Embed Size (px)
Citation preview

Developmental Expression of the Actin DepolymerizingFactor ADF in the Mouse Inner Ear and Spiral GangliaMichel K. Herde,1 Eckhard Friauf,2 and Marco B. Rust1*1Neurobiology/Neurophysiology Group, Department of Biology, University of Kaiserslautern, 67663 Kaiserslautern, Germany2Animal Physiology Group, Department of Biology, University of Kaiserslautern, 67663 Kaiserslautern, Germany
ABSTRACTHair cells, the inner ear’s sensory cells, are characterizedby tens to hundreds of actin-rich stereocilia that form thehair bundle apparatus necessary for mechanoelectricaltransduction. Both the number and length of actin fila-ments are precisely regulated in stereocilia. Proper co-chlear and vestibular function also depends on actin fila-ments in nonsensory supporting cells. The formation ofactin filaments is a dynamic, treadmill-like process inwhich actin-binding proteins play crucial roles. However,little is known about the presence and function of actinbinding molecules in the inner ear, which set up, and main-tain, actin-rich structures and regulate actin turnover.Here we examined the expression and subcellular locationof the actin filament depolymerizing factor (ADF) in thecochlea and vestibular organs. By means of immunocyto-chemistry and confocal microscopy, we analyzed whole-
mount preparations and cross-sections in fetal and post-natal mice (E15–P26). We found a transient ADFexpression in immature hair cells of the organ of Corti, theutricle, and the saccule. Interestingly, the stereocilia werenot labeled. By P26, ADF expression was restricted to sup-porting cells. In addition, we localized ADF in presynapticterminals of medio-olivocochlear projections after hearingonset. A small population of spiral ganglion neuronsstrongly expressed ADF. Based on their relative number,peripheral location within the ganglion, smaller somasize, and coexpression of neurofilament 200, we identi-fied these cells as Type II spiral ganglion neurons. Thedevelopmentally regulated ADF expression suggests atemporally restricted function in the stereocilia and, thus,a hitherto undescribed role of ADF. J. Comp. Neurol. 518:1724–1741, 2010.
© 2009 Wiley-Liss, Inc.
INDEXING TERMS: cochlea; planar cell polarity; actin treadmilling; stereocilia; efferent innervations
Actin depolymerizing factor (ADF) is a member of theADF/cofilin family of actin-binding proteins. ADF/cofilinbind cooperatively to filamentous actin (F-actin) (Hawkinset al., 1993; Hayden et al., 1993), resulting in filamentsevering and dissociation of ADP-bound actin subunits atthe filament’s minus end (Carlier et al., 1997; McGough etal., 1997). ADF/cofilin thus speeds up actin dynamics asthey increase the number of actin filaments that can eitherelongate or shorten, depending on the local set of actinbinding proteins (Condeelis, 2001). Furthermore, ADF/cofilin accelerate actin treadmilling, a permanent processof ATP-actin incorporation at the plus end as well as hydro-lysis and ADP-actin dissociation at the minus end (Bam-burg and Wiggan, 2002).
ADF and its homolog n-cofilin display a broad tissuedistribution with substantial overlap (Gurniak et al., 2005;Bellenchi et al., 2007). Whereas inactivation of n-cofilinresults in severe phenotypes, such as defective neuraltube closure and a lissencephaly-like disorder (Gurniak et
al., 2005; Bellenchi et al., 2007), deletion of ADF induces arather mild phenotype of corneal hypertrophy, combinedwith stromal neovascularization (Bellenchi et al., 2007).These defects have also been described for a spontaneousmouse mutant (Ikeda et al., 2003).
Functional inactivation of twinstar, the Drosophila ho-molog of ADF/cofilin, produces defective planar cell polar-ity in several epithelia, including wing and eye (Blair et al.,2006). Planar cell polarization, a process of tissue organi-zation in which cells adopt a uniform orientation within thehorizontal plane of an epithelium, occurs in a great numberof epithelia throughout the metazoans and is regulated by
Additional Supporting Information may be found in the online versionof this article.
Grant sponsor: German National Academic Foundation (to M.K.H.).
*CORRESPONDENCE TO: Marco B. Rust, PhD, Neurobiology/Neurophysiology Group, Department of Biology, University of Kaiserslau-tern, Erwin-Schrodinger-Stra�e 13, D-67663 Kaiserslautern, Germany.E-mail: [email protected]
Received 28 May 2009; Revised 14 August 2009; Accepted 21 November2009DOI 10.1002/cne.22298Published online December 17, 2009 in Wiley InterScience (www.interscience.wiley.com).© 2009 Wiley-Liss, Inc.
RESEARCH ARTICLE
1724 The Journal of Comparative Neurology � Research in Systems Neuroscience 518:1724–1741 (2010)

a conserved signaling cascade that also includes the rear-rangement and polarization of cytoskeletal structures(Lewis and Davies, 2002; Kelly and Chen, 2007; Zallen,2007).
The organ of Corti, the mammalian auditory sensory ep-ithelium, features a remarkable form of planar cell polarityas various cell types are precisely arranged in lines alongthe length of the cochlear duct. Moreover, cochlear haircells possess several rows of highly organized stereociliathat form a “V”-shaped hair bundle in a staircase patternwith the vertices pointing away from the center of thecochlea (review: Kelly and Chen, 2007). The structuralbackbone of stereocilia is formed by actin filaments(Drenckhahn et al., 1991), and actin undergoes permanentremodeling in the stereocilia of cochlear hair cells (Schnei-der et al., 2002). Interestingly, the velocity of actin tread-milling is tightly coupled to the stereocilia length, whichimplies a concerted activity of various actin-regulating pro-teins, including actin depolymerizing activity (Manor andKachar, 2008). Thus, several findings, like defective planarcell polarity in twinstar mutants and the importance ofplanar cell polarity signaling cascades during inner ear de-velopment, as well as the persistent precise regulation ofactin turnover in stereocilia, imply an important role ofactin depolymerization for inner ear development andmaintenance.
ADF expression was recently shown in the avian audi-tory sensory epithelium (Oh et al., 2002). ADF and its in-teraction partner WDR1 (WD40 repeat protein 1), the ver-tebrate homologue of yeast actin-interacting protein 1AIP1, are upregulated in cochlear structures subsequentto acoustic overstimulation (Oh et al., 2002), suggestingthat ADF is important for cochlear function and regenera-tive processes taking place in the avian inner ear. How-ever, the expression of ADF has not been investigated asyet in the mammalian inner ear.
Choosing the mouse as a model system, we performedan in-depth characterization of the developmental andspatial expression pattern of ADF in the mammalian co-chlea and in the vestibular sensory epithelia. By exploitingan ADF-specific antibody, we show that ADF is transientlyexpressed in developing hair cells of the organ of Corti andvestibular sensory epithelia, namely, the utricular maculaand crista ampullaris. Whereas ADF expression was broadin mature vestibular organs, it became restricted to sup-porting cells in the mature cochlea. Moreover, after hear-ing onset, ADF was localized to presynaptic structures ofmedio-olivocochlear (MOC) projections innervating outerhair cells (OHCs). This synaptic location was time-paralleled by a strong ADF expression in the somata of asubset of spiral ganglion neurons (SGNs), which we iden-tified as Type II SGNs. We therefore conclude that ADF is a
useful molecular tool for marking the small population ofType II SGNs.
MATERIALS AND METHODSAnimals
C57BL/6 mice of both genders were kept with access tofood and water ad libitum according to local and NationalInstitutes of Health (NIH) guidelines. At least two animalswere investigated at each age. If not indicated specifically,chemicals and antibodies were purchased from Sigma Al-drich (Munich, Germany).
Primary antibodies used in the present study are listedin Table 1. The ADF antibody (GV13, #D8815, Sigma Al-drich) recognized a single band of 19 kD on Western blotsof mouse brain, cochlear, and modiolar lysates. No bandwas detectable in brain lysates of ADF-deficient mice (Bel-lenchi et al., 2007) (see also Fig. 2).
Specificity of the calbindin antibody (clone CL-300,#C8666, Sigma Aldrich) was controlled by 2D immuno-blotting in which the antibody specifically stained a 28 kDspot that was identified as calbindin (manufacturer’s tech-nical information). In Western blots the antibody recog-nizes a single 28 kD band in rabbit cerebellum (Celio et al.,1990). Its specificity was further confirmed by failure todetect immunoreactivity in various tissues of calbindinknockout mice (Airaksinen et al., 1997; Wassle et al.,1998). Within the cochlea the antibody yielded hair cell-specific staining identical to that reported before (Rabie etal., 1983; Dechesne and Thomasset, 1988).
The neurofilament 200 antibody (clone NE14, #N5389,Sigma Aldrich) specifically recognizes phosphorylatedneurofilaments of 200 kD in immunohistochemistry. Theantibody shows no immunoreactivity with dephosphory-lated neurofilaments or with other intermediate filamentproteins (manufacturer’s technical information). In immu-nohistochemistry the antibody stained neurofilament-richstructures with an identical pattern of immunoreactivitywhen compared to other antibodies directed against neu-rofilament 200 (Mansour et al., 1989). The antibody isfrequently used in immunohistochemical analyses (Peichland Gonzalez-Soriano, 1993; Paysan et al., 2000; Leclereet al., 2007). In our study the antibody was used as amarker for Type II SGNs and, within the inner ear, yielded apattern of immunoreactivity similar to what was shownbefore with different neurofilament 200 antibodies (Ber-glund and Ryugo, 1991; Hafidi, 1998).
The parvalbumin antibody (clone PA-235, #P3171,Sigma Aldrich) was raised in mouse using purified carpmuscle parvalbumin as the immunogen. Specificity of theantibody was controlled by immunoblotting in which a sin-gle 12 kD band identified as parvalbumin was detected(manufacturer’s technical information). The antibody does
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1725
not react with other members of the EF-hand family ofcalcium-binding proteins (Rymar and Sadikot, 2007).Preadsorption experiments using purified carp muscleparvalbumin eliminated parvalbumin staining in rat braintissue (Heizmann and Celio, 1987). Within the cochlea theantibody yielded hair cell-specific staining identical to thatreported before (Pack and Slepecky, 1995; Eybalin et al.,2002).
Specificity of the SK2 antibody (#P0483, Sigma Aldrich)was verified by loss of immunoreactivity in myocytes isolatedfrom SK2-deficient mice and by loss of immunoreactivity inmyocytes of wildtype mice after preincubation with the re-spective antigenic peptide (Li et al., 2009). Within the organ ofCorti the antibody yielded a specific staining at postsynapticstructures of medio-olivocochlear/outer hair cell synapses asreported before (Oliver et al., 2000; Ruttiger et al., 2004;Winter et al., 2007).
In Western blots using human or mouse protein lysatesthe Sox2 antibody (Y-17, #sc-17320, Santa Cruz Biotech-nology, Santa Cruz, CA) recognizes a single band at 34 kD(manufacturer’s technical information). Specificity wasverified by using a Sox2 blocking peptide that abolishedimmunoreactivity of the antibody in the mouse cochlea(Hume et al., 2007). Within the postnatal mouse cochleathe antibody yielded supporting cell nuclei-specific stain-ing identical to that reported before (Hume et al., 2007;Oesterle et al., 2008).
In immunoblotting the synaptophysin antibody (#sc-7568, Santa Cruz) recognizes a single band at 38 kD inmouse and rat brain lysates (manufacturer’s technical in-formation). Based on mass spectrometry we revealed thesequence of the synaptophysin blocking peptide (#sc-
7568P, Santa Cruz), which is identical to the antigen usedfor immunization, as PQGGYQPDYGQPAGSGGSGYGP (cor-responding to aa 270–291 of human synaptophysin). Theantibody is frequently used to label synaptophysin-richpresynaptic structures in immunohistochemical analyses(Leshchyns’ka et al., 2006; Maas et al., 2006; Sytnyk et al.,2006). The synaptophysin1 antiserum (#101002, SynapticSystems, Goettingen, Germany) stains a single band of�40 kD in synaptosomal fraction of rat brain (manufactur-er’s technical information). The specificity of the antibodyhas been confirmed using immunoblot and immunocyto-chemistry analysis of hippocampal neurons (Synaptic Sys-tems technical information). Specificity was confirmed invarious immunohistochemical studies using mammalianretina or mouse brain (Von Kriegstein et al., 1999; Leitch etal., 2009). Our staining results in mouse cochlea produceda similar pattern of immunoreactivity for both antibodies.Moreover, these patterns are in accordance with previousstudies in which different synaptophysin antibodies wereused as markers for presynaptic structures of outer haircell-innervating medio-olivocochlear projections (Liber-man et al., 1990; Engel et al., 2006).
Western blot analysisProtein lysates were prepared from tissue of at least three
animals per age. In case of the embryonic day (E) 15 sample,the cochlear cartilage was removed and the entire tissue con-tent was collected. At older ages the stria vascularis wasremoved and the organ of Corti and the modiolus were col-lected and processed separately. Tissue was homogenized in20 mM TRIS-HCl (pH 8.0), 100 mM NaCl, 5 mM EGTA, 2 mMEDTA, and 0.5% Triton X-100 containing the protease inhibitor
TABLE 1.Primary Antibodies Used in this Study
Antibody Immunogen Species Catalog No. Lot No. Supplier Dilution
ADF hADF, aa 153–165; Hm. Rb. D8815 032K4885 Sigma-Aldrich 1:400 (IHC) 1:1,500 (IB)Calbindin-D Calbindin-D (28 kD) purified from
chicken gut; Ch.Ms. C8666 074H4816 Sigma-Aldrich 1:1,000
Neurofilament-200 Neurofilament purified from pigspinal cord; Pg.
Ms. N5389 056K4810 Sigma-Aldrich 1:80
Parvalbumin Parvalbumin purified from carpmuscle; Cp.
Ms. P3171 017H4821 Sigma-Aldrich 1:5,000
Phalloidin-TRITC Isolated from Amanita phalloides,fluorescent conjugate
x P1951 Sigma-Aldrich 1:500
SK2 Synthetic peptide correspondingto aa 542–559 of rtSK2; Rt.
Rb. P0483 066K1451 Sigma-Aldrich 1:100
Sox2 Synthetic peptide correspondingto aa 277–293 of hmSOX2;Hm.
Gt. sc-17320 I1708 Santa Cruz 1:200
Synaptophysin Synthetic peptide correspondingto aa 270–291 of hmSYP; Hm.
Gt. sc-7568 E2308 Santa Cruz 1:50
Synaptophysin1 Synthetic peptide correspondingto aa 301–313 of hmSYP; Hm.
Rb. 101002 101002/14 Synaptic Systems 1:100
aa, amino acids; IB, immunoblot; IHC, immunohistochemistry; Ms., mouse; Rb., rabbit; Gt., goat; Hm., human; Rt., rat; Cp., carp; Ch., chicken; Pg., pig.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1726 The Journal of Comparative Neurology � Research in Systems Neuroscience

complete mini (Roche Diagnostics, Mannheim, Germany).Cell debris was removed by centrifuging at 20,800g for 20minutes at 4°C. Sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) was performed and stained withCoomassie to adjust for equal protein amounts of the sam-ples. Incubation of the ADF antibody was performed in 5%milk powder in TRIS-buffered saline (20 mM TRIS-HCl pH 7.5,150 mM NaCl, and 0.1% Tween-20) at 4°C overnight. Afterrinsing with TRIS-buffered saline, the secondary antibodygoat antirabbit/HRP (Pierce, Bonn, Germany #31460;1:5,000) was incubated at room temperature for 1 hour. Fi-nally, the blot was developed using chemiluminescence(PerkinElmer, Waltham, MA).
Tissue preparationFor cryostat sections, animals were sacrificed by cervi-
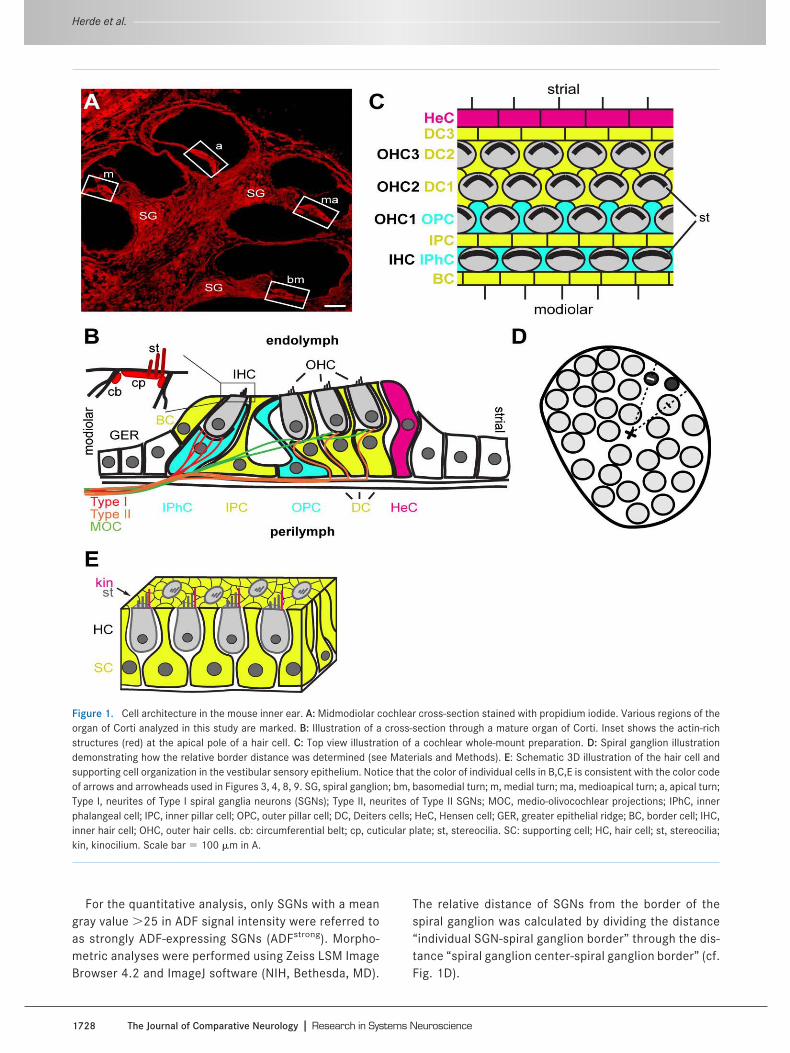
cal dislocation and the inner ears were dissected and fixedin 2% paraformaldehyde (PFA) in phosphate-buffered sa-line (PBS, pH 7.0) for 2 hours at room temperature. Co-chleae of animals older than postnatal day (P)8 were de-calcified using RDO Rapid Bone Decalcifier (ApexEngineering, Aurora, IL), and duration of decalcificationwas calculated according to the following equation: age (indays)/2 � duration (in minutes). Subsequently, cochleaewere cryoprotected at 4°C overnight in 25% sucrose in a 1mM HEPES-buffered solution containing (in mM) 5.36 KCl,0.5 MgCl2, 0.4 MgSO4, 141.7 NaCl, 1.56 CaCl2, 3.42L-glutamine, and 6.3 D-glucose. Inner ears of mice youngerthan P8 were cryoprotected without decalcification at 4°Covernight in 25% sucrose in PBS. Midmodiolar sections of10 �m thickness were prepared using a cryostat (LeicaCM3000, Leica Microsystems, Wetzlar Germany). A mid-modiolar cochlear cross-section stained with propidiumiodide is shown in Figure 1A and various regions of interestare marked. The organization of hair cells and supportingcells is illustrated in Figure 1B,E as seen in cochlear andvestibular cross-sections.
For whole-mount preparations, the inner ear was re-moved as described above. The oval window and the heli-cotrema were opened and the tissue was fixed in 4% PFA/PBS for 2 hours at room temperature. The organ of Cortiwas dissected by removing the cochlear bone, stria vascu-laris, Reissner membrane, and tectorial membrane and bypulling off the basilar membrane from the modiolus. Figure1C illustrates schematically the arrangement of variouscell types of the organ of Corti in a top view. Vestibularepithelia were carefully dissected by removing overlyingmembranes.
ImmunohistochemistryImmunohistochemistry with antibodies against ADF,
calbindin, neurofilament-200, and parvalbumin was per-formed after permeabilizing in 0.5% Triton X-100 in PBS for
10 minutes. After rinsing with PBS, nonspecific bindingwas blocked in 4% goat serum (Invitrogen, Karlsruhe, Ger-many) in PBS for 30 minutes. Primary antibodies were in-cubated at 4°C overnight in PBS containing 1% goat se-rum, 2% NaCl, and 0.1% Triton X-100. After rinsing withPBS, secondary antibodies (goat antirabbit/Alexa-488 #A-11008, 1:1,000; goat antimouse/Alexa-546 #A-11003,1:500; both obtained from Invitrogen, Karlsruhe, Germany)were incubated for 1 hour at room temperature. In someexperiments a nuclear counterstaining with propidium io-dide (Sigma Aldrich #P4170; 1:1,000) was performed for10 minutes in PBS. Whole-mounts were counterstainedwith the F-actin marker Phalloidin-TRITC (Sigma Aldrich#P1951, 1:500) for 20 minutes in PBS. Specimens werethen mounted using a homemade mounting medium basedon glycerol, polyvinylalcohol, and DABCO.
Immunohistochemical labeling with antibodies againstSK2 and synaptophysin was performed by permeabilizingin 0.1% Triton X-100 in PBS for 3 minutes. After rinsing withPBS, nonspecific binding was blocked in 1% bovine serumalbumin (BSA) in PBS for 30 minutes. Primary antibodieswere incubated at 4°C overnight in PBS containing 0.5%BSA. After rinsing with PBS, secondary antibodies (donkeyantirabbit/Cy3 1:1,500, Jackson Laboratories, WestGrove, PA; #711-166-152; donkey antigoat/Alexa-4881:500, Invitrogen #A-11055; goat antimouse/Alexa-546,1:500, Invitrogen #A-11003) were incubated for 1 hour atroom temperature. Specimens were mounted as de-scribed above.
Sox2 staining was performed by permeabilizing in 0.1%saponin and 0.1% Tween-20 in PBS for 30 minutes. Afterrinsing with PBS, nonspecific binding was blocked in 4%BSA, 0.03% saponin, and 0.1% Triton X-100 in PBS for 1hour. Primary antibodies were incubated at 4°C overnightin PBS containing 0.2% BSA, 0.03% saponin and 0.1% Tri-ton X-100. After rinsing with PBS, secondary antibodies(donkey antigoat/Alexa-546 1:500, Invitrogen #A-11056;goat antirabbit/Alexa-488 1:1,000, Invitrogen #A-11008)and Hoechst 33342 (Invitrogen #H1399; 1:1,000) wereapplied for 1 hour at room temperature. Specimens weremounted as described above.
Microscopy and image analysisImages were acquired on a Zeiss LSM 510 confocal
microscope (Carl Zeiss, Gottingen, Germany) using 10�/0.3, 40�/1.3, and 63�/1.4 objectives. Image stackswere projected with the Zeiss LSM Image Browser 4.2 inthe maximum intensity mode. Images were then exportedto the tiff-format and processed for brightness and con-trast with the Adobe Photoshop CS4 software (San Jose,CA). Figure panels were generated using Adobe IllustratorCS4 software.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1727
For the quantitative analysis, only SGNs with a meangray value �25 in ADF signal intensity were referred toas strongly ADF-expressing SGNs (ADFstrong). Morpho-metric analyses were performed using Zeiss LSM ImageBrowser 4.2 and ImageJ software (NIH, Bethesda, MD).
The relative distance of SGNs from the border of thespiral ganglion was calculated by dividing the distance“individual SGN-spiral ganglion border” through the dis-tance “spiral ganglion center-spiral ganglion border” (cf.Fig. 1D).
Figure 1. Cell architecture in the mouse inner ear. A: Midmodiolar cochlear cross-section stained with propidium iodide. Various regions of theorgan of Corti analyzed in this study are marked. B: Illustration of a cross-section through a mature organ of Corti. Inset shows the actin-richstructures (red) at the apical pole of a hair cell. C: Top view illustration of a cochlear whole-mount preparation. D: Spiral ganglion illustrationdemonstrating how the relative border distance was determined (see Materials and Methods). E: Schematic 3D illustration of the hair cell andsupporting cell organization in the vestibular sensory epithelium. Notice that the color of individual cells in B,C,E is consistent with the color codeof arrows and arrowheads used in Figures 3, 4, 8, 9. SG, spiral ganglion; bm, basomedial turn; m, medial turn; ma, medioapical turn; a, apical turn;Type I, neurites of Type I spiral ganglia neurons (SGNs); Type II, neurites of Type II SGNs; MOC, medio-olivocochlear projections; IPhC, innerphalangeal cell; IPC, inner pillar cell; OPC, outer pillar cell; DC, Deiters cells; HeC, Hensen cell; GER, greater epithelial ridge; BC, border cell; IHC,inner hair cell; OHC, outer hair cells. cb: circumferential belt; cp, cuticular plate; st, stereocilia. SC: supporting cell; HC, hair cell; st, stereocilia;kin, kinocilium. Scale bar � 100 �m in A.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1728 The Journal of Comparative Neurology � Research in Systems Neuroscience

Statistical analysisStatistical significance was calculated using Student’s
t-test. Values P � 0.05 were referred to as statisticallysignificant and marked by an asterisk (*P � 0.05). Data inFigure 7A–C are presented as mean � standard error ofthe mean.
RESULTSADF expression in the inner ear duringembryonic and postnatal development
In a first step, the expression of ADF in the fetal andpostnatal murine inner ear was investigated by immuno-blot analysis, using total cochlear protein lysates of em-bryos at day 15 and protein lysates of the modiolus as wellas the organ of Corti at various postnatal stages (P0–P26).The specificity of the antibody was confirmed by usingtotal brain lysates of wildtype (WT) and ADF-deficient(ADF/) mice (Fig. 2A, left lanes). In the cochlea, ADFwas present throughout fetal and postnatal development.Whereas the expression became upregulated in the modi-olus after hearing onset (Fig. 2A) which, in mice, occurs atabout P10–12 (Mikaelian et al., 1965; Ehret, 1976), itgradually decreased in the organ of Corti (Fig. 2B). Wetherefore decided to investigate the expression and loca-tion of ADF in the modiolus and various turns of the organof Corti in more detail. To do so, immunohistochemistrywas performed on both cochlear whole-mount prepara-tions and cross-sections.
ADF expression in hair cells and supportingcells during organ of Corti development
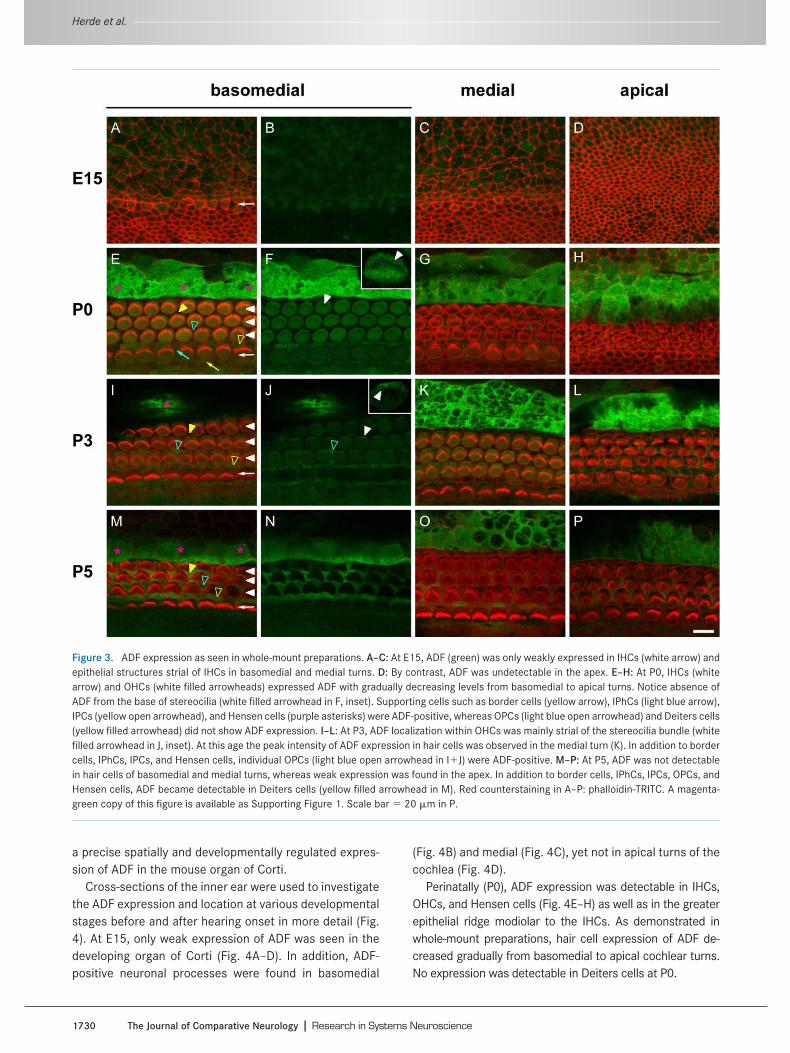
In whole-mount preparations of the organ of Corti atE15, weak ADF expression occurred in inner hair cells
(IHCs) of basomedial turns (Fig. 3A,B) and, to some extent,in medial turns (Fig. 3C). Furthermore, ADF was observedin epithelial structures strial to IHCs. No ADF expressionwas observed in apical turns of the cochlea (Fig. 3D),where cochlear development is delayed compared to thebasal parts (Bruce et al., 1997).
At P0, both IHCs and outer hair cells (OHCs) showedADF expression with a gradual decrease in signal intensityfrom basomedial to apical turns (Fig. 3E–H). Within haircells, ADF was absent from the base of stereociliary bun-dles (Fig. 3F). Around birth, ADF expression was not re-stricted to sensory hair cells. Hensen cells, which consti-tute the outer border of the organ of Corti, showed asubstantial amount of ADF expression in all turns of thecochlea (Fig. 3E–H). Aside from Hensen cells, inner pillarcells (IPCs), inner phalangeal cells (IPhCs), and border cellsalso expressed ADF. No ADF was found in Deiters cells andouter pillar cells (OPCs).
During early postnatal development (P3), peak expressionof ADF in hair cells shifted from basomedial to medial turns ofthe cochlea (Fig. 3I–L). Within OHCs, the ADF signal was mostprominent strial to stereocilia bundles (Fig. 3J). Again, ADFexpression was seen in Hensen cells, IPCs, IPhCs, and bordercells. At P3, Deiters cells showed no ADF expression,whereas individual OPCs expressed ADF.
During subsequent development (P5), no ADF expres-sion was detectable in hair cells of basomedial and medialturns and only weak expression was found in hair cells ofthe apex (Fig. 3M–P). Aside from Hensen cells, IPCs,IPhCs, and border cells, ADF expression became obviousin Deiters cells and OPCs. Taken together, our lateembryonic/early postnatal whole-mount analysis revealed
Figure 2. Developmental expression of ADF in the mouse inner ear. A: ADF presence in brain lysates of wildtype mice (WT) and absence in brainlysates of ADF-deficient mice (ADF/), demonstrating the specificity of the antibody. At E15, ADF was found in total cochlea protein lysates.Analysis of modiolus protein lysates revealed stronger ADF expression after hearing onset (P18, P26) compared to the younger stages (P0, P9).B: During postnatal development, ADF expression gradually decreased in the organ of Corti until P26.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1729
a precise spatially and developmentally regulated expres-sion of ADF in the mouse organ of Corti.
Cross-sections of the inner ear were used to investigatethe ADF expression and location at various developmentalstages before and after hearing onset in more detail (Fig.4). At E15, only weak expression of ADF was seen in thedeveloping organ of Corti (Fig. 4A–D). In addition, ADF-positive neuronal processes were found in basomedial
(Fig. 4B) and medial (Fig. 4C), yet not in apical turns of thecochlea (Fig. 4D).
Perinatally (P0), ADF expression was detectable in IHCs,OHCs, and Hensen cells (Fig. 4E–H) as well as in the greaterepithelial ridge modiolar to the IHCs. As demonstrated inwhole-mount preparations, hair cell expression of ADF de-creased gradually from basomedial to apical cochlear turns.No expression was detectable in Deiters cells at P0.
Figure 3. ADF expression as seen in whole-mount preparations. A–C: At E15, ADF (green) was only weakly expressed in IHCs (white arrow) andepithelial structures strial of IHCs in basomedial and medial turns. D: By contrast, ADF was undetectable in the apex. E–H: At P0, IHCs (whitearrow) and OHCs (white filled arrowheads) expressed ADF with gradually decreasing levels from basomedial to apical turns. Notice absence ofADF from the base of stereocilia (white filled arrowhead in F, inset). Supporting cells such as border cells (yellow arrow), IPhCs (light blue arrow),IPCs (yellow open arrowhead), and Hensen cells (purple asterisks) were ADF-positive, whereas OPCs (light blue open arrowhead) and Deiters cells(yellow filled arrowhead) did not show ADF expression. I–L: At P3, ADF localization within OHCs was mainly strial of the stereocilia bundle (whitefilled arrowhead in J, inset). At this age the peak intensity of ADF expression in hair cells was observed in the medial turn (K). In addition to bordercells, IPhCs, IPCs, and Hensen cells, individual OPCs (light blue open arrowhead in IJ) were ADF-positive. M–P: At P5, ADF was not detectablein hair cells of basomedial and medial turns, whereas weak expression was found in the apex. In addition to border cells, IPhCs, IPCs, OPCs, andHensen cells, ADF became detectable in Deiters cells (yellow filled arrowhead in M). Red counterstaining in A–P: phalloidin-TRITC. A magenta-green copy of this figure is available as Supporting Figure 1. Scale bar � 20 �m in P.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1730 The Journal of Comparative Neurology � Research in Systems Neuroscience
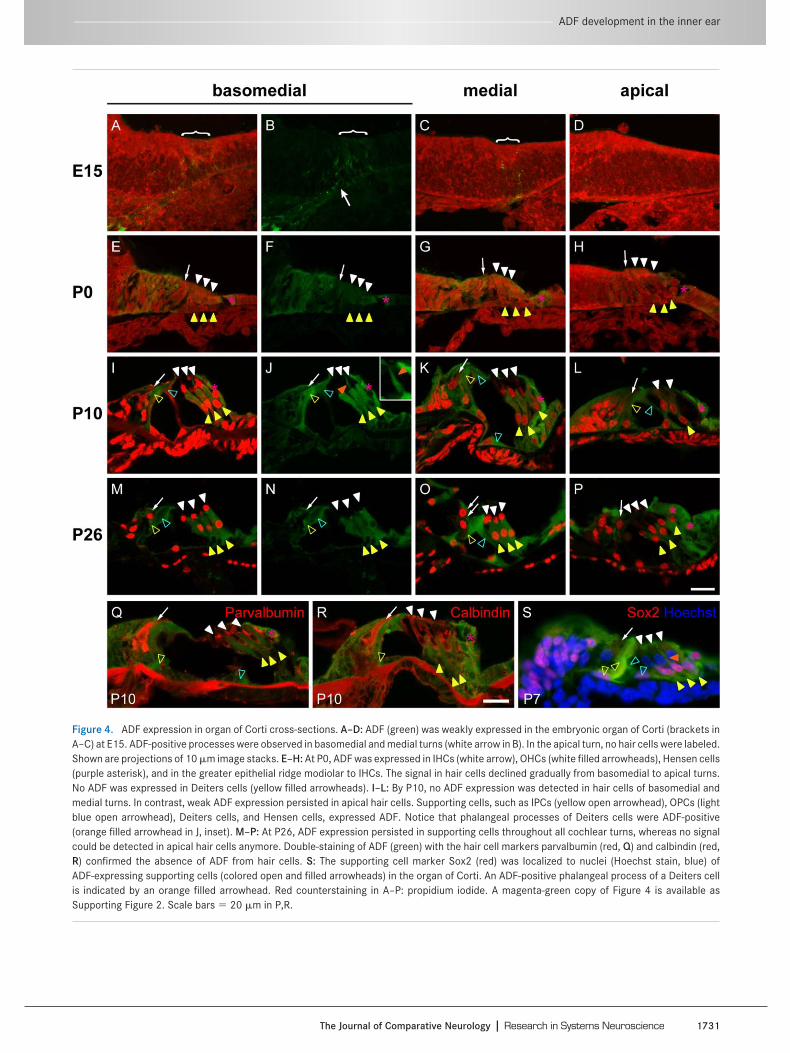
Figure 4. ADF expression in organ of Corti cross-sections. A–D: ADF (green) was weakly expressed in the embryonic organ of Corti (brackets inA–C) at E15. ADF-positive processes were observed in basomedial and medial turns (white arrow in B). In the apical turn, no hair cells were labeled.Shown are projections of 10 �m image stacks. E–H: At P0, ADF was expressed in IHCs (white arrow), OHCs (white filled arrowheads), Hensen cells(purple asterisk), and in the greater epithelial ridge modiolar to IHCs. The signal in hair cells declined gradually from basomedial to apical turns.No ADF was expressed in Deiters cells (yellow filled arrowheads). I–L: By P10, no ADF expression was detected in hair cells of basomedial andmedial turns. In contrast, weak ADF expression persisted in apical hair cells. Supporting cells, such as IPCs (yellow open arrowhead), OPCs (lightblue open arrowhead), Deiters cells, and Hensen cells, expressed ADF. Notice that phalangeal processes of Deiters cells were ADF-positive(orange filled arrowhead in J, inset). M–P: At P26, ADF expression persisted in supporting cells throughout all cochlear turns, whereas no signalcould be detected in apical hair cells anymore. Double-staining of ADF (green) with the hair cell markers parvalbumin (red, Q) and calbindin (red,R) confirmed the absence of ADF from hair cells. S: The supporting cell marker Sox2 (red) was localized to nuclei (Hoechst stain, blue) ofADF-expressing supporting cells (colored open and filled arrowheads) in the organ of Corti. An ADF-positive phalangeal process of a Deiters cellis indicated by an orange filled arrowhead. Red counterstaining in A–P: propidium iodide. A magenta-green copy of Figure 4 is available asSupporting Figure 2. Scale bars � 20 �m in P,R.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1731
At P10, i.e., shortly before hearing onset, ADF expres-sion in IHCs and OHCs was absent in basomedial and me-dial turns (Fig. 4I–K) and barely detectable in apical turns(Fig. 4L). As shown in whole-mount preparations, varioustypes of supporting cells, such as Hensen cells, Deiterscells, IPCs, and OPCs, expressed ADF at substantial levels.In Deiters cells, ADF was found in somata and phalangealprocesses (Fig. 4J).
At P26, roughly 2 weeks after hearing onset, ADF ex-pression remained dominant in supporting cells and wasabsent from hair cells (Fig. 4M–P). The absence of ADFfrom IHCs and OHCs was confirmed via colocalization ex-periments using the Ca2-binding proteins parvalbumin(Fig. 4Q) and calbindin (Fig. 4R) as hair cell markers(Dechesne and Thomasset, 1988; Xiang et al., 1998). Con-currently, labeling for Sox2 as a marker for supporting cells
(Hume et al., 2007) confirmed prominent ADF expressionin Deiters cells, Hensen cells, and pillar cells (Fig. 4S).
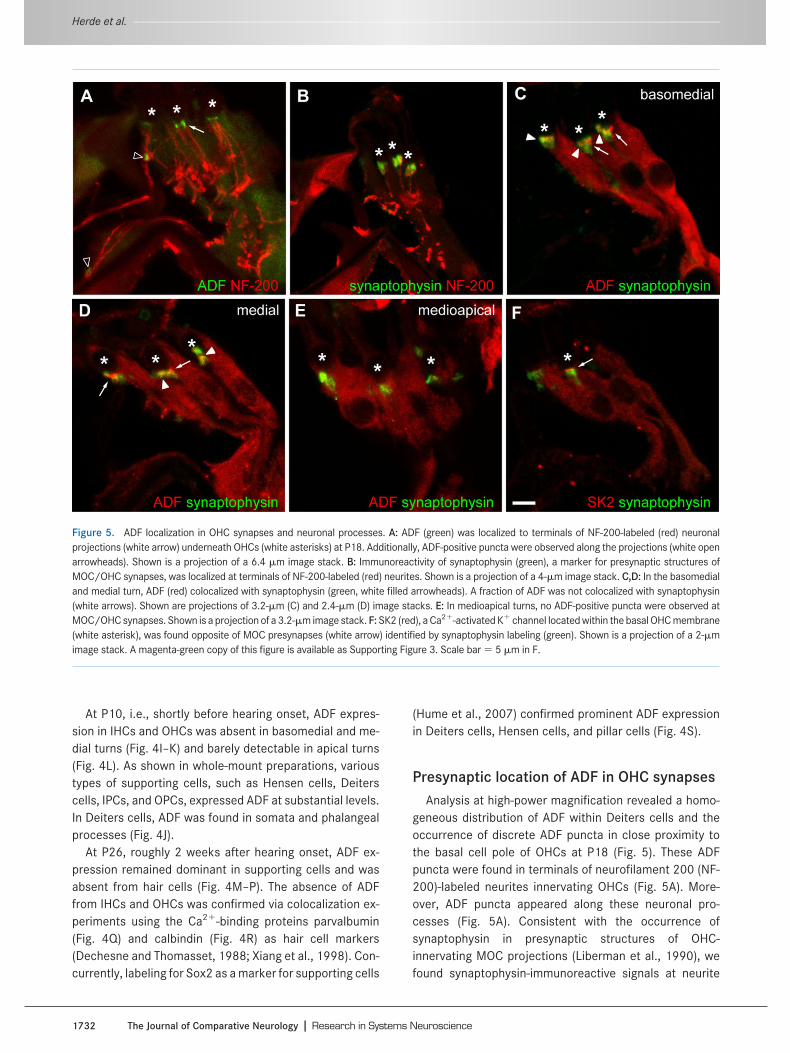
Presynaptic location of ADF in OHC synapsesAnalysis at high-power magnification revealed a homo-
geneous distribution of ADF within Deiters cells and theoccurrence of discrete ADF puncta in close proximity tothe basal cell pole of OHCs at P18 (Fig. 5). These ADFpuncta were found in terminals of neurofilament 200 (NF-200)-labeled neurites innervating OHCs (Fig. 5A). More-over, ADF puncta appeared along these neuronal pro-cesses (Fig. 5A). Consistent with the occurrence ofsynaptophysin in presynaptic structures of OHC-innervating MOC projections (Liberman et al., 1990), wefound synaptophysin-immunoreactive signals at neurite
Figure 5. ADF localization in OHC synapses and neuronal processes. A: ADF (green) was localized to terminals of NF-200-labeled (red) neuronalprojections (white arrow) underneath OHCs (white asterisks) at P18. Additionally, ADF-positive puncta were observed along the projections (white openarrowheads). Shown is a projection of a 6.4 �m image stack. B: Immunoreactivity of synaptophysin (green), a marker for presynaptic structures ofMOC/OHC synapses, was localized at terminals of NF-200-labeled (red) neurites. Shown is a projection of a 4-�m image stack. C,D: In the basomedialand medial turn, ADF (red) colocalized with synaptophysin (green, white filled arrowheads). A fraction of ADF was not colocalized with synaptophysin(white arrows). Shown are projections of 3.2-�m (C) and 2.4-�m (D) image stacks. E: In medioapical turns, no ADF-positive puncta were observed atMOC/OHC synapses. Shown is a projection of a 3.2-�m image stack. F: SK2 (red), a Ca2-activated K channel located within the basal OHC membrane(white asterisk), was found opposite of MOC presynapses (white arrow) identified by synaptophysin labeling (green). Shown is a projection of a 2-�mimage stack. A magenta-green copy of this figure is available as Supporting Figure 3. Scale bar � 5 �m in F.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1732 The Journal of Comparative Neurology � Research in Systems Neuroscience

terminals (Fig. 5B). Double-labeling with synaptophysindemonstrated presynaptic location of ADF in basomedialand medial turns (Fig. 5C,D), but not in medioapical (Fig.5E) and apical turns (not shown). However, a fraction ofADF did not colocalize with synaptophysin; instead it wasfound rather lateral of the synaptophysin signal (Fig. 5C,D).SK2 is a Ca2-activated K channel that, in the organ ofCorti, is located at postsynaptic structures of MOC/OHCsynapses (Oliver et al., 2000). Indeed, we found SK2 op-posing the synaptophysin-labeled presynapses (Fig. 5F).Taken together, we localized ADF in presynaptic structuresof MOC/OHC synapses. Moreover, by comparing ADF-and SK2-immunoreactive signals with synaptophysin-labeled structures, we could exclude ADF from postsynap-tic structures of MOC/OHC synapses.
Developmental expression of ADF in spiralganglion neurons
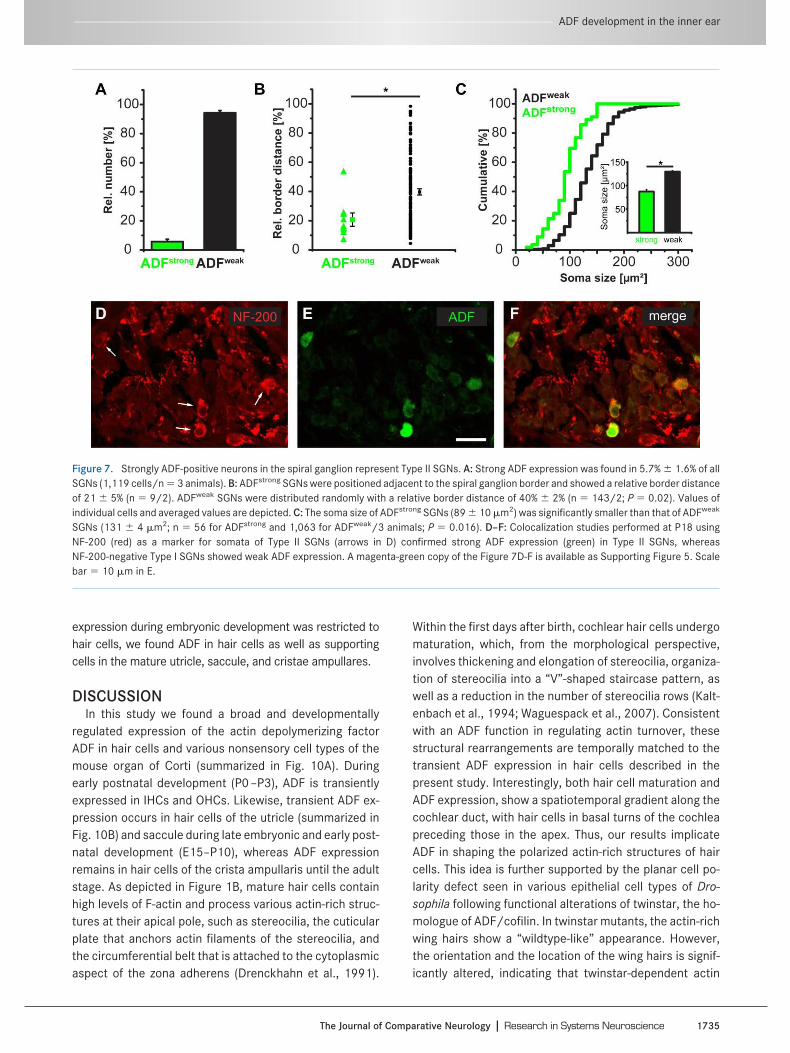
There is a broad neuronal expression of ADF in the cen-tral nervous system (Bellenchi et al., 2007). We thereforeinvestigated the expression of ADF in peripheral auditoryneurons by analyzing cochlear cross-sections at twostages: before (P0, P10) and after hearing onset (P18,P26). Before hearing onset, ADF expression in the spiralganglion was only minor. At P0, no ADF expression wasobserved throughout any cochlear turn (Fig. 6A–D). AtP10, weak ADF expression became detectable in a smallsubpopulation (6%) of SGNs predominantly in the basome-dial part (Fig. 6E) and somehow weaker also in medial (Fig.6G) and apical parts (Fig. 6F–H).
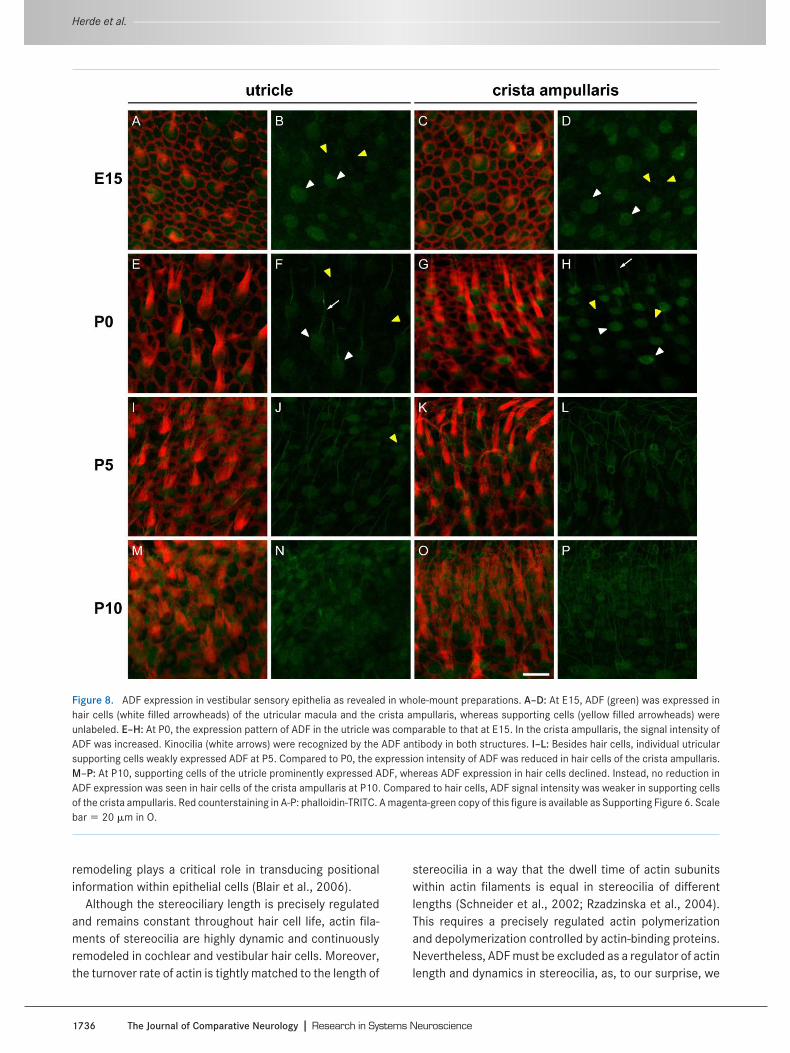
ADF expression in SGNs increased further after hearingonset. At P18 and P26, a few neurons preferentially at thespiral ganglion border showed substantial ADF expression(Fig. 6I–P). In contrast, ADF expression remained ratherweak in the majority of SGNs. This pointed to the presenceof two populations of SGNs, a minority of SGNs with strongADF expression (referred to as ADFstrong) more located inthe spiral ganglion periphery, and a majority of SGNs withweak ADF expression (ADFweak). ADFweak SGNs were ran-domly distributed throughout the ganglion. A peripherallocation and a relative amount of 5%–10% are hallmarks forType II SGNs (Schwartz et al., 1983; Raphael and Alt-schuler, 2003). We therefore asked whether ADFstrong neu-rons indeed represent Type II SGNs and investigated thenumber and spatial distribution of both neuron populationsin more detail. This analysis was performed at P26 andrevealed that ADFstrong neurons amounted for 5.7% � 1.6%of all SGNs (Fig. 7A; n � 1,119 cells/3 mice) and wereconcentrated at the periphery of the spiral ganglion ratherthan in central aspects (n � 9 cells/2 mice, Fig. 7B). Theremaining 94.3% � 1.6% neurons displayed a weak ADFexpression and were randomly distributed throughout thespiral ganglion (n � 143 cells/2 mice; P � 0.02). A third
parameter, previously used to distinguish Type II from TypeI SGNs, is their soma size: Type II SGNs are smaller (cf. Fig.3 of Berglund and Ryugo, 1991). Analysis of the SGN somaarea in cross-sections revealed a mean value of 89 � 10�m2 (n � 56 cells/3 mice) for ADFstrong neurons (Fig. 7C).These cells were significantly smaller than ADFweak neu-rons whose cell body size was 131 � 4 �m2 (n � 1,063cells/3 mice; P � 0.016). NF-200 was previously shown tolabel somata of Type II SGNs but not of Type I SGNs (Berg-lund and Ryugo, 1991; Hafidi, 1998). We therefore per-formed double-labeling experiments of ADF and NF-200 inthe spiral ganglion and found strong ADF expression exclu-sively in somata of NF-200-positive SGNs (Fig. 7D–F), im-plying that ADF can be used to specifically identify Type IIneurons in the spiral ganglion.
Developmental expression of ADF in thevestibular system
Besides the cochlea, the inner ear contains five addi-tional sensory epithelia that consist of hair cells and sup-porting cells: the utricle and saccule which detect transla-tional acceleration, and the sensory organs of the threesemicircular canals (called cristae ampullares) which de-tect rotational acceleration. Given the parallels in struc-ture, we asked whether ADF is also expressed in the dif-ferent cell types of the vestibular epithelia. As performedfor the cochlea, we assessed the expression of ADF in theutricle, the saccule, and the cristae ampullares at variousdevelopmental stages by analyzing both whole-mountpreparations and cross-sections.
Immunostaining of whole-mount preparations at E15 re-vealed ADF expression in hair cells of the utricle (Fig.8A,B), the crista ampullaris (Fig. 8C,D), and the saccule(not shown). At this stage, ADF was not present in support-ing cells in any of these structures (Fig. 8A–D). During earlypostnatal development (P0–P5), ADF expression was de-tectable in hair cells, but not in supporting cells, by analy-sis of whole-mount preparations (Fig. 8E–L). Aside from adiffuse cytoplasmic signal, we found ADF labeling in thekinocilia of hair cells (Fig. 8F,H). Unlike stereocilia, kino-cilia represent true cilia that mainly consist of microtu-bules (Kikuchi et al., 1989). As there is no actin in thesestructures, the localization of ADF in kinocilia was unex-pected and surprising. Indeed, preliminary control experi-ments performed by us in ADF/ mice indicated that thisADF signal is an artifact, as kinocilia were stained in theknockouts too (not shown). Our result is in line with thelabeling pattern obtained with other antibodies directedagainst proteins that are not thought to be present in ki-nocilia, for example, glial fibrillary acidic protein (GFAP; MKHerde and KP Steel, unpubl. obs.). The ADF expressionpattern in vestibular sensory epithelia had changed byP10, when the immunofluorescent signals became weaker
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1733
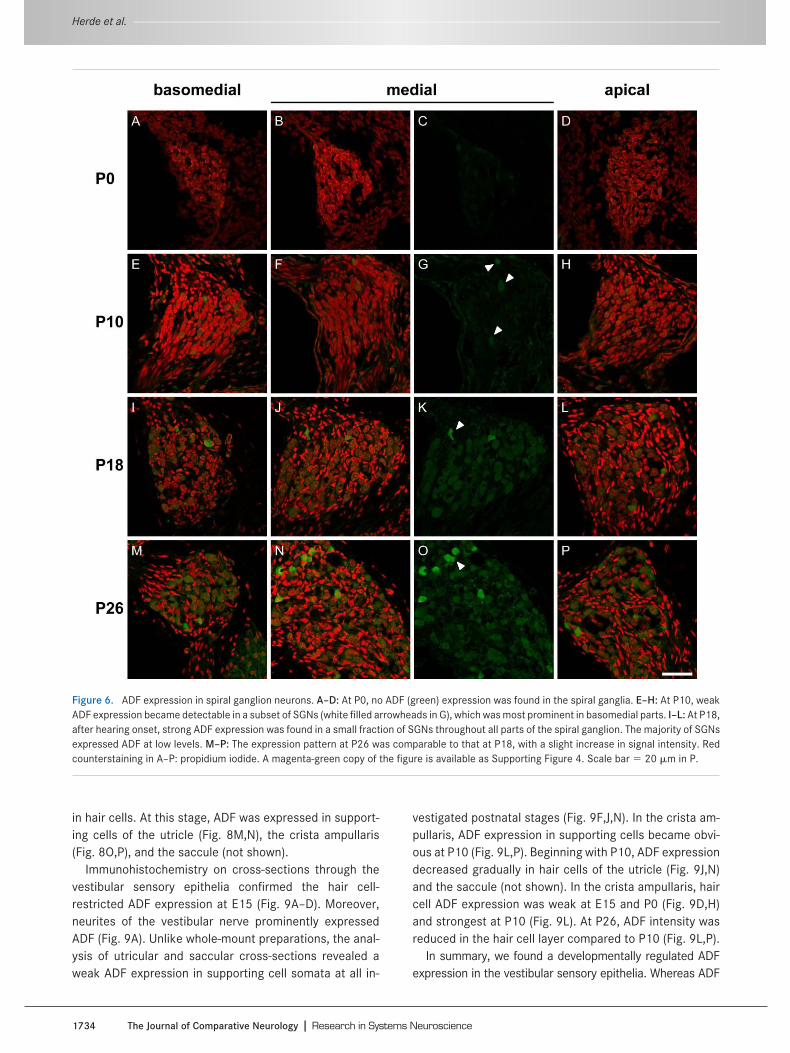
in hair cells. At this stage, ADF was expressed in support-ing cells of the utricle (Fig. 8M,N), the crista ampullaris(Fig. 8O,P), and the saccule (not shown).
Immunohistochemistry on cross-sections through thevestibular sensory epithelia confirmed the hair cell-restricted ADF expression at E15 (Fig. 9A–D). Moreover,neurites of the vestibular nerve prominently expressedADF (Fig. 9A). Unlike whole-mount preparations, the anal-ysis of utricular and saccular cross-sections revealed aweak ADF expression in supporting cell somata at all in-
vestigated postnatal stages (Fig. 9F,J,N). In the crista am-pullaris, ADF expression in supporting cells became obvi-ous at P10 (Fig. 9L,P). Beginning with P10, ADF expressiondecreased gradually in hair cells of the utricle (Fig. 9J,N)and the saccule (not shown). In the crista ampullaris, haircell ADF expression was weak at E15 and P0 (Fig. 9D,H)and strongest at P10 (Fig. 9L). At P26, ADF intensity wasreduced in the hair cell layer compared to P10 (Fig. 9L,P).
In summary, we found a developmentally regulated ADFexpression in the vestibular sensory epithelia. Whereas ADF
Figure 6. ADF expression in spiral ganglion neurons. A–D: At P0, no ADF (green) expression was found in the spiral ganglia. E–H: At P10, weakADF expression became detectable in a subset of SGNs (white filled arrowheads in G), which was most prominent in basomedial parts. I–L: At P18,after hearing onset, strong ADF expression was found in a small fraction of SGNs throughout all parts of the spiral ganglion. The majority of SGNsexpressed ADF at low levels. M–P: The expression pattern at P26 was comparable to that at P18, with a slight increase in signal intensity. Redcounterstaining in A–P: propidium iodide. A magenta-green copy of the figure is available as Supporting Figure 4. Scale bar � 20 �m in P.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1734 The Journal of Comparative Neurology � Research in Systems Neuroscience
expression during embryonic development was restricted tohair cells, we found ADF in hair cells as well as supportingcells in the mature utricle, saccule, and cristae ampullares.
DISCUSSIONIn this study we found a broad and developmentally
regulated expression of the actin depolymerizing factorADF in hair cells and various nonsensory cell types of themouse organ of Corti (summarized in Fig. 10A). Duringearly postnatal development (P0–P3), ADF is transientlyexpressed in IHCs and OHCs. Likewise, transient ADF ex-pression occurs in hair cells of the utricle (summarized inFig. 10B) and saccule during late embryonic and early post-natal development (E15–P10), whereas ADF expressionremains in hair cells of the crista ampullaris until the adultstage. As depicted in Figure 1B, mature hair cells containhigh levels of F-actin and process various actin-rich struc-tures at their apical pole, such as stereocilia, the cuticularplate that anchors actin filaments of the stereocilia, andthe circumferential belt that is attached to the cytoplasmicaspect of the zona adherens (Drenckhahn et al., 1991).
Within the first days after birth, cochlear hair cells undergomaturation, which, from the morphological perspective,involves thickening and elongation of stereocilia, organiza-tion of stereocilia into a “V”-shaped staircase pattern, aswell as a reduction in the number of stereocilia rows (Kalt-enbach et al., 1994; Waguespack et al., 2007). Consistentwith an ADF function in regulating actin turnover, thesestructural rearrangements are temporally matched to thetransient ADF expression in hair cells described in thepresent study. Interestingly, both hair cell maturation andADF expression, show a spatiotemporal gradient along thecochlear duct, with hair cells in basal turns of the cochleapreceding those in the apex. Thus, our results implicateADF in shaping the polarized actin-rich structures of haircells. This idea is further supported by the planar cell po-larity defect seen in various epithelial cell types of Dro-sophila following functional alterations of twinstar, the ho-mologue of ADF/cofilin. In twinstar mutants, the actin-richwing hairs show a “wildtype-like” appearance. However,the orientation and the location of the wing hairs is signif-icantly altered, indicating that twinstar-dependent actin
Figure 7. Strongly ADF-positive neurons in the spiral ganglion represent Type II SGNs. A: Strong ADF expression was found in 5.7% � 1.6% of allSGNs (1,119 cells/n � 3 animals). B: ADFstrong SGNs were positioned adjacent to the spiral ganglion border and showed a relative border distanceof 21 � 5% (n � 9/2). ADFweak SGNs were distributed randomly with a relative border distance of 40% � 2% (n � 143/2; P � 0.02). Values ofindividual cells and averaged values are depicted. C: The soma size of ADFstrong SGNs (89 � 10 �m2) was significantly smaller than that of ADFweak
SGNs (131 � 4 �m2; n � 56 for ADFstrong and 1,063 for ADFweak/3 animals; P � 0.016). D–F: Colocalization studies performed at P18 usingNF-200 (red) as a marker for somata of Type II SGNs (arrows in D) confirmed strong ADF expression (green) in Type II SGNs, whereasNF-200-negative Type I SGNs showed weak ADF expression. A magenta-green copy of the Figure 7D-F is available as Supporting Figure 5. Scalebar � 10 �m in E.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1735
remodeling plays a critical role in transducing positionalinformation within epithelial cells (Blair et al., 2006).
Although the stereociliary length is precisely regulatedand remains constant throughout hair cell life, actin fila-ments of stereocilia are highly dynamic and continuouslyremodeled in cochlear and vestibular hair cells. Moreover,the turnover rate of actin is tightly matched to the length of
stereocilia in a way that the dwell time of actin subunitswithin actin filaments is equal in stereocilia of differentlengths (Schneider et al., 2002; Rzadzinska et al., 2004).This requires a precisely regulated actin polymerizationand depolymerization controlled by actin-binding proteins.Nevertheless, ADF must be excluded as a regulator of actinlength and dynamics in stereocilia, as, to our surprise, we
Figure 8. ADF expression in vestibular sensory epithelia as revealed in whole-mount preparations. A–D: At E15, ADF (green) was expressed inhair cells (white filled arrowheads) of the utricular macula and the crista ampullaris, whereas supporting cells (yellow filled arrowheads) wereunlabeled. E–H: At P0, the expression pattern of ADF in the utricle was comparable to that at E15. In the crista ampullaris, the signal intensity ofADF was increased. Kinocilia (white arrows) were recognized by the ADF antibody in both structures. I–L: Besides hair cells, individual utricularsupporting cells weakly expressed ADF at P5. Compared to P0, the expression intensity of ADF was reduced in hair cells of the crista ampullaris.M–P: At P10, supporting cells of the utricle prominently expressed ADF, whereas ADF expression in hair cells declined. Instead, no reduction inADF expression was seen in hair cells of the crista ampullaris at P10. Compared to hair cells, ADF signal intensity was weaker in supporting cellsof the crista ampullaris. Red counterstaining in A-P: phalloidin-TRITC. A magenta-green copy of this figure is available as Supporting Figure 6. Scalebar � 20 �m in O.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1736 The Journal of Comparative Neurology � Research in Systems Neuroscience

found ADF neither in the stereocilia of cochlear nor ofvestibular hair cells at any developmental age. Further-more, ADF is not expressed at detectable levels in maturecochlear hair cells. Thus, other proteins with actin depoly-merizing activity, such as the broadly expressed n-cofilin(Gurniak et al., 2005), are potential candidates for regulat-ing the actin filament length and dynamics in stereocilia.
Before hearing onset, but also in the fully developedorgan of Corti, ADF is prominently expressed in varioustypes of supporting cells, including Hensen cells, Deiterscells, and both types of pillar cells. Likewise, ADF is ex-pressed in supporting cells of postnatal and mature ves-tibular sensory epithelia. Supporting cells are critically in-volved in K-recycling, which is essential for proper
Figure 9. ADF expression in vestibular sensory epithelia as revealed in cross-sections. A–D: At E15, hair cells of the utricle and the cristaampullaris expressed ADF (white filled arrowheads), but there was no ADF signal in supporting cells (yellow filled arrowhead). Neuronalprojections (white arrows) were prominently ADF-positive. Shown are projections of 12-�m (A,B) and 6-�m (C,D) image stacks. E–H: At P0, inaddition to hair cells, weak ADF expression was found in supporting cells of the utricle, but not of the crista ampullaris. I–L: At P10, decreased ADFlevels were found in the utricle, whereas signal intensity increased in the crista ampullaris. At this age, structures beneath the epithelia becamelabeled by the ADF antibody. M–P: At P26, the expression of ADF further decreased in the utricle. In the crista ampullaris, the ADF intensity in thehair cell layer was reduced compared to P10, whereas the apical surface and the supporting cell layer were strongly ADF-positive. Redcounterstaining in A-P: propidium iodide. A magenta-green copy of this figure is available as Supporting Figure 7. Scale bars � 20 �m in M (utricle)and O (crista ampullaris).
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1737

mechano-electrical transduction (Boettger et al., 2002,2003). Moreover, supporting cells display distinctive mor-phological features that are critical for the structural integ-rity of the organ of Corti and for its mechanical flexibilitythat allows the movement of the sensory epithelium inresponse to mechanical stimuli (Raphael and Altschuler,2003; Kelly and Chen, 2007). Deiters cells and pillar cellspossess actin-rich phalangeal (“finger-like”) protrusionsthat form tight contacts with hair cells at the surface of theorgan of Corti (Raphael and Altschuler, 2003). The tightjuxtaposition of hair cells and supporting cells is essentialfor sustaining the positive endocochlear potential, be-cause it separates the endolymph (scala media) from theperilymph (scala tympani; Kelly and Chen, 2007). It wasspeculated that actin plays a role in maintaining a contin-uous epithelial barrier between both fluid compartments(Oh et al., 2002). Indeed, a close physical interaction of theactin cytoskeleton with cell adhesion proteins of epithelialtight junctions is well known and intensively described formany cell types (review: Miyoshi and Takai, 2008). Wefound ADF in Deiters cells and pillar cells at substantiallevels before and after hearing onset and localized theprotein to phalangeal processes of these cells. Thus, acontribution of ADF in organizing the structural backboneof phalangeal processes is likely, suggesting that ADF isinvolved in regulating the endolymph/perilymph barrier.
Upon sound stimulation, Deiters cells and Hensen cellsreversibly move toward the center of the cochlear turn in a
contractile fashion (Flock et al., 1999). This movementcoincides with a reduced sensitivity to acoustic stimula-tion. Therefore, it was speculated that Deiters cells andHensen cells are actively involved in protecting the organof Corti against trauma during high-intensity sound expo-sure. The phalangeal processes of Deiters cells undergostructural rearrangements, and flexion of these processeswas implied in the movement of Deiters cells and the at-tached Hensen cells (Dulon et al., 1994; Flock et al., 1999).Both the reversible movement of supporting cells and theflexion of phalangeal processes require a highly dynamiccytoskeleton. As an actin depolymerizing factor, ADF sev-ers actin filaments and enhances the dissociation rate atthe filament minus end and therefore accelerates actindynamics (Bamburg and Wiggan, 2002). The strong ex-pression of ADF in Deiters cells and Hensen cells suggeststhat ADF is involved in regulating cochlear micromechan-ics during intense sound stimulation and, as a result, inprotection from acoustic trauma.
Besides the expression of ADF in sensory and nonsen-sory cells of the organ of Corti, we found developmentallyregulated ADF expression in SGNs. The spiral ganglion iscomposed of two populations of neurons: 1) Type I SGNsinnervated by IHCs, comprising the majority (90%–95%) ofSGNs; 2) Type II SGNs forming a small population (5%–10%) and receiving input from OHCs (Raphael and Alt-schuler, 2003). Type II SGNs are located at the periphery ofthe spiral ganglion (Schwartz et al., 1983) and are distin-
Figure 10. Semischematic illustrations summarizing the differential expression of ADF during development of mouse inner ear sensory epithelia.Depicted are three developmental stages of the basomedial turn of the organ of Corti and the spiral ganglion (P0, P5, P26) (A) and of the utricle(E15, P0, P26) (B). White arrows at P26 in A point to the synaptic localization of ADF. Notice that gray values in A,B reflect the intensity of the ADFsignal in individual cells and structures, with dark gray implying a high ADF expression.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1738 The Journal of Comparative Neurology � Research in Systems Neuroscience

guishable from Type I SGNs by their smaller cell bodies (cf.fig. 3 in Berglund and Ryugo, 1991). Furthermore, specificmarkers such as neurofilament NF-200, which are ex-pressed in somata of Type II, but not of Type I SGNs, areuseful probes to distinguish both populations of SGNs(Berglund and Ryugo, 1991). In the mature cochlea wefound a strong ADF expression in roughly 6% of all SGNs,whereas the majority of SGN neurons showed only weakADF staining. Based on their relative number, but also ontheir smaller soma size and localization to more peripheralparts within the spiral ganglion, we identified ADFstrong
neurons as Type II SGNs. This finding was confirmed bydouble-labeling experiments with ADF and NF-200.Whereas no NF-200 was detected in somata of ADFweak
neurons, ADFstrong SGNs showed expression of the Type IISGN marker NF-200. In concert with the ADF expression inType II SGNs, we found discrete ADF-positive puncta inclose proximity to the OHCs basal pole, whereas no suchsignals were observed at IHCs. These puncta appearedafter hearing onset and mostly colocalized tosynaptophysin-positive presynaptic structures of MOCprojections that innervate OHCs (Raphael and Altschuler,2003). Deflections of stereocilia that cause transductioncurrents induce a dual response in OHCs. On the one hand,OHCs display a prestin-based rapid change in length andstiffness that provides a region-specific amplification inthe movement of the basilar membrane relative to thetectorial membrane (Zheng et al., 2000; Oliver et al., 2001)and, consequently, an increased sensitivity and specificityin coding auditory information (Raphael and Altschuler,2003). On the other hand, OHCs innervate Type II SGNsprojecting to the cochlear nucleus whose neurons, in turn,project to the superior olivary complex, a pathway thatmodulates MOC efferents (Raphael and Altschuler, 2003).Inputs from MOC efferents hyperpolarize OHCs, which rep-resents an effective mechanism of regulating OHC motilityand their amplifier activity in response to strong acousticstimulation (Maison and Liberman, 2000). We found sub-stantial ADF expression in the somata of Type II SGNs andat presynaptic structures of MOC/OHC synapses, whichwere identified by synaptophysin immunoreactivity. Thisresult implies a functional role of ADF in modulating thestrength of the MOC feedback loop and, therefore, OHCs’activity. While the specific function of ADF in the somata ofType II SGNs remains elusive, the localization in MOC/OHC synapses suggests that ADF may be relevant for syn-aptic transmission. Interestingly, functional inactivation ofthe LIM kinase-1, a kinase that effectively inactivates ADF,results in impaired synaptic function of hippocampal CA1synapses, including defects in neurotransmitter release(Meng et al., 2002). Therefore, it is tempting to speculatethat ADF is relevant for synaptic transmission of MOC/OHC synapses.
Taken together, we found ADF in various cell types ofthe mammalian inner ear sensory epithelia, including atransient expression in immature hair cells of the organ ofCorti, the utricle, and the saccule. Moreover, we found ADFin supporting cells of the fully developed cochlea and ves-tibular sensory structures as well as in Type II SGNs. Inaddition, we localized ADF to MOC/OHC synapses afterhearing onset. Based on the broad ADF expression de-scribed in the present study, we assume a relevance ofADF for inner ear development and in various aspects ofauditory and vestibular function. Our study implicates theADF gene as a candidate locus for human mutations result-ing in cochlear or vestibular dysfunction. Future analysis ofADF knockout mice will help to clarify the physiologicalrelevance of this actin-binding protein within the inner ear.
ACKNOWLEDGMENTSWe thank K. Ociepka for excellent technical assistance
and Thomas Schulenborg for analyzing the synaptophysinblocking peptide. We thank Dr. Marlies Knipper and Dr.Karen P. Steel for generously training MKH in generatinginner ear cross-sections and whole-mounts. We also thankDr. Christine B. Gurniak and Dr. Walter Witke for providingbrain lysates and inner ear preparations of ADF/ mice.
LITERATURE CITEDAiraksinen MS, Eilers J, Garaschuk O, Thoenen H, Konnerth A,
Meyer M. 1997. Ataxia and altered dendritic calcium signalingin mice carrying a targeted null mutation of the calbindinD28k gene. Proc Natl Acad Sci U S A 94:1488–1493.
Bamburg JR, Wiggan OP. 2002. ADF/cofilin and actin dynamics indisease. Trends Cell Biol 12:598–605.
Bellenchi GC, Gurniak CB, Perlas E, Middei S, Ammassari-TeuleM, Witke W. 2007. N-cofilin is associated with neuronal mi-gration disorders and cell cycle control in the cerebral cortex.Genes Dev 21:2347–2357.
Berglund AM, Ryugo DK. 1991. Neurofilament antibodies andspiral ganglion neurons of the mammalian cochlea. J CompNeurol 306:393–408.
Blair A, Tomlinson A, Pham H, Gunsalus KC, Goldberg ML, LaskiFA. 2006. Twinstar, the Drosophila homolog of cofilin/ADF, isrequired for planar cell polarity patterning. Development 133:1789–1797.
Boettger T, Hubner CA, Maier H, Rust MB, Beck FX, Jentsch TJ.2002. Deafness and renal tubular acidosis in mice lacking theK-Cl co-transporter Kcc4. Nature 416:874–878.
Boettger T, Rust MB, Maier H, Seidenbecher T, Schweizer M,Keating DJ, Faulhaber J, Ehmke H, Pfeffer C, Scheel O, LemckeB, Horst J, Leuwer R, Pape HC, Volkl H, Hubner CA, Jentsch TJ.2003. Loss of K-Cl co-transporter KCC3 causes deafness,neurodegeneration and reduced seizure threshold. EMBO J22:5422–5434.
Bruce LL, Kingsley J, Nichols DH, Fritzsch B. 1997. The develop-ment of vestibulocochlear efferents and cochlear afferents inmice. Int J Dev Neurosci 15:671–692.
Carlier MF, Laurent V, Santolini J, Melki R, Didry D, Xia GX, HongY, Chua NH, Pantaloni D. 1997. Actin depolymerizing factor(ADF/cofilin) enhances the rate of filament turnover: implica-tion in actin-based motility. J Cell Biol 136:1307–1322.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1739

Celio MR, Baier W, Scharer L, Gregersen HJ, de Viragh PA, Nor-man AW. 1990. Monoclonal antibodies directed against thecalcium binding protein calbindin D-28k. Cell Calcium 11:599–602.
Condeelis J. 2001. How is actin polymerization nucleated in vivo?Trends Cell Biol 11:288–293.
Dechesne CJ, Thomasset M. 1988. Calbindin (CaBP 28 kDa) ap-pearance and distribution during development of the mouseinner ear. Brain Res 468:233–242.
Drenckhahn D, Engel K, Hofer D, Merte C, Tilney L, Tilney M.1991. Three different actin filament assemblies occur in everyhair cell: each contains a specific actin crosslinking protein.J Cell Biol 112:641–651.
Dulon D, Blanchet C, Laffon E. 1994. Photo-released intracellularCa2 evokes reversible mechanical responses in supportingcells of the guinea-pig organ of Corti. Biochem Biophys ResCommun 201:1263–1269.
Ehret G. 1976. Development of absolute auditory thresholds inthe house mouse (Mus musculus). J Am Audiol Soc 1:179–184.
Engel J, Braig C, Ruttiger L, Kuhn S, Zimmermann U, Blin N, Saus-bier M, Kalbacher H, Munkner S, Rohbock K, Ruth P, Winter H,Knipper M. 2006. Two classes of outer hair cells along thetonotopic axis of the cochlea. Neuroscience 143:837–849.
Eybalin M, Renard N, Aure F, Safieddine S. 2002. Cysteine-stringprotein in inner hair cells of the organ of Corti: synaptic ex-pression and upregulation at the onset of hearing. Eur J Neu-rosci 15:1409–1420.
Flock A, Flock B, Fridberger A, Scarfone E, Ulfendahl M. 1999.Supporting cells contribute to control of hearing sensitivity.J Neurosci 19:4498–4507.
Gurniak CB, Perlas E, Witke W. 2005. The actin depolymerizingfactor n-cofilin is essential for neural tube morphogenesis andneural crest cell migration. Dev Biol 278:231–241.
Hafidi A. 1998. Peripherin-like immunoreactivity in type II spiralganglion cell body and projections. Brain Res 805:181–190.
Hawkins M, Pope B, Maciver SK, Weeds AG. 1993. Human actindepolymerizing factor mediates a pH-sensitive destruction ofactin filaments. Biochemistry 32:9985–9993.
Hayden SM, Miller PS, Brauweiler A, Bamburg JR. 1993. Analysisof the interactions of actin depolymerizing factor with G- andF-actin. Biochemistry 32:9994–10004.
Heizmann CW, Celio MR. 1987. Immunolocalization of parvalbu-min. Methods Enzymol 139:552–570.
Hume CR, Bratt DL, Oesterle EC. 2007. Expression of LHX3 andSOX2 during mouse inner ear development. Gene Expr Pat-terns 7:798–807.
Ikeda S, Cunningham LA, Boggess D, Hawes N, Hobson CD,Sundberg JP, Naggert JK, Smith RS, Nishina PM. 2003. Aber-rant actin cytoskeleton leads to accelerated proliferation ofcorneal epithelial cells in mice deficient for destrin (actin de-polymerizing factor). Hum Mol Genet 12:1029–1037.
Kaltenbach JA, Falzarano PR, Simpson TH. 1994. Postnatal devel-opment of the hamster cochlea. II. Growth and differentiationof stereocilia bundles. J Comp Neurol 350:187–198.
Kelly M, Chen P. 2007. Shaping the mammalian auditory sensoryorgan by the planar cell polarity pathway. Int J Dev Biol 51:535–547.
Kikuchi T, Takasaka T, Tonosaki A, Watanabe H. 1989. Fine struc-ture of guinea pig vestibular kinocilium. Acta Otolaryngol 108:26–30.
Leclere PG, Norman E, Groutsi F, Coffin R, Mayer U, Pizzey J,Tonge D. 2007. Impaired axonal regeneration by isolectin B4-binding dorsal root ganglion neurons in vitro. J Neurosci 27:1190–1199.
Leitch B, Shevtsova O, Kerr JR. 2009. Selective reduction in syn-aptic proteins involved in vesicle docking and signalling at
synapses in the ataxic mutant mouse stargazer. J Comp Neu-rol 512:52–73.
Leshchyns’ka I, Sytnyk V, Richter M, Andreyeva A, Puchkov D,Schachner M. 2006. The adhesion molecule CHL1 regulatesuncoating of clathrin-coated synaptic vesicles. Neuron 52:1011–1025.
Lewis J, Davies A. 2002. Planar cell polarity in the inner ear: howdo hair cells acquire their oriented structure? J Neurobiol 53:190–201.
Li N, Timofeyev V, Tuteja D, Xu D, Lu L, Zhang Q, Zhang Z, Singa-puri A, Albert TR, Rajagopal AV, Bond CT, Periasamy M, Adel-man J, Chiamvimonvat N. 2009. Ablation of a Ca2-activatedK channel (SK2 channel) results in action potential prolon-gation in atrial myocytes and atrial fibrillation. J Physiol 587:1087–1100.
Liberman MC, Dodds LW, Pierce S. 1990. Afferent and efferentinnervation of the cat cochlea: quantitative analysis with lightand electron microscopy. J Comp Neurol 301:443–460.
Maas C, Tagnaouti N, Loebrich S, Behrend B, Lappe-Siefke C,Kneussel M. 2006. Neuronal cotransport of glycine receptorand the scaffold protein gephyrin. J Cell Biol 172:441–451.
Maison SF, Liberman MC. 2000. Predicting vulnerability to acous-tic injury with a noninvasive assay of olivocochlear reflexstrength. J Neurosci 20:4701–4707.
Manor U, Kachar B. 2008. Dynamic length regulation of sensorystereocilia. Semin Cell Dev Biol 19:502–510.
Mansour H, Bignami A, Labkovsky B, Dahl D. 1989. Neurofila-ment phosphorylation in neuronal perikarya following axo-tomy: a study of rat spinal cord with ventral and dorsal roottransection. J Comp Neurol 283:481–485.
McGough A, Pope B, Chiu W, Weeds A. 1997. Cofilin changes thetwist of F-actin: implications for actin filament dynamics andcellular function. J Cell Biol 138:771–781.
Meng Y, Zhang Y, Tregoubov V, Janus C, Cruz L, Jackson M, Lu WY,MacDonald JF, Wang JY, Falls DL, Jia Z. 2002. Abnormal spinemorphology and enhanced LTP in LIMK-1 knockout mice.Neuron 35:121–133.
Mikaelian D, Alford BR, Ruben RJ. 1965. Cochlear potentials and8 nerve action potentials in normal and genetically deaf mice.Ann Otol Rhinol Laryngol 74:146–157.
Miyoshi J, Takai Y. 2008. Structural and functional associations ofapical junctions with cytoskeleton. Biochim Biophys Acta1778:670–691.
Oesterle EC, Campbell S, Taylor RR, Forge A, Hume CR. 2008.Sox2 and JAGGED1 expression in normal and drug-damagedadult mouse inner ear. J Assoc Res Otolaryngol 9:65–89.
Oh SH, Adler HJ, Raphael Y, Lomax MI. 2002. WDR1 colocalizeswith ADF and actin in the normal and noise-damaged chickcochlea. J Comp Neurol 448:399–409.
Oliver D, Klocker N, Schuck J, Baukrowitz T, Ruppersberg JP,Fakler B. 2000. Gating of Ca2-activated K channels con-trols fast inhibitory synaptic transmission at auditory outerhair cells. Neuron 26:595–601.
Oliver D, He DZ, Klocker N, Ludwig J, Schulte U, Waldegger S,Ruppersberg JP, Dallos P, Fakler B. 2001. Intracellular anionsas the voltage sensor of prestin, the outer hair cell motorprotein. Science 292:2340–2343.
Pack AK, Slepecky NB. 1995. Cytoskeletal and calcium-bindingproteins in the mammalian organ of Corti: cell type-specificproteins displaying longitudinal and radial gradients. Hear Res91:119–135.
Paysan J, Conroy WG, Coggan JS, Berg DK. 2000. The neurofila-ment infrastructure of a developing presynaptic calyx. J CompNeurol 425:284–294.
Peichl L, Gonzalez-Soriano J. 1993. Unexpected presence of neu-rofilaments in axon-bearing horizontal cells of the mammalianretina. J Neurosci 13:4091–4100.
Herde et al. ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
1740 The Journal of Comparative Neurology � Research in Systems Neuroscience

Rabie A, Thomasset M, Legrand C. 1983. Immunocytochemicaldetection of calcium-binding protein in the cochlear and ves-tibular hair cells of the rat. Cell Tissue Res 232:691–696.
Raphael Y, Altschuler RA. 2003. Structure and innervation of thecochlea. Brain Res Bull 60:397–422.
Ruttiger L, Sausbier M, Zimmermann U, Winter H, Braig C, EngelJ, Knirsch M, Arntz C, Langer P, Hirt B, Muller M, Kopschall I,Pfister M, Munkner S, Rohbock K, Pfaff I, Rusch A, Ruth P,Knipper M. 2004. Deletion of the Ca2-activated potassium(BK) alpha-subunit but not the BKbeta1-subunit leads to pro-gressive hearing loss. Proc Natl Acad Sci U S A 101:12922–12927.
Rymar VV, Sadikot AF. 2007. Laminar fate of cortical GABAergicinterneurons is dependent on both birthdate and phenotype.J Comp Neurol 501:369–380.
Rzadzinska AK, Schneider ME, Davies C, Riordan GP, Kachar B.2004. An actin molecular treadmill and myosins maintainstereocilia functional architecture and self-renewal. J Cell Biol164:887–897.
Schneider ME, Belyantseva IA, Azevedo RB, Kachar B. 2002.Rapid renewal of auditory hair bundles. Nature 418:837–838.
Schwartz AM, Parakkal M, Gulley RL. 1983. Postnatal develop-ment of spiral ganglion cells in the rat. Am J Anat 167:33–41.
Sytnyk V, Leshchyns’ka I, Nikonenko AG, Schachner M. 2006.NCAM promotes assembly and activity-dependent remodel-
ing of the postsynaptic signaling complex. J Cell Biol 174:1071–1085.
Von Kriegstein K, Schmitz F, Link E, Sudhof TC. 1999. Distributionof synaptic vesicle proteins in the mammalian retina identifiesobligatory and facultative components of ribbon synapses.Eur J Neurosci 11:1335–1348.
Waguespack J, Salles FT, Kachar B, Ricci AJ. 2007. Stepwisemorphological and functional maturation of mechanotrans-duction in rat outer hair cells. J Neurosci 27:13890 –13902.
Wassle H, Peichl L, Airaksinen MS, Meyer M. 1998. Calcium-binding proteins in the retina of a calbindin-null mutantmouse. Cell Tissue Res 292:211–218.
Winter H, Braig C, Zimmermann U, Engel J, Rohbock K, Knipper M.2007. Thyroid hormone receptor alpha1 is a critical regulatorfor the expression of ion channels during final differentiationof outer hair cells. Histochem Cell Biol 128:65–75.
Xiang M, Gao WQ, Hasson T, Shin JJ. 1998. Requirement forBrn-3c in maturation and survival, but not in fate determina-tion of inner ear hair cells. Development 125:3935–3946.
Zallen JA. 2007. Planar polarity and tissue morphogenesis. Cell129:1051–1063.
Zheng J, Shen W, He DZ, Long KB, Madison LD, Dallos P. 2000.Prestin is the motor protein of cochlear outer hair cells. Na-ture 405:149–155.
-------------------------------------------------------------------------------------------------------------------------------------------------- ADF development in the inner ear
The Journal of Comparative Neurology � Research in Systems Neuroscience 1741
![Arabidopsis ACTIN-DEPOLYMERIZING FACTOR3 Is Required for … · Arabidopsis ACTIN-DEPOLYMERIZING FACTOR3 Is Required for Controlling Aphid Feeding from the Phloem1[OPEN] Hossain A](https://img.pdfslide.us/doc/110x75/5f7dd59be0afd940a23b8977/arabidopsis-actin-depolymerizing-factor3-is-required-for-arabidopsis-actin-depolymerizing.jpg)






![CYTOSKELETON NEWS - fnkprddata.blob.core.windows.net · Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)]](https://img.pdfslide.us/doc/110x75/609edd2b88630103265d18ee/cytoskeleton-news-dynamic-remodeling-of-the-actin-cytoskeleton-ie-rapid-cycling.jpg)










![Review Actin-targeting natural products: structures ... · actin-binding proteins actively break or ‘sever’ actin filaments [e.g. actin-depolymerizing factor (ADF) and cofilin]](https://img.pdfslide.us/doc/110x75/5f0f85bd7e708231d44494d0/review-actin-targeting-natural-products-structures-actin-binding-proteins-actively.jpg)







![[ A ] SPIRITS ADF [ADF] VODKA - BASIC](https://img.pdfslide.us/doc/110x75/6169d8c211a7b741a34c063e/-a-spirits-adf-adf-vodka-basic.jpg)


