Embed Size (px)
Citation preview

LETTERS
Structure and mechanism of the M2 proton channel ofinfluenza A virusJason R. Schnell1 & James J. Chou1
The integral membrane protein M2 of influenza virus forms pH-gated proton channels in the viral lipid envelope1. The low pH ofan endosome activates the M2 channel before haemagglutinin-mediated fusion. Conductance of protons acidifies the viral inter-ior and thereby facilitates dissociation of the matrix protein fromthe viral nucleoproteins—a required process for unpacking of theviral genome2. In addition to its role in release of viral nucleopro-teins, M2 in the trans-Golgi network (TGN) membrane preventspremature conformational rearrangement of newly synthesizedhaemagglutinin during transport to the cell surface by equilibrat-ing the pH of the TGN with that of the host cell cytoplasm3.Inhibiting the proton conductance of M2 using the anti-viral drugamantadine or rimantadine inhibits viral replication4–7. Here wepresent the structure of the tetrameric M2 channel in complexwith rimantadine, determined by NMR. In the closed state, fourtightly packed transmembrane helices define a narrow channel, inwhich a ‘tryptophan gate’ is locked by intermolecular interactionswith aspartic acid. A carboxy-terminal, amphipathic helixoriented nearly perpendicular to the transmembrane helixforms an inward-facing base. Lowering the pH destabilizes thetransmembrane helical packing and unlocks the gate, admittingwater to conduct protons, whereas the C-terminal base remainsintact, preventing dissociation of the tetramer. Rimantadine bindsat four equivalent sites near the gate on the lipid-facing side of thechannel and stabilizes the closed conformation of the pore. Drug-resistance mutations are predicted to counter the effect of drugbinding by either increasing the hydrophilicity of the pore orweakening helix–helix packing, thus facilitating channel opening.
M2 is a 97-residue single-pass membrane protein that has itsamino and carboxy termini directed towards the outside and insideof the virion, respectively; it is a homotetramer in its native state8,9.The four transmembrane helices form a channel in which His 37 isthe pH sensor and Trp 41 is the gate6,10,11. The adamantane-baseddrugs amantadine and rimantadine, which target the M2 channel,have been used as first-choice antiviral drugs against communityoutbreaks of influenza A viruses for many years, but resistance tothe adamantanes has recently become widespread. Many structuralmodels of this channel have been built, based on sequence analysis,mutagenesis and solid-state NMR8,11,12. Many of these studies havebeen done on inherently unstable transmembrane-only constructs,however, leading to conflicting structural conclusions.
Although the transmembrane-only peptide fails to form a stabletetramer, a construct of residues 18–60 (M2(18–60)), which includes15 residues of the C terminus in addition to the transmembraneregion, forms a stable tetramer in dihexanoyl-phosphatidyl-choline(DHPC) detergent micelles and yields high-resolution NMR spectra(Supplementary Fig. 1). In the closed conformation at pH 7.5,M2(18–60) is a homotetramer in which each subunit has anunstructured N terminus (residues 18–23), a channel-forming
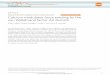
transmembrane helix (residues 25–46), a short flexible loop (residues47–50) and a C-terminal amphipathic helix (residues 51– 59). Thetransmembrane helices assemble into a four-helix bundle with a left-handed twist angle of ,23u and a well defined pore (Fig. 1). A ring ofmethyl groups from Val 27 constricts the N-terminal end of the poreto ,3.1 A (inner diameter). In agreement with proposed models11,13,His 37 and Trp 41 are inside the pore. A three-bond, 15N–13Cc scalarcoupling (3JNCc) value of 1.5 Hz (Supplementary Table 1) shows theHis 37 x1 rotamer to be predominantly trans, but with significantrotameric averaging. The x1 of Trp 41 is essentially locked in the transposition, as determined by a 3JNCc of 2.6 Hz, whereas the x2 is alsofixed at around 2120u by the side chain He1–Ne1 dipolar couplingand nuclear Overhauser effects (NOEs). The Trp 41 indole rings areat van der Waals distance from each other, prohibiting passage ofwater or ions (Fig. 1c). The indole He1 of one subunit is on average3.5 A from the Asp 44 carboxyl carbon of the adjacent subunit. Thetwo residues can form an intermolecular hydrogen bond that stabi-lizes the closed Trp 41 gate. The side chain of Arg 45 probably parti-cipates in an intermolecular interaction with Asp 44. These findingsare consistent with the increased pH-modulated activity of channelsin which asparagine has replaced Asp 44 (ref. 14).
The C-terminal end of the channel extends into a loop (residues47–50) that connects the transmembrane domain to the C-terminalamphipathic helix. Residual dipolar couplings (RDCs) and intra- andinter-monomer NOEs show that the amphipathic helices lie roughlyperpendicular (,82u) to the transmembrane helices and assemblehead-to-tail using a right-handed packing mode to form the base ofthe channel. The orientation and amphipathic character of theamphipathic helices suggest that the C-terminal base lies on thesurface of the membrane.
Residues 47–50 give no NOE peaks and do not have a stable,hydrogen-bonded structure in the detergent micelles used in ourwork. We believe that this segment adopts a more stable conforma-tion in the viral membrane because Cys 50, which we mutated toserine to avoid disulphide formation, is normally palmitoylated15.Modelling shows that extending the transmembrane helix toPhe 48 would place residue 50 facing the membrane, allowing forinsertion of the palmitoyl acyl chain into the lipid bilayer. This minorrearrangement would also move the amphipathic helices closer to thetransmembrane domain.
Drug binding stabilizes the closed conformation. On addition ofdrug, the resonances of residues 43–46 at the C terminus of thechannel, which are severely exchange-broadened in the drug-freesample, became significantly sharper and more homogeneous(Supplementary Fig. 2). The protein–drug NOEs collected fromfour different NOE spectroscopy (NOESY) spectra (SupplementaryFig. 3) place the binding site between adjacent helices at theC-terminal end of the transmembrane domain near the Trp 41 gate,on the membrane side of the channel (Fig. 1d). We could not detect
1Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, Massachusetts 02115, USA.
Vol 451 | 31 January 2008 | doi:10.1038/nature06531
591Nature Publishing Group©2008

drug NOEs in other parts of the protein, including the widely pro-posed drug-binding site in the pore of the channel. The amine head-group of rimantadine is in contact with the polar side chains ofAsp 44 and Arg 45, and with the indole amine of Trp 41. The sidechains of Ile 42 from one helix and Leu 40 and Leu 43 from anotherhelix form the hydrophobic walls of the binding pocket that interactwith the adamantane group of rimantadine. Thus, rimantadinecovers a unique polar patch in the otherwise hydrophobic envir-onment of the transmembrane domain. Interactions between riman-tadine and the channel are consistent with structure–activityrelationships of the adamantane group16. In particular, the basicnitrogen group and size limits at the methyl site are critical. Theserequirements are the result of the interactions with Asp 44 and thesmall hydrophobic pocket around Ile 42, respectively.
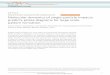
Water NOEs measured in the 110-ms 15N-separated NOESYexperiments give a clear picture of water distribution relative to thechannel (Fig. 2). The lipid-facing surface of the transmembraneregion is largely protected from water by the DHPC micelle. In the
closed channel pore, the Val 27 ring at the N terminus and the Trp 41gate at the C terminus essentially block water from freely diffusinginto the pore from either side of the membrane. Within the trans-membrane region, only the amides of Ser 31 and Ile 32 have NOEcrosspeaks at the chemical shift of water, probably corresponding tothe hydroxyl proton of Ser 31 in exchange with water. A polar residueis present at position 31 in all sequenced variants of M2, suggestingthat proton conduction requires water to be bound to this site. Thiswater may serve to bridge the proton relay from the N-terminal endof the pore to the His 37 pH sensor. Water was detected at the Cterminus of the transmembrane region, beginning at Arg 45. The He1of the Trp 41 indole ring, which points towards the C-terminal side ofthe pore, also has a strong NOE to water, indicating that the base ofthe channel is accessible to bulk water.
Lowering the pH from 7.5 to 6.5 broadens most of the NMRresonances corresponding to the transmembrane helix (Fig. 3a).The resonance broadening could not be attributed to proteinaggregation, because the self-diffusion coefficients were essentiallyunchanged between pH 7.5 and pH 6.5. Thus, activation of the chan-nel is coupled to increased conformational exchange in the trans-membrane domain. In contrast, the resonances of the amphipathichelices are essentially unaffected by lowering the pH, indicating thatthe C-terminal base of the tetramer remains intact as the channelopens.
In addition to destabilizing helix–helix packing in the transmem-brane domain, channel activation must also correlate with increaseddynamics of the Trp 41 gate. Because the indole amide resonance ofTrp 41 remained strong as the pH was lowered from 7.5 to 6.0, itserves as a useful NMR probe for monitoring opening of the channel.We compared the millisecond timescale dynamics of the Trp 41indole ring between the closed and open states by carrying outrelaxation-compensated Carr–Purcell–Meiboom–Gill (CPMG)experiments17 at pH 7.5, pH 7.0 and pH 6.0. A two-site exchangemodel fits the dependence of 15N relaxation caused by chemical shiftexchange on the frequency of refocusing (1/tcp) of chemical shiftevolution (Fig. 3b), implying that the gate switches between twoconfigurations at any given pH. As the pH was lowered from 7.5 to6.0, the rate of fluctuation increased by more than fourfold (Fig. 3b),indicating that channel activation ‘unlocks’ the gate. Adding riman-tadine to the channel at an intermediate pH of 7.0 slowed the time-scale of the gate motion to nearly that of a drug-free gate at pH 7.5(Fig. 3c). These results confirm that the reconstituted channels in the
~30 Å
Tran
smem
bra
ne c
hann
el
C-terminal base
Asp 44 OδTrp 41 Hε1
d
b
c
a
Figure 1 | Structure of the M2 channel. a, An ensemble of 15 low-energystructures derived from NMR restraints. Because residues 47–50 areunstructured, the transmembrane helices (residues 25–46) and theamphipathic helices (residues 51–59) are superimposed separately. Thebackbone r.m.s. deviations for the transmembrane and amphipathic helicesare 0.30 A and 0.56 A, respectively. b, A ribbon representation of a typicalstructure from the ensemble in a, showing the left-handed packing of thetransmembrane helices, right-handed packing of the amphipathic helices,the side chains of His 37 and Trp 41, and the drug rimantadine (coloured inred). c, A close-up view from the C-terminal side of the channel showing theTrp 41 gate and how it is stabilized by the inter-monomer hydrogen bondbetween Trp 41 He1 of one transmembrane helix and Asp 44 carboxyl of theadjacent transmembrane helix. d, The surface representation of therimantadine-binding pocket, showing the Asp 44, the indole amine ofTrp 41, and Arg 45, which form the polar patch, as well as the hydrophobicwall composed of Leu 40, Ile 42 and Leu 43.
1.4 Å Trp 41 indoles
3.1 Å Val 27 methyls
1.7 Å His 37 imidazole
6.7 Å Gly 34
4.4 Å Ala 30 methylsSer 31
ba
Figure 2 | Water accessibility of the M2 channel. a, Distribution of waterNOEs relative to the structure. Amide protons coloured in blue have a NOEcrosspeak to water. Those that do not are coloured red. b, The pore surfacecalculated using the program HOLE. The region of the channel coloured ingreen is only wide enough to allow passage of a water molecule, whereas theblue portion can accommodate two or more water molecules. The orangeregion is too narrow to allow any ions to pass through.
LETTERS NATURE | Vol 451 | 31 January 2008
592Nature Publishing Group©2008

NMR sample are pH-gated, and are consistent with the location ofthe rimantadine site proximal to the gate.
The structure of the M2 proton channel thus reveals a simple yeteffective gating. The tight packing of the four transmembrane helicesbrings the bulky indole rings of Trp 41 into van der Waals contact toform the channel gate. The gate is further stabilized by inter-subunithydrogen bonds with Asp 44. Lowering the pH protonates theimidazole rings of His 37, destabilizing helix–helix packing by elec-trostatic repulsion. This conformational rearrangement breaks inter-actions between Trp 41 and Asp 44 and allows the gate to flip open. A
pair of conserved N-terminal cysteines have been shown to formintermolecular disulphides in vivo9. Thus, the transmembrane helicesare tethered at one end by N-terminal disulphides and at the otherend by the C-terminal base, ensuring that destabilization of the four-helix bundle during channel activation does not cause dissociation ofthe tetramer (Fig. 4). Indeed, truncation of the amphipathic helixresults in channels that rapidly lose channel activity18.
The discovery of the external drug-binding site was unexpected.Drug-resistance mutations seemed to suggest that the drug-bindingsite was inside the pore, because, in early models of the channel,residues that lead to drug resistance were predicted to be pore-lining.The known mutations that confer drug resistance are L26F, V27A,A30T, S31N, G34E and L38F. Mapping these residues onto thestructure (Supplementary Fig. 7) reveals that Val 27, Ala 30 andGly 34 are pore-lining, but that Leu 26, Ser 31 and Leu 38 are in thehelix–helix packing interface. Moreover, these mutations are spreadout over more than three turns of the transmembrane helix, coveringa distance much larger than the dimensions of amantadine or riman-tadine. The authors of ref. 19 pointed out that having a cork-plugging-the-bottle model is insufficient to explain all the resultsof electrophysiology studies. For example, drug inhibition is moreeffective when applied to the closed channel than to the open chan-nel, which is not expected of a pore-blocking mechanism5. Several ofthe drug-resistance mutations in pore-lining residues have beenshown to retain drug binding20. Although a pore blocker is expectedto fit tightly in the pore, channel inhibition is unusually tolerant ofmodifications to the adamantane scaffold16. Together, the aboveobservations suggest an allosteric inhibition mechanism.
Is the external binding site consistent with all drug-resistancemutations? Although the exact structural effects of resistance muta-tions are difficult to predict, what they do have in common is thatthey either perturb the helix–helix interface (L26F, V27A, S31N,L38F) or increase the hydrophilicity of the pore (A30T, G34E).
0.5 1.0 1.5 2.015
20
25
30
1/τCP (ms–1)1/τCP (ms–1)
15
20
25
30
35
a
0.5 1.0 1.5 2.0
8.6 8.2 7.8 7.41H (p.p.m.)
126
124
122
120
118
116
114
112
110
15N (p
.p.m
.)
8.6 8.2 7.8 7.48.6 8.2 7.8 7.48.6 8.2 7.8 7.4
(–Rim.)(–Rim.)(–Rim.)(–Rim.)pH 6.0pH 6.5pH 7.0pH 7.5
40
R2
(τC
P) (
s–1 )
c
R2
(τC
P) (
s–1 )
bpH 6.0, kex = 2,300 s–1
pH 7.0, kex = 1,400 s–1
pH 7.5, kex = 500 s–1
–Rimantadine, kex = 1,400 s–1
+Rimantadine, kex = 740 s–1
Figure 3 | Low-pH-induced destabilization of the channel and opening ofthe Trp 41 gate. a, 1H-15N TROSY spectra of reconstituted M2(18–60)tetramer at pH 6.0, 6.5, 7.0 and 7.5, in the absence of rimantadine (2Rim.),recorded at 500 MHz 1H frequency and 30 uC. Green, transmembrane helix;pink, amphipathic helix; black, N-terminal loop. b, The 15N R2 (pureR2 1 Rex) of the Trp 41 Ne1 as a function of the frequency of refocusing (1/
tcp) of chemical shift evolution obtained at pH 7.5, 7.0 and 6.0, showingfaster timescale motion of the Trp 41 gate as the channel is activated.c, Comparison between R2(tcp) at pH 7.0 in the absence (blue) and presence(black) of rimantadine, demonstrating that the drug slows down the gateflipping at this pH.
N
C
H37 H37
W41
D44
W41
TMTM TM
BaseBase
Low-pH activation
Mem
bra
ne
Mem
brane
+ +
TM
C
Disulphide DisulphideN
Figure 4 | Schematic illustration of M2 channel activation. At high pH, thetransmembrane (TM) helices are packed tightly and the tryptophan gate islocked through intermolecular interactions with Asp 44. At low pH,protonation of the His 37 imidazoles destabilizes the transmembrane helixpacking, allowing hydration of the channel pore and proton conductance.The C-terminal base of the tetramer and N-terminal disulphide bonds keepthe channel from completely disassembling. For clarity, only two of the fourmonomers are shown.
NATURE | Vol 451 | 31 January 2008 LETTERS
593Nature Publishing Group©2008

From this observation, and from detection of a conformationalexchange among multiple states at lowered pH, we propose an allos-teric inhibition mechanism that can account for all of the mutations.In our model, drug binding makes the closed channel harder to open,whereas drug-resistance mutations destabilize the closed channel,making it easier to open. Replacing Val 27 with alanine enlarges theN-terminal opening and weakens helix–helix packing, and thereforemay facilitate channel opening. Ala 30 and Gly 34 are inside the pore,and replacing them with threonine and glutamate, respectively, mayfacilitate pore hydration, and, in turn, channel opening. Leu 26,Ser 31 and Leu 38 are helix–helix interface residues; their mutationsprobably perturb helix–helix packing and lower the energetic cost ofchannel opening.
Why, then, are no drug-resistant mutations observed near thedrug-binding site? In fact, few mutations are ever observed in thisregion of the channel (drug-resistant or otherwise), which is notsurprising owing to the functional constraints placed on these resi-dues in proximity to the channel pH sensor (His 37) and the channelgate (Trp 41). As the structure illustrates, intermolecular contactsbetween Asp 44 and Arg 45 form an integral part of the channel gate,along with Trp 41. The residues that form the hydrophobic walls ofthe binding pocket—Leu 40, Ile 42 and Leu 43—are on the lipid faceof the channel and must retain hydrophobicity for membrane parti-tioning. To accommodate the Trp 41 indole rings within the channel,the helices splay slightly at the C terminus of the transmembranedomain, and interhelical contacts below Leu 38—with the exceptionof the Trp 41 side chains—are no longer important for channelassembly. Thus, residues essential to channel assembly are in theN-terminal half of the transmembrane helix, exactly where drug-resistance mutations occur.
Binding from the membrane side is consistent with the high mem-brane partition coefficient of adamantane drugs, which effectivelyconcentrates them in the membrane and lowers their level in theaqueous phase21,22. Adamantanes interact with a number of otherion channels, including viroporins from hepatitis C23, the potassiumchannel Kcv of the chlorella virus PBCV-1 (ref. 24) and the humanNMDA receptors25. Hanatoxin, an allosteric inhibitor of voltage-gated K1 channels with a high membrane partition coefficient, alsohas an external binding site26,27. Membrane-side binding may thus bea feature of many channel inhibitors. This mode of inhibition couldbe advantageous for drug design because drug molecules are typicallymuch larger than hydrated ions selected by ion channels, and there-fore the energy barrier for the drug to find a blocking site inside thechannel pores would be much higher than targeting a functional sitefrom the membrane side of the channel.Note added in proof: We note that, in a separate X-ray study of thetransmembrane domain of M2, an electron density, which was pro-posed to be from amantadine, was observed inside the channel pore(see ref. 28).
METHODS SUMMARY
The M2(18–60) polypeptide construct was expressed as a C-terminal fusion to
bacterial trpLE with an N-terminal His9 tag in the pMM-LR6 vector29. The
M2(18–60) tetramer was reconstituted by dissolving peptide in a solution con-
taining 50 mM sodium phosphate, 6 M guanidine HCl and 150 mM DHPC,
dialyzing against a solution containing 40 mM sodium phosphate (pH 7.5)
and 30 mM glutamate, and concentrating. Rimantadine was added to the recon-
stituted protein. The final NMR sample used for structure determination
contained 0.75 mM M2(18–60) (monomer), ,300 mM DHPC and 40 mM
rimantadine. Given that the DHPC has an aggregation number of 27 (ref. 30)
and the strong partition coefficient of rimantadine in phospholipids (rimanta-
dine aqueous solubility is very low, ,50 mM), locally, there are about four
rimantadine molecules per micelle compartment in which the channel resides.
The NMR protocol used was similar to that described previously31. An
extensive set of structural restraints (including 230 3 4 intra- and 27 3 4
inter-molecular distance restraints derived from NOEs, 27 3 4 orientation
restraints from residual dipolar couplings (RDCs), and 23 3 4 side-chain rota-
mers from three-bond scalar couplings) were used to generate an ensemble of 15
low-energy structures with a backbone root mean square (r.m.s.) deviation of
0.30 A for the channel region and of 0.89 A for all structured regions (Fig. 1a).
The refinement statistics and NMR-derived restraints are summarized in
Supplementary Table 1. Structure calculation was accomplished in two steps,
in which the overall tetramer conformation was first defined by NOE-derived
distance restraints and J-coupling-derived dihedral restraints using a high-
temperature simulated annealing protocol, and was subsequently refined against
RDCs at low temperature.
Full Methods and any associated references are available in the online version ofthe paper at www.nature.com/nature.
Received 15 July; accepted 3 December 2007.
1. Lamb, R. A., Holsinger, L. J. & Pinto, L. H. Receptor-Mediated Virus Entry into Cells(ed., Wimmer, E.) 303–321 (Cold Spring Harbor Laboratory Press, Cold SpringHarbor, 1994).
2. Helenius, A. Unpacking the incoming influenza-virus. Cell 69, 577–578 (1992).3. Ciampor, F. et al. Evidence that the amantadine-induced, M2-mediated
conversion of influenza A virus hemagglutinin to the low pH conformation occursin an acidic trans Golgi compartment. Virology 188, 14–24 (1992).
4. Hay, A. J., Wolstenholme, A. J., Skehel, J. J. & Smith, M. H. The molecularbasis of the specific anti-influenza action of amantadine. EMBO J. 4, 3021–3024(1985).
5. Wang, C., Takeuchi, K., Pinto, L. H. & Lamb, R. A. Ion channel activity of influenza Avirus M2 protein: characterization of the amantadine block. J. Virol. 67,5585–5594 (1993).
6. Pinto, L. H., Holsinger, L. J. & Lamb, R. A. Influenza virus M2 protein has ionchannel activity. Cell 69, 517–528 (1992).
7. Chizhmakov, I. V. et al. Selective proton permeability and pH regulation of theinfluenza virus M2 channel expressed in mouse erythroleukaemia cells. J. Physiol.(Lond.) 494, 329–336 (1996).
8. Sugrue, R. J. & Hay, A. J. Structural characteristics of the M2 protein of influenza Aviruses: evidence that it forms a tetrameric channel. Virology 180, 617–624(1991).
9. Holsinger, L. J. & Lamb, R. A. Influenza virus M2 integral membrane protein is ahomotetramer Stabilized by formation of disulfide bonds. Virology 183, 32–43(1991).
10. Tang, Y., Zaitseva, F., Lamb, R. A. & Pinto, L. H. The gate of the influenza virus M2proton channel is formed by a single tryptophan residue. J. Biol. Chem. 277,39880–39886 (2002).
11. Pinto, L. H. et al. A functionally defined model for the M2 proton channel ofinfluenza A virus suggests a mechanism for its ion selectivity. Proc. Natl Acad. Sci.USA 94, 11301–11306 (1997).
12. Wang, J. F., Kim, S., Kovacs, F. & Cross, T. A. Structure of the transmembraneregion of the M2 protein H1 channel. Protein Sci. 10, 2241–2250 (2001).
13. Kukol, A., Adams, P. D., Rice, L. M., Brunger, A. T. & Arkin, I. T. Experimentallybased orientational refinement of membrane protein models: A structure for theinfluenza A M2 H1 channel. J. Mol. Biol. 286, 951–962 (1999).
14. Betakova, T., Ciampor, F. & Hay, A. J. Influence of residue 44 on the activity of theM2 proton channel of influenza A virus. J. Gen. Virol. 86, 181–184 (2005).
15. Sugrue, R. J., Belshe, R. B. & Hay, A. J. Palmitoylation of the influenza A virus M2protein. Virology 179, 51–56 (1990).
16. Aldrich, P. E. et al. Antiviral agents. 2. Structure–activity relationships ofcompounds related to 1-adamantanamine. J. Med. Chem. 14, 535–543 (1971).
17. Loria, J. P., Rance, M. & Palmer, A. G. A relaxation-compensatedCarr–Purcell–Meiboom–Gill sequence for characterizing chemical exchange byNMR spectroscopy. J. Am. Chem. Soc. 121, 2331–2332 (1999).
18. Tobler, K., Kelly, M. L., Pinto, L. H. & Lamb, R. A. Effect of cytoplasmic tailtruncations on the activity of the M(2) ion channel of influenza A virus. J. Virol. 73,9695–9701 (1999).
19. Pinto, L. H. & Lamb, R. A. Understanding the mechanism of action of the anti-influenza virus drug amantadine. Trends Microbiol. 3, 271 (1995).
20. Astrahan, P., Kass, I., Cooper, M. A. & Arkin, I. T. A novel method of resistancefor influenza against a channel-blocking antiviral drug. Proteins 55, 251–257(2004).
21. Subczynski, W. K., Wojas, J., Pezeshk, V. & Pezeshk, A. Partitioning andlocalization of spin-labeled amantadine in lipid bilayers: an EPR Study. J. Pharm.Sci. 87, 1249–1254 (1998).
22. Wang, J. F., Schnell, J. R. & Chou, J. J. Amantadine partition and localization inphospholipids membrane: a solution NMR study. Biochem. Biophys. Res. Commun.324, 212–217 (2004).
23. Griffin, S. D. et al. The p7 protein of hepatitis C virus forms an ion channelthat is blocked by the antiviral drug, Amantadine. FEBS Lett. 535, 34–38(2003).
24. Plugge, B. et al. A potassium channel protein encoded by chlorella virus PBCV-1.Science 287, 1641–1644 (2000).
25. Svensson, T. H. Dopamine release and direct dopamine receptor activation in thecentral nervous system by D-145, an amantadine derivative. Eur. J. Pharmacol. 23,232–238 (1973).
LETTERS NATURE | Vol 451 | 31 January 2008
594Nature Publishing Group©2008

26. Swartz, K. J. & MacKinnon, R. Hanatoxin modifies the gating of a voltage-dependent K1 channel through multiple binding sites. Neuron 18, 665–673(1997).
27. Lee, S.-Y. & MacKinnon, R. A membrane-access mechanism of ion channelinhibition by voltage sensor toxins from spider venom. Nature 430, 232–235(2004).
28. Stouffer, A. L. et al. Structural basis for the function and pharmaceutical inhibitionof an influenza virus proton channel. Nature doi:10.1038/nature06528 (thisissue).
29. Call, M. E. et al. The structure of the ff transmembrane dimer reveals featuresessential for its assembly with the T cell receptor. Cell 127, 355–368 (2006).
30. Chou, J. J., Baber, J. L. & Bax, A. Characterization of phospholipids mixed micellesby translational diffusion. J. Biomol. NMR 29, 299–308 (2004).
31. Oxenoid, K. & Chou, J. J. The structure of phospholamban pentamer reveals achannel-like architecture in membranes. Proc. Natl Acad. Sci. USA 102,10870–10875 (2005).
Supplementary Information is linked to the online version of the paper atwww.nature.com/nature.
Acknowledgements We thank M. Berardi for many discussions, and S. Harrison fordiscussion and assisting with the manuscript. This work was supported by the NIHand the Pew Scholars Program in the Biomedical Sciences awarded to J.J.C. J.R.S. issupported by an NIH F32 postdoctoral fellowship.
Author Contributions J.R.S. and J.J.C. designed research, performed research,analysed data and wrote the paper.
Author Information The structures have been deposited in the Protein Data Bankunder the accession number 2RLF. Reprints and permissions information isavailable at www.nature.com/reprints. The authors declare competing financialinterests: details accompany full-text HTML version of the paper onwww.nature.com/nature. Correspondence and requests for materials should beaddressed to J.J.C. ([email protected]).
NATURE | Vol 451 | 31 January 2008 LETTERS
595Nature Publishing Group©2008

METHODSSample preparation. M2(18–60) was expressed into inclusion bodies as a fusion
to His9–trpLE32. The M2(18–60) peptide was released from the fusion protein by
cyanogen bromide digestion in 70% formic acid (2 h, 0.2 g ml21). The digest was
dialyzed to water, lyophilized, and loaded onto a C4 column (Grace–Vydac) in
2:1:2 hexafluoroisopropanol:formic acid:water and separated on a gradient of
3:2 isopropanol:acetonitrile. The lyophilized peptide was refolded at 250mM by
dissolving in 6 M guanidine and 150 mM DHPC and dialyzing against the final
NMR buffer containing 40 mM sodium phosphate and 30 mM glutamate. The
sample was concentrated to a final M2(18–60) concentration of 0.75 mM(monomer). Rimantadine was added after concentrating. The concentration
of DHPC was determined from 1H NMR spectroscopy to be around 300 mM.
NMR spectroscopy. NMR experiments were conducted at 30 uC on spectro-
meters equipped with cryogenic probes (Bruker). Sequence-specific assignment
of backbone 1HN, 15N, 13Ca and 13Cb chemical shifts were accomplished using
the TROSY versions of the HNCA (1HN, 15N and 13Ca correlation spectroscopy)
and HNCACB (1HN, 15N and 13Cb correlation spectroscopy) experiments on a15N-, 13C- and 85% 2H-labelled protein. Side-chain x1 and x2 rotamers were
obtained from measurements of the three-bond scalar couplings including 3JNCc,3JC9Cc and 3JCaCd (refs 33, 34) 1H-15N RDCs were measured for the protein–
detergent complex weakly aligned in radially compressed polyacrylamide gel35.
Intramonomer NOEs involving both backbone and side-chain protons were
assigned using the three-dimensional 15N-edited and 13C-edited NOESYs
recorded with NOE mixing times of 110 and 150 ms, respectively, on a sample
containing 15N-, 13C-labelled protein, rimantadine, and deuterated DHPC
(D35-DHPC; Avanti Polar Lipids, Inc.).
For identifying contacts between adjacent monomers, intramonomer NOEs
were first assigned to completion. This is possible owing to the low complexity ofthe NOESY spectra of M2(18–60) (see Supplementary Figs 4 and 5). The remain-
ing NOEs involving residues of the transmembrane and amphipathic helices,
which could not be explained by intramonomer distances based on the known
secondary structures, were identified to be intermonomer NOEs. The assign-
ment of intermonomer distance restraints and structure calculation was
carried out iteratively until all NOE cross peaks in the NOESY spectra were
self-consistent in the tetrameric structure. Protein–drug NOEs were first iden-
tified using the 15N-edited and 13C-edited NOESYs described above, and sub-
sequently confirmed by a 15N-edited NOESY recorded with 500-ms mixing time
on a sample containing uniform 15N- and 2H-labelled protein, rimantadine, and
D35-DHPC.
Structure determination. Structures were calculated using the program
XPLOR-NIH36. The secondary structure of the monomer was first calculated
from its random coil using intramonomer NOEs, backbone dihedral restraints
derived from chemical shifts (TALOS)37 and side-chain x1 and x2 restraints
shown in Supplementary Table 2. A total of 20 monomer structures were
calculated using a standard high-temperature simulated annealing protocol in
which the bath temperature was cooled from 1,000 K to 200 K. To obtain aninitial set of tetramer structures, four copies of the lowest-energy monomer
structure calculated above were used. The same high-temperature simulated
annealing run was performed in the presence of intermonomer NOEs and all
other intramonomer restraints except RDCs. For each experimental intermono-
mer NOE between two adjacent subunits, four identical distance restraints were
assigned, respectively, to all pairs of neighbouring subunits to satisfy the con-
dition of C4 rotational symmetry. During the annealing run, the bath was cooled
from 1,000 K to 200 K with a temperature step of 20 K, and 6.7 ps of Verlet
dynamics at each temperature step, using a time step of 3 fs. A total of 100
tetramer structures were calculated and independently cross-validated by1H-15N RDCs.
Fitting of RDCs to structures was done by singular value decomposition using
the program PALES38. The goodness of fit was assessed by Pearson correlation
coefficient (r) and quality factor (Q). Among the 100 structural models, 15
structures in which the individual subunits have on average the best agree-
ment with RDCs (r of ,0.91 and Q of ,0.25) were selected for a final, low-
temperature refinement against RDCs in the presence of all NOE and dihedral
restraints. During the refinement, the bath was cooled from 200 K to 20 K with
a temperature step of 10 K, and 6.7 ps of Verlet dynamics at each temperature
step, using a time step of 3 fs. The force constants for NOE and experimental
dihedral restraints were fixed at 100 kcal mol21 A22 and 40 kcal mol21 rad22,
respectively. RDC restraint force constant was ramped from 0.01 to
0.125 kcal mol21 Hz22. The force constant for RDCs was set to a small value
to prevent violation of NOE restraints. For each of the 15 structures validated by
RDCs, 10 RDC-refined structures were generated. From that set, the structure
with the lowest total energy was added to the final ensemble to describe the
structural diversity of the solution structure. The structure with heavy atom
conformation closest to the mean was chosen to represent the final ensemble.
Without violating any NOE restraints, the final subunit structures fit RDCs to r
of 0.98 and Q of 0.15, with Da 5 14.2 Hz and Rh 5 0.24. Final backbone r.m.s.
deviation from the mean in the transmembrane and amphipathic domains were
0.30 A and 0.56 A, respectively.
Measurement of chemical exchange in the Trp 41 indole ring. The timescale
of chemical shift exchange of the Trp 41 side chain was measured using a
relaxation-compensated CPMG experiment17 in 1D mode at 1H frequency of
600 MHz. The dependence of 15N relaxation owing to chemical exchange on the
frequency of refocusing (1/tcp) of chemical shift evolution was fitted to a two-site
exchange model given by Rex!1{ 2tex
�tcp
� �tanh tcp
�2tex
� �, where Rex is the
contribution to transverse relaxation owing to chemical shift exchange, and tex is
the correlation time of the process that is generating the chemical shift
exchange39.
32. Blacklow, S. C. & Kim, P. S. Protein folding and calcium binding defects arisingfrom familial hypercholesterolemia mutations of the LDL receptor. Nature Struct.Biol. 3, 758–762 (1996).
33. Bax, A. et al. Measurement of homo- and heteronuclear J couplings fromquantitative J correlation. Methods Enzymol. 239, 79–105 (1994).
34. MacKenzie, K. R., Prestegard, J. H. & Engelman, D. M. Leucine side-chain rotamersin a glycophorin A transmembrane peptide as revealed by three-bond carbon-carbon couplings and 13C chemical shifts. J. Biomol. NMR 7, 256–260 (1996).
35. Chou, J. J., Gaemers, S., Howder, B., Louis, J. M. & Bax, A. A simple apparatus forgenerating stretched polyacrylamide gels, yielding uniform alignment of proteinsand detergent micelles. J. Biomol. NMR 21, 377–382 (2001).
36. Schwieters, C. D., Kuszewski, J., Tjandra, N. & Clore, G. M. The Xplor-NIH NMRmolecular structure determination package. J. Magn. Reson. 160, 66–74 (2002).
37. Cornilescu, G., Delaglio, F. & Bax, A. Protein backbone angle restraints fromsearching a database for chemical shift and sequence homology. J. Biomol. NMR13, 289–302 (1999).
38. Zweckstetter, M. & Bax, A. Prediction of sterically induced alignment in a diluteliquid crystalline phase: aid to protein structure determination by NMR. J. Am.Chem. Soc. 122, 3791–3792 (2000).
39. Allerhand, A. & Thiele, E. Analysis of Carr–Purcell spin-echo NMR experiments onmultiple-spin systems. II. The effect of chemical exchange. J. Chem. Phys. 45,902–916 (1966).
doi:10.1038/nature06531
Nature Publishing Group©2008

Schnell and Chou, “Structure and Mechanism of the M2 Proton Channel of Influenza A Virus” Supplementary Methods Sample preparation Validation of M2 construct and NMR sample condition. Several constructs of varying length, ranging from the TM peptide to the full-length protein, were tested for tetramerization and NMR spectral quality in different detergent solutions. The reconstituted full-length protein yields poor quality NMR spectra at physiologically relevant conditions, whereas the TM-only peptide, M2(18-46), does not assemble to a tetramer (as indicated by SDS-PAGE and chemical cross-linking). The region consisting of residues 18-60 [M2(18-60)], which includes the TM domain as well as 15 residues of the C-terminal region (Fig. S1a), forms a stable tetramer in dihexanoyl-phosphatidyl-choline (DHPC) detergent micelles while yielding high-resolution NMR spectra. At very low peptide concentrations (~20 μM), chemical cross-linking using dithiobis(succinimidyl)propionate (DSP) resulted in a homogeneous tetramer (Fig. S1b). At high protein concentrations used for NMR experiments (0.75 mM monomer), M2(18-60) runs as a homogeneous tetramer in SDS-PAGE in the absence of cross-linkers (Fig. S1c), indicating a very stable assembly. The reconstituted tetramer is pH-sensitive as lowering the pH led to dramatic changes in the NMR spectrum (Fig. 3a). The tetramer also interacts with the drug, rimantadine, as addition of the drug led to the specific perturbation of NMR resonances (Supplementary Fig. S2). The spectra do not change significantly between pH 7.4 and 8.3, indicating that the closed conformation is stable over this pH range. We therefore collected spectra for structure determination at pH 7.5 for the M2(18-60) tetramer in complex with rimantadine and DHPC micelles. The final sample condition, which yielded a high-resolution 1H-15N correlation spectrum (Fig. S1d), are described in the METHODS SUMMARY section. Identification protein-drug NOEs We first identified the rimantadine-binding site using a 13C-edited NOESY with 150 ms mixing time. Distinct rimantadine NOEs were present for the methyl groups of Leu42 and Ile43 (Fig. S3). We then independently confirmed the site by recording a 15N-edited NOESY with 500 ms mixing time on a sample containing uniform 15N- and 2H-labeled protein, deuterated detergent, and protonated rimantadine. This spectrum contains strong and weak crosspeaks to the two magnetically identical adamantane protons at 1.58 ppm for the backbone amides of Leu40 and Leu43, respectively (Fig. S3). The site assignment is consistent with the appearance of Ile43, Asp44, and Arg45 backbone amide resonances upon addition of rimantadine. We could not detect drug NOEs in other parts of the protein, including the widely proposed drug-binding site in the pore of the channel.
SUPPLEMENTARY INFORMATION
doi: 10.1038/nature06531
www.nature.com/nature 1

NMR spectroscopy NMR data analyses Data processing and spectra analyses were done in NMRPipe1 and CARA2. The program TALOS3 was used to predict backbone dihedral angles from characteristic chemical shifts. Fitting of residual dipolar couplings (RDCs) to structures was done by singular value decomposition (SVD)4, using the program PALES5. Analysis of the relaxation-compensated CPMG experiment6 was done using the program CPMGfit from Art Palmer (http://www.palmer.hs.columbia.edu/software/cpmgfit.html). Assignment of backbone and sidechain resonances Complete sequence specific assignment of backbone 1HN, 15N, 13Cα, and 13Cβ chemical shift were accomplished by performing a suite of standard triple resonance experiments, including the TROSY version of HNCA and HNCACB7, 8 on a 15N-, 13C, and 85% 2H-labeled protein sample at 600 MHz 1H frequency. Having the residue-specific chemical shift of 1HN and 15N, a 3D 15N-edited NOESY, recorded with 110 ms mixing time on a sample containing uniform 15N-, 13C-labeled protein, rimantadine, and deuterated DHPC (1,2-Dihexanoyl(D22)-sn-Glycero-3-Phosphocholine-1,1,2,2-D4-N,N,N-trimethyl-D9) (Avanti Polar Lipids, Inc.), was used to correlate the backbone amide and sidechain aliphatic and aromatic 1H resonances. Since the amide resonances are well resolved (see Fig. S1, pH 7.5) and all structured regions of M218-60 are α-helical, as indicated by 13Cα and 13Cβ chemical shifts (TALOS) and the characteristic local NOE patterns of α helix, assignment of intra-residue and sequential NOEs in the 15N-separated NOESY spectrum was straightforward (Fig. S2). Using the same approach, the assigned chemical shifts of aliphatic and amide protons were then used to assign the methyl 1H and 13C resonances, which are also mostly resolved in a 2D 1H-13C HSQC spectrum recorded with 28 ms constant-time (CT) 13C evolution (see Supplementary Fig. S4). This was accomplished using a 3D 13C-edited NOESY, recorded with 150 ms mixing time and 28 ms of CT 13C evolution on the same sample with deuterated detergent (Fig. S3). Specific stereo assignment of the gamma 13C of valine and delta 13C of leucine were obtained from a 10% 13C-labeled protein sample by recording a 1H-13C HSQC with 28 ms CT 13C evolution as previously described9. Measurement of 3-bond J couplings for determination of sidechain rotamers For determining whether the sidechain χ1 rotamers are trans for amino acids other than Thr, Val, and Ile, 3JNCγ coupling constants were measured using 2D spin-echo difference methods based on the 1H-15N constant-time TROSY experiment performed on 15N-, 13C-, and 85% 2H-labeled protein at 1H frequency of 750 MHz10, 11. For 3JC’Cγ and 3JNCγ of Thr, Val, and Ile, and 3JCαCδ of Leu and Ile, 2D spin-echo difference methods based on 1H-13C constant-time HSQC experiments were employed12-14. These spectra were recorded at 1H frequency of 600 MHz. All 3-bond J couplings are shown in Supplementary Table 2. Residual dipolar coupling measurements Weak alignment of the DHPC-reconstituted M218-20 relative to the magnetic field was accomplished using a modified approach15 of the strain-induced alignment in a gel (SAG) method16, 17. The protein/detergent solution was soaked into a cylindrically shaped polyacrylamide gel (4.5% acrylamide concentration and acrylamide/bisacrylamide molar ratio of 80), initially of 6 mm diameter and 9 mm length, which was subsequently radially compressed to fit within the 4.2 mm inner diameter of a NMR
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 2

tube with open ends (http://newera-spectro.com). The RDCs were obtained from subtracting J of the unaligned sample from J + D of aligned sample. The sign of dipolar couplings follows the convention that |1JNH + 1DNH| < 90 Hz when 1DNH is positive. The 1H-15N RDCs were obtained from 1JNH / 2 and (1JNH + 1DNH) / 2, which were measured at 600 MHz (1H frequency) by interleaving a regular gradient-enhanced HSQC and a gradient-selected TROSY18, both acquired with 80 ms of 15N evolution. On the basis of the length of the time domain data and the signal to noise ratio19, the accuracy of the measured RDCs is expected to be at ±0.5 Hz (1DNH). Structure calculation Structure calculation was done using the program XPLOR-NIH20. The secondary structure of the monomer was first calculated from random coil using intramonomer NOEs, backbone dihedral restraints derived from chemical shifts (TALOS)3 and sidechain χ1 and χ2 restraints shown in Supplementary Table 1. This was done using the following high-temperature simulated annealing (SA) protocol21. The intramonomer NOE restraints were enforced by flat-well harmonic potentials, with the force constant fixed at 50 kcal mol-1 Å-2. For sidechain χ1 and χ2 angles that are not assigned as rotamer averaging in Supplementary Table 1, flat-well (±30˚) harmonic potentials were applied with force constant fixed at 30 kcal mol-1 rad-2. Other force constants, commonly used in NMR structure calculation, were: k(vdw) = 0.02 4.0 kcal mol-1 Å-2, k(impr) = 0.1 1.0 kcal mol-1 degree-2, and k(bond angle) = 0.4 1.0 kcal mol-1 degree-2. During the annealing run, the bath was cooled from 1000 to 200 K with a temperature step of 20 K, and 6.7 ps of Verlet dynamics at each temperature step, using a time step of 3 fs. A total of 20 monomer structures were calculated using this protocol. The lowest energy structure was chosen for subsequent tetramer assembly.
To obtain an initial set of tetramer structures, four copies of the lowest-energy subunit structure calculated above were used. A high-temperature SA protocol similar to that of above was performed in the presence of intermonomer NOEs and all other intramonomer restraints except RDCs. For each experimental intermonomer NOE between two adjacent subunits, four identical distance restraints were assigned respectively to all pairs of neighboring subunits to satisfy the condition of C4 rotational symmetry. These restraints were enforced by flat-well (±0.6 Å) harmonic potentials, with the force constant ramped from 25 to 100 kcal mol-1 Å-2. During the annealing run, the bath was cooled from 1000 to 200 K with a temperature step of 20 K, and 6.7 ps of Verlet dynamics at each temperature step, using a time step of 3 fs. A total of 100 tetramer structures were generated for independent validation by RDCs.
The 100 structures calculated above were independently validated by 1H-15N RDCs. Fitting of RDCs to structures was done by singular value decomposition (SVD)4, using the program PALES5. The goodness of fit was assessed by both Pearson correlation coefficient (r) and the quality factor (Q)22. Among the 100 structural models, 15 structures of which the individual subunits have on average the best agreement with RDCs (r ~ 0.91 and Qfree ~ 0.25) have been selected for a second round, low-temperature refinement against RDCs in the presence of all other NOE and dihedral restraints. For RDC refinement in the XPLOR-NIH program, approximate initial values of the magnitude (Da) and rhombicity (Rh) of the alignment tensor must be specified. We initially used the average Da (14.0 Hz) and Rh (0.20) obtained from the best SVD fits. In theory, for a tetramer with a C4 rotational symmetry, the alignment tensor is axially symmetric23, or Rh = 0. However, this equation is only true in an ideal case for a completely rigid macromolecular assembly, but no longer holds in systems having some degrees of internal flexibility. In fact, the deviation from an axially symmetric tensor was also observed in the phospholamban homo-pentamer24. Since Rh ≠ 0, the RDCs assigned equivalently to the four
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 3

subunits could not be refined against a single alignment tensor. Instead, the RDCs of the four subunits were subject to four separate alignment tensors during XPLOR refinement.
During the refinement, the bath was cooled from 200 to 20 K with a temperature step of 10 K, and 6.7 ps of Verlet dynamics at each temperature step, using a time step of 3 fs. The force constants for NOE and experimental dihedral restraints were fixed at 100 kcal mol-1 Å-2 and 40 kcal mol-1 rad-2, respectively. In addition to the experimental χ1 and χ2 restraints, a weak database-derived ‘Rama’ potential function in XPLOR25 was ramped from 0.02 to 0.2 (dimensionless force constant) for the general treatment of sidechain rotamers. RDC restraint force constant was ramped from 0.01 to 0.125 kcal mol-1 Hz-2 (normalized for the 1DNH couplings). All other force constants were the same as before. For each of the 15 structures validated by RDCs, an ensemble of 10 RDC-refined structures were generated and the structure with the lowest total energy was chosen to represent the ensemble. In the end, we obtained 15 structures, each having the lowest energy in its corresponding ensemble, for describing the structural diversity of the NMR structure.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 4

Supplementary Table 1. NMR structural statistics and atomic r.m.s. differences1 Quantity Number of restraints Violations per structure
NOEs 257 0.0 (> 0.3 Å)
Intra-monomer 230 0.0 Inter-monomer 20 0.0 Drug-protein 7 0.0
Dihedral angle restraints 23 0.3 (> 5˚) χ1 15 0.3 χ2 8 0.0 Dipolar coupling restraints (Hz) 2 NH 27 1.04 +/- 0.09 Deviations from idealized covalent geometry Bonds (Å) 0.00187 +/- 0.00003 Angles (deg) 0.27 +/- 0.01 Impropers (deg) 0.40 +/- 0.01 Coordinate precision (Å) 3
All heavy atoms 1.25 Backbone heavy atoms 0.89 Ramachandran plot statistics (%) 4 Most favored regions 94.6 Additional allowed regions 5.4 1 Statistics are calculated and averaged over an ensemble of 15 lowest energy tetramers. The number of experimental restraints and violations given are per monomer. 2 Violations are given as the r.m.s. difference (in Hz) between individual sets of experimental dipolar couplings and those predicted by the 15 final structures by means of SVD fit. 3 The precision of the atomic coordinates is defined as the average r.m.s. difference between the 15 final structures and their mean coordinates. Unstructured residues at the terminii and the loop (residues 47-50) were excluded. 4 As evaluated for residues Leu26 to Leu59, excluding the flexible loop (residues 47-50) with the program PROCHECK26.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 5

Supplementary Table S2. Three-bond J couplings used for assigning sidechain χ1 and χ2 rotamers. The capital letter ‘A’ indicates rotameric averaging for which no dihedral restraints were used during structure calculation. Rotamer information was extracted from the couplings according to analyses previously described14, 27, 28.
Residue 3J(NCγ)
(Hz) 3J(C'Cγ)
(Hz) 3J(CδCα)
(Hz) χ1
(degrees) χ2
(degrees) Arg18 Cys19 Asn20 Asp21 Ser22 Ser23 Asp24 Pro25
Leu26-δ1 1.31 2.80 A A Leu26-δ2 2.84 Val27-γ1 2.35 0 180˚ Val27-γ2 0 2.82 Val28-γ1 1.16 0 180˚ Val28-γ2 0 3.40
Ala29 Ala30 Ser31
Ile32-γ2 2.15 0 -60˚ 180˚ Ile32-δ1 3.56 Ile33-γ2 2.44 2.07 -60˚ 180˚ Ile33-δ1 3.72 Gly34
Ile35-γ2 1.83 1.52 -60˚ 180˚ Ile35-δ1 3.26
Leu36-δ1 1.63 - 180˚ A Leu36-δ2 -
His37 1.52 180˚ Leu38-δ1 1.65 - 180˚ A Leu38-δ2 - Ile39-γ2 1.91 0 -60˚ 180˚ Ile39-δ1 3.58
Leu40-δ1 0.71 - A A Leu40-δ2 -
Trp41 2.63 180˚
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 6

Ile42-γ2 1.01 0.63 A 180˚ Ile42-δ1 4.14
Leu43-δ1 1.67 - 180˚ A Leu43-δ2 -
Asp44 Arg45 - A
Leu46-δ1 - 3.42 A 180˚ Leu46-δ2 -
Phe47 Phe48 Lys49 Ser50
Ile51-γ2 1.94 1.49 -60˚ 180˚ Ile51-δ1 3.40 Tyr52 0.90 A Arg53 1.32 A Phe54 1.33 A Phe55 0 A Glu56 1.42 A His57 0.84 A Gly58
Leu59-δ1 - - A A Leu59-δ2 -
Lys60 1.29 A
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 7

Supplementary Figure S1. Characterization of the M2(18-60) polypeptide construct. a, Amino acid sequence of M2 (A/Udorn). Residues 18 – 60 are underlined. Cysteines 19 and 50 were mutated to serines. The TM and AP helical regions are colored in blue and red, respectively. b, Urea-PAGE of reconstituted M2(18-60) at very low concentration (20 µM monomer) with and without chemical cross-linking. Lanes from left to right are: 1) molecular weight (MW) markers, 2) NMR sample without DSP cross-linker, 3) the sample in lane 2 subject to 15 min of DSP cross-linking reaction. c, SDS-PAGE of a typical NMR sample. Lanes from left to right are: 1) MW markers, 2) HPLC-purified M2(18-60) peptide dissolved directly into gel loading buffer solution followed by 5 min boiling, 3) NMR sample of M2(18-60) reconstituted using the protocol described in METHODS, 4) the sample used in lane 3 with rimantadine, and 5) the sample in 4 after 5 min of boiling, for demonstrating the stability of the tetrameric assembly. d, A 1H-15N transverse relaxation-optimized spectroscopy (TROSY) spectrum of uniform 15N-, 85% 2H-labeled M2(18-60) tetramer (0.75 mM monomer) in 300 mM DHPC and 40 mM rimantadine, recoreded at 600 MHz 1H frequency, pH 7.5, and 30 °C. Each peak represents a backbone NH moiety with residue number labeled.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 8

Supplementary Figure S2. The 1H-15N TROSY spectra of reconstituted M2(18-60) tetramer in the absence (black) and presence (red) of rimantadine at pH 6.0, 6.5, 7.0, and 7.5, recorded at 500 MHz 1H frequency and 30 °C.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 9

Supplementary Figure S3. Selected strips showing intermolecular NOEs between the protein and rimantadine. Experiments are (i) 15N-separated NOESY (500 ms NOE mixing) on 15N-, 2H-labeled M2(18-60), (ii) 13C-filtered, 13C-separated NOESY (200 ms mixing) on 15N-, 13C-labeled M2(18-60), (iii) 15N-separated NOESY (110 ms mixing), and (iv) 13C-separated NOESY (150 ms mixing).
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 10

Supplementary Figure S4. The 1H-15N strips corresponding to residues 26 – 46 from the 3D 15N-edited NOESY with water-gate readout pulse scheme, recorded at 1H frequency of 600 MHz on a sample containing 0.75 mM (monomer) M2(18-60), 40 mM rimantadine, 300 mM D35-DHPC, 40 mM sodium phosphate (pH 7.5) and 30 mM glutamate. The spectrum was acquired with 110 ms NOE mixing time, 36 ms of 15N evolution and 19 ms 1H evolution in the indirect dimension.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 11

Supplementary Figure S5. The 1H-13C strips corresponding to the methyl groups of residues 26 – 46 from the 3D 15C-edited NOESY with gradient coherence selection, recorded at 1H frequency of 600 MHz on a sample containing 0.75 mM (monomer) M2(18-60), 40 mM rimantadine, 300 mM D35-DHPC, 40 mM sodium phosphate (pH 7.5) and 30 mM glutamate. The spectrum was acquired with 150 ms NOE mixing time, 28 ms constant-time 13C evolution, and 26 ms 1H evolution in the indirect dimension.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 12

Supplementary Figure S6. The methyl regions of a high-resolution 1H-13C HSQC spectrum recorded with 28 ms of constant-time 13C evolution using a sample containing 0.75 mM (monomer) M2(18-60), 300 mM D35-DHPC, 40 mM rimantadine, and 30 mM glutamate in a 40 mM sodium phosphate buffer with a pH of 7.5. The labels on the spectrum are the complete assignments of methyl resonances.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 13

Supplementary Figure S7. Positions of known drug resistance mutations in the closed channel structure. Pore-lining residues are colored in cyan and residues that are in the helix-helix packing interface are colored in gray.
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 14

Supplementary References 1. Delaglio, F. et al. NMRPipe: a multidimensional spectral processing system based on UNIX
pipes. J. Biomol. NMR 6, 277-293 (1995). 2. Keller, R. (2004). 3. Cornilescu, G., Delaglio, F. & Bax, A. Protein backbone angle restraints from searching a
database for chemical shift and sequence homology. J. Biomol. NMR 13, 289-302 (1999). 4. Losonczi, J. A., Andrec, M., Fischer, M. W. F. & Prestegard, J. H. Order matrix analysis of
residual dipolar couplings using singular value decomposition. J. Magn. Reson. 138, 334-342 (1999).
5. Zweckstetter, M. & Bax, A. Prediction of sterically induced alignment in a dilute liquid crystalline phase: aid to protein structure determination by NMR. J. Am. Chem. Soc. 122, 3791-3792 (2000).
6. Loria, J. P., Rance, M. & Palmer, A. G. A relaxation-compensated Carr-Purcell-Meiboom-Gill sequence for characterizing chemical exchange by NMR spectroscopy. Journal of the American Chemical Society 121, 2331-2332 (1999).
7. Salzmann, M., Wider, G., Pervushin, K. & Wuthrich, K. Improved sensitivity and coherence selection for [N-15,H-1]-TROSY elements in triple resonance experiments. J. Biomol. NMR 15, 181-184 (1999).
8. Kay, L. E., Ikura, M., Tschudin, R. & Bax, A. Three-dimensional triple resonance NMR spectroscopy of isotopically enriched proteins. J. Magn. Reson. 89, 496-514 (1990).
9. Szyperski, T., Neri, D., Leiting, B., Otting, G. & Wuthrich, K. Support of 1H NMR assignments in proteins by biosynthetically directed fractional 13C-labeling. J. Biomol. NMR 2, 323-334 (1992).
10. Hu, J.-S., Grzesiek, S. & Bax, A. chi1 angle information from a simple two-dimensional NMR experiment which identifies trans 3JNCg couplings in isotopically enriched proteins. J. Biomol. NMR 9, 323-328 (1997).
11. Hu, J.-S., Grzesiek, s. & Bax, A. Two-dimensional NMR methods for determining c1 angles of aromatic residues in proteins from three-bond JC'Cg and JNCgcouplings. J. Am. Chem. Soc. 119, 1803-1804 (1997).
12. Vuister, G. W., Wang, A. C. & Bax, A. Measurement of three-bond nitrogen-carbon J couplings in proteins uniformly enriched in nitrogen-15 and carbon-13. J. Am. Chem. Soc. 115, 5334-35 (1993).
13. Grzesiek, S., Vuister, G. W. & Bax, A. A Simple and Sensitive Experiment for Measurement of Jcc Couplings Between Backbone Carbonyl and Methyl Carbons in Isotopically Enriched Proteins. J. Biomol. NMR 3, 487-493 (1993).
14. MacKenzie, K. R., Prestegard, J. H. & Engelman, D. M. Leucine side-chain rotamers in a glycophorin A transmembrane peptide as revealed by three-bond carbon-carbon couplings and 13C chemical shifts. J. Biomol. NMR 7, 256-260 (1996).
15. Chou, J. J., Gaemers, S., Howder, B., Louis, J. M. & Bax, A. A simple apparatus for generating stretched polyacrylamide gels, yielding uniform alignment of proteins and detergent micelles. J. Biomol. NMR 21, 377-382 (2001).
16. Tckyo, R., Blanco, F. J. & Ishii, Y. Alignment of biopolymers in strained gels: A new way to create detectable dipole-dipole couplings in high-resolution biomolecular NMR. J. Am. Chem. Soc. 122, 9340-9341 (2000).
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 15

17. Sass, H. J., Musco, G., Stahl, S. J., Wingfield, P. T. & Grzesiek, S. Solution NMR of proteins within polyacrylamide gels: Diffusional properties and residual alignment by mechanical stress or embedding of oriented purple membranes. Journal of Biomolecular Nmr 18, 303-309 (2000).
18. Weigelt, J. Single Scan, Sensitivity- and Gradient-Enhanced TROSY for Multidimensional NMR Experiments. J. Am. Chem. Soc. 120, 10778-9 (1998).
19. Kontaxis, G., Clore, G. & Bax, A. Evaluation of cross-correlation effects and measurement of one-bond couplings in proteins with short transverse relaxation times. J. Magn. Reson. 143, 184-196 (2000).
20. Schwieters, C. D., Kuszewski, J., Tjandra, N. & Clore, G. M. The Xplor-NIH NMR molecular structure determination package. J. Magn. Reson. 160, 66-74 (2002).
21. Nilges, M., Clore, G. M. & Gronenborn, A. M. Determination of three-dimensional structures of proteins from interproton distance data by dynamical simulated annealing from a random array of atoms. Circumventing problems associated with folding. FEBS Lett 239, 129-36 (1988).
22. Cornilescu, G., Marquardt, J. L., Ottiger, M. & Bax, A. Validation of protein structure from anisotropic carbonyl chemical shifts in a dilute liquid crystalline phase. J. Am. Chem. Soc. 120, 6836-37 (1998).
23. Jain, N. U., Noble, S. & Prestegard, J. H. Structural characterization of a mannose-binding protein-trimannoside complex using residual dipolar couplings. J. Mol. Biol. 328, 451-62 (2003).
24. Oxenoid, K. & Chou, J. J. The structure of phospholamban pentamer reveals a channel-like architecture in membranes. Proc Natl Acad Sci U S A 102, 10870-5 (2005).
25. Kuszewski, J., Gronenborn, A. M. & Clore, G. M. Improvements and extensions in the conformational database potential for the refinement of NMR and X-ray structures of proteins and nucleic acids. J. Magn. Reson. 125, 171-177 (1997).
26. Laskowski, R. A., MacArthur, M. W., Moss, D. S. & Thornton, J. W. PROCHECK: a program to check the stereochemical quality of protein structures. J. Appl. Cryst. 26, 283-291 (1993).
27. Bax, A. et al. Measurement of homo- and heteronuclear J couplings from quantitative J correlation. Methods Enzymol. 239, 79-105 (1994).
28. Chou, J. J., Case, D., Bax, A. Insights in mobility of methyl-bearing sidechains in proteins from Jcc and Jcn couplings. J. Am. Chem. Soc. 125, 8959-8966 (2003).
doi: 10.1038/nature06531 SUPPLEMENTARY INFORMATION
www.nature.com/nature 16