Embed Size (px)
Citation preview

Plaiit Physiol. (196/7), 42. 1284-1287
Detecting Mutants That Have Impaired Photosynthesisby Their Increased Level of Fluorescence'
Pierre Bennoun2 and R. P. LevineThe Biological Laboratories, Harvard University, Cambridge, Masisachusetts 02138
Received June 21, 1967.
Summary. A technique is described for detecting mutants of Chlaryvdoiimoias rciniar'di
that lhave impaired photosynthesis. It is based on the fact that when photosyntheticelectron transport is blocked there may be an increase in the level of fluorescence similarto that obtained in the presence of an inhibitor of photosynthetic electron transport such
as dichloropohenyldimethylurea.
One way to obtain information about the mecha-nism of photosynthesis is to study mutant organismsthat are unable to carry out normal photosynthesis.The unicellular green algae Chlarnydowiionas recinliardiand Scenedesmnius obliqutus have been used in this way(1-3, 6). A prerequisite for the research with theseorganisms is an efficient and rapid method of screen-ing for the appropriate mutations. A reliable nmethodhas been described (4), but it is not as rapid asdesiredl. In this paper we describe a technique forChlamvwdomloiias reinhardi that is based upon the factthat the steady level of fluorescence reached tindercontinuiotus illuminiation may serve as an in(lex ofelectron flow tlhroughI the photosynthetic electrontransport chain. When photosynthetic electron trans-port is blocked there imav be ani increase in the levelof fluorescence similar to thiat obtained in the presenceof an inhllibitor of electron transport such as dichloro-phenyldimethylurea (DCMU). For colonies of C.reinhlardi comprised of mutant cells unable to carryout nlormiial photosynthesis tlhere is the expectationthat somiie of them xvill showN a steadxs level offluorescence that is highler than that shown l)v acolony of xwil(l-type cells. This expectation has beenrealized via the techniqtie described beloxx.
Materials and Methods
'I'lie \\-ilcl-type strain ( 137c) of C. rcoihlta-d,i xvasused. It w\-as ctiltured anid harxested under conditionspreviously d-rs,cribed (2). and imutations were indutcedwith mllethvlm1ethane stilfonate ( 8). Cells, in thelogar;thl;mic phase of growx th, were harvested ancdxvashled inl 0.068 M potassium phosphlate buffer, pH7.0, ani( resuspended in 10 mil of the lbffer contained
1 This investi-ation xvas supported by -rants GB 2955froml the Nationial Science Foutmdlationi and GM\1 12336from the National Instiiutes of Healtlh.
2 Preselnt address: Institut (le Biologie Physico-Chimiclque, 13 ru-te Picrre Cturie, 75 Paris 5e, France.
in a test tube. The cell concenitr-ationi xas about107 per ml. The test tube was placed in a waterbath nmaintained at 220 and the suspension xwas agi-tated. After equilibrationi to the temperature, methyl-methane sulfonate was added to a final conientratiolnof 0.14 WI. The cells xvere exposed to the muttagenfor 3.5 hoturs. The action of the mutagen wasstopped by dilutiing the cell suspension 20-f0old in thephosphate buffer. The diltited suspens.on xvas thenwvashed twice in the buffer. After washing, the cellsuspension was adjtisted to have approximately1 X 104 cells per ml and 0.2 ml aliquots xvere platedto minimal agar supplemented \\ith 0.2 % socliuniiacetate and 0.4 % yeast extract. Survival xas about15 % under the conditions desicr,ibed here.
The fluorescence of the colonies that formecl afterplating xxvas activated by illuminating themi xvith anOsram HBO 200 highl-pressure mercury arc lamilp ina Zeiss lamip hotising. 'Tlhe light was passed throtiglha xvater filter to remove ox' and infra-red irradiationand then throuighl a Corning glass filtel- #4305 thatcuit off all Xvavelengths above 640 nni. h'lec inltenlsityof the lighlt xx as 4000 loix at the surface of the plate.The fluorescence of colonies was detectable by eye,btut the most sensitive xvav to record the fluorescencexvas to photograplh the colonies tusing red-sensitixePolaroidl film type 413 in a Polaroid type 800 camera.'IThe lens was fitted xxith a Corning glass filter #2030so that only Iight abox'e 640 nmn was transimittedl tothe film. A neutral denisity filter (1.2) xxas alsoplaced over the lens to adjust the exposu-re time toabout ; secondIs. Thle plates xvere illtminilliated for1 minute before the picture xvas taken.
Carhon dioxide fixation by xhole cells of thewxild-type and muttant strains xvas meastured as theincorporation of 14CO2. T-xvo mlI of cells, stuspenidedin minimal meditim (9) xere placed in a 25 ml flaskanid aerated for 10 miniultes at 25° tinder incidentillumination of 40,000 ltox. 'T'hell 0.2 ml of NaaH14(CO1('10 ftmoles, 5 uc) xvas injected inlto the cell stospen-sion, and 0.2 ml samples wereix'ithdraxvn every 20seconds over a period of 180 seconds. The cells in
1284 www.plantphysiol.orgon September 16, 2018 - Published by Downloaded from
Copyright © 1967 American Society of Plant Biologists. All rights reserved.
BENNOUN AND LEVINE-IMPAIRED PHOTOSYNTHESIS-INCREASED FLUORESCENCE
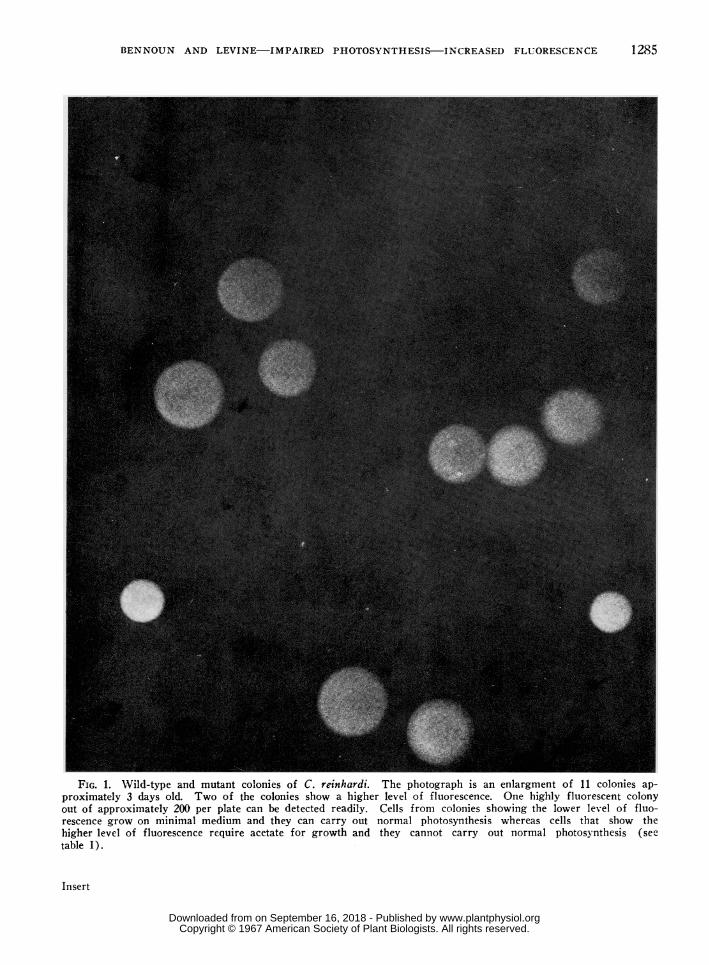
FIG. 1. Wild-type and mutant colonies of C. reinhardi. The photograph is an enlargment of 11 colonies ap-proximately 3 days old. Two of the colonies show a higher level of fluorescence. One highly fluorescent colonyout of approximately 200 per plate can be detected readily. Cells from colonies showing the lower level of fluo-rescence grow on minimal medium and they can carry out normal photosynthesis whereas cells that show thehigher levcl of fluorescence require acetate for growth and they cannot carry out normal photosynthesis (seetable I).
Insert
1285
www.plantphysiol.orgon September 16, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

BENNOUN AND LEVINE-IMPAIRED PHOTOSYNTHESIS-INCREASED FLUORESCENCE
Table I. CO, Fixation and the Photoreduction of DPIP and NADP by the Wild-Type and Muttanit StrainsFor CO., fixation the reaction mixture contained cells equivalent to 100 ug of chlorophyll suspelnded in 2 ml
of minimal meditum. Ten dumoles of NaH14CO3 (5 Ac) were added after the suspension had been aerated in thelight for 10 minutes. The light intensity was 40,000 lux.
NADP-a is the photoreduction of NADP with water as the electron donor, and NADP-b is the photoreductionof NADP withi reduced DPIP as the electron donor.
For NADP-a the reaction mixture (2.0 ml) contained chloroplast fragments (20-25 ug chlorophyll) and thefollowing in umoles: potassium phosphate buffer, pH 7.0, 20; KCI, 40; MgCl2, 5; ferredoxin purified from wild-type C. rcinhardi, 0.0005; and NADP, 0.5. For NADP-b the reaction mixture contained, in addition to the above,the followitng in Anmoles: DPIP, 0.1; sodium ascorbate, 10; and DCM1IU, 0.02. The reaction was run in a cuvetteplaced in the saml-ple compartment of the spectrophotometer. The cuvette in the reference compartment containeda control reactioni mixture from which ferredoxin and NADP were omitted.
For the plhoLoreduction of DPIP, the reaction mixture (2.0 ml) contained chloroplast fragments (20 /ug chloro-phyll) aind the following in /umoles: potassium phosphate buffer, pH 7.0, 20; KCl, 40; MgCl2, 5; and DPIP, 0.1.The DPIP was omitted from the control reaction mixture.
All reactionis wxere run at 250.
Strain CO., Fixation,uXfoles hr-1 mg-, chlorophyll
Wild TypeH 34FH 54FH 54
DPIP
150<1<1<1
2980
4540
each sample were killed with 0.5 ml of a mixturecontaining glacial acetic acid and concentrated HClin a volume-to-volume ratio of 4:1. An aliquot ofeach sample was plated on stainless steel planchettesand counted under a gas flow counter.
The photoreduction of 2,6-dichlorophenolindophenol(DPIP) and of NADP by chloroplast fragmentswere measured according to the mzthod described byGorman and Levine (3).
Resultsand Discussion
It was found that 2 types of colonies appeared onthe plates after treatment of cells with the mutagen;namely, a majority with a low steady level of fluores-cence and a minority with a higher steady level offluorescence (fig 1). In the presence of DCMUall colonlies shlxosed the higher level of fluorescence.Colon es showing the higher level of fluorescence inthe absen,ce of DCMU were selected. These werefound to be comprised of cells that required acetatefor growtlh and were deficient in their capacity tofix carbon dioxide by photosynthesis. Furthermore,when thes)e cells were crossed to cells of the wild-typestrain, the pattern of Mendelian segregation for asingle-gene mutation was obtained. Frolmi these re-sults we conclude that we have obtained mutant strailnsthat have impaired photosynthesis. Fifty mutantstrains have been isolated by the procedure describedabove. Some of the photos)-nthetic properties of 10of the strains have been characterized. Table I sum-mlarizes the results relating to 3 typical strains. Ingeneral, the photosynthetic properties of the 10 mu-tant strains obtained by screening for their fluior-es-cence are similar to those obtained by the earlielmethod (2, 3, 5-7), but importantly the new methodis simpler and more rapid.
,uMoles of oxidant photoreducedhr'1 mg'1 chlorophyll
NADP-a
178<2<2<2
NADP-b
782565<2
Literature Cited
1. BiSHop, N. I. 1964. Mutations of uniicellulargreen algae and their application to studies onthe mechanism of photosynthesis. Record Chem.Progr. 25: 181-95.
2. GORMAN, D. S. AND R. P. LEVINE. 1965. Cyto-chrome f and plastocyanin: their sequence in thephotosynthetic electron transport chain of Chlamy-domiionas reinthardi. Proc. Natl. Acad. Sci. U.S.54: 1665-69.
3. GORMAN, D. S. AND R. P. LEVINE. 1966. Photo-synthetic electron transport chain of Chlamydo-miionas reinhardi. VI. Electron transport in mu-tant strains lackinig either cytochrome 553 orplastocyanin. Plant Physiol. 41: 1648-56.
4. LEVINE, R. P. 1960. A screening technique forphotosynthetic mutants in unicellular algae. Na-ture 188: 339-40.
5. LEVINE, R. P. 1963. The electron transport sys-tem of photosynthesis deduced from experimentswith mutants of Chlanmydonmonas reinhardi. Pho-tosynthetic AMechanisms in Green Plants. NAS-NRC Publ. 1145, Washington, D. C. 158-73.
6. LEVINE, R. P. AND D. S. GORMAN. 1966. Photo-synthetic electron transport chain of Chlanuuydo-mo ias r-einhaardi. III. Light-induced absorbancechanges in chloroplast fragments of the wi!d-typeamid mutant strains. Plant Physiol. 41: 1293-1300.
7. LEVINE, R. P. AND R. M. SMILIAE. 1903. Thephotosynthetic electron transport chain ofChlmam'Ydoinionas reinthardi. I. Triphosphopyri-dine nucleotide photoreductioni in wild-type andmutant strains. J. Biol. Chem. 238: 4052-57.
8. LOPPES, R. 1966. Damage induccd by methyl-metlhanesulfon1ate (MMS) in ChlaniYdontonasreimmhairdi. Z. Vererbungslehre 98: 193-202.
9. SUEOKA, N. 1960. Mitotic replication of deoxy-ribonucleic aci(d in Chlanmvdononmas erci,hardi.Proc. Nat]. Acadl. Sci. U. S. 46: 83--91.
1287
www.plantphysiol.orgon September 16, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.













![Evolution from Primitive Life to Homo sapiens Based on ...carried out in bacteria by Noboru Sueoka [27]. Then, my group investigated the cellular amino acid composition not only of](https://img.pdfslide.us/doc/110x75/5e6b41cf0a650825432d215c/evolution-from-primitive-life-to-homo-sapiens-based-on-carried-out-in-bacteria.jpg)