationof the populations as giant sulphur bacteria; indeed, we know of nobacteria that produce acanthomorphic spores of the size, morpho-logy and preservability exhibited by the Doushantuo structures12.The preserved processes observed cannot be bacterial fimbriaebecause they are hollow and arise from the surface of an encom-passing cyst, not from surfaces interpreted as cell walls in ref. 8. In
contrast, the combined morphological evidence of ornamented wallsand multicellular contents is fully consistent with the interpretationof the Doushantuo fossils as animals and suggests that a resting stagewas part of early-animal life cycles. Other large, spinose microfossilsin Doushantuo phosphorites also preserve multicellular contents13,so perhaps a greater record of animal diversity exists among thediverse acanthomorphic cyst walls preserved in Doushantuo andother Ediacaran rocks12,18.
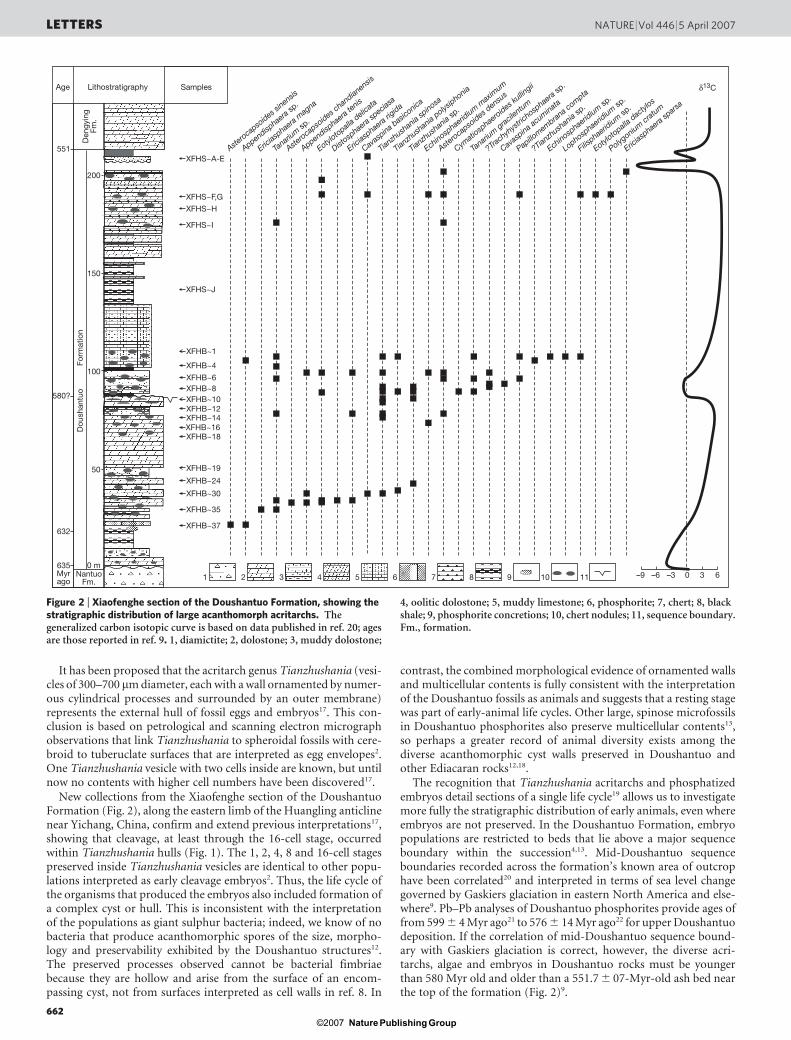
The recognition that Tianzhushania acritarchs and phosphatizedembryos detail sections of a single life cycle19 allows us to investigatemore fully the stratigraphic distribution of early animals, even whereembryos are not preserved. In the Doushantuo Formation, embryopopulations are restricted to beds that lie above a major sequenceboundary within the succession4,13. Mid-Doushantuo sequenceboundaries recorded across the formation’s known area of outcrophave been correlated20 and interpreted in terms of sea level changegoverned by Gaskiers glaciation in eastern North America and else-where9. Pb–Pb analyses of Doushantuo phosphorites provide ages offrom 599 6 4 Myr ago21 to 576 6 14 Myr ago22 for upper Doushantuodeposition. If the correlation of mid-Doushantuo sequence bound-ary with Gaskiers glaciation is correct, however, the diverse acri-tarchs, algae and embryos in Doushantuo rocks must be youngerthan 580 Myr old and older than a 551.7 6 07-Myr-old ash bed nearthe top of the formation (Fig. 2)9.
Dou
shan
tuo
Form
atio
n
NantuoFm.
δ13CAge Lithostratigraphy Samples
1 2 3 4 5 6 7 8 9 10 11
XFHS~F,G
XFHS~A-E
XFHS~I
XFHS~J
XFHS~H
XFHB~1
XFHB~4
XFHB~6XFHB~8XFHB~10XFHB~12XFHB~14XFHB~16XFHB~18
XFHB~19
XFHB~35
XFHB~30
XFHB~24
XFHB~37
551
635Myrago
632
580?
Den
gyin
gFm
.
0 63–3–6–90 m
50
100
150
200
Tianzh
ushan
ia sp
inosa
Asteroca
psoides
sinen
sis
Appendisp
haera
sp.
Tanari
um sp.
Ericias
phaera
magna
Appendisp
haera
tenis
Eotylotopall
a deli
cata
Asteroca
psoides
chan
dianen
sis
Distosp
haera
spec
iasa
Ericias
phaera
rigida
Cavas
pina bas
iconica
Tianzh
ushan
ia polys
iphonia
Tianzh
ushan
ia sp
.
Asteroca
psoides
densu
s
Echinosp
haerid
ium max
imum
Cymati
osphae
roides ku
llingii
Tanari
um gracile
ntum
Cavas
pina acu
minata
?Trach
yhys
trichosp
haera
sp.
Papillo
membran
a compta
?Tianzh
ushan
ia sp
.
Echinosp
haerid
ium sp.
Lophosphae
ridium sp
.
Filisphae
ridium sp
.
Eotylotopall
a dac
tylos
Polygonium cr
atum
Ericias
phaera
spars
a
Figure 2 | Xiaofenghe section of the Doushantuo Formation, showing thestratigraphic distribution of large acanthomorph acritarchs. Thegeneralized carbon isotopic curve is based on data published in ref. 20; agesare those reported in ref. 9. 1, diamictite; 2, dolostone; 3, muddy dolostone;
4, oolitic dolostone; 5, muddy limestone; 6, phosphorite; 7, chert; 8, blackshale; 9, phosphorite concretions; 10, chert nodules; 11, sequence boundary.Fm., formation.
LETTERS NATURE | Vol 446 | 5 April 2007
662Nature ©2007 Publishing Group

A bed-by-bed census of microfossils throughout the 220 mXiaofeng succession indicates that Tianzhushania acritarchs occurlow in the formation, within chert and phosphorite nodules in dolo-mitic shales only about 30 m above glaciogenic rocks of the under-lying Nantuo Formation (Fig. 2; see also ref. 23). U–Pb zirconanalyses provide a date for an ash bed within the cap carbonate atthe formation’s base of 635.2 6 0.6 Myr ago; a second ash bed 5 mbelow the earliest Tianzhushania fossils is dated as being 632.5 6 0.5Myr old9. Thus, organisms that produced hulls that are indistinguish-able from those containing embryos, higher in the Doushantuo suc-cession, existed within about 3 million years of snowball ice sheetdecay24. Other taxa of large acanthomorphic microfossils occur stilllower in the section, below the 632.5 6 0.5-Myr-old ash bed (Fig. 2)23.
In summary, the discovery that large, morphologically complexcyst walls or hulls contain fossils that are interpreted as the earlycleavage stages of animal embryos supports interpretation of bothembryos and cyst walls as metazoan. The expanded set of searchimages for early animal fossils afforded by this association providesevidence that at least stem group metazoans were components ofearliest Ediacaran ecosystems, later expanding markedly, perhapsin association with rising oxygen levels25–27.
Received 8 January; accepted 13 February 2007.
1. Xiao, S., Zhang, Y. & Knoll, A. H. Three-dimensional preservation of algaeand animal embryos in a Neoproterozoic phosphorite. Nature 391, 553–558(1998).
2. Xiao, S. & Knoll, A. H. Phosphatized animal embryos from the NeoproterozoicDoushantuo Formation at Weng’an, Guizhou, South China. J. Paleontol. 74,767–788 (2000).
3. Chen, J. et al. Small bilaterian fossils from 40 to 55 million years before theCambrian. Science 305, 218–222 (2006).
4. Dornbos, S. Q. et al. Environmental controls on the taphonomy of phosphatizedanimals and animal embryos from the Neoproterozoic Doushantuo Formation,southwest China. Palaios 21, 3–14 (2006).
5. Raff, E. C., Villinski, J. T., Turner, F. R., Donoghue, P. C. J. & Raff, R. A. Experimentaltaphonomy shows the feasibility of fossil embryos. Proc. Natl Acad. Sci. USA 103,5846–5851 (2006).
6. Hagadorn, J. W. et al. Cellular and subcellular structure of Neoproterozoic animalembryos. Science 314, 291–294 (2006).
7. Donoghue, P. C. J. et al. Fossilized embryos are widespread but the record istemporally and taxonomically biased. Evol. Dev. 8, 232–238 (2006).
8. Bailey, J. V., Joye, S. B., Kalanetra, K. M., Flood, B. E. & Corsetti, F. A. Evidence ofgiant sulfur bacteria in Neoproterozoic phosphorites. Nature 445, 198–201(2007).
9. Condon, D. et al. U-Pb ages from the Neoproterozoic Doushantuo Formation,China. Science 308, 95–98 (2005).
10. Donoghue, P. C. J. Embryonic identity crisis. Nature 445, 155–156 (2007).
11. Xiao, S., Hagadorn, J. W., Zhou, C. & Yuan, X. Rare helical spheroidal fossils fromthe Doushantuo Lagerstatte: Ediacaran animal embryos come of age? Geology 35,115–118 (2007).
12. Knoll, A. H., Javaux, E. J., Hewitt, D. & Cohen, P. Eukaryotic organisms inProterozoic oceans. Phil. Trans. Roy. Soc. B 361, 1023–1038 (2006).
13. Zhang, Y., Yin, L., Xiao, S. & Knoll, A. H. Permineralized fossils from the terminalProterozoic Doushantuo Formation, south China. J. Paleontol. 72, (Suppl. to no. 4),1–52 (1998).
14. Yuan, X. & Hofmann, H. J. New micropaleontological data from NeoproterozoicSinica Doushantuo phosphorite rocks, Weng’an, Guizhou Province, southwesternChina. Alcheringa 22, 189–222 (1998).
15. Van Waveren, I. & Marcus, N. H. Morphology of recent copepod egg envelopesfrom Turkey Point, Gulf of Mexico, and their implications for acritarch affinity.Spec. Pap. Paleontol. 48, 111–124 (1993).
16. Buckland-Nicks, J. Hull cupules of chiton eggs: parachute structures and spermfocusing devices? Biol. Bull. 184, 269–276 (1993).
17. Yin, C., Bengtson, S. & Yue, Z. Silicified and phosphatized Tianzhushania,spheroidal microfossils of possible animal origin from the Neoproterozoic ofSouth China. Acta Palaeontol. Pol. 49, 1–12 (2004).
18. Grey, K. Ediacaran palynology of Australia. Mem. Assoc. Australas. Palaeontologists31, 1–439 (2005).
19. Marcus, N. H. & Boero, F. Minireview: The importance of benthic–pelagic couplingand the forgotten role of life cycles in coastal aquatic systems. Limnol. Oceanogr.43, 763–768 (1998).
20. Zhu, M., Zhang, J. & Yang, A. Integrated Ediacaran (Sinian)chronostratigraphy of South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. (in thepress).
21. Barfod, G. H. et al. New Lu-Hf and Pb-Pb age constraints on the earliest animalfossils. Earth Planet. Sci. Lett. 201, 203–212 (2002).
22. Chen, D., Dong, W. Q., Qi, L., Chen, G. Q. & Chen, X. P. Pb-Pb ages ofNeproterozoic Doushantuo phosphorites in South China: constraints on earlymetazoan evolution and glaciation events. Precambr. Res. 132, 123–222 (2004).
23. Zhou, C., Xie, G., Mcfadden, K., Xiao, S. & Yuan, X. The diversification andextinction of Doushantuo–Pertatataka acritarchs in South China: causes andbiostratigraphic significance. Geol. J. 41, 1–34 (2006).
24. Hoffman, P. F., Kaufman, A. J., Halverson, G. P. & Schrag, D. P. A Neoproterozoicsnowball earth. Science 281, 1342–1346 (1998).
25. Voreb’eva, N. G., Sergeev, V. N. & Knoll, A. H. Diverse Ediacaran acritarchs fromthe margin of the East European Platform. Geology (submitted).
26. Canfield, D. E., Poulton, S. W. & Narbonne, G. M. Late Neoproterozoicdeep-ocean oxygenation and the rise of animal life. Science 10.1126/science.1135013 (2006).
27. Fike, D. A., Grotzinger, J. P., Pratt, L. M. & Summons, R. E. Oxidation of theEdiacaran ocean. Nature 444, 744–747 (2006).
Acknowledgements This work was supported by the Chinese Academy ofSciences, the National Natural Science Foundation of China, the Major BasicResearch Projects of MST of China, and an NSF Grant. We thank S. Xiao andP. Cohen for discussions.
Author Information Reprints and permissions information is available atnpg.nature.com/reprintsandpermissions. The authors declare no competingfinancial interests. Correspondence and requests for materials should beaddressed to L.Y. ([email protected]).
NATURE | Vol 446 | 5 April 2007 LETTERS
663Nature ©2007 Publishing Group