Embed Size (px)
Citation preview

International Journal of Food Microbiology 126 (2008) 180–185
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Development of biocontrol agents from food microbial isolates for controllingpost-harvest peach brown rot caused by Monilinia fructicola
Ting Zhou a,⁎, Karin E. Schneider b, Xiu-Zhen Li a
a Guelph Food Research Centre, Agriculture and Agri-Food Canada, 93 Stone Road West, Guelph, Ontario, Canada N1G 5C9b Southern Crop Protection and Food Research Centre, 4902 Victoria Avenue, Vineland Station, Ontario, Canada L0R 2E0
⁎ Corresponding author. Tel.: +1 519 780 8036; fax: +E-mail address: [email protected] (T. Zhou).
0168-1605/$ – see front matter © 2008 Published by Edoi:10.1016/j.ijfoodmicro.2008.05.020
A B S T R A C T
A R T I C L E I N F OArticle history:
An unconventional strategy Received 24 April 2007Received in revised form 16 May 2008Accepted 16 May 2008Keywords:BacillusBiological controlBrown rotFood microorganismPrunus persicaStone fruit
of screening food microbes for biocontrol activity was used to develop biocontrolagents for controlling post-harvest peach brown rot caused by Monilinia fructicola. Forty-four microbialisolates were first screened for their biocontrol activity on apple fruit. Compared with the pathogen-onlycheck, seven of the 44 isolates reduced brown rot incidence by N50%, including four bacteria: Bacillus sp. C06,Lactobacillus sp. C03-b and Bacillus sp. T03-c, Lactobacillus sp. P02 and three yeasts: Saccharomyces delbrueckiiA50, S. cerevisiae YE-5 and S. cerevisiae A41. Eight microbial isolates were selected for testing on peaches bywound co-inoculation with mixtures of individual microbial cultures and conidial suspension of M. fructicola.Only two of them showed significant biocontrol activity after five days of incubation at 22 °C. Bacillus sp. C06suppressed brown rot incidence by 92% and reduced lesion diameter by 88% compared to the pathogen-onlycheck. Bacillus sp.T03-c reduced incidence and lesion diameter by 40% and 62%, respectively. The two isolateswere compared with Pseudomonas syringae MA-4, a biocontrol agent for post-harvest peach diseases, byimmersing peaches in an aliquot containing individual microbial isolates and the pathogen conidia.Treatments with isolates MA-4, C06 and T03-c significantly controlled brown rot by 91, 100, and 100%respectively. However, only isolates MA-4 and C06 significantly reduced brown rot by 80% and 15%,respectively when bacterial cells alone were applied. On naturally infected peaches, both the bacterial cultureand its cell-free filtrate of the isolate C06 significantly controlled peach decay resulting in 77 and 90%reduction, respectively, whereas the treatment using only the bacterial cells generally had no effect. IsolateC06 is a single colony isolate obtained from a mesophilic cheese starter, and has been identified belonging toBacillus amyloliquefaciens. The results have clearly demonstrated that isolate C06 has a great potential forbeing developed into a biocontrol agent.
© 2008 Published by Elsevier B.V.
1. Introduction
Brown rot caused by Monilinia fructicola (Wint.) Honey is a majordisease on stone fruit including apricots, cherries and peaches inNorth America. Although the disease may affect blossoms and twigs, itis highly destructive on fruit, and can ruin half or more of the cropbefore harvest, with the remaining fruit subject to post-harvest decay.In a two-year survey conducted in Southern Ontario, 20 to 80% ofcommercially ripe peaches harvested from local orchards developedbrown rot decay after only 4–5 days incubation at room temperature(Zhou and Sholberg, 2002). Both the industry and consumers urgentlydemand effective strategies for prolonging shelf life of peaches.
In the past two decades, substantial efforts have been made onfinding effective biocontrol agents for controlling brown rot of stonefruit from plant epiphytic or endophytic microorganisms. Bacillussubtilis, from apple rhizosphere, which produced an antibioticsubstance toxic to M. fructicola (Pusey and Wilson, 1984), performed
1 519 829 2600.
lsevier B.V.
as effectively as fungicides in controlling peach brown rot in the pilottests (Pusey et al., 1988). Certain isolates of Pseudomonas syringaeisolated from apple leaves were effective in suppressing both brownrot and rhizopus rot in peaches (Zhou et al., 1999). Pantoeaagglomerans EFS125 isolated from the surface of a pear fruit controlledstone fruit decay effectively as did the fungicides (Bonaterra et al.,2003). Antagonistic yeasts have also been investigated as biocontrolagents for brown rot of stone fruit. Fifty-one endophytic isolates ofAureobasidium pullulans were obtained from the flesh of sweetcherries and extensively evaluated for their biocontrol activity againstpost-harvest rots of sweet cherries; one of the isolates (533) reducedfruit decay by up to 80% (Schena et al., 2003). Among a total of 586natural yeasts isolated from grape berries, two strains of S. cerevisiaeand one of Zygosaccharomyces showed a broad spectrum of antag-onistic activity against 10 fungal pathogens in in vitro tests (Suzziet al., 1995). A total of 103 yeasts isolated from peach fruit woundswere screened for biocontrol activity against M. fructicola and one ofthe best isolates DR52, identified as Kloeckera apiculata, showedsignificant reduction of brown rot on wound-inoculated peaches(Karabulut and Baykal, 2003).

181T. Zhou et al. / International Journal of Food Microbiology 126 (2008) 180–185
Whether or not biocontrol products will be widely used willdepend not only on their performance, but also on their acceptance bythe consumers as safe alternatives to chemical fungicides (Chalutz andDroby, 1998). Certain microorganisms, commonly called “foodmicrobes”, including bacteria, yeasts and filamentous fungi, havehistorically been used to produce special foods and beverages such asbread, cider, beer, wine etc. and as natural preservatives forfermenting fruit, vegetables, meat and dairy products. These beneficialmicrobes are generally considered safe and have long been acceptedby consumers. Many food microbes possess the capability ofproducing antimicrobials, such as bacteriocins and substances withantifungal activities, which may have a great potential to be used incontrolling fresh fruit decays (Davidson and Hoover, 1993; Rodgers,2001). Certain strains of lactic acid bacteria were active against Peni-cillium expansum, a pathogen of apple blue mold (Florianowicz, 2001).The screening of 150 industrial yeasts for control of Penicillium rot inlemons has resulted in seven with antagonistic activity toward Peni-cillium digitatum; among them, one Saccharomyces cerevisiae andthree Kluyveromyces isolates provided complete control of Penicilliumrot in lemons (Cheah and Tran,1995). However, it is unknownwhetherthose microbial isolates may be antagonistic toward M. fructicola. Inaddition to lactic acid bacteria and yeasts, species of Bacillus are alsocommonly used in food processing and are likely to have anti-microbial activity (Han et al., 2001; Kiers et al., 2000). The objectivesof this research were to evaluate various food microbial isolates fortheir antifungal activity against M. fructicola and to determine theefficacy and potential applications of the selected food microbialisolates in controlling post-harvest decay of peach fruit.
2. Materials and methods
2.1. Fungal pathogen
M. fructicola was obtained from peach (Prunus persica (L.) Batsch)fruit with typical brown rot symptoms. The single-spore isolates werecultured on Potato Dextrose Agar (PDA) slants and stored at 4 °C.Conidia of the pathogen were produced on V-8 agar (Dhingra andSinclair, 1995) at 22–24 °C for 7–10 days.
2.2. Food microbial isolates
Food microbial isolates evaluated in this research (Table 1) wereeither purchased from different commercial retailers as culturestarters for wine, beer, cheese and yogurt making, or isolated in thelaboratory from food products bought from local food stores. Beforeuse in the experiments, the microbial isolates were cultured ineither Potato Dextrose Broth (PDB) or Tryptic Soy Broth (TSB) at22–24 °C on a rotary shaker at 150 rpm for 3 days. When requested,freshly grown cultures of the microbial isolates were centrifuged toseparate the cells from the supernatant, and the latter was passedthrough an ultra-filter (0.2 µm) resulting in the cell-free filtrate.The cell pellets were re-suspended in 0.5% CaCl2 and stored at 4 °Cuntil use.
2.3. Evaluation of food microbial isolates on apples
All food microbial isolates were evaluated for their efficacy insuppressing the brown rot pathogen, M. fructicola on apples (Malusdomestica Borkh.) ‘Jonagold’ or ‘Golden Delicious’ apples wereobtained from a commercial cold storage facility in Simcoe, Ontario,where the apples were stored under controlled atmosphereconditions. Apples were soaked in 10% household bleach (5%sodium hypochlorite) and 0.01% Tween 20 (Fisher Scientific) for4 min and rinsed in reverse osmosis water for 4 min. Once thesurface was dry, the disinfested apples were wounded four timeswith a sterile nail-like pointer (5 mm diameter and 5 mm in depth)
and then placed on a plastic packing tray contained in a plastic box.Suspensions of the individual microbial isolates were mixed withequal amounts of the conidial suspension of M. fructicola. The finalmixture consisted of 1×107 CFU ml−1 of a microbial isolate and1×104 pathogen conidia ml−1, estimated based on optical density(OD) reading and determined by using a hemacytometer, respec-tively. A drop of 20 µL of the mixture was applied to each wound.The experiment was arranged as a completely randomized designwith three replications, and five apples (i.e. 20 wounds) weretreated for each microbial isolate in each replicate. Once treated,apples were incubated in an environmental chamber at 22–24 °Cwith 85% relative humidity. Disease incidence (% wounds withbrown rot lesions) and severity (lesion size) were assessed after5 days post-treatment by measuring diameters of rotten area usinga caliper.
2.4. Co-inoculation of peaches with pathogen and food microbial isolates
Peaches of cultivar ‘Redhaven’ (P. persica (L.) Batsch) wereharvested when commercially ripe from research orchards at JordanStation, Ontario. The harvested fruit were stored at 4 °C prior totreatment. The orchards did not receive any preharvest fungicideapplication.
A method similar to the evaluation on apples was used todetermine efficacy of selected microbial isolates in suppressingbrown rot on peaches. Harvested peaches were placed in a plasticpacking insert (20 fruit master) contained in a wooden tray(45×32×9 cm), and 12 peaches per tray. Each fruit was woundedtwice and inoculated within 5 min with 20 μL of inoculant containingeither a conidial suspension of M. fructicola or the pathogensuspension mixed with individual microbial isolates. The finalconcentrations were 1×104 conidia ml−1 and 1×108 CFU ml−1 forthe pathogen and microbial isolates, respectively. Each treatment wasapplied to peaches in four trays (replicates). The treated peaches wereincubated in a growth room at 22 °C with 85% RH for 5 days. Theexperiment was repeated once.
2.5. Immersion treatments of artificially inoculated peaches
‘Harrow Beauty’ peaches were also harvested commercially ripefrom research orchards at Jordan Station, Ontario. The peaches weresoaked for 2 min in different suspensions (treatments) all containing1×105 conidia ml−1 of M. fructicola. The treatments included eitherwhole culture or cells only of individual microbial isolates (Fig. 1) atthe concentration of 1×108 CFU ml−1. A solution of 0.5% CaCl2 was setas the control for the cells only treatments since the obtained cellswere re-suspended in the solution. Fifteen peaches were treated foreach treatment within a replicate and there were four replications.After removal from the treatment suspension the peaches wereallowed to be air-dried for 5 min, prior to being placed in the plasticpacking inserts in the wooden tray described above, and incubated at22 °C with 85% RH for 5 days.
2.6. Immersion treatments of naturally infected peaches
Commercially unblemished ‘Redhaven’ peaches were harvestedfrom an orchard with severe brown rot at Jordan Station, Ontario.The peaches were treated similarly to the inoculated experimentdescribed above. The treatments included whole culture, cells only,and cell-free filtrate of individual microorganisms (Fig. 2). Water,0.5% CaCl2, and PDB (medium used for culturing the microorgan-isms) were set as controls corresponding to the differenttreatments. The experiment also used 15 peaches for eachtreatment within a replicate and four replications. The treatedpeaches were arranged and incubated in the same manner asdescribed above.

Table 1List of food microbial isolates evaluated for antifungal activity against Monilinia fructicola
Microbial isolates Product Origin Source
BacteriaC03-b Lactobacillus sp. Ricotta cheese/single colony isolation Tre Stelle, Montreal, Canada Food Store, OntarioC06 Mesophilic bacteria Cheese starter/single colony isolation University of Guelph, ON, CanadaM3200 Pedicococcus cerevisiae Lambic beer culture Belgium The Yeast Culture Kit Company, Madison, MI, USAP01 Lactobacillus sp. Pickled vegetables/single colony isolation Home made, Ontario, CanadaP02 Lactobacillus sp. Kimchi–cabbage/single colony isolation Korea Food Store, OntarioT01-a Bacillus sp. Preserved Beancurd (Furu)/single colony isolation Panyu, China Food Store, OntarioT01-b Bacillus sp. Preserved Beancurd (Furu)/single colony isolation Panyu, ChinaT02 Bacillus sp. Preserved Beancurd (Furu)/single colony isolation TaiwanT03-c Bacillus sp. Preserved Beancurd (Furu)/single colony isolation GuangzhouT04-b Bacillus sp. Pickled bean cake/single colony isolation TaiwanT05-a Bacillus sp. Tofu cake/single colony isolation Taiwan
YeastsA01 Saccharomyces cerevisiae American Ale Beer yeast culture California The Yeast Culture Kit Company, Madison, MI, USAA04 S. cerevisiae German Ale Beer yeast culture GermanA06 S. cerevisiae Stout Ale Beer yeast culture IrelandA08 S. cerevisiae English Barley wine Beer yeast culture DorchesterA13 S. cerevisiae Irish Ale Beer yeast culture IrelandA15 S. cerevisiae English Ale Beer yeast culture EnglandA34 S. cerevisiae Scotch Ale Beer yeast culture EdinburghA35 S. cerevisiae Belgian Wit Beer yeast culture BelgiumA37 Saccharomyces sp. Altbier Beer yeast culture BavariaA41 S. cerevisiae Wit Beer yeast culture BelgiumA42 S. cerevisiae Tadcaster Beer yeast culture BelgiumA50 Saccharomyces delbrueckii Weizen Beer yeast culture BavariaA64 S. cerevisiae Faversham Beer yeast culture FavershamL02 Saccharomyces uvarum Bavarian Lager Beer yeast culture BavarianL09 S. uvarum German Lager Beer yeast culture BavariaL17 S. uvarum Pilsen Lager Beer yeast culture PilzenM3220 Brettanomyces lambicus Lambic beer culture BelgiumS104 S. cerevisiae Tekna (Reisling) Wine yeast culture Home Brewer's Yeast Service, Saskatoon, CanadaS105 S. cerevisiae Red Star Wine yeast cultureS201 S. cerevisiae British Ale Beer yeast cultureS203 S. cerevisiae Irish Ale Beer yeast cultureS204 S. cerevisiae London Ale Beer yeast cultureS205 S. cerevisiae Nottingham Ale Beer yeast cultureS206 S. cerevisiae Fleur de Lys Ale Beer yeast cultureS302 S. uvarum Tekna (German Lager II) Beer yeast cultureS303 S. uvarum Steam Lager Beer yeast cultureS501 Saccharomyces sp. Greenwood Cider Hard cider yeast Ontario apple ciderW01-a S. cerevisiae Rice wine starter Shanghai Food Store, OntarioW06 S. cerevisiae Champagne Wine yeast culture Montreal The Yeast Culture Kit CompanyW22 S. cerevisiae Aromaic Mead Wine yeast culture FranceYE-2 S. cerevisiae 71B-1122 Lalvinu yeast culture Narbonne Lallemand Inc., Ontario, CanadaYE-3 S. cerevisiae Bourgovin RC 212- Lalvin yeast culture BurgundyYE-4 S. cerevisiae Souche ICV D47 Lalvin yeast culture Côte de RhôneYE-5 S. cerevisiae K1-V1116 Lalvin yeast culture MontpellierYE-6 Saccharomyces bayanus EC1118 Lalvin yeast cultureYE-7 S. cerevisiae Coopers Brewery Coopers Brewery — Pure Brewer's Yeast
182 T. Zhou et al. / International Journal of Food Microbiology 126 (2008) 180–185
2.7. Taxonomy identification of isolate C06
The bacterial isolate was identified based on two tests: BIOLOGmicrobial identification system (http://BIOLOG.com) and Microbial IDdetection using 16 S rRNA gene sequencing method. All tests wereconducted according to the manufacture's instructions by LaboratoryServices, University of Guelph, Guelph, Ontario, Canada.
2.8. Data analysis
All data were analyzed using Unistat 5.5 software (Unistat Ltd.London, UK). General Linear Model (GLM) procedures were used forthe analysis of variance. Data from experimental repetitions weretested for homogeneity of variance (Pb0.05) prior to pooling.Percentage data with a range greater than 40% were subjected toarcsine-square-root transformation before the analysis of variance.Differences among treatments were determined by Duncan's MultipleRange test. Percent disease reductions relative to the corresponding
controls were calculated from disease incidence or lesion diametersaccording to: 100×(C−T)/C, where C=pathogen-only check or CaCl2check; T=individual treatment.
3. Results
3.1. Biocontrol activity of food microbial isolates evaluated on apples
Comparedwith the pathogen-only check, seven of the 44microbialisolates tested against M. fructicola on apples reduced diseaseincidence (percentage of wounds developing brown rot lesions)by more than 50% (Table 2). Of them, four are bacteria (Bacillussp. C06, Lactobacillus sp. C03-b Bacillus sp. T03-c, and Lactobacillussp. P02) and three are yeasts (Saccharomyces delbrueckii A50, S.cerevisiae YE-5 and S. cerevisiae A41). The best isolates were P02and A50, which reduced disease incidence by over 80%. IsolatesP02, A50 and C06 were only three of the 44 isolates reducing lesiondiameter by over 50% compared to the check, and their reduction
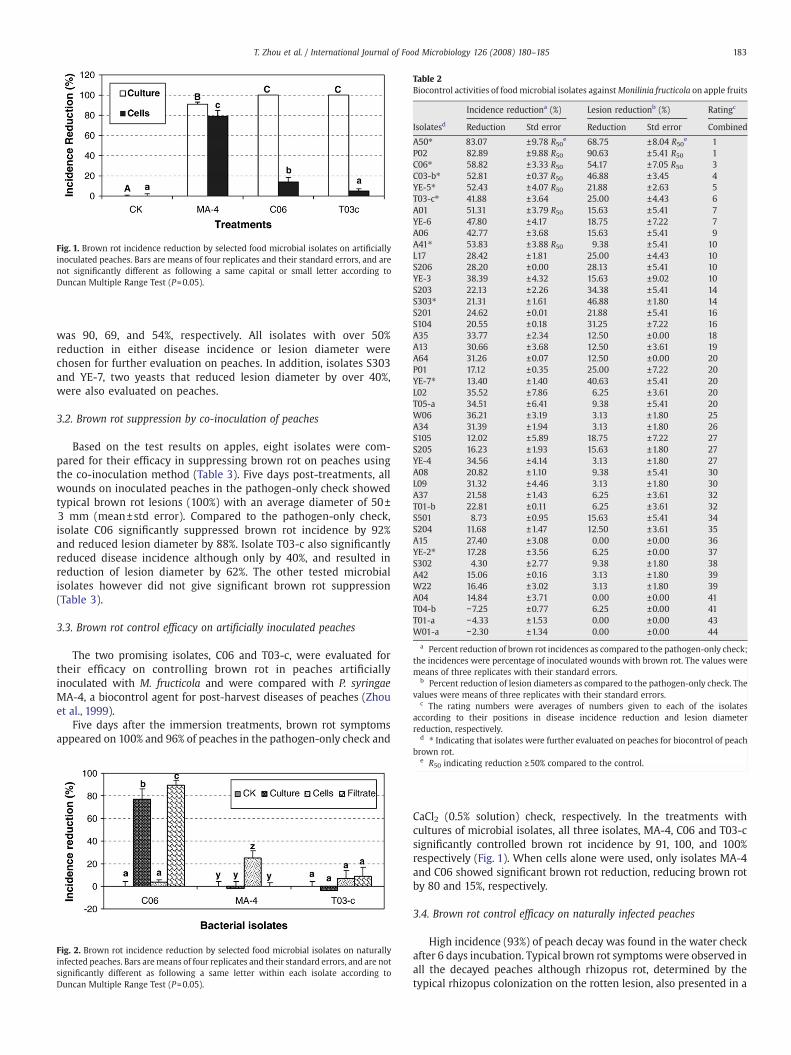
Table 2Biocontrol activities of foodmicrobial isolates againstMonilinia fructicola on apple fruits
Incidence reductiona (%) Lesion reductionb (%) Ratingc
Isolatesd Reduction Std error Reduction Std error Combined
A50⁎ 83.07 ±9.78 R50e 68.75 ±8.04 R50
e 1P02 82.89 ±9.88 R50 90.63 ±5.41 R50 1C06⁎ 58.82 ±3.33 R50 54.17 ±7.05 R50 3C03-b⁎ 52.81 ±0.37 R50 46.88 ±3.45 4YE-5⁎ 52.43 ±4.07 R50 21.88 ±2.63 5T03-c⁎ 41.88 ±3.64 25.00 ±4.43 6A01 51.31 ±3.79 R50 15.63 ±5.41 7YE-6 47.80 ±4.17 18.75 ±7.22 7A06 42.77 ±3.68 15.63 ±5.41 9A41⁎ 53.83 ±3.88 R50 9.38 ±5.41 10L17 28.42 ±1.81 25.00 ±4.43 10S206 28.20 ±0.00 28.13 ±5.41 10YE-3 38.39 ±4.32 15.63 ±9.02 10S203 22.13 ±2.26 34.38 ±5.41 14S303⁎ 21.31 ±1.61 46.88 ±1.80 14S201 24.62 ±0.01 21.88 ±5.41 16S104 20.55 ±0.18 31.25 ±7.22 16A35 33.77 ±2.34 12.50 ±0.00 18A13 30.66 ±3.68 12.50 ±3.61 19A64 31.26 ±0.07 12.50 ±0.00 20P01 17.12 ±0.35 25.00 ±7.22 20YE-7⁎ 13.40 ±1.40 40.63 ±5.41 20L02 35.52 ±7.86 6.25 ±3.61 20T05-a 34.51 ±6.41 9.38 ±5.41 20W06 36.21 ±3.19 3.13 ±1.80 25A34 31.39 ±1.94 3.13 ±1.80 26S105 12.02 ±5.89 18.75 ±7.22 27S205 16.23 ±1.93 15.63 ±1.80 27YE-4 34.56 ±4.14 3.13 ±1.80 27A08 20.82 ±1.10 9.38 ±5.41 30L09 31.32 ±4.46 3.13 ±1.80 30A37 21.58 ±1.43 6.25 ±3.61 32T01-b 22.81 ±0.11 6.25 ±3.61 32S501 8.73 ±0.95 15.63 ±5.41 34S204 11.68 ±1.47 12.50 ±3.61 35A15 27.40 ±3.08 0.00 ±0.00 36YE-2⁎ 17.28 ±3.56 6.25 ±0.00 37S302 4.30 ±2.77 9.38 ±1.80 38A42 15.06 ±0.16 3.13 ±1.80 39W22 16.46 ±3.02 3.13 ±1.80 39A04 14.84 ±3.71 0.00 ±0.00 41T04-b −7.25 ±0.77 6.25 ±0.00 41T01-a −4.33 ±1.53 0.00 ±0.00 43W01-a −2.30 ±1.34 0.00 ±0.00 44
a Percent reduction of brown rot incidences as compared to the pathogen-only check;the incidences were percentage of inoculated wounds with brown rot. The values weremeans of three replicates with their standard errors.
b Percent reduction of lesion diameters as compared to the pathogen-only check. Thevalues were means of three replicates with their standard errors.
c The rating numbers were averages of numbers given to each of the isolatesaccording to their positions in disease incidence reduction and lesion diameterreduction, respectively.
d ⁎ Indicating that isolates were further evaluated on peaches for biocontrol of peach
Fig. 1. Brown rot incidence reduction by selected food microbial isolates on artificiallyinoculated peaches. Bars are means of four replicates and their standard errors, and arenot significantly different as following a same capital or small letter according toDuncan Multiple Range Test (P=0.05).
183T. Zhou et al. / International Journal of Food Microbiology 126 (2008) 180–185
was 90, 69, and 54%, respectively. All isolates with over 50%reduction in either disease incidence or lesion diameter werechosen for further evaluation on peaches. In addition, isolates S303and YE-7, two yeasts that reduced lesion diameter by over 40%,were also evaluated on peaches.
3.2. Brown rot suppression by co-inoculation of peaches
Based on the test results on apples, eight isolates were com-pared for their efficacy in suppressing brown rot on peaches usingthe co-inoculation method (Table 3). Five days post-treatments, allwounds on inoculated peaches in the pathogen-only check showedtypical brown rot lesions (100%) with an average diameter of 50±3 mm (mean±std error). Compared to the pathogen-only check,isolate C06 significantly suppressed brown rot incidence by 92%and reduced lesion diameter by 88%. Isolate T03-c also significantlyreduced disease incidence although only by 40%, and resulted inreduction of lesion diameter by 62%. The other tested microbialisolates however did not give significant brown rot suppression(Table 3).
3.3. Brown rot control efficacy on artificially inoculated peaches
The two promising isolates, C06 and T03-c, were evaluated fortheir efficacy on controlling brown rot in peaches artificiallyinoculated with M. fructicola and were compared with P. syringaeMA-4, a biocontrol agent for post-harvest diseases of peaches (Zhouet al., 1999).
Five days after the immersion treatments, brown rot symptomsappeared on 100% and 96% of peaches in the pathogen-only check and
brown rot.e R50 indicating reduction ≥50% compared to the control.
Fig. 2. Brown rot incidence reduction by selected food microbial isolates on naturallyinfected peaches. Bars aremeans of four replicates and their standard errors, and are notsignificantly different as following a same letter within each isolate according toDuncan Multiple Range Test (P=0.05).
CaCl2 (0.5% solution) check, respectively. In the treatments withcultures of microbial isolates, all three isolates, MA-4, C06 and T03-csignificantly controlled brown rot incidence by 91, 100, and 100%respectively (Fig. 1). When cells alone were used, only isolates MA-4and C06 showed significant brown rot reduction, reducing brown rotby 80 and 15%, respectively.
3.4. Brown rot control efficacy on naturally infected peaches
High incidence (93%) of peach decay was found in the water checkafter 6 days incubation. Typical brown rot symptomswere observed inall the decayed peaches although rhizopus rot, determined by thetypical rhizopus colonization on the rotten lesion, also presented in a

Table 3Brown rot control by selected food microorganisms on wound-inoculated peaches
Isolates Incidencea reduction (%) Lesion diameterb reduction (%)
(means±std error) (means±std error)
C06 92.42 ±5.95 A 87.64 ±7.86 aT03-C 39.55 ±10.35 B 61.96 ±4.40 bYE-7 1.85 ±1.85 C 7.97 ±2.80 cA41 0.00 ±0.00 C 2.56 ±2.82 cA50 0.00 ±0.00 C 1.86 ±2.27 cC03-b 0.00 ±0.00 C 0.78 ±2.72 cS303 0.00 ±0.00 C 1.30 ±2.04 cYE-2 0.00 ±0.00 C 6.51 ±4.39 cYE-5 0.00 ±0.00 C 2.48 ±2.02 cCheck 0.00 ±0.00 C 0.00 ±0.00 c
a Incidence was the percentage of wound with typical brown rot lesion assessed fivedays after the treatments. The incidence in the pathogen-only check was 100%. Themeans were averages of six replicates pooled from two experiments, and are notsignificantly different when followed with the same letter (Duncan Multiple Range TestP=0.05).
b Lesion diameter was the diameter of brown rot lesion around the inoculated woundmeasures five days after the treatments. The mean of lesion diameter in the pathogen-only check was 50 mm. The means were averages of six replicates pooled from twoexperiments, and are not significantly different when followed with the same letter(Duncan Multiple Range Test P=0.05).
184 T. Zhou et al. / International Journal of Food Microbiology 126 (2008) 180–185
few cases. Both the culture and cell-free filtrate of isolate C06significantly controlled peach decay resulting in 77 and 90% reduction,respectively (Fig. 2), while the cells only treatment generally had nosignificant effect on controlling peach decay. In contrast to isolate C06,the cells only treatment of isolate MA-4 controlled peach decay by25%, but the culture and cell-free filtrate treatments failed to give anysignificant control. Isolate T03-c performed very poorly on thenaturally infected peaches; none of its treatments showed significantdecay reduction as compared to the water check. Although no specifictreatments were set to evaluate if the isolates may cause anysymptom, attention was paid in this aspect. No abnormal symptomwas observed as peaches treated with the bacterial isolate werecompared with untreated ones.
3.5. Taxonomy identification of isolate C06
Isolate C06 was identified using BIOLOG as a strain of Bacillusamyloliquefaciens (prob=96; SIM=0.65); its partial 16S rRNAsequences have 99% similarity with B. amyloliquefaciens (GenBankaccession number AY651023).
4. Discussion
The research used an untraditional strategy to develop biocontrolagents from food microbial isolates. The approach of isolatingbiocontrol agents from naturally occurring microorganisms has beencommonly used in developing biocontrol agents of post-harvestdiseases with a considerable number of successes (Janisiewicz andKorsten, 2002; Zhou et al., 2007). Microorganisms isolated from thefruit surface generally are able to colonize and adapt more readily to aniche similar to where they originated, resulting in a significantcompetition against fruit pathogens (Chalutz and Droby, 1998).However, the strategy to develop biocontrol agents from foodmicrobes has certain obvious advantages. Since consumers willdirectly handle and consume fresh fruit, safety and acceptability areextremely important for a biocontrol product targeting post-harvestdecay of fruit. The safety of foodmicrobes has historically been proved,therefore, no major obstacles are anticipated by their use as biologicalcontrol agents. In addition, many food microbes have been wellstudied and clearly characterized physiologically and genetically(Davidson and Hoover, 1993; Rey et al., 2004; Schweizer, 2004) andare already mass-produced in industrial scales.
An in vivo bioassay was developed to preliminarily screen theforty-four food microbial isolates for their biocontrol activity. Since invitro testing often does not always correlate with in situ biocontrolresults (Schisler and Slininger,1997), an appropriate bioassay is criticalfor screening biocontrol agents from a relatively large number ofcandidates. Due to seasonal availability and rapid physiologicaldeterioration of peach fruit, apple fruit were used for the preliminaryscreening of the food microbes. The reasons for choosing applesincluded their susceptibility to the pathogen, long storability withrelatively stable physiology, and availability all year long. Thedistinguishing appearance of disease lesions on light-colored applefruit and their relative firm texture were other advantages of usingapples for the bioassay. In addition, apple assay has been commonlyused for testing pathogenicity of different plant pathogens (Melzerand Boland, 1999). The results of the bioassay with apple fruit showedthat more than half of the food microbial isolates (23 out of 44)reduced brown rot incidence by over 25%, seven by over 50%, and twoby over 80%, proving the bioassay was an effective tool for screeningbiocontrol agents.
Isolate C06, a single colony isolated from a mesophilic cheesestarter, significantly controlled brown rot in all three experimentsconducted on peaches. The experiments were designed to evaluateefficacy of the selected promising microbial isolates in controllingpost-harvest brown rot resulting from different types of M. fructicolainfections: wound infections in the orchards or during harvest,secondary infections during packaging and marketing, and latentinfections in the growing season (Ogawa et al., 1995). The effective-ness of isolate C06 in all experiments indicated it has great potential tobe developed as a biocontrol agent for post-harvest brown rot.Another bacterial isolate T03-c, a single colony isolate from preservedbeancurd, reduced brown rot significantly in the wound co-inocula-tion and the artificial inoculation experiments. However, it failed tocontrol brown rot caused by latent (natural) infections, which are themost important cause of brown rot (Zhou and Sholberg, 2002).Although several food yeast isolates showed biocontrol activity onapple fruit, none of the six yeast isolates tested on peaches reducedbrown rot incidence and severity. The variation in biocontrol activitybetween peaches and apples might be due to the difference inpathogen aggressiveness on different hosts; high aggressiveness of apathogen often results in low efficacy of a biocontrol agent (Francéset al., 2006). Antifungal activity against post-harvest fungal pathogensby wine yeasts has been reported elsewhere (Cheah and Tran, 1995;Suzzi et al., 1995), however, their effectiveness toward M. fructicolawas not tested. Wine yeasts may be used as biocontrol agents aftertheir improvement through gene transformation (Jones and Prusky,2002).
In this research, the promising food microbial isolates werecompared with a known biocontrol agent, P. syringae MA-4, whichwas effective in controlling post-harvest diseases of apples andpeaches (Zhou et al., 1999, 2001, 2002). In the experiment withartificially inoculated peaches, bacterial cultures of isolates C06 andT03-c controlled brown rot slightly better than isolateMA-4. However,the cells only treatments with isolates C06 and T03-c performed verypoorly by giving almost no disease reduction, while the cells onlytreatment of MA-4 controlled brown rot by 80%. In the experimentwith naturally infected peaches, although a culture of isolate C06significantly controlled brown rot by close to 80%, the cells onlytreatment generally did not provide any disease reduction. Thedifference in origins of the two isolates might explain the differentperformance of their cells only treatments. Isolate MA-4 originatedfrom apple leaves, and therefore might have been able to colonize andadapt to the peach fruit surface more effectively than cells of isolateC06, which was isolated from a cheese starter and might have haddisadvantages in surviving on the fruit surface. However, further studyis needed to determine the population dynamic of the isolate C06 inthe wound and on the surface of peach fruit.

185T. Zhou et al. / International Journal of Food Microbiology 126 (2008) 180–185
The cell-free filtrate of the isolate C06 showed strong biocontrolactivity on naturally infected peaches, whereas the cells alonetreatment was ineffective; indicating that it was the extracellularsubstance produced by isolate C06 and not the bacteria, whicheffectively reduced the fruit decay. When tested in vitro, the filtratecompletely inhibited conidial germination of M. fructicola (data notshown), implying that it might contain antifungal compound(s).However, the filtrate might also contain compound(s) with otherfunctions, such as inducing fruit resistance and inhibiting the activityof pathogenesis enzymes of the pathogen. These factors might playimportant roles in controlling peach brown rot (El Ghaouth et al.,2003).
Isolate C06 was identified as B. amyloliquefaciens. This bacterialspecies is known for its production of α-amylase and is often used incheese starter to promote flavour production in cheddar cheese. Theresults presented here have clearly demonstrated that isolate C06 hasgreat potential for being developed into a biocontrol agent althoughevaluations on a larger scale are needed to confirm its efficacy. A clearidentification and characterization of the active ingredient(s) secretedby the bacterial cells are necessary for further development of the foodmicrobial isolate into a biocontrol product, and this research iscurrently underway.
Acknowledgements
The authors would like to thank the Canada Agricultural Adapta-tion Council, Ontario Tender Fruit Producers' Marketing Board for thefinancial support, and Todd Larmet and Karen Michaud for theirtechnical assistance.
References
Bonaterra, A., Mari, M., Casalini, L., Montesinos, E., 2003. Biological control of Monilinialaxa and Rhizopus stolonifer in postharvest of stone fruit by Pantoea agglomeransEPS125 and putative mechanisms of antagonism. International Journal of FoodMicrobiology 84, 93–104.
Chalutz, E., Droby, S., 1998. Biological control of postharvest disease. In: Boland, G.J.,Kuykendall, L.D. (Eds.), Plant-Microbe Interactions and Biological Control. MarcelDekker, Inc., New York, pp. 157–170.
Cheah, L.H., Tran, T.B., 1995. Postharvest biocontrol of Penicillium rot of lemons withindustrial yeasts. In: Popay, A.J. (Ed.), Proceedings of the Forty Eighth New ZealandPlant Protection Conference, Angus Inn, Hastings, New Zealand, August 8–10, 1995,pp. 155–157.
Davidson, P.M., Hoover, D.G., 1993. Antimicrobial components from lactic acid bacteria.In: Salminen, S., von Wright, A. (Eds.), Lactic Acid Bacteria. Marcel Dekker, Inc, NewYork, pp. 127–160.
Dhingra, O.D., Sinclair, J.B., 1995. Basic Plant Pathology Methods, 2nd Edition. CRC Press,Inc., Boca Raton, FL, USA, p. 363.
El Ghaouth, A., Wilson, C.L., Wisniewski, M., 2003. Control of postharvest decay of applefruit with Candida saitoana and induction of defense responses. Phytopathology 93,344–348.
Florianowicz, T., 2001. Antifungal activity of some microorganisms against Penicilliumexpansum. European Food Research and Technology 212, 282–286.
Francés, J., Bonaterra, A., Moreno, M.C., Cabrefiga, J., Badosa, E., Montesinos, E., 2006.Pathogen aggressiveness and postharvest biocontrol efficiency in Pantoea agglom-erans. Postharvest Biology and Technology 39, 299–307.
Han, B.Z., Rombouts, F.M., Nout, M.J.R., 2001. A Chinese fermented soybean food.International Journal of Food Microbiology 65, 1–10.
Janisiewicz, W.J., Korsten, L., 2002. Biological control of postharvest diseases of fruits.Annual Review of Phytopathology 40, 411–441.
Jones, R.W., Prusky, D., 2002. Expression of an antifungal peptide in Saccharomyces: anew approach for biological control of the postharvest disease caused by Colleto-trichum coccodes. Phytopathology 92, 33–37.
Karabulut, O.A., Baykal, N., 2003. Biological control of postharvest diseases of peachesand nectarines by yeasts. Journal of Phytopathology 151, 130–134.
Kiers, J.L., Van Laeken, A.E.A., Rombouts, F.M., Nout, M.J.R., 2000. In vitro digestibility ofBacillus fermented soya bean. International Journal of Food Microbiology 60,163–169.
Melzer, M.S., Boland, G.J., 1999. CHV3-type dsRNAs and the GH2 genotype in apopulation of Cryphonectria parasitica in Ontario. Canadian Journal of PlantPathology 21, 248–255.
Ogawa, J.M., Zehr, E.I., Bird, G.W., Ritchie, D.F., Uriu, K., Uyemoto, J.K., 1995. Compendiumof Stone Fruit Diseases. APS press, St. Paul. MN.
Pusey, P.L., Wilson, C.L., 1984. Postharvest biological control of stone fruit brown rot byBacillus subtilis. Plant Disease 68, 753–756.
Pusey, P.L., Hotchkiss, M.W., Dulmage, H.T., Baumgardner, R.A., Zehr, E.I., Reilly, C.C.,Wilson, C.L., 1988. Pilot tests for commercial production and application of Bacillussubtillis (B-3) for postharvest control of peach brown rot. Plant Disease 72, 622–626.
Rey,M.W., Ramaiya, P., Nelson, B.A., Brody-Karpin, S.D., Zaretsky, E.J., Tang,M., Lopez de Leon,A.,Xiang,H.,Gusti, V., Clausen, I.G., Olsen, P.B., Rasmussen,M.D.,Andersen, J.T., Jørgensen,P.L., Larsen, T.S., Sorokin, A., Bolotin, A., Lapidus, A., Galleron, N., Ehrlich, S.D., Berka, R.M.,2004. Complete genome sequence of the industrial bacterium Bacillus licheniformis andcomparisons with closely related Bacillus species. Genome Biology 5, R77.
Rodgers, S., 2001. Preserving non-fermented refrigerated foodswithmicrobial cultures— areview. Trends in Food Science & Technology 12, 276–284.
Schweizer, M. (Ed.), 2004. The Metabolism and Molecular Physiology of Saccharomycescerevisiae. Taylor & Francis Group, London, p. 480.
Schisler, D.A., Slininger, P.J., 1997. Microbial selection strategies that enhance thelikelihood of developing commercial biological control products. Journal ofIndustrial Microbiology & Biotechnology 19, 172–179.
Schena, L., Nigro, F., Pentimone, I., Ligorio, A., Ippolito, A., 2003. Control of postharvestrots of sweet cherries and table grapes with endophytic isolates of Aureobasidiumpullulans. Postharvest Biology and Technology 30, 209–220.
Suzzi, G., Romano, P., Ponti, I., Montuschi, C., 1995. Natural wine yeasts as biocontrolagents. Journal of Applied Bacteriology 78, 304–308.
Zhou, T., Sholberg, P., 2002. Monilinia fructicula (Winter) Honey, Brown Rot(Hyphomycetes). In: Mason, P.G., Huber, J.T. (Eds.), Biological Control Programmesin Canada, 1981–2000. CABI Publishing, New York, pp. 468–471.
Zhou, T., Northover, J., Schneider, K., 1999. Biological control of postharvest diseases ofpeach with phyllosphere isolates of Pseudomonas syringae. Canadian Journal ofPlant Pathology 21, 375–381.
Zhou, T., Chu, C.L., Liu, W.T., Schneider, K.E., 2001. Postharvest control of blue mold andgray mold on apples using isolates of Pseudomonas syringae. Canadian Journal ofPlant Pathology 23, 246–252.
Zhou, T., Northover, J., Schneider, K.E., Lu, X., 2002. Interaction between Pseudomonassyringae MA-4 and cyprodinil in the control of blue mold and gray mold of apple.Canadian Journal of Plant Pathology 24, 154–161.
Zhou, T., Yu, H., Errampalli, D., 2007. Strategies for biolocontrol of fungal diseases oftemperate fruits. In: Chincholkar, S.B., Mukerji, K.G. (Eds.), Biological Control ofPlant Diseases. The Haworth Press, Inc., Binghamton, NY, pp. 239–281.





























