Embed Size (px)
Citation preview

Caspase-mediated apoptosis and caspase-independent celldeath induced by irofulven in prostate cancer cells
Huiyun Liang,1 Richard A. Salinas,1
Belinda Z. Leal,1 Teresa Kosakowska-Cholody,2
Christopher J. Michejda,2 Stephen J. Waters,3
Terence S. Herman,1 Jan M. Woynarowski,1 andBarbara A. Woynarowska1
1Department of Radiation Oncology, University of TexasHealth Science Center, San Antonio, Texas; 2National CancerInstitute, Frederick, Maryland; and 3MGI Pharma, Inc.,Bloomington, Minnesota
AbstractIrofulven (hydroxymethylacylfulvene) is a novel antitumordrug, which acts by alkylating cellular macromolecular tar-gets. The drug is a potent inducer of apoptosis in varioustypes of tumor cells, whereas it is nonapoptotic in normalcells. This study defined molecular responses to irofulveninvolving mitochondrial dysfunction and leading to deathof prostate tumor LNCaP-Pro5 cells. Irofulven caused early(2–5 hours) translocation of the proapoptotic Bax fromcytosol to mitochondria followed by the dissipation ofmitochondrial membrane potential and cytochrome c re-lease at 4 to 12 hours. These effects preceded caspaseactivation and during the first 6 hours were not affectedby caspase inhibitors. Processing of caspase-9 initiated thecaspase cascade at f6 hours and progressed over time.The activation of the caspase cascade provided a positivefeedback loop that enhanced Bcl-2-independent translo-cation and cytochrome c release. General and specificcaspase inhibitors abrogated irofulven-induced apoptoticDNA fragmentation with the following order of potency:pan-caspase z caspase-9 > caspase-8/6 > caspase-2 >caspase-3/7 > caspase-1/4. Abrogation of caspase-mediated DNA fragmentation failed to salvage irofulven-treated cells from growth inhibition and loss of viability,demonstrating a substantial contribution of a caspase-
independent cell death. Monobromobimane, an inhibitor ofalternative caspase-independent apoptotic pathway that ismediated by mitochondrial permeability transition, antag-onized both apoptosis, measured as phosphatidylserineexternalization, and cytotoxicity of irofulven. Collectively,the results indicate that irofulven-induced signaling isintegrated at the level of mitochondrial dysfunction. Theinduction of both caspase-dependent and caspase-inde-pendent death pathways is consistent with pleiotropic ef-fects of irofulven, which include targeting of cellular DNAand proteins. [Mol Cancer Ther 2004;3(11):1385–96]
IntroductionApoptosis induction is a desirable property for variousanticancer drugs including those that target cellular DNA.Apoptosis induction in response to DNA-targeting drugshas been extensively studied and the role of DNA damageas a significant proapoptotic stimulus is well established(1). It is less understood what factors are responsible forthe high variability of the timing and magnitude of apo-ptosis induction among different DNA-reactive drugs andin various cell models. We have postulated that such dif-ferences could reflect in part the interplay of DNA dam-age with drug effects on other cellular targets (2, 3). Thisidea takes into account the fact that various anticancerdrugs classified as DNA-damaging alkylating agents tendto react mainly with cellular proteins, although their DNAadducts constitute only a small proportion of total cellularadducts (4, 5).
One of the novel anticancer drugs that targets both DNAand proteins is a semisynthetic sesquiterpenoid irofulven(hydroxymethylacylfulvene, MGI 114, or NSC 683863;Fig. 1; refs. 2, 3, 5, 6). Irofulven, which is currently underinvestigation in several phase I and II clinical trials (7, 8),shows in vivo antitumor activity against a broad spectrumof human tumor xenografts (9–13). In addition, the drugremains efficacious against various models of multidrug-resistant tumors (11, 14). Tumor shrinkage and completecurative effects were frequently observed (12, 13).
Consistent with its ability to cause tumor regression(12, 13), irofulven exhibits potent proapoptotic properties invarious tumor cell systems (15–17), including models withapoptotic deficiencies, such as the lack of caspase-3 (18) andoverexpression of antiapoptotic Bcl-2 (19). In contrast, sev-eral normal cell lines tested showed remarkable resistanceto apoptosis induction by irofulven despite the formationof drug-macromolecule adducts at levels comparable withthose in tumor cells (16, 20). For equivalent cellular druguptake, irofulven was 3- to 4-fold less growth inhibitoryto normal cells than to tumor cells (20). Moreover, drug-treated normal cells remained viable (nonapoptotic) andcapable of resuming growth after a lag period (16, 20).
Received 7/8/04; revised 8/23/04; accepted 9/15/04.
Grant support: National Cancer Institute grants CA78706 andCA112175, and MGI Pharma, Inc.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
Note: A preliminary account of this study has been presented in part atthe 92nd Annual Meeting of AACR, March 24–28, 2001, New Orleans,Proc. AACR 42, 2001, 640.
Requests for reprints: Jan M. Woynarowski, Department of RadiationOncology, University of Texas Health Science Center, IDD Building,14960 Omicron Drive, San Antonio, TX 78245. Phone: 210-677-3832;Fax: 210-677-0058. E-mail: [email protected]; or Barbara A. Woynarowska,Phone: 210-677-3846; Fax: 210-677-0058. E-mail: [email protected]
Copyright C 2004 American Association for Cancer Research.
Molecular Cancer Therapeutics 1385
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

Biological activities of irofulven seem to be related to thealkylating properties of this drug (5, 21–23). Although thedrug molecule has several potentially reactive electrophiliccenters (21, 22), irofulven forms probably only monoad-ducts with cellular DNA (3, 5). DNA monoadducts are, ingeneral, considered low lethality lesions (24). Accordingly,unlike irofulven, other DNA-reactive agents of clinicalanticancer significance form either interstrand or intra-strand cross-links or double-strand breaks (24). In additionto DNA reactivity, however, irofulven readily alkylatessulfhydryl groups (5, 21–23). In cellular systems, covalentbinding of irofulven to protein sulfhydryls exceeds 3- to 10-fold drug binding to DNA (5, 25). This substantial proteinbinding, which notably involves targeting of key redoxcontrolling proteins, could explain the ability of irofulvento distort protein redox homeostasis (2, 25).
Mitochondria are believed to play a central role indetermining cell survival or death in response to diverseapoptotic stimuli including agents that cause either DNAdamage or protein damage or distort protein redox status(1, 26–28). Responses triggered by DNA damage ofteninvolve a shift in the balance between proapoptotic andantiapoptotic members of the Bcl-2 family, which in turnincreases the permeability of the outer mitochondrialmembrane (26–29). The resulting release of cytochromec and other apoptogenic components initiates caspase acti-vation and triggers caspase-mediated apoptotic DNA frag-mentation and eventually cell death (29). The Bax-mediatedouter membrane permeabilization and the downstreamcaspase-mediated cell death tend to be inhibitable by Bcl-2overexpression (19, 30).
In addition to the classic caspase-mediated apoptosis,mammalian cells can undergo caspase-independent apo-ptosis that is mediated by the dissipation of the innermitochondrial membrane potential DCm (inner membranepermeability transition, MPT) and the release of apoptosis-inducing factor (AIF; refs. 28, 31). The caspase-independent,AIF-mediated cell death pathway has been documentedin various death models including exposure to someagents that affect protein redox status (28). Specifically,oxidative cross-linking of redox-sensing inner membraneproteins was found to directly cause Bcl-2-independentMPT and AIF release (28, 31–33). The triggering of caspase-independent mechanisms may circumvent the apo-ptotic resistance of those cancer cells that are insensitiveto DNA damage induced by ionizing radiation andetoposide (34).
We hypothesized that the consequences of irofulvenreactivity with its diverse cellular targets converge andintegrate at the level of mitochondrial dysfunction. Tobetter understand the nature of irofulven-induced apo-ptosis, we attempted to discern the primary apoptotic re-sponses from the downstream effects and to identifyfactors/pathways that determine the fate of irofulven-treated cancer cells. Using prostate tumor cells LNCaP-Pro5, we show that irofulven-induced apoptosis involvesearly translocation of proapoptotic Bax from cytosol tomitochondria followed by mitochondrial dysfunction,cytochrome c release, caspase activation, and DNA frag-mentation. Although caspase inhibition abrogates DNAfragmentation, it does not salvage irofulven-treated cellsfrom growth inhibition and loss of viability. In contrast, asubstantial reduction of irofulven-induced apoptosis andantagonized cytotoxicity is observed after blocking mito-chondrial proteins involved in MPT. Thus, mitochondrialdysfunction seems to reflect a decisive point in the actionof irofulven, resulting in cancer cell death via an interplayof caspase-dependent and caspase-independent routes.
Materials andMethodsCell Culture and DrugTreatmentLNCaP-Pro5 cells (from Dr. C.A. Pettaway, MD Ander-
son Cancer Center, Houston, TX) were maintained in RPMI1640 supplemented with 10% FCS, 2% MEM vitamin solu-tion (Life Technologies, Grand Island, NY), 1% nonessentialamino acids, sodium pyruvate (1 mmol/L), and glutamine(2 mmol/L) as described previously (16, 20). Cell numberswere determined by cell counting in a hemocytometer orin a model ZM Coulter counter (Beckman Coulter, Inc.,Fullerton, CA). Cell viability (cell membrane integrity) wasdetermined based on cell ability to exclude trypan blue.Stock solution of irofulven (MGI Pharma, Inc., Blooming-ton, MN) was prepared in DMSO and stored at �20jCprotected from light. Unless stated otherwise, cells weretreated for 4 hours with 10 Amol/L drug. At the end ofdrug treatment, cell monolayers were either harvested orwashed twice with prewarmed medium and postincubatedin fresh drug-free medium as indicated. When caspaseinhibitors were used, they were added to cell cultures 1hour prior to drug addition and were present during drugtreatment as well as after incubation in irofulven-freemedium. Inhibitors of caspase-1/4 [acetyl-Tyr-Val-Ala-Asp-chloromethyl ketone (Ac-YVAD-cmk)], caspase-2[N-benzyloxycarbonyl-Val-Ala-Val-Ala-Asp(OMe)-fluoro-methyl ketone (Z-VDVAD-fmk)], caspase-3/7 [acetyl-Asp-Glu-Val-Asp-chloromethyl ketone (Ac-DEVD-cmk)],caspase-9 [N -benzyloxycarbonyl-Leu-Glu(OMe)-His-Asp(OMe)-fluoromethyl ketone (Z-LEHD-fmk)], and thebroad-spectrum inhibitor, N-benzyloxycarbonyl-Val-Ala-Asp(OMe)-fluoromethyl ketone (Z-VAD-fmk), were fromAlexis Biochemicals (San Diego, CA). Caspase-8/6 inhibitor[N -benzyloxycarbonyl-Ile-Glu(OMe)-Thr-Asp(OMe)-fluoromethyl ketone (Z-IETD-fmk)] was purchased fromBIOMOL Research Labs (Plymouth Meeting, PA).
Figure 1. Structure of irofulven.
Pathways of Cell Death by Irofulven1386
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

Cytotoxic ActivityGrowth inhibitory activity was assayed as described
previously (15) using the standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Expo-nentially growing cells were incubated with the drug forthe indicated time and subjected to colorimetric reactionwith MTT. The combinations of irofulven and inhibitorswere evaluated for synergy/antagonism according to themedian effect method of Chou and Talalay (35) usingCalcuSyn program.BaxTranslocation and Cytochrome c ReleaseAt the end of treatments, attached cells harvested by
gentle trypsinization were combined with detached cellsand washed twice by centrifugation (5 minutes at 200 � g)with ice-cold PBS. The cytosolic and mitochondrial frac-tions were prepared according to Kluck et al. (36) withslight modification. Briefly, cell pellets were suspended(107 cells/200 AL) in isotonic buffer containing 10 mmol/LHEPES (pH 6.9), 200 mmol/L mannitol, 70 mmol/Lsucrose, 1 mmol/L EGTA, 100 Amol/L phenylmethylsul-fonyl fluoride, 10 Ag/mL aprotinin, and 10 Ag/mLleupeptin, and incubated on ice for 15 minutes followedby homogenization in a Dounce homogenizer (20 strokeswith type B pestle). After centrifugation at 1,000 � g for10 minutes, supernatants were recentrifuged at 10,000 � gfor 20 minutes at 4jC to pellet mitochondria. Thepostmitochondrial supernatants (cytosolic fractions) weresequentially passed through 0.2 and 0.1 Am filter toremove possible contaminating membrane fragments (36).The mitochondrial pellets were washed once with theisotonic buffer and solubilized in radioimmunoprecipita-tion assay buffer [1% NP40, 0.5% sodium deoxycholate,0.1% SDS in PBS (pH 7.2)] supplemented with proteaseinhibitors (100 Amol/L phenylmethylsulfonyl fluoride,10 Ag/mL aprotinin, 10 Ag/mL leupeptin). Protein con-centrations in cytosol were determined by Bradford assayusing bovine serum albumin as a standard, whereasthe concentrations of mitochondrial proteins in thepresence of 0.6% SDS were determined spectrophotomet-rically at 280 nm (assuming that A280 = 0.21 correspondsto 10 mg/mL protein).
Both cytosolic and mitochondrial fractions containingequal amounts of protein were subjected to reducing SDS-PAGE, transferred to nitrocellulose, and probed withantibodies specific for Bax (rabbit anti-human, 1:1,000,PharMingen, San Diego, CA) or cytochrome c (mouse anti-human, 2 Ag/mL, PharMingen) for 2 hours at roomtemperature. Following additional 1-hour incubation withhorseradish peroxidase–conjugated secondary antibodies(goat anti-rabbit IgG 1:500, rabbit anti-mouse IgG 1:2,000),specific protein bands were visualized by chemiluminescenceusing enhanced chemiluminescence system (AmershamBiosciences, Piscataway, NJ). Bands recorded on X-ray filmwere scanned using a Molecular Dynamics densitometer(Sunnyvale, CA) and their intensity was quantitated withIQ software (Molecular Dynamics). Bcl-2-independent andcytochrome c signals were normalized to the h-actin signalin the analyzed samples, which was also detected by
Western blotting. The integrity of the isolated mitochondriapreparations was assessed by the lack of release ofcytochrome c oxidase detected with appropriate antibodyin the cytosolic fractions.
Caspase ProcessingThe cleavage of pro-caspases from inactive to active
forms was monitored by Western blotting, similar to theprocedure described for Bcl-2-independent determinations.After treatment with drug as indicated, cells wereharvested as described for Bax translocation determina-tions, washed with ice-cold PBS, and resuspended inHEPES (25 mmol/L, pH 7.4) containing 0.1% Triton X-100, 10% glycerol, DTT (5 mmol/L), and protease inhibitors(1 mmol/L phenylmethylsulfonyl fluoride, 10 Ag/mLleupeptin, 10 Ag/mL aprotinin). After 10 minutes ofincubation on ice followed by three cycles of freezing-thawing (in liquid nitrogen and at 37jC), samples werecentrifuged at 10,000 � g for 10 minutes at 4jC. Equalamounts of proteins were resolved by reducing SDS-PAGE.After protein transfer, nitrocellulose membranes wereprobed with the antibodies against human caspase-9(mouse IgG, 1 Ag/mL, Oncogene Research Products,Boston, MA), caspase-7 (mouse IgG, 1 Ag/mL, PharMin-gen), caspase-8 (mouse IgG, 1 Ag/mL, Oncogene), andcaspase-3 (rabbit polyclonal, 1:1,000, PharMingen). Follow-ing incubation with respective horseradish peroxidase–conjugated secondary antibodies, caspase were visualizedusing enhanced chemiluminescence system and their signalquantitated as described for Bax.DEVDCleavageActivityA spectrophotometric assay was used to measure the
release of p-nitroanilide (pNA) from peptide conjugateAc-DEVD-pNA (Alexis Biochemicals), a substrate forcaspase-3 and caspase-7. Cell extracts obtained as describedabove for caspase processing were used as enzyme sourcefor these determinations. Reactions were carried out inPIPES (20 mmol/L, pH 7.2) containing NaCl (100 mmol/L),10% sucrose, 0.1% Triton X-100, and h-mercaptoethanol(20 mmol/L). The reaction mixture containing Ac-DEVD-pNA (150 Amol/L) and cellular extract (40 AL) in the finalvolume of 100 AL was incubated at 37jC and absorbance at405 nm was read after 0, 1, 2, and 3 hours of incubation.Parallel reactions containing activated caspase-3 (AlexisBiochemicals) were used as positive controls and DEVDproteolytic activity inhibitor (Ac-DEVD-cmk, 10 Amol/L)was used to assess nonspecific hydrolysis. The results areexpressed as nanomoles of pNA per milligram of proteinper hour with pNA as a standard.
Flow Cytometry Determinations of DCm
DCm was determined as described previously (19)using the lipophilic cationic probe 5,5V,6,6V-tetrachloro-1,1V,3,3V-tetraethylbenzimidazolyl-carbocyanine iodide (JC-1),which emits red fluorescence when bound to mitochondrialmembranes with high DCm and green fluorescence in cellswith depolarized mitochondrial membranes (37).
Briefly, irofulven-treated cells were harvested as de-scribed for Bax determinations and washed with PBS andwith serum-free RPMI 1640 without phenol red and
Molecular Cancer Therapeutics 1387
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

glutamine. Finally, cells were resuspended in PBS. Aliquotsof cell suspension were incubated for 25 minutes with JC-1containing binding buffer (ApoAlert Mitochondrial Mem-brane Sensor, BD Biosciences Clontech, Palo Alto, CA) asper manufacturer’s protocol and analyzed immediately byflow cytometry on a Coulter EPICS ELITE flow cytometer.Typically, 10,000 events were collected using excitation/emission wavelengths of 488/525 and 488/675 nm forgreen and red fluorescence, respectively. Events with highred and high green fluorescence were gated as measuresof cells with high and low DCm, respectively.
AnnexinVBindingPhosphatidylserine externalization, a marker of early
apoptotic events, was detected by binding of FITC-conjugated Annexin V, whereas counterstaining withpropidium iodide (PI) allowed for the detection of cellswith permeable cell membrane (19). Cells were treated withirofulven alone or in combination with inhibitors asindicated and harvested by trypsinization, washed withserum-free medium, and suspended in PBS at the density1 � 106 cells/mL. Aliquots of 1 � 106 cells were suspendedin binding buffer (500 AL, Annexin V-FITC staining kit,PharMingen). This cell suspension (100 AL) was stainedwith mouse anti-human Annexin V antibody (mIgG type,5 AL) and PI (500 Ag/mL, 10 AL) for 15 minutes in thedark. The immunostaining was terminated by addition ofbinding buffer and cells were immediately analyzed byflow cytometry. The negative control included stainingwith mIgG only. Typically, 10,000 events were collectedusing excitation/emission wavelengths of 488/525 and488/675 nm for Annexin V and PI, respectively.
QuantitativeApoptotic DNAFragmentationThe quantitative DNA fragmentation assay detects both
early, high molecular weight DNA fragments and lateresidual oligonucleosomal-sized fragments (15, 16, 19, 20).Briefly, 14 C-thymidine-prelabeled cells treated with drugas indicated were permeabilized in a hypotonic bufferfollowed by the extraction of fragmented DNA andcollection of these fragments by centrifugation in respectivesupernatants, whereas nondegraded DNA remains in thenuclear pellet fraction. The results are expressed as thepercentage of the total DNA released in the supernatants,corrected for the radioactivity released from untreatedcontrols. Irofulven-generated double-stranded fragmentswere shown previously to peak at f40 to 50 kbp after a24-hour incubation of drug-treated cells (18, 38).
ResultsThis study centered on mitochondrial dysfunction and itsconsequences in the model of prostate cancer LNCaP-Pro5cells in which irofulven has been shown previously to be apotent apoptosis inducer (16, 20). To identify early drugeffects, we used a 4-hour exposure to irofulven (10 Amol/L)and a postincubation in drug-free medium, depending on aspecific end point, for up to 12 to 16 hours elapsed time.These treatment conditions correspond to substantial but
still incomplete growth inhibition (<90% inhibition; datanot shown). To determine drug effects on cell growth andviability, longer times and a lower range of drug concen-trations were used.
Translocation of Proapoptotic Bax toMitochondriaThe recruitment of the proapoptotic Bax protein to
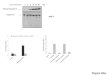
mitochondria is one of the early events that are essentialfor apoptosis triggering in various systems (29, 39). Theeffects of irofulven on the translocation of Bax from cytosolto mitochondria were examined by Western blotting usingcytosolic and mitochondrial fractions from drug-treatedcells (Fig. 2A). The quantification shows Bax signals in bothfractions normalized to respective h-actin signals. Prior todrug treatment, Bax was detected predominately in thecytoplasmic fraction. A 2 to 4 hours of incubation withirofulven (10 Amol/L) resulted in f2-fold increase inmitochondria-associated Bax. Bax translocation progressedrapidly over time. At 8 hours elapsed time, there was anf12-fold elevation of Bax signal in the mitochondrialfraction. Concurrently, the level of cytosolic Bax decreasedmarkedly below the initial level (Fig. 2A). Whereas in somesystems Bax translocation is promoted by the proteolyticcleavage of Bax molecule (40), Bax cleavage was notdetected in irofulven-treated LNCaP-Pro5 cells.
Figure 2. Translocation of proapoptotic Bax from cytosol to mitochon-dria. LNCaP-Pro5 cells were incubated with irofulven (10 Amol/L) for up to4 hours and postincubated in drug-free medium as indicated followed byWestern blot determination of total Bax protein in cytosolic andmitochondrial fractions. A, time course of Bax translocation. A represen-tative Western blot along with the quantification of Bax levels inmitochondrial (n, top ) and cytosolic (., bottom ) fractions. B, effects ofthe pan-caspase inhibitor Z-VAD-fmk (25 Amol/L added 1 hour beforeirofulven) on irofulven-induced changes in cytosolic Bax at 12 hourselapsed time. Data in both A and B are normalized to untreated control asratio of Bax signals to the respective actin signals. Points, mean [treatedversus control (T/C )] of at least two independent experiments; bars, SE(A). Columns, mean (treated versus control) of five to six independentexperiments; bars, SE (B). *, P < 0.02, significant reduction of irofulveneffect by Z-VAD-fmk (two-tailed t test).
Pathways of Cell Death by Irofulven1388
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

To assess whether irofulven-induced Bax translocation isupstream from the caspase cascade, the levels of cytosolicBax in irofulven-treated LNCaP-Pro5 cells were monitoredin presence of the broad-spectrum caspase inhibitorZ-VAD-fmk and the initiator caspase-9 inhibitor Z-LEHD-fmk. The initial depletion of cytosolic Bax (up to 6 hourselapsed time) was not prevented by either inhibitor(data not shown). At 12 hours, however, Z-VAD-fmksignificantly (P < 0.02) reduced the magnitude of iro-fulven-induced loss of Bax from the cytosolic fraction(Fig. 2B).Dissipation of DCm
The translocation of proapoptotic Bax translocation tomitochondria is expected to compromise mitochondrialmembrane integrity, which may include the dissipation ofDCm (41). Accordingly, flow cytometry determinationsshowed that irofulven affects DCm (Fig. 3). Whereas themajority of untreated LNCaP-Pro5 cells featured a highDCm, the number of cells with low DCm after a 6-hourexposure to irofulven (10 Amol/L) slightly increased from11.4 F 1.4% to 20.2 F 2.7% (Fig. 3A). The effect was furtherintensified at 12 hours, with 47 F 4.9% cells showing lowDCm (Fig. 3B).
The addition of Z-VAD-fmk prior to irofulven treat-ment had marginal effect on the fraction of cells with low
DCm at 6 hours elapsed time (Fig. 3A). At 12 hours,however, Z-VAD-fmk partially prevented the dissipation ofDCm as illustrated by the decrease in the number of cellswith low DCm from 47 F 4.9% for irofulven alone to 30 F0.5% in presence of Z-VAD-fmk (Fig. 3B). These findingssuggest that the activated caspases facilitate the collapseof DCm.
Cytochrome c ReleaseMitochondrial dysfunction promoted by Bax transloca-
tion usually leads to the leakage of cytochrome c frommitochondria (41). Accordingly, irofulven treatment ofLNCaP-Pro5 cells increased the levels of cytoplasmiccytochrome c. The effect was detectable after 4 hours andprogressed to reach a plateau at 6 to 12 hours (Fig. 4).Similar timing and progression of cytochrome c release wasalso detected in a related subline (LNCaP-LN3). Themagnitude of this effect, however, was less pronouncedthan in LNCaP-Pro5 (data not shown), consistent with thelower apoptotic propensity of LNCaP-LN3 in response toirofulven (16).
The preincubation of cells with either the pan-caspaseinhibitor or the caspase-9 inhibitor revealed that caspasedependency of cytochrome c release changes distinctlyover time (Fig. 4). Both inhibitors failed to preventirofulven-induced release of cytochrome c at 4 hours. At6 hours elapsed time, however, cytochrome c release be-came partly caspase dependent as indicated by a significantinhibitory effect of both Z-VAD-fmk and Z-LEHD-fmk(78% and 60% inhibition, respectively). Partial inhibition byZ-VAD-fmk was also observed at 12 hours (40% inhibition;data not shown). Thus, irofulven-induced early release of
Figure 3. Irofulven-induced dissipation of DCm. LNCaP-Pro5 cells weretreated continuously with irofulven (10 Amol/L) for 6 (A) and 12 (B) hoursin the presence and absence of the pan-caspase inhibitor Z-VAD-fmk(100 Amol/L) before flow cytometry measurements with JC-1 fluorescentprobe. Green and red fluorescence signals were gated as measures of cellswith high (open columns ) and low (closed columns ) DCm, respectively.
Figure 4. Irofulven-induced cytochrome c release into cytoplasm.LNCaP-Pro5 cells were exposed to irofulven (10 Amol/L) for 4 hoursfollowed by 2 and 8 hours postincubation in drug-free medium (6 and 12hours elapsed times). Cytochrome c was measured in cytosolic fractionfrom drug-treated cells by Western blotting. The pan-caspase inhibitorZ-VAD-fmk or the caspase-9 inhibitor Z-LEHD were added as indicated.Results were normalized to actin signals in the same samples andexpressed as the ratio of normalized signals in drug-treated samples overnormalized signals in control samples (treated versus control). Columns,mean of at least two independent experiments; bars, SE.
Molecular Cancer Therapeutics 1389
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

cytochrome c was not affected by the caspase inhibitorsand preceded loss of DCm. The temporal switch fromcaspase-independent to partially caspase-dependent re-lease of cytochrome c parallels analogous switch for thedissipation of DCm.Induction of Caspase CascadeCytochrome c released to cytosol forms a multimeric
complex with apoptotic protease-activating factor 1 andpro-caspase-9 leading to the activation of caspase-9 anddownstream caspases (42). We thus examined the effects ofirofulven on the processing of caspase-9 and of caspase-7and caspase-3, which are directly activated by caspase-9(42, 43). The temporal sequence of caspase processing/activation was assessed based on the conversion (cleavage)of the inactive pro-caspases to the enzymatically activetruncated forms (Fig. 5A and B).
The activation of caspase-9 became apparent after6 hours elapsed time (coinciding with the timing ofprofound cytochrome c release) and progressed at leastup to 16 hours (Fig. 5A and B). Processing of caspase-7shows a rapid progression and is essentially complete by 12hours. The most delayed and least effective is theprocessing of the effector caspase-3, which is in line withthe previously shown dispensability of that caspase forirofulven effects (18). As expected, caspase-9 inhibitorZ-LEHD-fmk and pan-caspase inhibitor Z-VAD-fmk abro-gated downstream processing of caspase-7 and caspase-3(Fig. 5C and D).
To further address the possible role of the executionerDEVD caspases 7/3, we examined the induction of DEVD-targeted proteolytic activity in the extracts from irofulven-treated cells using Ac-DEVD-pNA as the substrate. DEVDproteolytic activity increased from 7.6 F 2.6 pmol pNA/Agprotein/h in cell extracts from control cells to 54.4 F 1.9pmol pNA/Ag protein/h in cells treated with irofulven(10 Amol/L) for 4 hours. Consistent with the processing ofcaspase-3 and caspase-7, addition of a caspase-3/7 inhibitor(Ac-DEVD-CHO) at 10 Amol/L reduced DEVD proteolyticactivity almost to the control level (12.1 F 1.5 pmol pNA/Ag protein/h).Caspase-Dependent DNAFragmentationActivation of executioner caspases (such as caspase-3 and
caspase-7) normally precedes the manifestation of apopto-sis as massive DNA fragmentation (44). Consistent with theprevious findings (16), DNA fragmentation induced byirofulven (5 Amol/L) progressed with time reaching thelevel of 43.6 F 1.4% of total DNA after 12 hours (Fig. 6A)and 77.4 F 5.2% after 24 hours. This irofulven-inducedDNA fragmentation was profoundly suppressed (by 80.9 F4%) in presence of the pan-caspase inhibitor Z-VAD-fmk(Fig. 6B). Nearly as potent were the inhibitors of caspase-9and caspase-8/6 (Z-LEHD-fmk and Z-IETD-fmk), produc-ing 70.7 F 1.3% and 77.2 F 6.3% suppression, respectively.A more modest inhibition was produced by caspase-2inhibitor Z-VDVAD-fmk (42.3 F 6.0% inhibition) andby caspase-3 and caspase-7 inhibitor Ac-DEVD-fmk (32.9F 4.1% inhibition). Caspase-1 and caspase-4 inhibitor
Ac-YVAD-cmk had marginal effect (11.2 F 2.4% inhibi-
tion). These data further corroborate the initiator role ofcaspase-9 and show that irofulven-induced apoptotic DNAfragmentation results mostly from the caspase-mediatedpathway.
Inhibitionof theCaspaseCascadeDoesNotSuppresstheAntiproliferative Effects of Irofulven
If the fate of irofulven-treated cells depended solely onthe caspase-mediated route, the inhibition of the caspasecascade would be expected to rescue cells from death. Toresolve this question, we compared the loss of cell viability(by trypan blue exclusion) and cell growth inhibitioninduced by irofulven (5 Amol/L) in the presence andabsence of caspase inhibitors (Fig. 6C and D, respectively).
Consistent with the data reported previously (20), cellstreated with irofulven alone showed reduced viability at8 hours from the beginning of drug insult, with strong
Figure 5. Irofulven-induced caspase processing. Time course ofcaspase processing (A and B) and effects of inhibitors (C and D).LNCaP-Pro5 cells were treated continuously with irofulven (10 Amol/L). A,representative Western blots for the time course. B, relative fractions ofpro-caspases (top ) and activated caspases (bottom ) based on quantifica-tion of Western blots. Caspase-9 (E), caspase-7 (.), and caspase-3 (n).C, representative blot of caspase-3 and caspase-7 processing in thepresence of Z-LEHD-fmk (100 Amol/L) and Z-VAD-fmk (100 Amol/L) after4 hours of drug treatment and 12 hours postincubation. Lane 1, control;lane 2, irofulven (10 Amol/L); lane 3, irofulven (10 Amol/L) with Z-LEHD-fmk (100 Amol/L); lane 4, irofulven (10 Amol/L) with Z-VAD-fmk(100 Amol/L). D, quantitation of caspase processing in presence ofZ-LEHD-fmk and Z-VAD-fmk. Columns, mean of at least two independentexperiments; bars, SE.
Pathways of Cell Death by Irofulven1390
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

progression through 12 and 24 hours (Fig. 6C). Z-VAD-fmkand other caspase inhibitors had marginal to slight effectson the irofulven-induced loss of cell viability. The effects ofcaspase inhibitors remained marginal also when related tothe number of remaining cells (Fig. 6D). After a 24-hourexposure to irofulven (5 Amol/L) alone, the total cellnumber was reduced by 32 F 5% compared with untreatedcontrol decreases observed of either Z-VAD-fmk orZ-LEHD-fmk (42% and 50%, respectively). Consequently,the estimated numbers of viable cells are only marginallyhigher in the presence of caspase inhibitors. Furthermore,Z-VAD-fmk had negligible effects on cell growth inhibitionmeasured by the MTT assay (Fig. 6E; data not shown).After 12, 24, and 48 hours, irofulven concentrationsinhibiting the MTT signal by 50% (IC50) in the absenceversus presence of Z-VAD-fmk amounted to 1.2 versus 1.9,0.19 versus 0.23, and 0.10 versus 0.11 Amol/L, respectively.The inability of Z-VAD-fmk to rescue cells indicates asignificant contribution of a caspase-independent compo-nent in irofulven effects on cell growth and viability.Monobromobimane Antagonizes Cell Growth Inhibi-
tion and Suppresses Apoptosis by IrofulvenIrofulven-induced, caspase-independent cell death might
reflect the known alternative route of apoptosis, whichoriginates from MPT mediated by oxidative changes incritical redox-sensing mitochondrial membrane proteins(32, 34, 45). To test this possibility, we used monobromo-bimane, a protein reagent that prevents conformationalchanges in mitochondrial membrane proteins that arerequired to precipitate MPT (46). Thus, by blocking theearly step in caspase-independent, AIF-mediated apopto-sis, monobromobimane can serve as a mechanistic markerfor this alternative route.
The ability of monobromobimane to interfere withirofulven-induced apoptosis was assessed using Annexin
V binding assay. High Annexin V signal in this assaycorresponds to phosphatidylserine externalization in cellmembranes, which is a characteristic marker of bothcaspase-dependent and caspase-independent routes ofapoptosis (47). The flow cytometry histograms forLNCaP-Pro5 cells incubated with irofulven (2.5 Amol/L)show large numbers of cells with high Annexin V signaland low PI signal (that are indicative of early apoptosis) aswell as cells with high signal for both Annexin and PI(indicative of late apoptosis; Fig. 7A). Addition of mono-bromobimane 1 hour prior to irofulven attenuated theintensity of the Annexin signals. The quantification of flowcytometry data (Fig. 7B) showed that apoptotic cells aremarkedly reduced in the presence of monobromobimane.
Figure 6. Effects of caspase inhibitors on irofulven-induced apoptoticDNA fragmentation and cell growth. The following caspase inhibitors,when used as indicated below, were added at 25 Amol/L concentration1 hour prior to the addition of irofulven: Z-VAD-fmk (the pan-caspaseinhibitor), Z-LEHD-fmk (caspase-9), Z-IETD-fmk (caspase-8 and caspase-6),Z-VDVAD-fmk (caspase-2), Ac-DEVD-fmk (caspase-3 and caspase-7), andAc-YVAD-cmk (caspase-1 and caspase-4). LNCaP-Pro5 cells wereexposed to irofulven (5 Amol/L) for the indicated times in the presenceor absence of caspase inhibitors followed by the determinations ofrespective apoptotic end points. A and C, untreated cells (!); irofulvenalone (.); and irofulven in the presence of Z-VAD-fmk (E), Z-IETD-fmk(5), or Z-LEHD-fmk (o). A, time course of apoptotic DNA fragmentationinduced by irofulven (5 Amol/L) in the absence and presence of caspaseinhibitors. Values of percentage of fragmented DNA were corrected forrespective values in cultures without irofulven. B, inhibition of DNAfragmentation induced after 12-hour incubation with irofulven (5 Amol/L)by the indicated caspase inhibitors. C, time course of changes in cellviability. D, remaining cells after 24-hour exposure to irofulven (10 Amol/L)normalized to the number of cells in untreated cultures. TOTAL cells basedon electronic cell counting represent both viable and nonviable cells.Values for viable cells are estimated by multiplying total cell number byrespective relative viability values from C. Columns, mean of twoexperiments in duplicates; bars, SE. E, cytotoxicity of irofulven (HMAF )in the absence and presence of Z-VAD-fmk (25 Amol/L) measured usingthe MTT assay after 48-hour continuous exposure to the drug.
Molecular Cancer Therapeutics 1391
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

Because monobromobimane should mainly antago-nize the MPT-mediated, caspase-independent route, mono-bromobimane was expected to be less antagonistic withregard to DNA fragmentation, which is largely mediatedby the caspase cascade. Determinations of DNA fragmen-tation in the presence of monobromobimane corroboratedthis expectation. Compared with irofulven alone, mono-bromobimane exerted some protective effect, but itsmagnitude (f14% at 1 Amol/L monobromobimane; datanot shown) seemed insufficient to account for the profoundprotection against apoptosis monitored by the Annexin Vassay. Thus, the antagonistic effects of monobromobimaneon irofulven-induced apoptosis are consistent with theinhibition of MPT/AIF pathway.
To determine whether monobromobimane antagonizesgrowth inhibition of irofulven-induced cells, cells werepretreated with monobromobimane for 1 hour prior to thedrug addition for 48 hours. Following the recommendationfor the median effect approach (35), these experiments werecarried out using a constant ratio of irofulven to mono-bromobimane as opposed to fixed monobromobimaneconcentration in the Annexin assay. Cell growth wasmeasured by the MTT assay and data were analyzed byplotting the combination index versus the fraction affected(Fig. 7C). In this plot, low to modest fraction affected valuescorrespond to noncytotoxic to slightly cytotoxic concen-trations of monobromobimane (data not shown). Theinterpretation of monobromobimane antagonism needs tobe limited to this range, because combination index valuesat higher fraction affected values are likely to be influencedby nonspecific cytotoxicity of monobromobimane. Thecombination index values observed at fraction affectedvalues up to f0.35 are clearly in the range indicative ofstrong antagonistic or antagonistic interaction betweenirofulven and monobromobimane as per criteria of Chouand Talalay (35). This result further strengthens thepossibility that the antiproliferative effects of irofulvenoriginate in part from the MPT/AIF–mediated alternativeapoptotic route.
DiscussionThe consistently potent induction of apoptosis in varioustumor systems is the hallmark of the novel anticancer drug,irofulven (15, 16, 18, 19). Previous investigations haveidentified DNA and proteins as major targets for alkylationby this drug in cancer cells (5). Various studies implicatedboth reactivities in apoptotic properties of irofulven (2, 5,25). This report defines key molecular events that integrateirofulven-induced apoptotic signals and lead to cell death.
Mitochondrial dysfunction plays a central role inirofulven-induced apoptosis in prostate cancer LNCaP-Pro5 cells. An early step in this process is the translocationof proapoptotic Bax to mitochondria (Fig. 2) followed bythe dissipation of DCm (Fig. 3) and the leakage ofcytochrome c (Fig. 4). The released cytochrome c ap-parently promotes the activation of caspases (Fig. 5) andgives rise to caspase-dependent DNA fragmentation
Figure 7. Effect of monobromobimane on irofulven-induced apoptosisand cytotoxicity. A, representative bivariate flow cytometry histograms ofAnnexin V and PI binding to LNCaP-Pro5 cells after a 24-hour exposure toirofulven (2.5 Amol/L; IROF ) in the absence and presence of monobromo-bimane (0.5 Amol/L; MBB ). B, quantification of early and late apoptosisbased on flow cytometry analysis. Columns, mean corrected for respec-tive values for untreated cells from two experiments in duplicates; bars,SE. C, plot of combination index (CI ) versus fraction affected (Fa) forLNCaP-Pro5 cells incubated for 48 hours with either agent alone or incombination followed by MTT assay. Combination index and fractionaffected values were computed from the MTT data according to themedian effect method of Chou and Talalay (35). Regression points andcalculated confidence limits represent data for mutually exclusive model(35). Ranges of combination index values corresponding to antagonisticand supra-additive interactions between the studied compounds areindicated using the criteria of Chou and Talalay (35).
Pathways of Cell Death by Irofulven1392
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

(Fig. 6). Importantly, the apoptotic fate of irofulven-treatedcells is determined not only by the caspase-mediatedpathway but also via a coexisting caspase-independentroute (Figs. 6 and 7). The nature and attributes of theirofulven-induced apoptotic cascade resulting in these tworoutes are discussed below in the context of the likelyinterplay between the drug-induced DNA damage signal-ing and the consequences of protein damage.
The observation that irofulven induces Bax translocationis consistent with the known effects of DNA damagesignaling, although other apoptotic stimuli may alsoproduce such effect (27, 29, 39). Especially relevant to theprotein-targeting activities of irofulven are the reports thatthe oxidation of intracellular thiols facilitate the proapop-totic rearrangements of Bax/Bcl-2 balance (48, 49). Thus,irofulven-induced pro-oxidative changes in the proteinredox status might additionally enhance Bax translocationresulting from DNA damage signaling.
Irofulven-induced Bax translocation seems to precedeboth cytochrome c release and loss of the inner membranepotential DCm, which might reflect a causative role of Baxtranslocation. However, the initial DCm dissipation is stillrelatively small at the time of significant cytochrome crelease. This pattern suggests that, whereas the initialcytochrome c release depends largely on Bax translocation,DCm collapse might be in part Bax/Bcl-2 independent.Such interpretation is supported by our previous findings,which showed that forced overexpression of Bcl-2 had mar-ginal effect on early irofulven-induced DCm collapse (19).
Bax/Bcl-2-independent DCm collapse resulting in therelease of AIF is a well-established pathway in the action ofagents that oxidize the critical thiol groups in mitochon-drial membrane proteins (28, 50). In addition to globallydistorting protein redox status (25, 51), irofulven directlyreacts with a subset of mitochondrial proteins.4 Furtherstudies are needed to clarify whether these effects indeedplay a role in drug-induced inner membrane permeabilitytransition. Consistent with such a role, however, is theobservation that irofulven causes the release of AIF (inleukemic CEM cells).4
There is no doubt that irofulven-induced mitochondrialdysfunction is upstream from the caspase cascade, as Baxtranslocation, changes in DCm, and cytochrome c releasewere consistently independent of caspase inhibitors at earlytimes. However, the emergence of partial caspase depen-dency at later times strongly suggests that the activatedcaspases provide a positive feedback loop, which furthercompromises mitochondrial integrity. Such interpretation isconsistent with other reports that implicated caspase ac-tivation as a factor potentiating the collapse of DCm (52, 53).
The timing of caspase processing in response to irofulvenand the effects of caspase inhibitors point to the activationof caspase-9 upstream in the cascade followed by theactivation of caspase-7 and caspase-3. This order isconsistent with the established role of caspase-9 as an
initiator caspase (42, 43). To activate caspase-9, cytochromec is thought to interact first with an adaptor moleculeapoptotic protease-activating factor 1 and next with pro-caspase-9 (44). Such interaction can be efficient in LNCaP-Pro5 cells, which have clearly detectable levels of apoptoticprotease-activating factor 1 that are unaffected by exposureto irofulven.4 Noteworthy, a recent study reported that theactivation of caspase-9 is inhibited by the antiapoptoticredox-regulating protein thioredoxin (54). The ability ofirofulven to interfere with the function of the thioredoxinsystem (25) may thus facilitate caspase-9 activation. Thispossibility indicates yet another level at which a crosstalkbetween irofulven effects on multiple cellular targets mightself-potentiate apoptotic signaling.
Massive fragmentation of DNA in irofulven-treatedLNCaP-Pro5 cells is clearly caspase dependent asevidenced by its near complete abrogation by the pan-caspase inhibitor. General and specific caspase inhibitorsabrogated irofulven-induced apoptotic DNA fragmenta-tion with the following order of potency: pan-caspasez caspase-9 > caspase-8/6 > caspase-2 > caspase-3/7 >caspase-1/4. The potent inhibitory effect of the caspase-9inhibitor corroborates the role of this caspase as the maininitiator of the caspase cascade in cells exposed toirofulven. It is less clear which caspases serve as execu-tioners. Any significant role of caspase-1/4 can be ruledout, because the inhibitor of these caspases had marginaleffect on irofulven-induced apoptotic DNA fragmentation.Although caspase-3 is commonly regarded as majorexecutioner caspase, it was less efficiently processed inirofulven-treated LNCaP-Pro5 cells than other caspases.The modest inhibition of irofulven-induced DNA fragmen-tation in LNCaP-Pro5 cells observed in this study for theinhibitor of DEVD caspase-3 and caspase-7 is consistentwith virtually no effect of this inhibitor in leukemic CEMcells (3). Moreover, previous studies with caspase-3-deficient MCF-7 cells showed that caspase-3 is dispensablefor irofulven effects (18). The activation of caspase-7accompanied apoptosis by irofulven in LNCaP-Pro5 cells(this study) and in pancreatic cancer cells (17) but wasnegligible in two breast cancer cell lines (18). Thus,although caspase-7 may contribute to the cascade in somemodels, it is unlikely to be universally critical for irofulven-induced apoptosis.
In contrast to weak effects of the caspase-3/7 inhibitor, apotent suppression of DNA fragmentation was caused byZ-IETD-fmk, the inhibitor of caspase-8/6. Irofulven wasreported to induce caspase-8 processing in pancreaticcancer cells (17). In LNCaP-Pro5 cells, however, we couldnot detect any caspase-8 processing (by Western blotting)or any caspase-8 enzymatic activity in cell extracts fromdrug-treated cells (data not shown). These results suggestthat the inhibition of DNA fragmentation by Z-IETD-fmkreflects the inactivation of caspase-6 rather than caspase-8.The possibility that caspase-6 could be an important exe-cutioner caspase in irofulven-induced apoptotic DNAfragmentation is consistent with analogous inference basedon results in breast cancer cell lines (18).4 Liang and Woynarowska, unpublished data.
Molecular Cancer Therapeutics 1393
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

The findings of this report and data in other models (15,16, 18, 19) show clearly that the death process initiatedby irofulven-induced mitochondrial dysfunction has dis-tinct attributes of apoptosis: early changes in cellmembrane phospholipids, altered nuclear morphology,and DNA fragmentation, all of which precede the lossof cell membrane integrity (cell ‘‘viability’’). It is importantto underscore, however, that blocking the caspasecascade, which largely abrogates DNA fragmentation,minimally protects against the loss of membrane integ-rity and does not rescue irofulven-treated cells fromgrowth inhibition. These observations provide unequivocalevidence that irofulven-induced apoptosis comprisesnot only caspase-dependent pathway but also caspase-independent death.
Although the classic route of apoptosis involves caspase-mediated phenomena, in a broader sense, apoptosis refersto systemic cell degradation ‘‘from inside’’ that precedesthe loss of cell membrane integrity. The specific term‘‘caspase-independent apoptosis’’ is increasingly used inthe literature to denote a defined alternative apoptoticroute (31, 32). This route involves membrane permeabilitytransition that is initiated by changes in redox-sensingmitochondrial membrane proteins and proceeds in a Bcl-2-independent manner. MPT leads to the release of apopto-genic factors, notably AIF. AIF translocates to the nucleus,where it affects a long-range disruption of nuclear chro-matin but not oligonucleosomal level DNA fragmentation.Like caspase-mediated apoptosis, AIF-mediated, caspase-independent apoptosis may also be induced by variousstimuli. Nonetheless, the activation of caspase-independentapoptotic route, in addition to the caspase-mediated effects,is a characteristic feature of protein-reactive agents, suchas diamide (31, 32, 55), but is not observed for DNA-damaging agents, such as etoposide, that do not react withproteins (34).
The pattern of irofulven effects is consistent with thecontribution of the above-mentioned mechanistic attributesof caspase-independent apoptosis. First, preventing thereactivity of irofulven with the redox-sensing sulfhydrylsin the mitochondrial membrane proteins (by preincubationwith monobromobimane) partially antagonizes the apo-ptotic and antiproliferative effects of irofulven (Fig. 7).Secondly, as mentioned above, irofulven-induced mem-brane permeability transition seems to be partly Bcl-2independent. Finally, irofulven-induced apoptosis islargely Bcl-2 independent, unlike apoptosis by etoposidethat is almost completely abrogated by Bcl-2 overexpres-sion (19). The idea that caspase-independent cell deathinduced by irofulven indeed reflects the AIF-mediatedalternative apoptotic pathway (as suggested in the dia-gram in Fig. 8) is highly likely, but it still needs to beconfirmed unequivocally. Further studies are needed, inparticular, to explore drug effects on redox-sensitivemitochondrial proteins that might initiate AIF-mediatedcell death.
Taken together, the reported findings reveal that the fateof irofulven-treated cells is most likely determined by a
complex interplay among multiple signaling routes origi-nated from drug effects on its diverse cellular targets (DNAand proteins). These signals converge and integrate at themitochondrial level as the decisive point and diverge againdownstream from mitochondria into two coexisting deathroutes (Fig. 8). The coinduction of multiple signalingpathways and apoptotic routes could play essential rolein the ability of irofulven to preempt prosurvival attemptsand facilitate the efficient elimination of cancer cells,including apoptosis-resistant models (2, 16, 18, 19). Theirofulven paradigm of consistently potent apoptosis incancer cells underscores the potential benefits of polytar-geted anticancer approaches.
Acknowledgments
We thank Emily Van Laar for critical reading of the article, insightfulcomments, and valuable suggestions.
References
1. Norbury CJ, Zhivotovsky B. DNA damage-induced apoptosis. Oncogene2004;23:2797–808.
2. Woynarowska BA, Woynarowski JM. Preferential targeting of apo-ptosis in tumor versus normal cells. Biochim Biophys Acta 2002;1587:309–17.
Figure 8. Possible integration of irofulven-induced death signalingbased on this report and other investigations. Apoptotic signals resultingfrom DNA damage and protein reactivity of irofulven involve translocationof Bax to mitochondria, cytochrome c release, activation of the caspasecascade, and apoptotic DNA fragmentation. However, only minor portionof irofulven-induced DNA fragmentation is inhibitable by Bcl-2 over-expression (19), indicating contribution of other factors. Protein reactivityof irofulven results in pro-oxidative distortion of protein redox status(25, 51). These changes may contribute to membrane permeabilitytransition (the dissipation of DCm; MPT ), which seems to be independentof Bax/Bcl-2 at early times (19). Irofulven effects culminate not only incaspase-dependent apoptosis but also in caspase-independent cell death.The mechanism of caspase-independent death promoted by irofulvenremains unknown but is likely to involve the known alternative apoptoticpathway mediated by MPT and AIF release.
Pathways of Cell Death by Irofulven1394
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

3. Woynarowski JM, Koester S, Woynarowska B, et al. Is drug-inducedDNA damage sufficient for apoptosis induction? Proc AACR 1999;40:736.
4. Akaboshi M, Kawai K, Maki H, et al. The number of platinum atomsbinding to DNA, RNA and protein molecules of HeLa cells treatedwith cisplatin at its mean lethal concentration. Jpn J Cancer Res 1992;83:522–6.
5. Herzig MCS, Arnett B, MacDonald JR, Woynarowski JM. Drug uptakeand cellular targets of hydroxymethylacylfulvene (HMAF). BiochemPharmacol 1999;58:217–25.
6. Kelner MJ, McMorris TC, Montoya MA, et al. Characterization ofMGI 114 (HMAF) histiospecific toxicity in human tumor cell lines. CancerChemother Pharmacol 1999;44:235–40.
7. Urien S, Alexandre J, Raymond E, et al. Phase I population pharmaco-kinetics of irofulven. Anticancer Drugs 2003;14:353–8.
8. Bayes M, Rabasseda X, Prous JR. Gateways to clinical trials. March2003. Methods Find Exp Clin Pharmacol 2003;25:145–68.
9. MacDonald JR, Muscoplat CC, Dexter DL, et al. Preclinical antitumoractivity of 6-hydroxymethylacylfulvene, a semisynthetic derivative of themushroom toxin illudin S. Cancer Res 1997;57:279–83.
10. Kelner MJ, McMorris TC, Estes L, et al. Anti-leukemic action of thenovel agent MGI 114 (HMAF) and synergistic action with topotecan.Leukemia 2000;14:136–41.
11. Kelner MJ, McMorris TC, Estes LA, et al. Efficacy of MGI 114 (HMAF)against the MRP+ metastatic MV522 lung carcinoma xenograft. Anti-cancer Drugs 2000;11:217–24.
12. Van Laar ES, Roth S, Weitman S, MacDonald JR, Waters SJ. Activityof irofulven against human pancreatic carcinoma cell lines in vitro andin vivo . Anticancer Res 2004;24:59–65.
13. Van Laar ES, Weitman S, MacDonald JR, Waters SJ. Antitumoractivity of irofulven monotherapy and in combination with mitoxantroneor docetaxel against human prostate cancer models. Prostate 2004;59:22–32.
14. Kelner MJ, McMorris TC, Estes L, et al. Efficacy of MGI 114(6-hydroxymethylacylfulvene, HMAF) against the mdr1/gp170 meta-static MV522 lung carcinoma xenograft. Eur J Cancer 1998;34:908–13.
15. Woynarowski JM, Napier C, Koester SK, et al. Effects on DNAintegrity and apoptosis induction by a novel antitumor sesquiterpenedrug, 6-hydroxymethylacylfulvene (HMAF, MGI 114). Biochem Pharmacol1997;54:1181–93.
16. Woynarowska BA, Woynarowski JM, Herzig MCS, et al. Differentialcytotoxicity and induction of apoptosis in tumor and normal cells byhydroxymethylacylfulvene (HMAF). Biochem Pharmacol 2000;59:1217–26.
17. Wang W, Waters SJ, MacDonald JR, et al. Irofulven (6-hydroxyme-thylacylfulvene, MGI 114) induces caspase 8 and 9-mediated apoptosisin human pancreatic adenocarcinoma cells. Anticancer Res 2001;21:1789–94.
18. Herzig MCS, Liang H, Johnson AE, Woynarowska B, WoynarowskiJM. Irofulven induces apoptosis in breast cancer cells regardless ofcaspase-3 status. Breast Cancer Res Treat 2002;71:133–43.
19. Herzig MC, Trevino AV, Liang H, et al. Apoptosis induction by thedual-action DNA- and protein-reactive antitumor drug irofulven is largelyBcl-2-independent. Biochem Pharmacol 2003;65:503–13.
20. Woynarowska BA, Roberts K, Woynarowski JM, MacDonald JR,Herman TS. Targeting apoptosis by hydroxymethylacylfulvene in combi-nation with g radiation in prostate tumor cells. Radiat Res 2000;154:429–38.
21. McMorris TC, Elayadi AN, Yu J, Hu Y, Kelner MJ. Metabolism ofantitumor hydroxymethylacylfulvene by rat liver cytosol. Drug MetabDispos 1999;27:983–5.
22. McMorris TC, Yu J, Ngo HT, Wang H, Kelner MJ. Preparationand biological activity of amino acid and peptide conjugates of anti-tumor hydroxymethylacylfulvene. J Med Chem 2000;43:3577–80.
23. Dick RA, Yu X, Kensler TW. NADPH alkenal/one oxidoreductaseactivity determines sensitivity of cancer cells to the chemotherapeuticalkylating agent irofulven. Clin Cancer Res 2004;10:1492–9.
24. Lawley PD, Phillips DH. DNA adducts from chemotherapeutic agents.Mutat Res 1996;355:13–40.
25. Woynarowska BA, Woynarowski JM, Liang H, et al. Irofulven binding
and inactivation of purified and cellular redox-controlling proteins. ProcAACR 2004;45:A1510.
26. Henry-Mowatt J, Dive C, Martinou JC, James D. Role of mitochon-drial membrane permeabilization in apoptosis and cancer. Oncogene 2004;23:2850–60.
27. van Gurp M, Festjens N, van Loo G, Saelens X, Vandenabeele P.Mitochondrial intermembrane proteins in cell death. Biochem Biophys ResCommun 2003;304:487–97.
28. Cande C, Vahsen N, Garrido C, Kroemer G. Apoptosis-inducingfactor (AIF): caspase-independent after all. Cell Death Differ 2004;11:591–5.
29. Tsujimoto Y. Cell death regulation by the Bcl-2 protein family in themitochondria. J Cell Physiol 2003;195:158–67.
30. Kuwana T, Newmeyer DD. Bcl-2-family proteins and the role of mito-chondria in apoptosis. Curr Opin Cell Biol 2003;15:691–9.
31. Cregan SP, Dawson VL, Slack RS. Role of AIF in caspase-dependent and caspase-independent cell death. Oncogene 2004;23:2785–96.
32. Costantini P, Belzacq AS, Vieira HL, et al. Oxidation of a critical thiolresidue of the adenine nucleotide translocator enforces Bcl-2-independentpermeability transition pore opening and apoptosis. Oncogene 2000;19:307–14.
33. McStay GP, Clarke SJ, Halestrap AP. Role of critical thiol groups onthe matrix surface of the adenine nucleotide translocase in the mechanismof the mitochondrial permeability transition pore. Biochem J 2002;367:541–8.
34. Joseph B, Marchetti P, Formstecher P, et al. Mitochondrial dysfunc-tion is an essential step for killing of non-small cell lung carcinomasresistant to conventional treatment. Oncogene 2002;21:65–77.
35. Chou TC, Talalay P. Quantitative analysis of dose-effect relationships:the combined effects of multiple drugs or enzyme inhibitors. Adv EnzymeRegul 1984;22:27–55.
36. Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The releaseof cytochrome c from mitochondria: a primary site for Bcl-2 regulation ofapoptosis [see comments]. Science 1997;275:1132–6.
37. Salvioli S, Ardizzoni A, Franceschi C, Cossarizza A. JC-1, but notDiOC6(3) or rhodamine 123, is a reliable fluorescent probe to assess y c
changes in intact cells: implications for studies on mitochondrial func-tionality during apoptosis. FEBS Lett 1997;411:77–82.
38. Woynarowska B, Woynarowski JM, Higdon AL, Munoz R. Inductionof apoptosis in prostate tumor cells by hydroxymethylacylfulvene (HMAF)and oxaliplatin. Proc AACR 1999;40:488.
39. Jia L, Patwari Y, Srinivasula SM, et al. Bax translocation is crucial forthe sensitivity of leukemic cells to etoposide-induced apoptosis. Oncogene2001;20:4817–26.
40. Gao G, Dou QP. N-terminal cleavage of bax by calpain generates apotent proapoptotic 18-kDa fragment that promotes bcl-2-independentcytochrome c release and apoptotic cell death. J Cell Biochem 2000;80:53–72.
41. Costantini P, Jacotot E, Decaudin D, Kroemer G. Mitochondrion as anovel target of anticancer chemotherapy. J Natl Cancer Inst 2000;92:1042–53.
42. Slee EA, Harte MT, Kluck RM, et al. Ordering the cytochromec -initiated caspase cascade: hierarchical activation of caspases-2, -3, -6,-7, -8, and -10 in a caspase-9-dependent manner. J Cell Biol 1999;144:281–92.
43. Kuida K, Haydar TF, Kuan CY, et al. Reduced apoptosis and cyto-chrome c -mediated caspase activation in mice lacking caspase 9. Cell1998;94:325–37.
44. Kim R, Tanabe K, Uchida Y, et al. Current status of the molecularmechanisms of anticancer drug-induced apoptosis. The contribution ofmolecular-level analysis to cancer chemotherapy. Cancer ChemotherPharmacol 2002;50:343–52.
45. Halestrap AP, Brennerb C. The adenine nucleotide translocase: acentral component of the mitochondrial permeability transition pore andkey player in cell death. Curr Med Chem 2003;10:1507–25.
46. Pallis M, Grundy M, Turzanski J, Kofler R, Russell N. Mitochondrialmembrane sensitivity to depolarization in acute myeloblastic leukemia isassociated with spontaneous in vitro apoptosis, wild-type TP53, andvicinal thiol/disulfide status. Blood 2001;98:405–13.
Molecular Cancer Therapeutics 1395
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

47. Vermes I, Haanen C, Reutelingsperger C. Flow cytometry of apoptoticcell death. J Immunol Methods 2000;243:167–90.
48. Honda T, Coppola S, Ghibelli L, et al. GSH depletion enhancesadenoviral bax-induced apoptosis in lung cancer cells. Cancer Gene Ther2004;11:249–55.
49. Maser RS, Antoku K, Scully WJ Jr, Cho RL, Johnson DE. Analysisof the role of conserved cysteine residues in the bcl-2 oncoprotein.Biochem Biophys Res Commun 2000;277:171–8.
50. Halestrap AP, McStay GP, Clarke SJ. The permeability transitionpore complex: another view. Biochimie 2002;84:153–66.
51. Liang H, Herzig MCS, Salinas R, et al. Pro-oxidative distortion ofthe cellular redox-homeostasis in irofulven-induced apoptosis. Clin CancerRes 2001;7:3723s.
52. Chen Q, Gong B, Almasan A. Distinct stages of cytochrome c releasefrom mitochondria: evidence for a feedback amplification loop linkingcaspase activation to mitochondrial dysfunction in genotoxic stress in-duced apoptosis. Cell Death Differ 2000;7:227–33.
53. Gao CF, Ren S, Zhang L, et al. Caspase-dependent cytosolic releaseof cytochrome c and membrane translocation of Bax in p53-inducedapoptosis. Exp Cell Res 2001;265:145–51.
54. Katoh I, Tomimori Y, Ikawa Y, Kurata S. Dimerization and processingof procaspase-9 by redox stress in mitochondria. J Biol Chem 2004;279:15515–23.
55. Zamzami N, Marzo I, Susin SA, et al. The thiol crosslinking agentdiamide overcomes the apoptosis-inhibitory effect of Bcl-2 by enforc-ing mitochondrial permeability transition. Oncogene 1998;16:1055–63.
Pathways of Cell Death by Irofulven1396
Mol Cancer Ther 2004;3(11). November 2004
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from

2004;3:1385-1396. Mol Cancer Ther Huiyun Liang, Richard A. Salinas, Belinda Z. Leal, et al. death induced by irofulven in prostate cancer cellsCaspase-mediated apoptosis and caspase-independent cell
Updated version
http://mct.aacrjournals.org/content/3/11/1385
Access the most recent version of this article at:
Cited articles
http://mct.aacrjournals.org/content/3/11/1385.full#ref-list-1
This article cites 54 articles, 8 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/3/11/1385.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mct.aacrjournals.org/content/3/11/1385To request permission to re-use all or part of this article, use this link
Research. on August 29, 2020. © 2004 American Association for Cancermct.aacrjournals.org Downloaded from