Embed Size (px)
Citation preview

JPET #184275
1
Distinct Effects of Imperatorin on Allergic Rhinitis: Imperatorin Inhibits
Caspase-1 Activity In Vivo and In Vitro System.
Hyun-A Oh, Hyung-Min Kim*, Hyun-Ja Jeong*
Department of Pharmacology, College of Oriental Medicine, Kyung Hee University,
Seoul,130-701, Republic of Korea (H.A.O., H.M.K); and Biochip Research Center, Hoseo
University 165, Sechul-ri, Baebang-myun, Asan, Chungnam, 336-795, Republic of Korea
(H.J.J)
JPET Fast Forward. Published on July 5, 2011 as DOI:10.1124/jpet.111.184275
Copyright 2011 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
2
Running title: Effect of imperatorin on allergic rhinitis
Address correspondence to: Dr. HJ Jeong ( ) and HM Kim
HJ Jeong, Biochip Research Center, Hoseo University 165, Sechul-ri, Baebang-myun, Asan,
Chungnam, 336-795, Republic of Korea, Tel.: +82 41 540 9681, Fax: +82 41 542 9681, e-
mail: [email protected].; HM Kim, Department of Pharmacology, College of Oriental
Medicine, Kyung Hee University, Seoul,130-701, Republic of Korea, e-mail:
The number of text page is 31
The number of figure is 8
The number of reference is 34
The number of words in the abstract is 249
The number of words in the introduction is 705
The number of words in the discussion is 634
ABBREVIATIONS: AR, allergic rhinitis; IPT, imperatorin; DEX, dexamethasone; OVA,
ovalbumin; HMC-1 cell, human mast cell line; IMDM, iscove’s modified dulbecco’s
medium; PBS, phosphate-buffered saline; PBST, 1 × PBS containing 0.05% tween-20;
PMACI, PMA and calcium ionophore A23187; ANOVA, analysis of variance.
A recommended section: Inflammation, Immunopharmacology, and Asthma
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
3
ABSTRACT
Because imperatorin (IPT), the furanocoumarins exhibits anti-inflammatory activity, we
reasoned that IPT might modulate the allergic rhinitis (AR). The aim of this study was to
analyze the regulation of AR by IPT. Here, we show the effect and mechanism of IPT in an
ovalbumin (OVA)-induced AR model. The number of rubs after the OVA challenge in the
OVA-sensitized mice was significantly higher than that in the OVA-unsensitized mice. The
increased number of rubs was inhibited by the oral administration of IPT. The increased
levels of IgE and histamine in the OVA-sensitized mice were reduced by IPT administration.
The levels of interferon-γ were enhanced while the levels of interleukin (IL)-4 were reduced
on the spleen tissue of the IPT-administered AR mice. Protein levels of IL-1β, macrophage
inflammatory protein-2, intercellular adhesion molecule -1, and cyclooxygenase-2 were
reduced by IPT administration in the nasal mucosa of the OVA-sensitized mice. In the IPT-
administered mice, the number of eosinophils and mast cells infiltration increased by OVA-
sensitization were also decreased. In addition, IPT inhibited caspase-1 activity in the same
nasal mucosa tissue. In activated human mast cells, the receptor interacting protein (RIP)2,
IκB kinase (IKK)-β, nuclear factor (NF)-κB/Rel A, and caspase-1 activation were increased,
but increased RIP2, IKK-β, NF-κB/Rel A, and caspase-1 activation were inhibited by the
treatment of IPT. In addition, IPT inhibited caspase-1 activity and IL-1β production in IgE-
stimulated bone marrow-derived mast cells. We can conclude that IPT exerts significant
effects by regulating of caspase-1 activation in AR animal and in vitro models.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
4
Introduction
Allergic rhinitis (AR) is a global health problem that causes major illness and disability
and common manifestation of allergic diseases, affecting approximately 500 million people
worldwide (Bousquet et al., 2008). Many of the symptoms of patients with AR, including
sneezing, itching, and respiratory obstruction cause a lot of pain. However, the symptoms of
AR do not end here. If prolonged, AR, can cause problems in the nasal voice box, and can
cause very severe eye and ear symptoms. These symptoms are due to the release of histamine
and other active substances by mast cells, which stimulate the dilation of blood vessels,
irritate nerve endings and increase the secretion of tears (Whitcup, 2006).
Since the discovery by Coffman and colleagues of two distinct types of Th in mice
(Mosmann et al., 1986), mutual regulation between Th1 cells and Th2 cells has been
considered important for homeostatic maintenance of the immune system in the whole body.
Dysregulated Th1 and Th2 responses lead to excessive Th1 cell or Th2 cell activation,
resulting in the development of autoimmune diseases associated with the accumulation of
Th1 cells or in an induction of allergic diseases due to the accumulation of Th2 cells,
respectively (Bach, 2002). In response to exposure to allergens, patients with AR present an
inflammatory Immunoglobulin (Ig)E-mediated response characterized by a Th2 immunologic
pattern with mast cells and eosinophils activation and the release of inflammatory mediators,
interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α (Johansson et al., 2001).
Leukotrienes and prostanoids produced by the 5-lipoxygenase and cyclooxygenase (COX)-2
pathways have potent pro-inflammatory and vascular actions that implicate them in the
allergic and inflammatory reactions (Montuschi et al., 2007). Eosinophils are innate effector
cells that are important in immune responses against helminth parasitic infections and
contribute to the pathology associated with allergic inflammatory conditions. Mast cells
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
5
contribute to the induction and/or maintenance of eosinophilic inflammation by a variety of
mechanisms, including IgE-dependent and IgE-independent processes (Pawankar et al., 2007).
The recruitment of these mast cells to inflammatory sites occurs in response to chemotactic
and activation signals (Bournazou et al, 2010). Minimal persistent inflammation is a
physiopathological phenomenon referring to the presence of an inflammatory cell infiltrate
(eosinophils and neutrophils) associated with the expression of intercellular adhesion
molecule-1 (ICAM-1) in the epithelial cells of the mucosa exposed to the allergen, in the
absence of clinical symptoms. ICAM-1 is still only expressed in the mucosal epithelial cells
of allergic patients. ICAM-1 was considered a marker of allergic inflammation (Montoro et al,
2007). Macrophage-inflammatory protein 2 (MIP-2) is a potent chemoattractant for immune
cells (Gupta et al., 1996).
Caspase-1 is a member of the cystein-aspartic acid protease (caspase) family (Stutz et al.,
2009). Caspase-1 is characterized by its ability to activate the inactive precursors of IL-1�
and IL-18 that are involved in inflammation. Caspase-1 contains an N-terminal caspase
recruitment domain (CARD). This CARD promotes the proteolytic activation of the recruited
caspase-1 in inflammation (Stutz et al., 2009). Caspase-1 is activated within inflammasome, a
large cytosolic protein complex that is induced by a growing number of endogenous,
microbial, chemical or environmental stimuli (Stutz et al., 2009). Specific adaptor molecules
of the receptor interacting protein-2 (RIP2, CARD containing kinase) regulate the activation
of caspase-1 through the CARD-CARD interaction (Kobayashi et al., 2002). RIP2 then
recruits the IκB kinase (IKK) complex through the direct interaction of its intermediate
domain with IKK-β, leading to the activation of the nuclear factor (NF)-κB (Inohara et al,
2000).
Imperatorin (IPT, Fig. 1A) is one of the furanocoumarins. Furanocoumarins are depicted
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
6
with miscellaneous biological functions including vasorelaxation of corpus cavernosum
(Chen et al., 2000), increased cell differentiation in osteoblasts (Kuo et al., 2005), anti-
convulsant (Luszczki et al., 2009), anti-diabetic (Liang et al., 2009), vascular vasodilation
(He et al., 2007), and reduction in liver steatosis (Ogawa et al., 2007). Lin et al.,
demonstrated that glutamate release was facilitated by IPT in rat hippocampal nerve terminals
(Lin et al., 2010). In addition, Adebajo et al., reported that the IPT had the main anti-
trichomonal activity (Adebajo et al., 2009).
This present study was designed to investigate the possibility of applying this IPT for the
regulation of AR. Furthermore, we aimed to validate a possible mechanism in the ovalbumin
(OVA)-induced AR models and activated mast cells.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
7
Material and methods
Materials. IPT, dexamethasone (DEX), OVA, phorbol 12-myristate 13-acetate (PMA),
A23187, O-phthaldialdehyde (OPA), avidin peroxidase (AP), 2'-azino-bis (3-
ethylbenzithiazoline-6-sulphonic acid) tablets substrate (ABTS), bicinchoninic acid (BCA),
anti-dinitrophenyl (DNP) IgE, DNP-human serum albumin (HSA), and other reagents were
purchased from Sigma (St. Louis, MO, USA). Fetal bovine serum (FBS), iscove’s modified
dulbecco’s medium (IMDM), and streptomycin were purchased from Gibco BRL (Grand
Island, NY, USA). Anti-mouse IgE/IL-1β (mature form detection Ab)/ΙL-4/IFN-
γ antibody (Ab), biotinylated anti-mouse IgE/IL-1β/IL-4/IFN-γ Ab, recombinant mouse (rm)
IgE/IL-1β/IL-4/IFN-γ, anti-human IL-1β Ab, biotinylated anti-human IL-1β Ab, and
recombinant human (rh) IL-1β were purchased from Pharmingen (Sandiego, CA, USA). Ab
for IKK-β, RIP2, caspase-1, COX-2, NF-κB/Rel A, IκBα, histon, and actin were obtained
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The mouse OVA-specific IgE kit
was purchased from DS Pharma Biomedical Co. Ltd. (Osaka, Japan) The caspase-1 assay kit
was supplied by R&D Systems Inc. (Minneapolis, MN, USA).
OVA-induced AR animal model. We maintained 6-week-old female BALB/c (Charles
River Technology) mice under pathogen-free conditions. Mouse care and experimental
procedures were performed under approval from the animal care committee of Kyung Hee
University [KHUASP (SE)-10-016]. The mice were sensitized on days 1, 5, and 14 by
intraperitoneal (i.p) injection of 100 μg OVA emulsified and 20 mg aluminum hydroxide
(Sigma) in a 100 μl phosphate-buffered saline (PBS) and challenged intranasally with 1.5 mg
OVA in 2 μl PBS or PBS. The mice were challenged intranasally with PBS in a similar
manner for the negative control. IPT (0.1 and 1 mg/kg), DEX (5 mg/ml), or a control vehicle
(distilled water.) was administrated orally for 10 days before the intranasal (i.n.) OVA
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
8
challenge (Fig. 1B). Bain et al., reported that OVA primed mice displayed a significant
increase in sneezing behavior when challenged intranasally (i.n.) with OVA (2011). In our
study, although the frequency of sneezing was increased by the OVA challenge, there were no
appreciable differences in the appearance of the mice between the OVA-unsensitized and
OVA-sensitized groups (data not shown). Nasal symptoms were only evaluated by counting
the number of nasal rubs that occurred in the 10 min after OVA i.n. provocation at the 10 day
mark after the challenge. We measured OVA-specific IgE as relevant end point for AR. The
numbers of mice in each group was 5.
Culture of HMC-1 cells. The human mast cell line (HMC-1) was generously provided by
Eichi Morri (Osaka University, Japan). HMC-1 cells were grown in IMDM supplemented
with 100 unit/ml penicillin, 100 mg/ml streptomycin and 10% heat-inactivated FBS at 37°C
5% CO2 and 95% humidity. HMC-1 cells (3 × 105 cells/ml) were treated with IPT (1 and 10
μg/ml) for 1 hr prior to stimulation with PMA and calcium ionophore A23187 (PMACI)
incubated for 2 hrs or 8 hrs.
Histamine assay. The histamine was measured from HMC-1 cells and serum according to
the manufacturer’s specifications using the histamine assay kit supplied by Oxford
Biomedical Research (Oxford, MI, USA).
Enzyme-linked immunosorbent assay (ELISA). HMC-1 cells (3 × 105) were treated
with IPT (1 and 10 μg/ml) for 1 h prior to stimulation with PMACI incubated for 8 h.
Cytokines of serum, mucosa, and spleen tissue, and supernatant were measured by an ELISA.
The ELISA was performed by coating 96-well plates with 1 μg/well of capture Ab. Before
the subsequent steps in the assay, the coated plates were washed twice with 1 × PBS
containing 0.05% tween-20 (PBST). All reagents and coated wells used in this assay were
incubated for 2 h at room temperature. The standard curve was generated from the known
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
9
concentrations of cytokine, as provided by the manufacturer. After exposure to the medium,
the assay plates were exposed sequentially to each of the biotin-conjugated secondary
antibodies, and an AP and ABTS substrate solution containing 30% H2O2. The plates were
read at 405 nm. Appropriate specificity controls were included, and all samples were run in
duplicate. The OVA-specific IgE was measured from the serum according to the
manufacturer’s specifications using an OVA-specific IgE kit. Cytokine levels in the spleen
and nasal mucosa were divided according to the total protein. The protein was estimated
using the BCA method with the BCA protein assay kit (Pierce, Rockford, IL, USA). This
method combines the reduction of cupric ions to cuprous ions by the protein in an alkaline
medium and then the subsequent reaction of the cuprous ions with two molecules of BCA to
give an intense purple color read at 560 nm.
Reverse transcription-polymerase chain reaction (RT-PCR). HMC-1 cells (3 × 106)
were treated with IPT (1 and 10 μg/ml) for 1 h prior to stimulation with PMACI incubated for
6 h. The total RNA was isolated from the cells and nasal mucosa according to the
manufacturer’s specification using an easy-BLUETM RNA extraction kit (iNtRON Biotech,
Korea). The concentration of total RNA in the final elutes was determined by
spectrophotometry. Total RNA (2.5 μg) was heated at 65°C for 10 min and then chilled on ice.
Each sample was reverse-transcribed to cDNA for 90 min at 37°C using a cDNA synthesis kit
(Amersham Pharmacia Biotech, Piscataway, NJ, USA). The PCR was performed with the
following primers for the mouse IL-1β (5’ AGG CCA CAG GTA TTT TGT CG 3’; 5’ GCC
CAT CCT CTG TGA CTC AT 3’), mouse GAPDH (5’TTC ACC ACC ATG GAG AAG GC
3’; 5’GGC ATG GAC TGT GGT CAT GA 3’), human IL-1β (5` GGG GTA CCT TAG GAA
GAC ACA AAT TG 3`; 5` CCG GAT CCA TGG CAC CTG TAC GAT CA 3`), and human
GAPDH (5` CCT GCT TCA CCA CCT TCT TG 3`; 5` CAA AAG GGT CAT CAT CTC TG
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
10
3`). GAPDH was used to verify whether equal amounts of RNA were used for reverse
transcription and PCR amplification from different experimental conditions. Saturation
curves for PCR were obtained from various experimental conditions (RNA concentrations,
annealing temperatures, and PCR cycle numbers). We determined the optimal amplification
conditions (annealing temperature and PCR cycle number) of primers for the PCR. The
annealing temperature was 50°C for mouse and human IL-1β and 60°C for mouse and human
GAPDH respectively. Products were electrophoresed on a 1.5% agarose gel and visualized by
staining with ethidium bromide.
Western blot analysis. HMC-1 cells (3 × 106) were treated with IPT (1 and 10 μg/ml) for
1 h prior to stimulation with PMACI incubated for 2 h. Western blot analysis was used for
nasal mucosa tissue extracts and cell extracts were prepared by a detergent lysis procedure.
Samples were heated at 95°C for 5 min, and briefly cooled on ice. Following the
centrifugation at 15,000 × g for 5 min, 50 μg aliquots were resolved by 10% SDS-PAGE. The
resolved proteins were electrotransferred overnight to nitrocellulose membranes in 25 mM
Tris, pH 8.5, 200 mM glycerin, 20% methanol at 25 V. Blots were blocked for at least 2 h
with PBST containing 5% nonfat dry milk and then incubated with primary antibodies for 1 h
at room temperature. Blots were developed by peroxidase-conjugated secondary antibodies,
and proteins were visualized by enhanced chemiluminescence procedures (Amersham
Bioseciences, Piscataway, NJ, USA) according to the manufacturer's instructions.
Histological examination. Tissue samples were immediately fixed with 10%
formaldehyde and embedded in paraffin. The sections of the nasal mucosa sample were 4 μm
thick. Each section was stained with hematoxylin and eosin (H&E, for eosinophils), alcian
blue and safranine O (A&S, for mast cells), CD4 (for T cells, Abbiotec, San diego, CA, USA),
F4/80 (for macrophages, eBioscience, San diego, CA, USA) or immunohistochemical stain
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
11
(for IL-1β) before dewaxing and dehydration. The numbers of eosinophils, mast cells, T cells,
macrophages, and IL-1β on both sides of the septal mucosa were counted. Sections were
coded and randomly analyzed by two blinded observers.
Myeloperoxidase (MPO) assay. To evaluate the effect of IPT on neutrophils infiltration,
the activity of tissue MPO was assessed. A biopsy was placed in 0.75 ml of 80 mM PBS, pH
5.4 containing 0.5% hexadecyltrimethylammonium bromide and homogenized (45 s at 0°C)
in a motor-driven homogenizer. The homogenate was decanted into a microfuge tube, and the
vessel was washed with a second 0.75 ml aliquot of hexadecyltrimethylammonium bromide
in a buffer. The wash was added to the tube, and the 1.5 ml sample was centrifuged at 12000
× g at 4°C for 15min. Samples of the resulting supernatant were added to 96-well microlitre
plates in triplicate at a volume of 30 μl. For the MPO assay, 200 μl of a mixture containing
100 μl of 80 mM PBS pH 5.4, 85 μl of 0.22 M PBS pH 5.4 and 15 μl of 0.017% hydrogen
peroxide were added to the wells. The reaction was started by the addition of 20 μl of 10 mM
O-dianisidine dihydrochloride in 80 mM PBS (pH 5.4). The plates were incubated at 37°C for
3 min and then placed on ice. The reaction was stopped by the addition of 30 μl of 1.46 M
sodium acetate, pH 3.0. Enzyme activity was determined colorimetrically using a late reader
set to measure absorbance at 460 nm and is expressed as OD mg per tissue.
Caspase-1 assay. HMC-1 cells (3 × 106) were treated with IPT (1 and 10 μg/ml) for 1 h
prior to stimulation with PMACI incubated for 2 h. The caspase-1 assay used nasal mucosa
tissue and cell extracts. Caspase-1 activity was measured according to the manufacturer’s
specifications using a caspase assay kit (R & D system). Equal amounts of the total protein
were quantified by a BCA protein quantification kit (Sigma) in each lysate. Catalytic activity
of caspase-1 from the cell lysate was measured by the proteolytic cleavage of WEHD-pNA or
YVAD-pNA (Biovision, Inc. California, USA) for 4 h or various times at 37°C. The plates
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
12
were read at 405 nm.
Transient transfection and luciferase assay. For the transfection, we seeded the HMC-1
cells (1 × 107) in a 100 mm culture dish. We then used Lipofectamine™ 2000 (Invitrogen,
Carlsbad, CA, USA) to transiently transfect reporter gene constructs into HMC-1 cells.
HMC-1 cells (3 × 106) were treated with IPT (1 and 10 μg/ml) for 1 h prior to stimulation
with PMACI incubated for 24 h. We mixed 20 μl of cell extract and 100 μl of the luciferase
assay reagent at room temperature. To measure the luciferase activity, we used a luminometer
(1420 luminescence counter, Perkin Elmer) in accordance with the manufacturer's protocol.
The transfection experiments were performed in at least three different experiments, all with
similar results. The relative luciferase activity was defined as the ratio of firefly luciferase
activity to renilla luciferase activity.
IKK-β assay. An IKK-β kinetic assay was performed according to the manufacturer’s
specifications using an IKK-β activity assay kit (EMD chemical, Gibbstown, NJ, USA).
Preparation of mouse bone marrow-derived mast cells (BMMCs). BMMCs were
generated from the femoral bone marrow cells of mice as described previously. Cells were
incubated in RPMI 1640 supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin,
100 µg/ml streptomycin, 100 μM 2-mercaptoethanol, 10 mM sodium pyruvate, 10 μM MEM
nonessential amino acid solution (Invitrogen), 100 U/m murine IL-3 (R&D Systems), and 0.5
U/ml murine stem cell factor (SCF; R&D Systems) at 37°C in a humidified atmosphere in the
presence of 5% CO2. After 4 weeks of being cultured, more than 96% of the cells were
identifiable as mast cells as determined by toluidine blue staining. BMMCs were treated with
anti-DNP IgE (10 μg/ml) and HSA (50 ng/ml).
Statistical analysis. The experiments shown are a summary of the data from at least three
experiments and statistical analyses were performed using SPSS statistical software (SPSS
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
13
11.5, USA). Treatment effects were analyzed by one-way ANOVA, offered by Tukey’s
multiple range tests, and P < 0.05 was used to indicate significance.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
14
Results
Effects of IPT on clinical symptoms and histamine, IgE, IL-4, IFN-γ, and IL-1β levels
in the AR model. To investigate the inhibitory effects of IPT in the AR model, we sensitized
mice on days 1, 5, and 14 by i.p. injections of 100 μg OVA emulsified in 20 mg aluminum
hydroxide and the challenged mice with 1.5 mg OVA. DEX (5 mg/kg) used as a positive
control. The numbers of nasal and ear rubs after the OVA challenge in the OVA-sensitized
mice were significantly higher than those in the OVA-unsensitized mice. Increased rub scores
were inhibited by treatment with IPT (Fig. 2A). The spleen weights after the OVA challenge
in the OVA-sensitized mice were significantly higher than those in the OVA-unsensitized
mice. Increased spleen weights were reduced by IPT administration (Fig. 2B). Histamine
levels in the serum were reduced by IPT (Fig. 2C). Levels of IgE in the AR mice were
significantly higher than those in the serum, spleen, and nasal mucosa tissues of the OVA-
unsensitized mice (Fig. 2D and E). Total IgE and OVA-specific IgE levels increased by the
OVA in the serum were reduced by IPT. To identify the Th1/Th2 immune reaction in IPT-
administered mice, we measured IL-4 and IFN-γ levels in the spleen. As shown in Fig. 2F
and G, the levels of IL-4 in the AR mice significantly increased compared to those in the
normal mice. IL-4 levels significantly decreased in the IPT-administered AR mice. However,
IFN-γ levels increased by OVA were not changed in the IPT-administered AR mice. The
protein levels of IL-1β in the serum and spleen tissue were increased in the AR mice
compared to those levels in the control mice (Fig. 2H). However, protein levels of IL-1β were
significantly reduced by IPT administration. IPT did not affect various allergic factors by
itself.
Effects of IPT on IL-1β, MIP-2, ICAM-1, and COX-2 levels in the nasal mucosa tissue
of the AR model. To evaluate the regulatory effect of IPT on IL-1β expression, we measured
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
15
the protein and mRNA levels of IL-1β in the AR model. The protein and mRNA levels of IL-
1β in the nasal mucosa tissue were increased in the AR mice compared to those levels in the
control mice (Fig. 3A and B). However, protein and mRNA levels of IL-1β were reduced by
IPT administration. Levels of MIP-2, ICAM-1, and COX-2 in the AR mice were significantly
higher than those in the nasal mucosa tissues of the OVA-unsensitized mice (Fig. 3C-F).
Increased levels of MIP-2, ICAM-1, and COX-2 were inhibited by IPT (Fig. 3C-F).
Effects of IPT on inflammation of nasal mucosa tissue. The respective numbers of
inflammatory cells (eosinophil, mast cell, neutrophils, macrophages, and T cells) in the nasal
mucosa in the AR mice were significantly higher than those in the control mice. In the IPT-
administered mice, the increase in eosinophil, mast cells, macrophages, and T cells
infiltration (MPO activation) by OVA sensitization was decreased. But IPT did not affect
neutrophil infiltration (Fig. 4B). Immunohistochemical analysis of the nasal mucosa sections
in the AR mice revealed that IL-1β is highly expressed; whereas in the IPT-administered mice,
it is decreased (Fig. 4A).
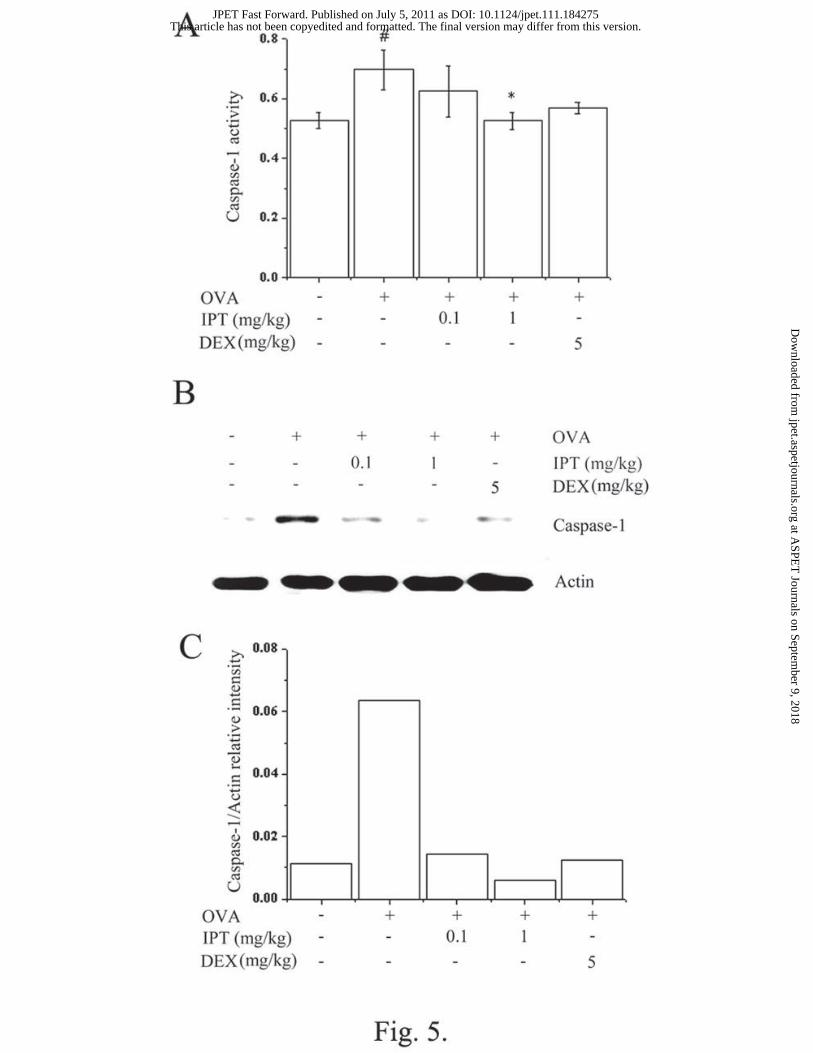
Effects of IPT on caspase-1 activation in the nasal mucosa tissues. Caspase-1 plays a
key role in inflammatory responses by cleaving pro-IL-1β into secreted pro-inflammatory
cytokines. To investigate the effect of IPT on caspase-1 activation, a caspase-1 assay was
performed with the nasal mucosa tissues. As shown in Fig. 5A, IPT (1 mg/kg) and DEX (5
mg/kg) inhibited OVA-induced caspase-1 activation. IPT also reduced the expression of
caspase-1 in the nasal mucosa tissue (Fig. 5B and C).
Effects of IPT on PMACI-induced IL-1β expression and histamine release in HMC-1
cells. Mast cells play a major role in the inflammatory reaction of AR (Pawankar et al., 2007).
Since IL-1β is a major cytokine released from mast cells after allergic responses, we
examined the effect of IPT on the expression of IL-1β in human mast cells. The protein and
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
16
mRNA levels of IL-1β were significantly inhibited by IPT (Fig. 6A and B). Also, the
inhibitory effects of IPT on PMACI-induced histamine release from HMC-1 cells are shown
in Fig. 6C. IPT did not affect IL-1β and histamine by itself. We examined cell viability using
a MTT assay and found that IPT had no effect on cell viability (Fig. 6D).
Effects of IPT on PMACI-induced NF-κB activation in HMC-1 cells. To assess the
regulatory mechanism of IPT on allergic inflammation in the in vitro model, we examined the
effect of IPT on PMACI-induced NF-κB activation, which is known to be important for
cytokine expression in HMC-1 cells. Because the suppression of NF-κB is linked with anti-
inflammation, we postulated that IPT mediates its effects at least partly through the
suppression of NF-κB activation. The pretreatment with IPT (1 and 10 μg/ml) inhibited the
PMACI-induced NF-κB/Rel A levels in the nuclear extract (Fig. 7A). As a marker of NF-κB
activation, the degradation of IκB-α in cell lysates was detected. We showed that IPT (1 and
10 μg/ml) inhibited the PMACI-induced IκB-α degradation (Fig. 7A). Histone and actin
expression levels were not changed by any treatment in the nuclear and cytoplasmic extracts.
We next examined whether IPT could modulate the luciferase expression specifically via the
NF-κB. The NF-κB luciferase reporter gene constructs (pNF-κB-LUC, plasmid containing
NF-κB binding site; STRATAGENE, La Jolla, CA, USA) were transiently transfected into
HMC-1 cells, which was treated by IPT and then stimulated by PMACI. As shown in Fig. 7B,
PMACI increased reporter gene activity. However, the increased activity was decreased by
the treatment of IPT (P < 0.05). IPT did not affect NF-κB activation by itself.
Effects of IPT on PMACI-induced RIP2/IKK-β/caspase-1 activation in HMC-1 cells.
RIP2 is a CARD-containing kinase that interacts with caspase-1 and plays an important role
in NF-κB activation (Kelsall, 2005). Apoptosis-associated speck-like protein containing a
CARD (ASC) is a PYRIN and CARD-containing molecule, important in the induction of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
17
apoptosis and caspase-1 activation (Sarkar et al., 2006). Expressions of RIP2 and IKK-β were
inhibited by IPT in HMC-1 cells (Fig. 8A). In the IKK-β assay, IKK-β activity was also
inhibited by IPT (Fig. 8B). To evaluate whether IPT regulated the caspase-1 activation, we
performed a Western blot analysis and caspase-1 assay. As shown in Fig. 8C and D, caspase-1
activity increased by PMACI was inhibited by IPT. A caspase-1 kinetic assay was used to
evaluate the binding affinity of IPT for the caspase-1 catalytic domain. Additionally,
recombinant caspase-1 (Biovision, Inc. California, USA) was also used to confirm the effect
of IPT in the kinetic assay. As shown in Fig. 8E, caspase-1 activity was increased after a
treatment with YVAD-pNA (caspase-1 substrate), but the IPT or caspase-1 inhibitor inhibited
the binding of YVAD-pNA with the caspase-1 catalytic domain. Interestingly, IPT (10 μg/ml)
exhibited a more potent binding affinity than did the caspase-1 inhibitor. In addition, caspase-
1 activity and IL-1β production increased by anti-DNP IgE and HSA were inhibited by IPT in
BMMCs (Fig. 8F and G). IPT did not affect the expression of RIP2, activation of IKK-β and
caspase-1, and production of IL-1β by itself.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
18
Discussion
In this study, IPT reduced the allergic inflammatory reaction in the AR model. AR is
characterized by a two-phase allergic reaction. In the early-phase inflammatory response
allergen-IgE dependent activation of mast cells and basophils results in the production of
pharmacologically active mediators such as histamine, prostaglandins, leukotrienes, and
cytokines which produce sneezing, rhinorrhea, and itching (Jeong et al., 2009). The late-
phase of AR show accumulations of mast cells, eosinophils, and basophils in the epithelium
and an accumulation of eosinophils in the deeper lamina propria (Fuentes-Beltrán et al, 2009).
Recruitment of inflammatory cells, including eosinophils, mast cells, basophils, and T cells,
results in the further release of histamine and leukotrienes, as well as in the release of other
compounds including proinflammatory cytokines, COX-2, and chemokines, which sustain the
allergic response and promote the late phase response (Fuentes-Beltrán et al., 2009; Fukui et
al., 2009). In previous studies, polyphenolic phytochemicals including rosmarinic acid have
been shown to inhibit the IgE response (Makino et al., 2001) and inflammation characterized
by polymorphonuclear leukocytes (eosinophils, and neutrophils) infiltration (Osakabe et al.,
2002). Glucocorticosteroid (GC) is the most effective drug for AR. GC inhibits the function
of infiltrating inflammatory cells and their recruitment into the nasal mucosa. GC inhibits the
maturation, cytokine production, COX-2 expression, FcεRI expression, and mediator release
of mast cells (Takano et al., 2004; Smith et al., 2002). We observed that IPT inhibited the IgE
production, inflammatory cytokine production, chemokine production, and COX-2
expression in the mice AR model. DEX also reduced the allergic and inflammatory reaction.
Therefore, our results suggest that the effect of IPT is similar to the mechanism of rosmarinic
acid or GC. We can also deduce that IPT has an anti-allergic effect.
Inflammasomes are multiprotein cytoplasmic complexes that mediate the activation of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
19
inflammatory caspase-1 (Stutz et al., 2009). Inflammasome regulates the activation and
secretion of caspase-1-regulated IL-1β and IL-18. Caspase-1–/– mice show the decreased
production of IL-6 after stimulation with lipopolysaccharide (Martinon, 2005). Grzegorczyk
et al., have reported a significant increase in caspase-1 level in serum from allergic asthmatic
patients as compared to a control group (Grzegorczyk et al., 2002). Caspase-1 activity was
increased in patients with the Netherton syndrome (Hosomi et al., 2008). In this study, we
observed that caspase-1 was activated in the AR mice. IPT inhibited caspase-1 activity and
IL-1β production. Therefore, we postulate that the inhibitory effect of IPT on inflammatory
cytokine production might be derived from the regulation of caspase-1 activation.
Mast cells arise from pluripotential stem cells and mature in the tissue. They have the
ability to generate immune reactions following exposure to a variety of receptor-mediated
signals initiated by both innate and acquired immune response mechanisms. Activated mast
cells release a broad spectrum of mediators including cytokines such as IL-1β (Johansson et
al., 2001; Howarth, 2003). Suppression of NF-κB activation has been linked with the
inhibition of inflammatory cytokine (IL-1β, IL-6, and TNF-α) expression (Inohara et al.,
2000). Activation by RIP2 induces caspase-1 oligomerization and promotes caspase-1
activation, with the latter inducing cytokine stimulation (Inohara et al., 2000). RIP2 and IKK
complexes may play an important role for NF-κB activation (Inohara et al., 2000). In this
study, IPT inhibited the IL-1β expression and production. We also confirmed that IPT
suppressed the RIP2/IKK-β/caspase-1 activation in mast cells. This result suggested that the
inhibitory effect of IPT on the allergic inflammatory reaction might be derived through the
regulation of RIP2/IKK-β/caspase-1 activation.
IPT has not been elucidated for the effect and mechanism on inflammatory reactions. We
first time observed that IPT can regulate the reduction of inflammatory cytokine expression
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
20
and the inhibition of caspase-1 activation, which causes the symptoms of AR, nasal itching
and eye's mucous membrane inflammation were found to alleviate symptoms. Therefore, the
author sees the potential as the drugs of IPT have on inhibitory action on AR.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
21
Authorship Contributions
Participated in research design: Kim HM and Jeong HJ.
Conducted experiments: Oh HA.
Performed data analysis: Jeong HJ and Oh HA.
Wrote or contributed to the writing of the manuscript: Kim HM, Jeong HJ, and Oh HA.
The authors have no conflicting financial interests.
`
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
22
References
Adebajo AC, Iwalewa EO, Obuotor EM, Ibikunle GF, Omisore NO, Adewunmi CO,
Obaparusi OO, Klaes M, Adetogun GE, Schmidt TJ, Verspohl EJ (2009) Pharmacological
properties of the extract and some isolated compounds of Clausena lansium stem bark:
anti-trichomonal, antidiabetic, anti-inflammatory, hepatoprotective and antioxidant effects.
J Ethnopharmacol 122:10-19.
Bach JF (2002) The effect of infections on susceptibility to autoimmune and allergic diseases.
N Engl J Med 347:911-920.
Bain G, Lorrain DS, Stebbins K, Broadhead A, Santini A, Prodanovich P, Darlington J, King
C, Lee C, Baccei C, Stearns B, Truong Y, Hutchinson J, Prasit P, Evans J (2011)
Pharmacology of AM211, a potent and selective DP2 receptor antagonist that is active in
animal models of allergic inflammation. J Pharmacol Exp Ther. Epub ahead of print.
Bournazou I, Mackenzie KJ, Duffin R, Rossi AG, and Gregory CD (2010) Inhibition of
eosinophil migration by lactoferrin. Immunol Cell Biol 88:220-223.
Bousquet J, Khaltaev N, Cruz AA, Denburg J, Fokkens WJ, Togias A, Zuberbier T, Baena-
Cagnani CE, Canonica GW, van Weel C, et al. (2008) World Health Organization; GA (2)
LEN; Allergen. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in
collaboration with the World Health Organization, GA (2) LEN and Allergen). Allergy
86:8-160.
Chen J, Chiou WF, Chen CC, and Chen CF (2000) Effect of the plant-extract osthole on the
relaxation of rabbit corpus cavernosum tissue in vitro. J Urol 163:1975-1980.
Fuentes-Beltrán A, Montes-Vizuet R, Valencia-Maqueda E, Negrete-García MC, García-Cruz
Mde L, and Teran LM (2009) Chemokine CC-ligand 5 production and eosinophil
activation into the upper airways of aspirin-sensitive patients. Clin Exp Allergy 39:491-499.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
23
Fukui N, Honda K, Ito E, and Ishikawa K (2009) Peroxisome proliferator-activated receptor
gamma negatively regulates allergic rhinitis in mice. Allergol Int 58:247-253.
Grzegorczyk J, Kowalski ML, Pilat A, and Iwaszkiewicz J (2002) Increased apoptosis of
peripheral blood mononuclear cells in patients with perennial allergic asthma/rhinitis:
relation to serum markers of apoptosis. Mediators Inflamm 11:225-233.
Gupta S, Feng L, Yoshimura T, Redick J, Fu SM, and Rose CE (1996) Intra-alveolar
macrophage-inflammation peptide 2 induces rapid neutrophil localization in the lung. Am J
Respir Cell Mol Biol 15:656-663.
He JY, Zhang W, He LC, and Cao YX (2007) Imperatorin induces vasodilatation possibly via
inhibiting voltage dependent calcium channel and receptor-mediated Ca2+ influx and
release. Eur J Pharmacol 573:170-175.
Hosomi N, Fukai K, Nakanishi T, Funaki S, and Ishii M (2008) Caspase-1 activity of stratum
corneum and serum interleukin-18 level are increased in patients with Netherton syndrome.
Br J Dermatol 159:744-746.
Howarth PH (2003) Allergic and nonallergic rhinitis. In: Middleton E, Reed C, Ellis E,
Adkinson N, Yunginger J, Busse W et al. Allergy, Principles and Practice, vol. 2, Allergy,
Principles and Practice. 6th edn. Philadelphia: Mosby 1391-410.
Inohara N, Koseki T, Lin J, del Peso L, Lucas PC, Chen FF, Ogura Y, and Núñez G (2000) An
induced proximity model for NF-kappa B activation in the Nod1/RICK and RIP signaling
pathways. J Biol Chem 275:27823-27831.
Jeong HJ, Moon PD, Kim SJ, Seo JU, Kang TH, Kim JJ, Kang IC, Um JY, Kim HM, Hong
SH (2009) Activation of hypoxia-inducible factor-1 regulates human histidine
decarboxylase expression. Cell Mol Life Sci 66:1309-1319.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
24
Johansson SG, Hiurihane JO, Bousquet J, Bruijnzeel-Koomen C, Dreborg S, Haahtela T,
Kowalski ML, Mygind N, Ring J, van Cauwenberge P et al. (2001) A revised nomenclature
for allergy: an EAACI position statement from the EAACI nomenclature task force.
Allergy 56:813-824.
Kobayashi K, Inohara N, Hernandez LD, Galán JE, Núñez G, Janeway CA, Medzhitov R,
Flavell RA (2002) RICK/Rip2/CARDIAK mediates signaling for receptors of the innate
and adaptive immune systems. Nature 416:194-199.
Kuo PL, Hsu YL, Chang CH, and Chang JK (2005) Osthole-mediated cell differentiation
through bone morphogenetic protein-2/p38 and extracellular signal-regulated kinase 1/2
pathway in human osteoblast cells. J Pharmacol Exp Ther 314:1290-1299.
Liang HJ, Suk FM., Wang CK, Hung LF, Liu DZ, Chen NQ, Chen YC, Chang CC, and Liang
YC (2009) Osthole, a potential anti-diabetic agent, alleviates hyperglycemia in db/db mice.
Chem Biol Interact 181:309-315.
Lin TY, Lu CW, Huang WJ, and Wang SJ (2010) Osthole or imperatorin-mediated facilitation
of glutamate release is associated with a synaptic vesicle mobilization in rat ippocampal
glutamatergic nerve endings. Synapse 64:390-396.
Luszczki JJ, Wojda E, Andres-MachM, Cisowski W, Glensk M, Glowniak K, and Czuczwar
SJ (2009) Anticonvulsant and acute neurotoxic effects of imperatorin, osthole and
valproate in the maximal electroshock seizure and chimney tests in mice: a comparative
study. Epilepsy Res 85:293-299.
Makino T, Furuta A, Fujii H, Nakagawa T, Wakushima H, Saito K, and Kano Y (2001) Effect
of oral treatment of Perilla frutescens and its constituents on type-I allergy in mice. Biol
Pharm Bull 24:1206-1209.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
25
Martinon F (2005) NLRs join TLRs as innate sensors of pathogens. Trends Immunol 26:447-
454.
Montuschi P, Sala A, Dahlen SE, and Folco G (2007) Pharmacological modulation of the
leukotriene pathway in allergic airway disease. Drug Discov Today 12:404-412.
Montoro J, Sastre J, Jáuregui I, Bartra J, Dávila I, A del Cuvillo, Ferrer M, Mullol J, and
Valero A (2007) Allergic rhinitis: Continuous or on demand antihistamine therapy? J
Investig Allergol Clin Immunol 17:21-27.
Mosmann TR, Cherwinski H, Bond MW, Giedin MA, and Coffman RL (1986) Two types of
murine helper T cell clone. I. Definition according to profiles of lymphokine activities and
secreted proteins. J Immunol 136:2348-2357.
Ogawa H, Sasai N, Kamisako T, and Baba K (2007) Effects of osthol on blood pressure and
lipid metabolism in stroke-prone spontaneously hypertensive rats. J Ethnopharmacol
112:26-31.
Osakabe N, Yasuda A, Natsume M, Sanbongi C, Kato Y, Osawa T, and Yoshikawa T (2002)
Rosmarinic acid, a major polyphenolic component of Perilla frutescens, reduces
lipopolysaccharide (LPS)-induced liver injury in D-galactosamine (D-GalN)-sensitized
mice. Free Radic Biol Med 33:798-806.
Pawankar R, Lee KH, Nonaka M, and Takizawa R (2007) Role of mast cells and basophils in
chronic rhinosinusitis. Clin Allergy Immunol 20:93-101.
Sarkar A, Duncan M, Hart J, Hertlein E, Guttridge DC, and Wewers MD (2006) ASC directs
NF-κB activation by regulating receptor interaction protein-2 (RIP2) caspase-1 interactions.
J immunol 176:4979-4986.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
26
Smith SJ, Piliponsky AM, Rosenhead F, Elchalal U, Nagler A, and Levi-Schaffer F (2002)
DEX inhibits maturation, cytokine production and Fc epsilon RI expression of human cord
blood-derived mast cells. Clin Exp Allergy 32:906-913.
Stutz A, Golenbock DT, and Latz E Inflammasomes: too big to miss. J Clin Invest 119:3502-
3511.
Takano H, Osakabe N, Sanbongi C, Yanagisawa R, Inoue K, Yasuda A, Natsume M, Baba S,
Ichiishi E, and Yoshikawa T (2004) Extract of Perilla frutescens enriched for rosmarinic
acid, a polyphenolic phytochemical, inhibits seasonal allergic rhinoconjunctivitis in
humans. Exp Biol Med 229:247-254.
Whitcup SM (2006) Recent advances in ocular therapeutics. Int Ophthalmol Clin 46:1-6.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
27
Footnotes
This research was supported by Basic Science Research Program through the National
Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and
Technology [2009-0090401].
*Kim HM and Jeong HJ contributed equally to this work.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
28
Legends for figure
Fig. 1. Structure of IPT (A) and Schematic protocols of AR animal model (B).
Fig. 2. Effects of IPT on clinical symptoms, spleen weight, and histamine, IgE, IL-4, IFN-γ,
and IL-1β levels in the AR model. We sensitized mice on days 1, 5, and 14 by intraperitoneal
injections of 100 μg OVA emulsified in 20 mg of aluminum hydroxide and we challenged
mice with 1.5 mg OVA. Mice received IPT before the intranasal OVA challenge for 10 days.
(A) The number of the nasal and ear rubs that occurred in the 10 min after the OVA intranasal
provocation. (B) Spleen weight, (C) Serum was isolated from blood and then assayed about
histamine. (D and E) IgE or OVA-specific IgE, (F) IL-4, (G) IFN-γ, and (H) IL-1β were
measured by the ELISA method. All parameters measured in the tissue homogenate were
presented as a ratio to the total protein levels in the tissue. .#P < 0.05; significantly different
from the OVA-unsensitized mice. *P < 0.05; significantly different from the OVA-sensitized
mice. N=5. DEX, dexamethasone.
Fig. 3. Effects of IPT on IL-1β, ΜIP-2, ICAM-1, and COX-2 expression in the nasal mucosa
of the AR mice. We sensitized mice on days 1, 5, and 14 by intraperitoneal injections of 100
μg OVA emulsified in 20 mg of aluminum hydroxide and we challenged the mice with 1.5
mg OVA. The mice received IPT for 10 days before the intranasal OVA challenge. (A) IL-1β,
(C) MIP-2, and (D) ICAM-1 were measured by the ELISA method in the nasal mucosa tissue.
(B) Messenger RNA was measured using the RT-PCR method. (E) COX-2 protein expression
was evaluated by using Western blot analysis. (F) The protein levels of COX-2 were
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
29
quantified by densitometry. #P < 0.05; significantly different from the OVA-unsensitized mice.
*P < 0.05; significantly different from the OVA-sensitized mice. N=5. DEX, dexamethasone.
Fig. 4. Effects of IPT on eosinophil and mast cell infiltration, and IL-1β expression in the AR
nasal mucosa tissue. (A) Nasal mucosa stained with H&E (for eosinophils = arrow head),
A&S (for mast cells = black arrow) and immunohistochemical DAB stain (for IL-1β = red
arrow). (B) Eosinophil, mast cells, neutrophil, macrophages, and T cells (MPO activity) were
counted by two individuals. After five randomly selected tissue sections per mouse were
counted. The absolute number of cells was counted as the mean ± standard error of the mean
(S.E.M.). #P < 0.05; significantly different from the OVA-unsensitized mice. *P < 0.05;
significantly different from the OVA-sensitized mice. DEX, dexamethasone. (Original
magnification × 400, scale bar=100 μm).
Fig. 5. Effects of IPT on caspase-1 activation in the nasal mucosa of the AR mice. We
sensitized mice on days 1, 5, and 14 by intraperitoneal injections of 100 μg OVA emulsified
in 20 mg of aluminum hydroxide and we challenged the mice with 1.5 mg OVA. Mice
received IPT for 10 days before the intranasal OVA challenge. (A) Protein was assayed about
caspse-1. (B) Caspase-1 protein expression was evaluated by using Western blot analysis. (C)
The protein levels were quantified by densitometry. Results are representative of three
independent experiments. #P < 0.05; significantly different from the OVA-unsensitized mice.
*P < 0.05; significantly different from the OVA-sensitized mice. N=5. DEX, dexamethasone.
Fig. 6. Effects of IPT on PMACI-induced IL-1β expression and histamine release in HMC-1
cells. (A) HMC-1 cells were treated with IPT (1 and10 μg/ml) for 1 h and then stimulated
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
30
with PMACI for 8 h. IL-1β was measured by the ELISA method. (B) Messenger RNA was
measured using the RT-PCR method. (C) Secreted histamine was assayed by a histamine
assay. (D) Cell viability was evaluated by a MTT assay. #P < 0.05; significantly different
from the unstimulated cells. *P < 0.05; significantly different from the PMACI-stimulated
cells. M, marker.
Fig. 7. Effects of IPT on PMACI-induced NF-κB activation in HMC-1 cells. (A) HMC-1
cells were treated with IPT (1 and 10 μg/ml) for 1 h and then stimulated with PMACI for 2 h.
Nuclear protein and cytoplasmic protein were prepared and analyzed for NF-κB and IκB-
α by Western blotting as described in the experimental procedures. Results are
representative of three independent experiments. (B) The NF-κB activity was assayed by a
luciferase assay. Values are the mean ± S.E.M. of duplicate determinations from three
separate experiments. #P < 0.05: significantly different from the unstimulated cells. * P <
0.05: significantly different from the PMACI-stimulated cells.
Fig. 8. Effects of IPT on PMACI-induced RIP2/IKK-β/caspase-1 activation in HMC-1 cells.
HMC-1 cells were pretreated with IPT for 1 h prior to PMACI stimulation for 2 hr. (A) RIP2
and IKK-β protein expressions were evaluated by using Western blot analysis. (B) An IKK-
β assay was performed. (C) The levels of caspase-1 were assayed by Western blot analysis.
(D) The enzymatic activity of caspase-1 was tested by a caspase-1 colorimetric assay. (E)
Catalytic activity of recombinant caspase-1 (8 units) was measured by YVAD-pNA
(caspase-1 substrate), IPT (1 and 10 μg/ml), or caspase-1 inhibitor (10 µM) for various
times. (F) The effect of IPT (10 μg/ml) on caspase-1 activity in IgE-stimulated BMMCs. (G)
The effect of IPT (10 μg/ml) on IL-1β production in IgE-stimulated BMMCs. Data represent
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #184275
31
the mean ± S.E.M. of the three independent experiments. #P < 0.05: significantly different
from the unstimulated cells. * P < 0.05: significantly different from the stimulated cells.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on July 5, 2011 as D
OI: 10.1124/jpet.111.184275
at ASPET Journals on September 9, 2018 jpet.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on July 5, 2011 as DOI: 10.1124/jpet.111.184275
at ASPE
T Journals on Septem
ber 9, 2018jpet.aspetjournals.org
Dow
nloaded from