Embed Size (px)
Citation preview

1
Bangarang January 2014 Backgrounder1
Designing Plankton Nets (A Manual)
Eric Keen
Abstract Introduction Background Starting Points The Open-Area Ratio Initial Filtration Efficiency (IFE) Sustained Filtration Efficiency (SFE) Net Model Case Study Designing the Net Step 1: First Steps Step 2: Mesh Size Porosity Step 3: Diameter Step 4: Tow Duration Constraints Practical Proxies for Tow Volume Results Step 5: Minimum Open-Area Ratio Green vs. Blue Waters Results Step 6: Solving for Net Length Conclusion Appendix 1: Case study Literature Cited Figures & Tables
1 Bangarang Backgrounders are imperfect but rigorous reviews – written in haste, not peer-reviewed – in an effort to organize and memorize the key information for every aspect of the project. They will be updated regularly as new learnin’ is incorporated.

2
Abstract A good net must perform in many ways, but foremost it must filter a sufficient volume of water for the right size of organism with consistent efficiency. One can achieve such a net by designing with the objective(s) of a study as the starting point and the directives of maximizing and maintaining filtration efficiency as a compass. The goal is to finish the design process with a selection of appropriate configurations from which to choose. While many decisions must be made to design a net in full, three key parameters are the most consequential and need to be decided first: mesh size, mouth diameter, and overall net length. The general shape of a net, the speed at which it is towed, and the care it receives will contribute to its initial (inherent) filtration efficiency. Then, by manipulating the “key three” variables, a net can be designed to sustain its initial filtration efficiency for a target tow volume. The design process begins by (Step 1) rigorously answering basic questions about study objectives, the study area, and the resources at hand. These answers will (2) determine the target groups of your study and the mesh size required to retain them. The highest porosity material of that mesh size can then be selected. Then, by considering target groups and the resources available, (3) a range of viable mouth diameters can be defined. Finally, to determine overall net length, (4) a target tow volume must be set, which (5) informs the minimum OAR permissible given mesh size, mouth diameter, and the qualities of the study area. 6) The one variable that remains is the overall net length, which can now be manipulated to achieve the minimum OAR. A case study is used to demonstrate the process in practice.

3
Introduction
A poorly chosen net can prove fatal to a plankton study. In the field, inappropriate nets will bring about inefficient use of ship time and space, the exhaustion of crew, clogging, and the extrusion, escape or avoidance of target species. During analysis, the wrong net may plague a study with insufficient, unreliable, and irrelevant data that renders those costly hours of fieldwork useless. Choosing the appropriate net begins with the same concerns fundamental to any field-based research: 1) the objectives of your study; 2) the conditions of your study area; and 3) the resources that are available to you. These considerations must be weighed to arrive at an appropriate configuration, which can be daunting for anyone with small budgets of time or money. This guide can help to leverage such constraints as tools in a decision-making process that, in the end, provides a range of optimal parameter combinations. This paper draws on articles throughout the rich history of field planktology (reviewed in Wiebe et al. 2003), but relies most heavily on four: Tranter & Smith (1968), Harris et al. (2000, esp. chapters 2 and 3), Bernardi (1984), and, most recently, Ohman (2013). These give readers a newfound respect for such questions as seemingly simple as, “Which net should I use?” They are also excellent guides for study design and organism preservation. However, upon finishing these papers readers might remain uncertain where to begin. Understandably, given these papers’ broad coverage of topics for a wide range of organisms, their advice cannot be too specific or concrete. This guide errs on the other side of that compromise, by attempting to provide specific, step-by-step instructions in net design at the cost of a substantially limited scope. This manual is intended for studies of zooplankton that invoke single- or dual-net samplers, and is based on the standard cylindrical-conical (“cyl-cone”) design advocated by UNESCO Working Party no. 2 (WP-2) 2. While not ideal for all circumstances, the WP-2 cyl-cone sampler has been a pillar of net design for zooplankton studies ever since. The WP-2 standard is intended for studies of plankton 200 micrometers to 10 mm in length within the upper 200m of ocean (Tranter & Fraser 1968, schematic on p. 155). As such, it is most readily helpful to small-scale, low-budget projects conducted by single researchers or small teams. However, the principles and process invoked here should be of use in a variety of applications. The scope here is limited to the process of deciding upon three key net parameters: 1) diameter of the net’s mouth, 2) overall net length, and 3) aperture of the mesh material (i.e., mesh size). While a net has many other parameters that can increase its efficacy (e.g., net color, bridle and flowmeter rigging, strobe light use, etc.), these “key three” are all that one needs to get started. They combine to influence a net’s filtration efficiency and a net’s propensity to clog. They are also the most readily observable when considering a candidate net, so it is convenient that the decision process is centered upon their interaction.
2 Working Party no. 2 was a team of scientists who designed the net in an attempt to standardize field equipment and best practices for a specific size class of organisms (Tranter & Fraser 1968).

4
Background
Starting Points___________________________________________________________________ To perform well and consistently, a good net must meet this ordered list of requisites:
1. Sample for a target duration before clogging deteriorates filtration efficiency and the size selectivity of the mesh.
2. Retain the organisms targeted by the study’s objectives, minimizing their ability to avoid the net and extrude through the mesh.
3. Filter a sufficient volume of water over sufficient depths to provide statistically robust results. 4. Be sustainably operable given the finite energies of the crew and resources at disposal.
The imperative upon which all other criteria hinge, and the concern that will organize and motivate the decision tree outlined below, is the need to maximize and maintain a net’s filtration efficiency (1). In the end, however, if a net suffers from a poor filtration efficiency to begin with and clogs easily, it will fail at both capturing target organisms (2) and filtering sufficient volumes of water at a sustained efficacy (3). The resources devoted to the study (4) would therefore be wasted. Filtration efficiency is the percent of water a net passes through that is actually sampled. As we will see, net performance can be discussed in terms of both initial and sustained filtration efficiency (IFE and SFE, respectively)3. In addition to diminishing the volume sampled, clogging over the course of a tow can cause a net to become increasingly size selective, which introduces a host of biases to results. Filtration efficiency can also affect rates of net avoidance and escapement. See Tranter & Smith (1968) for an excellent review.
The Open Area Ratio___________________________________________________________ Designing for high filtration efficiency means first maximizing IFE and then, within those design confines, maximizing SFE to the extent possible. The “open-area ratio” (OAR) is a useful metric that plays a role in both directives. OAR is the ratio of the filtering mesh area to the area of the net’s mouth opening4. The “filtering mesh area” refers to the total open area of the net’s mesh, which can be found by multiplying the net’s surface area by the mesh’s “porosity”, or percent open area.
OAR = β aA
β = porosity a = total area of net A= mouth area
A net’s surface area can be calculated using the net’s geometry (shape, total length and mouth diameter). Note that mouth diameter influences both the numerator and denominator of the open-area ratio. Hence our “key three” parameters combine to determine our net’s OAR, the defining metric of filtration performance. The OAR of a net plays a complimentary role in IFE but a defining role in sustaining that efficiency over the course of a tow (SFE). In general, the higher the OAR ratio of a net, the better for both IFE and SFE. The rub, however, is that other concerns constrain a net’s design, and a sufficient OAR can sometimes be difficult to attain. Net selection is therefore a game of trade-offs, driven by the conflicting priorities of maximizing OAR, maximizing sample size, and minimizing cost and effort.
3 Tranter & Smith (1968) abbreviated filtration efficiency as F. In an effort to emphasize the distinction between initial and sustained efficiency, we use a different notation here. 4 In Tranter & Fraser (1968) and other works, the open-area ratio is abbreviated simply as R. Here, we follow Ohman (2013).
(1)

5
Initial Filtration Efficiency (IFE)_____________________________________________ The shape of a net and the way it is used will give it an inherent, or initial, filtering efficiency. Generally speaking, a higher IFE means better use of sampling effort and more flexibility in the steps to come. IFE values for conventional net types range from 75% (simple conical nets) to upwards of 110% (reducing collar cyl-cone nets). The WP-2 has an IFE of 94% (Tranter & Fraser 1968). Some design for 100% IFE to simplify calculations (Tranter & Smith 1968). Net Shape The geometry of a net governs the way in which water flows through and around it. The hydrodynamics of this flow are sensitive to a slew of variables (reviewed in Tranter & Smith 1968) and can be summarized with the following principles of design: Simple cones are bad. Simple conical nets are among the least efficient, at about 75-85% IFE. They need to be longer (higher OAR) to sample the same volume of water as efficiently as a cylindrical-conical net of similar shape. Nonporous encasements are bad. Nets housed within nonporous encasements are not efficient (Fig. 6d in Tranter and Smith 1968). Cylindrical collars are good. Both IFE and SFE increase when a porous cylindrical section of mesh is added ahead of the cone. This is the basic net design used in this paper’s models (Fig. 1). Water rejected by the tapering area of the cone can escape through the cylinder’s mesh rather than out the mouth where animals would be lost (Currie 1963). A cylindrical collar also adds more filtering area while adding less to the overall length of the net than a conical form would. In other words, a cylindrical-conical net has a higher OAR than a conical net of the same overall length. Non-porous collars are good too. Non-porous collars, which may be necessary for some nets with choke lines (e.g., plummet nets, Heron 1982), can also yield high IFEs (Tranter and Smith 1968). Note, however, that the length of a non-porous collar cannot contribute to the filtering area used in OAR calculations for the design; that is, a non-porous cyl-cone net may be more efficient but it will have to be longer. Both porous and non-porous cylindrical collars alike tend to oscillate slightly underway, inducing a “self-cleaning” behavior (Smith and Clutter 1965). Mouth-reducing collars can be even better. Enlarging the terminal radius of the cylindrical section to create a “mouth reducing cone” or “collar” creates an area of low pressure behind the net mouth into which incoming water will accelerate (Fig. 6c in Tranter and Smith 1986). This increases IFE to 110% or more, among the most efficient of any design (Smith, Counts, and Clutter 1968). However, they are less commonly available and the goal here is to determine minimum pragmatic requirements. The net model in this guide will not incorporate mouth reducing cones. Be careful with side angles. IFE declines sharply when the net’s side angle, the angle of incidence of water striking the mesh (Θ in Fig. 1), falls below 75° (or when the ratio A/a rises above 0.2). It is therefore important to increase side angle to the extent possible, which translates into increasing net length. Similarly, the side angle in mouth-reducing cones should be maximized (the complement of this angle was called the “angle of expansion” in Tranter & Heron 1967). For top efficiency, the reduction cone’s side angle should be more than 86.5° (Pankhurst and Holder 1952). This is achieved by either keeping the discrepancy between the two radii of the cylindrical section small, or, preferably, by lengthening the cylindrical section. OAR has limited effect on IFE. As outlined above, the boosted IFE of a cyl-cone net can be explained by both mechanical effects (e.g. self-cleansing) and the higher OAR that comes with the addition of filtering area. However, due to hydrodynamic theory (which is beyond the scope of this paper) improving OAR only boosts IFE up to a certain point; the effect plateaus above an OAR of 3. Instead, OAR (along with the net features implicated in it: mouth diameter, mesh size, and overall length) becomes much more influential in matters of sustained filtering efficiency (see below).

6
Tow Speed Nets towed at very low speed (< .5 m/s, or 1 knot) filter at a lower efficiency (Tranter & Smith 1968). Again, however, the effect plateaus: at speeds greater than 0.5m/s, IFE is unaffected. In the case study’s literature review (Table 2), most tow speeds were between 1.5 knots (.77 m/s, Fiedler et al. 1998) and 2.5 knots (1.28 m/s, Mackas & Galbraith 2002, Schulenberger 1978). The median tow speed reported was 2 knots, or ~1 m/s. In an effort to standardize methods, Working Party No. 2 recommended that vertical tows – which are typically slower than oblique hauls -- be raised at 0.75m/s (Tranter & Fraser 1968). Higher tow speeds (> 1.5 m/s) may allow for shorter tow durations and may or may not minimize avoidance (Clutter & Anraku 1968), but the pressure drop across the mesh at high speeds can damage specimens. Investigators wishing to collect specimens for live experiments or precise morphological studies must prioritize minimal tow speed. Furthermore, the towing cable or deck rigging for the cable can also become strained at high speeds, and manual retrieval of the net is very tiresome at anything above idle speed – at least for this author. Samplers specifically designed for high tow speeds, such as continuous plankton recorders, are beyond the scope of this paper. Washing & Care Plankton dry more completely on monofilament nylon mesh (which has now become the industry standard) compared to older silk designs. The presence of remnant plankters and debris in successive tows can introduce variable IFEs over the course of a study (Tranter & Smith 1968). Thorough rinsing between tows with uncontaminated fresh water is critical to maintaining consistent OAR and IFE values. Harris et al. (2000) outlines best practices for net care and storage.
Sustained Filtration Efficiency (SFE)_____________________________________ Over the course of a tow, clogging will cause the net’s initial filtration efficiency to deteriorate. Suspended detritus, phytoplankton, and the target organisms themselves can have a nefarious impact on a study’s results. Clogging introduces more and more bias over the course of a tow, not only by reducing overall filtration efficiency, but by selectively sampling a progressively constrained size range of organisms (Ohman 2013). When testing the performance of a net, clogging can be monitored by comparing readings from two flowmeters, one mounted inside and the other outside of the net during a sequence of variable-duration tows (Tranter & Smith 1968). The rate at which clogging occurs is a function of the conditions of the study area (over which we have little control) and the design of our net. By convention, once filtration efficiency has dropped below 85% of initial, the net has become clogged (Smiths, Counts & Clutter 1968). Sampling enough water before this occurs is the central goal. SFE is governed by mouth area, mesh size and porosity, and net length. It is therefore directly related to a net’s OAR. Net shape can also have an affect; as explained above, the geometry of a net can introduce “self-cleaning” oscillations that increase the duration of efficient filtration. A cylindrical or mouth-reducing section can also add filtering area to the net, increasing the OAR and therefore SFE. Mesh Size The width of holes in mesh influence SFE in two ways. First, fine gauze will clog more readily than coarse gauze merely because it catches more particles. In fact, the effective tow duration of a net declines as a function of the square of the mesh width (Smith, Counts & Clutter 1968). In order to compensate for the inherently low SFE of fine mesh nets, OAR must dramatically increase. For mesh sizes greater than 300µ, Tranter & Smith (1968) recommend an overall OAR of 5 or higher, with an OAR of 3 in the conical portion and an OAR of 2 in the cylinder. They warn that any mesh smaller than that would require an OAR of 9 or higher, with 3 in the cone and

7
6 in the cylinder. However, in the same monograph, Working Party 2 proposed a standardized net schematic with 200µ mesh that had an OAR of only 6:1, 3 in the cylinder and 3 in the cone (Tranter & Fraser 1968). To err on the conservative side (sensu Ohman 2013), we will hold minimum OAR at 9:1 for mesh apertures less than 300µ. But Working Party No. 2 makes the good point that such arbitrary thresholds should be taken only as guidelines. Second, mesh size determines a net’s porosity, which is a term in the equation for OAR. While one would expect porosity to decrease linearly with shrinking mesh size, different widths of Nylon monofilament are used in weaves for various mesh sizes, such that porosity levels rise and fall abruptly as mesh size increases. This author was able to find two distributors that reported porosities for the Nitex mesh they sell: Wildlife Supply Company (WSC)5 and Dynamic-Aqua Supply, Ltd. (DAS)6. Since mesh of the same aperture can be available at different porosities (Fig. 2), it behooves investigators to seek out the most porous mesh available. Doing so may allow a net to be shorter7. While high-porosity material may not be as supple as thick-filament (low porosity) mesh, weave strength is not of great concern for small-scale studies with low tow speeds.
Designing for OAR_____________________________________________________________ Controlling for the size and abundance of particles in the water, the open-area ratio is a sensitive metric for the duration of filtration that is possible before clogging sets in. Minor design modifications can substantially change the effective duration of a tow. Smith, Counts and Clutter (1968) found that a doubling in their nets OAR from 3.2 to 6.4 increased the volume filtered efficiently by sixfold. The OAR of a net is a compromise between porosity, filtering area, and mouth area – which are products of a net’s mesh size, diameter, and overall length – all of which are constrained by the organisms of interest, target volume, and resources. As such, the OAR represents a game of trade-offs. Here is the OAR equation, repeated for convenience:
OAR = β aA
All else being held constant, lengthening a net will increase its OAR, as will choosing a mesh size that is more porous. Note that since diameter is implicated in both the numerator and denominator of the OAR equation – both of which involve exponents -- the nature of its impact on OAR is depends on the net size in question. Generally speaking, however, the largest OARs are achieved with the longest total length, smallest mouth openings, and most porous mesh. Conversely, the smallest OARs occur in the shortest nets with large mouth openings and low-porosity gauze.
Net Model__________________________________________________________________________ Parameter Bounds To maximize IFE, we will only consider configurations that yield an OAR greater than 3. As we manipulate net length relative to diameter, we will not allow the side angle of the net to fall below 75° (which would decrease IFE); in other words, for a given diameter, there is a certain minimum length the net must be. The tow speed we use in calculations of tow duration will not fall below 0.5 m/s nor exceed 1.5 m/s, and the average will be 1 m/s (2 knots). The average IFE for conventional cyl-cone net configurations reported by Tranter & Smith (1968,
5 www.wildco.com 6 http://www.dynamicaqua.com/nitex.html 7 Mesh aperture and porosity can be difficult to know if a net does not come with documentation. Smith, Counts, & Clutter (1968) provide instruction on measuring and calculating these values using a microscope.
(1)

8
Table 5, n=7) was 94.43% ±2.99%. To remain conservative, our model will have an IFE one standard deviation below that mean, at 91.4%. Geometry See Fig. 1 for the geometry of the model net used in this guide and Table 1 for a definition of its parameters. Cod-end: Because nets reduce not to a point but to a terminal cod-end bucket, the “cones” of nets are actually tapered cylinders. The mean cod-end radius of 63 nets sampled from the Scripps Institution of Oceanography Pelagic Invertebrate Collection (SIO PIC) was 5.475cm (sd=2.1cm, L. Sala, pers. comm.). A cod-end of radius 3.75cm was recommended by the Working Party No. 2 (1968), which is a value within the standard deviation of the PIC sample. Because the WP-2 radius ensures the model will err on the side of underestimating OAR, it will be used as the radius in the model net. Collar length: Because the cylindrical section contributes much to overall OAR with a comparatively minor addition of total length, the cone’s length will be minimized. The strict minimum of the side angle (Θ) is 75° (Tranter & Smith 1968), but we will keep our model’s at the side angle used in the WP2 net, 81.33°. With this angle immobilized, we can calculate the length of the conical section ( con) of any net by knowing only the radii of its mouth (R) and cod-end (r, given above as 3.5cm).
con = tanθ(R− r)
The length of the cylindrical section will comprise the remainder of the net’s overall length. This means that low OAR nets will have proportionally longer conical sections than those with a high OAR. In nets with an OAR of 10 or higher, the majority of its length will be composed of cylindrical section. In fact, the length ratio of the cylindrical and conical sections may swing from 2:3 to 6:3 as net length increases (Smith & Clutter 1965, Tranter & Smith 1968). The overall length of the net ( tot) will be the simple sum of the lengths of the cylindrical and conical sections ( cyl and con, respectively).
tot = con + tot
Similarly, the total filtering area of the net (a) will be the sum of the lateral surface areas of each section (acyl and acon).
a = acyl + acon
Using conventional geometric equations, these surface areas are calculated as follows:
acyl = 2πcyl
acon = πs(R+ r)
Where s is the side length of the conical section of net, which can be thought of the hypotenuse of a right triangle with sides of length con and R – r. As such, the Pythagorean relationship applies:
s = con2 + (R− r)2
These equations will be used to calculate overall net length, which shall be the last free variable after diameter, mesh size (porosity) and OAR are constrained. To verify that the above equations yield accurate results, the WP2 schematic provided in Tranter & Fraser (1968) was used as a check. The schematic displays a net with 200µ mesh aperture, 0.57m diameter, .95m cylinder length, 1.66m side length (different from cone length), and
(2)
(3)
(4)
(5)
(6)
(7)

9
an OAR of 6:1. The porosity for this mesh size is reported as 45% by Dynamic Aqua Ltd. (Fig. 2; the same value is given in Table 4 in Tranter & Smith 1968). By assuming this net has a 0.075m diameter cod-end, simple geometry can be used to calculate the cone’s side angle (81.33°) and the length of the cone (1.64m). The WP2’s total length is the sum of its two section lengths, or 2.59m. Using only the mesh size, diameter and OAR reported in the schematic, the above equations calculated that the overall length of the net would be 2.566m, with 0.961m in the cylinder and 1.604m in the cone. These values are within 2% of the actual dimensions.
Case Study_________________________________________________________________________ To illustrate each step of the decision making process, Appendix 1 details a case study set in the Great Bear Fjordland of northern British Columbia, Canada. The study’s primary objective is to document trophic interactions between rorqual whales and their prey. Primary target species include locally dominant euphausiids and large-bodied copepods, as well as amphipods and chaetognaths.

10
Designing the Net As outlined above, the need to maximize initial filtration efficiency has already yielded several constraints: 1) tow speed must be above 0.5 m/s; 2) the net should have a cylindrical collar or mouth-reducing cone; 3) the minimum acceptable OAR is 3. If mesh size is less than 300 microns, this minimum OAR increases to 9 (Tranter & Smith 1968). Sustaining this filtration efficiency, however, presents another procedural puzzle: The coupled interactions of the “key three” parameters (diameter, length, and mesh size) are such that several different configurations can result in the same OAR. They must therefore be constrained in a certain order according to a certain rationale. Each design decision in the process outlined below will inform the following step. The result should be an optimal combination (or range thereof) of the “key three” parameters. The figures associated with each step provide a quick way for readers to work through the process with their own study in mind.
Step 1: Organizing Questions______________________________________________ Determining the right net for your study first requires that you rigorously define your objectives and familiarize yourself with the study area. Having strict answers for the following questions will direct the remainder of the net selection process. 1. Objective: The primary motivator of any research initiative is its central question(s). Without it, there would be no study at all. What are the study’s motivating questions or hypotheses? Do they have to do with the diversity of an area, the density or abundance of certain species, the distribution of those species in space or time, or a combination of these objectives? 2. Target groups: In one form or another, all zooplanktoon studies set out to gather data in a certain area for a specific taxon, group of taxa, or groups within a taxon (e.g., life stages). Therefore, one should first identify target organisms and lifestages with inordinate specificity. 3. Study area: Where is the study area, and what bathymetric, geographic, and oceanographic features should be taken into account during study design? Are the waters very shallow or deep? Are there narrow channels? Are the waters turbid and productive (“green waters”) or clear and free of suspended debris (“blue waters”)? Will the weather be extremely cold and wet? 4. Resources: How will field conditions and equipment constraints limit the capacities of the crew? Will the net be deployed and retrieved manually, or with electrical or mechanical help? Is there adequate deck space for manual hauling and sample preservation? 5. Design: What kind of sampling design do these goals necessitate? Day-time or night-time sampling? Vertical tows or oblique tows? Will these tows be vertically integrated or stratified? What sampling frequency and coverage need to be planned for?
Step 2: Mesh Size______________________________________________________________ Once target groups have been identified and narrowed down to life stages of interest based on the study’s objectives, the mesh size can be determined. On one hand, you want a mesh size small enough to catch the study’s target group; if there is more than one species or ontogenetic stage of interest, that with the smallest width should be the benchmark. On the other hand, you want to minimize the capture of marine snow and smaller species that clog and burden sample analysis. Therefore, you should select a mesh size just small

11
enough to capture the target group that is “skinniest” at its widest (i.e., with the smallest cross-sectional size at its broadest orientation). If the mesh aperture you require is less than 300µ, remember to revise your minimum OAR to at least 9:1 (after Tranter & Smith 1968). Porosity An upper limit on your mesh aperture also provides a constrained range of mesh porosities to expect. Because Nitex mesh of different apertures is made from monofilament of varying thickness, it is not enough to assume that smaller mesh sizes have lower porosities. Refer back to Fig. 2 to determine the porosity of your maximum mesh aperture.
Step 3: Diameter________________________________________________________________ Mouth diameter will be constrained by both target organism behavior and the study’s resources. Net avoidance – zooplankton actively dodging the net -- is a serious concern in mesozooplankton studies and its severity depends upon many factors (Weibe et al. 1982). These include time of day (Fleminger and Clutter 1965); light regime (Isaacs 1965); size, shape, and color of the net (McGowan and Fraundorf 1966); speed of tow (Brinton 1967); species (Clutter and Anraku 1968); sex or developmental stage of the organisms; their physiological state (Laval 1974); and absolute density (Boyde et al. 1978). Avoidance is of special concern for euphausiids (Brinton 1962). Avoidance effects may also be mitigated during analysis; because euphausiid capture efficiency is believed to differ between day and night samples due to visual net avoidance (Shaw & Robinson 1998), some studies (e.g. Mackas et al. 2000) apply a correction factor to euphausiid results. Here, however, the question is how the key three parameters can be designed to minimize avoidance. A larger net is not inherently better at filtering, but the risk of avoidance makes it better at sampling certain taxa representatively (Pearcy 1983, Tranter 1963), especially rarer species (McGowan & Fraundorf 1966). McGowan & Fraundorf (1966) focused on the efficacy of different net sizes in sampling for diversity and abundance, and the susceptibility of various designs to biases introduced by species patchiness and ability to avoid. Mouth diameters in their study ranged from .2m to 1.4m. Their sampling design held other variables constant, including mesh size (550 micron), tow speed (3.4 km/hr, 1.85 knots, 0.9 m/s) and volume sampled, in order observe the sole effect of mouth opening diameter on net efficacy. Their analyses also untangled biases dye to the patchiness of plankton aggregations and those caused their active avoidance of the net. The size of the sampling device did in fact have an effect on estimates of zooplankton diversity. With regards to diversity sampling, the nets performed in the following ranked order: 1.4 > 1.0 = 0.4 = 0.8 > 0.6 > 0.2 m. In terms of abundance estimates, the nets also varied in performance: 1.4 > 1.0 > 0.8 > 0.6 > 0.4 > 0.2 m. We are presented with a suite of trade-offs. We must decide what to gain and what to lose based on our objectives, the study area, and our resources – in that order. Mouth diameter must be maximized to sample an adequate volume of water and to minimize the ability of target species to avoid the net. However, because diameter occurs in both area terms of the OAR equation, its impact on OAR is nonlinear and depends on the porosity of the mesh size used. There are also reasons to minimize diameter: a larger diameter net is a longer, more costly, and more cumbersome net with more drag under tow. This would change cable angle and sampling depth for a give tow speed (Tranter & Smith 1968). Other considerations include deck space, towing hardware, and the stamina of the crew retrieving the net multiple times a day for weeks at a time. Tows of large-diameter nets over short distances may sample the same volume of water as a longer tow with a small-diameter net. And while wide nets minimize the risk of avoidance, they are also more susceptible to the effects of patchiness (McGowan & Fraundorf 1966).

12
Step 4: Tow Duration__________________________________________________________ “Plankton nets are an efficient means to concentrate organisms from relatively large volumes of water” (Ohman 2013). With two of the key three parameters constrained, the remaining design question is net length. To answer, however, some ancillary issues must first be addressed. The first of these is target volumes, which will be constrained with knowledge about the habitat preferences of target taxa and the conditions in the study area. Without sufficient sample size, statistically robust analyses of our data would be impossible and the whole endeavor would be a waste. Many considerations go into a target sample volume. The first is maximum required tow volume. The second is the tow distances that could yield that volume given a certain diameter. The volume sampled (V) by a tow of a certain distance (D) is determined by the net’s mouth area (A) and its initial filtration efficiency (IFE):
V = IFE ⋅A ⋅D Constraints from Sampling Design Many brief tows, or one longer tow? For example, if you are interested in the horizontal distribution or patchiness of zooplankton aggregations, dividing your effort into a sufficient number of shorter tows rather than a single long tow would be more appropriate. However, this can drain preservation supplies and crew morale. Insights from the Literature What are typical tow volumes for various studies? Most of the published zooplankton studies relevant to our case study area did not report tow volumes. Exceptions were Mackas & Anderson (1986), who used a small diameter net for short oblique tows of 4-10m3 in a mesozooplankton survey, and Miller et al. (1984), who used a stratified vertical sampling regime. Based on their reported net size and towing distances, this author estimates that their tow volumes ranged from 50 to 380 m3. McGowan & Fraundorf (1966), in their study of net size efficacy, shot for a standard sampling volume throughout their tows of single nets with various mouth sizes. They averaged 368.65 m3 per tow (n=24 tows), with a standard deviation of 74.12 m3. Schulenberger (1978), in his study of central gyre hyperiid amphipods, had a target sample volume of 400 m3. CalCOFI (California Cooperative Oceanic Fisheries Investigations) has been conducting zooplankton tows at designated stations in southern California waters since 1949, the results of which are publicly available online (http://www.calcofi.org). This study operates from large oceanographic vessels, employs a variety of nets for different purposes, and has used different equipment over the course of its history (Ohman & Smith 1995). Investigators increased tow depths to 210m in 1969, and switched from a 1.0m bridled single net to a 0.71m bridleless BONGO net in 1969 (both with 510-550µ mesh)(Ohman & Smith 1995). However, their scientific objectives have remained more or less the same, and there is much that small-boat studies can learn from their records. Between 1951 and 2012 their 46,502 tows for large mesozooplankton have yielded an average tow volume of 434.7 m3, with a standard deviation of 147.5 m3 (Fig. 3). Constraints Imposed by Study Area How does the study area constrain my sampling volume? If, like our case study, sampling must occur within a complex of narrow coastal channels, large oblique tow distances may not be an option. All North Pacific studies of our primary taxa of interest (euphausiids and calanoid copepods) that used oblique tows sampled no shallower than 100m (Coyle & Pincuk 2005) and as deep as 500m (Trevarrow et al. 2005). Vertical tows are constrained more by water depth than by channel width. Among the papers this author reviewed, those using vertical tows had the following operating depth ranges: 100m (Coyle & Pincuk 2005), 185 m (in a fjord, Osgood & Frost 1994) and 250m (in a fjord, Tanasichuck 1998; Mackas 1992), and 1000m (Miller et al. 1984).
(8)

13
Practical Proxies for Tow Volume For small boats, it is typically impossible to know the volume of water that was sampled in a tow until you retrieve the net and check the flowmeter. In practice, therefore, a proxy for volume sampled, such as tow distance or duration, must be used to decide when to end a tow. A means of translating among tow volume, distance, and duration is therefore of great use. This can be done by equating the distance term in the conventional rate equation:
rtow =D
(tend − tstart )
rtow = tow speed-over-water D = distance of tow tend = end time of tow tstart = start time of tow
to that in the volume equation (Eq. 8), then solving for volume sampled (V). The result:
V = Artow (tend − tstart ) IFE
Assuming that no clogging occurs during a tow at 1m/s speed-over-water and that IFE is 85%, we can then predict the volume we sample from the duration of our tow (Fig. 4). An example of such relationships can be seen in the CalCOFI dataset (Fig. 5). With their net configuration, the average tow volume, ~430 m3, was obtained after approximately 15 minutes of towing (but note the bimodal distribution; two two protocols may have been used). These data representations can also display the effect of clogging at longer durations, which would manifest itself in a leveling off of the curve at greater durations. This did not appear to occur in the CalCOFI datasets. In order to correct for the effect of water current on the “apparent” tow speed of your net, it is important that speeds are recorded as speed over water. If speed over water is known, simple calculations provide a more practical proxy for volume that can be incorporated into protocols. It is essential to ground-truth these calculations in the field using a calibrated flowmeter, both before official sampling commences and repeatedly throughout. Results Once a maximum sampling volume is determined, use Fig. 6 to translate between towing distance and sample volume for different mouth diameters. This allows you to determine a range of expected tow distances, which will then be used to determine the open-area-ratio required of your net.
Step 5: Minimum Open-Area Ratio_________________________________________ With target tow volumes identified, determining minimum OAR (MOAR) is the last task before one can decide upon an overall length. Because all studies are confined by logistics and limited resources in some respect, determining the true minimum OAR specifically for a study is invaluable as one weighs the feasibility of sampling design and explores the equipment options available. MOAR for a net can be set by the IFE, the required mesh size, or the desired volume per sample. We’ve learned already that to maximize our IFE, the OAR must not fall below 3:1. If the mesh aperture is at least 300µ, Tranter & Smith (1968) recommended an OAR of 5:1; if aperture is smaller, they recommended a minimum of at least 9:1. Ohman (2013) set a minimum at 6:1 for all nets, but suggested that 10:1 may be needed in some
(9)
(10)

14
environments. Nets used by Mackas & Galbraith (2002) in the vicinity of the case study area reportedly had an OAR of 11.5. However, there is much room for deviation within these conventions, and many different configurations can yield a single OAR. Green vs. Blue Waters The volume you desire to sample can also establish the MOAR. Clogging becomes a bigger issue the farther a net is towed, at a rate that depends on the turbidity of the study area. In coastal “green waters” that are nutrient and detritus-rich, clogging occurs faster than in offshore “blue waters” and therefore require a higher OAR. The relationship between MOAR and the distance towed (by proxy, the volume sampled) has been described by two equations (Smith, Counts and Clutter 1968): For “Green waters”:
logOAR = 0.38(logVA)− 0.17
For “Blue waters”:
logOAR = 0.37(logVA)− 0.49
Where V is the volume of the sample and A is the area of the mouth opening. The V/A term (which is more or less equivalent to distance towed, if IFE is assumed to be 100%) adds some versatility to the equation; if you know only the distance you want to tow, or your desired sampling volume, or the diameter of the net mouth, you can still narrow your options. Results For each net diameter you are considering, find the distance you must tow in order to achieve the maximum sampling volume you hope to accommodate. Bring these distances to Fig. 7, and, with a sense of the productivity of your study area, determine the minimum OAR values that will be required of your net.
Step 6: Solving for Net Length______________________________________________ We have now set the target mesh size, diameter, and sampling volume for our study. In doing so, we have determined the porosity of our mesh and the minimum acceptable OAR for our net. Given these fixed or constrained parameters, we can now explore the length options that make that OAR possible. We constrain this parameter last because the other two, mesh size and diameter, stem directly from the basal objectives of the study. There are certainly reasons to minimize the length of the net, including the high cost of mesh yardage, the loss of deck space, and the difficulties of handling the drag. Remember, however, that it is better to purchase a costly, bulky but effective net than one that is cheap, easy to handle but scientifically useless. In Fig. 8, the designer can go to the frame that corresponds to her net’s minimum OAR. On the y-axis is the porosity range that corresponds to the mesh size of the net. Then the color-coded lines that best match the candidate diameters can be identified. The point of intersection of diameter and porosity lay over the necessary length for the net (x axis). Rarely will the answer be cut and dry; most of us will approach this final step with a range of mesh sizes and diameters still under consideration. It often requires returning to the roots of this process to weigh the candidate configurations against each other. If the net length required by a certain combination of mesh size, diameter, and sample volume is prohibitive, the constrained parameters may have to be compromised. If so, they should be reconsidered in the reverse order
(11)
(12)

15
that they were pinned down: first sample volume (Tranter & Smith 1968), then diameter, and finally, mesh size. If you must resort to changing the mesh aperture of your net, then the prudence of sampling with a net (or the sensibility of the sampling objectives in general), may need reconsideration as well. Hopefully it won’t come to that. Unless there is no other way, our questions should not be compromised by the methods at our disposal.
Conclusion With this information in hand, we can move forward with designing other features of the net. There is much more to appropriate net design than mesh size, diameter, and length. Many have been encountered during the above process: net shape, tow speed, and maintenance. There is also the fundamental question of whether a net is a better sampler than bottles or optical or acoustic methods in the first place (Ohman 2013 and Harris et al. 2000 are excellent resources here). There are rigging considerations too: Where will the bridles for the net be, and how will the net attach? Should you add weight to your net? Should the net close or open under your control? Where will the flowmeter be mounted? Should the net have a strobe light? Many decisions still need to be made. This guide’s scope was limited to the “key three” features because 1) it addresses the need for a paper that provides a clear starting point for the net design process, 2) those three features strongly influence the all-important filtration efficiency of the net and 3) they are the most readily obvious features of a net, and 4) if an investigator can gain the understanding required to constrain them, she will be empowered to grapple with the other aspects of net design with more confidence. It is then a mere matter of obtaining the net and putting it to good use.

16
Appendix 1 Case Study
Step 1: Organizing Questions Objectives Target Groups
Whale Diets Fin Whales Sei Whales Humpback Whales Killer Whales
Diets of Whale Prey Herring Sardine Pacific Hake Sand Lance
Guidance from Regional Zooplankton Studies Copepods Euphausiids Other Major Taxa
Sizes of Target Groups Target Copepods Target Euphausiids
Study Area Resources Study Design
Step 2: Maximum Mesh Size Step 3: Diameter Step 4: Tow Duration Step 5: Minimum Open-Area Ratio Step 6: Solving for Net Length Scenario 1: 333µ mesh Scenario 2: 200-230µ mesh Decision

17
Step 1: Organizing Questions Objective________________________________________________________________________________ The primary objective of this study, set in the northern fjords of British Columbia, is to document trophic interactions between large cetaceans, primarily fin whales (Balaenoptera physalus) and humpback whales (Megaptera novaeangliae), and their prey. The goals regarding zooplankton are as follows:
1. Monitor seasonal trends in relative density and patchiness of dominant zooplankton (and schooling fish) using echosounder imagery, ground-truthed by vertical tows.
2. Describe geographic patterns in shifts in zooplankton diversity and dominant taxa (e.g. offshore-onshore gradients, aggregation at sills, responses to tidal cycles, etc.).
3. By examining tow and echosounder results in the context of water column samples and predator surveys, describe zooplankton dynamics within the context of both “bottom-up” (environmental) and “top-down” (competition and predation) interactions.
Target Groups____________________________________________________________________________ Target groups for this study were chosen by consulting published studies of cetacean diet and zooplankton ecology from the region. We wanted to prioritize sampling for the species that resident whales were known to feed upon, as well as those species preyed upon by their prey. To summarize that review (below), the primary targets of our study will be Euphausia pacifica, Thysanoessa spinifera (all life stages), and the copepods Neocalanus cristatus, N. plumchrus, N. flemingeri, Calanus marshallae, and Metridia lucens (late naupliar and copepodite stages). Secondary targets included hyperiid amphipods, chaetognaths, and larvaceans. In addition to representing the primary zooplankton prey of resident whales, these taxa are also the dominant taxonomic groups of B.C. waters, monitoring their dynamics would also reflect general zooplankton community dynamics in the study area. Whale Diets Because the scope of our study is focused on trophic interactions, we must first determine which zooplankton taxa are implicated in the foraging ecology of local whales, either directly or by degrees of trophic separation. The best starting point, therefore, is the known diets of locally sampled whales. Fin Whales: Compared to fin whale stomach content data from other regions in the North Pacific, BC fin whales rely less on fish and more on zooplankton for their nutritional needs (Flinn et al. 2002). The primary plankton found in their stomachs was euphausiids (Flinn et al. 2002), but general North Pacific studies indicate that calanoid copepods are also primary prey (Kawamura 1980, 1982; Nemoto 1957, 1959; Nemoto and Kasuya 1965). The stomach contents of BC fin whale were primarily the euphausiids E. pacifica and T. spinifera and the copepod Neocalanus cristatus (Flinn et al. 2002). Kawamura (1982) hypothesized that fin whales in the nearby Gulf of Alaska prey switch from euphausiids (abundant in late spring and early summer) to copepods (most abundant in summer and fall). Other species known to be fin whale prey include the euphausiids T. longipes and T. inermis and the copepods Calanus cristatus, C. plumchrus, C. finmarchicus, and Metridia lucens. Sei Whales: The sei whale (Balaenoptera borealis), a rare offshore balaenopterid that has not yet been seen in British Columbia fjords but could feasibly occur within the study area, is more piscivorous. Stomach content records from the same study indicate that in addition to the above species, sei whales eat hyperiid amphipods, saury (Cololabis saira), walleye Pollock (Theragra chalcogramma), myctophids, and herring (Clupea harengis pallesi)(Flinn et al. 2002). Humpback Whales: Humpbacks in B.C. and Gulf of Alaska waters have been observed feeding upon sardine, herring, capelin, pollock, eulachon, Pacific mackerel (Scomber japonicas), and euphausiids (Nemoto 1959, Fisheries & Oceans Canada 2010). Stomach content records (summarized by Ford et al. 2009) were dominated by the euphausiids E. pacifica and T. spinifera. One stomach was found to contain a species of small squid (Ford et al. 2009). Pacific hake (Merlucius productus) is also dominantly abundant in coastal waters (Mackas et al. 1997) and may be preyed upon by humpbacks. Relative to the specialized diets of other rorquals, humpback

18
whales forage opportunistically (Calkins 1986). From previous experience in the study area, the author can attest that humpback whales are bubble-net feeding intensively on schooling fish in early and mid-summer. They may switch to a krill-dominated diet in the late summer (Janie Wray, pers. comm.). It has also been suggested that humpbacks can prey switch between years (Krieger & Wing 1985). Just north of the study area in Fredericka Sound, AK, the euphausiids T. raschi and E. pacifica constitute 50-80% of humpback diet (Dolphin 1987). T. longipes have also been found in stomachs in the Gulf of Alaska, sometimes hundreds of kilograms of the species (Tomilin 1957, in Russian; cited in Calkins 1986). Chinook Salmon: The main preference of northern resident killer whales within the study area is chinook (spring) salmon (Oncorhynchus tshawytsha; Ford et al. 1998). In British Columbia, Chinook salmon feed opportunistically in estuaries on a range of larvae and zooplankton (Healey 1980) and in open waters on small fishes (21 taxonomic groupings; Pritchard & Tester 1964). This includes herring (especially for large Chinook; Prakash 1962), sand lance (Ammodytes hexapterus) and pilchard (Sardinops sagax caerulea), a subspecies of Pacific sardine (Pritchard & Tester 1964). Invertebrate taxa seem to comprise a negligible percentage of chinook diet (Pritchard & Tester 1964). Diets of Whale Prey It takes a food web to attract large cetaceans to an area, and it would be impossible to design a sampling regime that adequately samples all of the zooplankton (or phytoplankton or fish, for that matter) tied up in that web. But it is nonetheless prudent to take the steps necessary to acknowledge what is being left out of the picture that we hope to paint with our data. Without such precautions, we cannot substantiate claims of which taxa are major “players”, and which it is acceptable not to monitor. In this vein, reviewing which zooplankton are consumed by larger constituents of cetacean diets is the least we can do. Herring: Adult pacific herring feed exclusively on euphausiids (Tanasichuk 1998). Juvenile herring from Prince William Sound were found to prey upon Cirrepedia nauplii, fish eggs, small and large calanoids, euphausiids, and larvaceans (Foy & Norcross 1998). Sardine: The most important prey items for the sardine (Sardinops sagax) in British Columbian waters are diatoms, euphausiids, euphausiid eggs, copepods, and oikopleurids (larvaceans; McFarlane et al. 2005). Pacific Hake: Of all the abundant schooling fish in B.C. waters, hake may exert the greatest predation pressure on their krill prey, Euphausia pacifica and Thysanoessa spinifera, by sheer dint of their biomass (Mackas et al. 1997). Hake and euphausiids are known to co-occur in great aggregations in coastal B.C. throughout summer months. Sand lance: Sand lances (reviewed thoroughly in Robards et al. 1999) are actively pelagic as they feed in the daytime, but they rest in the benthos at night. In the winter, a higher proportion of their diet tends to come from epibenthic prey (Rogers et al. 1979). During their vertical migration, they are preyed upon intensively. Larvae feed on phytoplankton, from diatoms to dinoflagellates (Trumble 1973). When longer than 10 cm, they feed upon the nauplii of copepods in the summer and euphausiids in the winter (Craig 1987). Adults feed predominantly on Calanus copepods, but also on a range of species, including chaetognaths, mysids, amphipods, and fish larvae (Field 1988, O’Connell and Fives 1995, Scott 1973). Regional Literature In addition to our consideration of focal predator diets, our sampling design must account for regionally dominant groups, regardless of their direct involvement in cetacean food webs. Below we outline the major zooplankton groups identified in the study area, as identified by the regional literature. As expected, there is considerable overlap between the primary prey groups identified above. Two dominant groups stand out: copepods and euphausiids (Mackas & Tsuda 1999). Copepods: The most numerically dominant zooplankters of at least the southern fjords of BC are the copepods (Trevorrow et al. 2005). The same goes for the nearby Gulf of Alaska (Cooney 1986). The most common species in BC coastal waters is Neocalanus plumchrus (Harrison et al. 1983). Because this species was split by Miller (1988) into two species, N. plumchrus and the slightly smaller N. flemingeri, studies previous

19
to that must be read with a grain of salt. Other dominant zooplankters include Neocalanus cristatus (another large-bodied copepod in offshore waters and the fringe of BC fjordland), Metridia lucens, and Calanus marshallae (Mackas et al. 2007). Eucalanus bungii is numerically dominant in the Gulf of Alaska (Cooney 1986). Euphausiids: Euphausiids are pelagic shrimp-like eucarids that aggregate in high densities and provide the basis of many trophic webs. Twelve or more euphausiid species occur off the outer B.C. coast (Mackas 1992, Brinton et al. 2000), but the three dominant species there, from the Juan de Fuca area (Mackas 1992) to Prince William Sound (Coyle & Pincuk 2005), are Euphausia pacifica, Thysanoessa spinifera, and T. inspinata (Mackas 1992). All three species are epipelagic, endemic to the North Pacific, and vertical migrators (Bollens et al. 1992). Other Major Taxa: Of course, groups other than these are present in the study area, occasionally in dominant numbers. Those most commonly mentioned in studies of predator diets and regional studies are amphipods, chaetognaths, and tunicates. Hyperiid amphipods: Of the few regional studies that were found (Lorz & Pearcy 1975; Schulenberger 1978; Yamada et al. 2004), sampling protocols were similar to euphausiid studies (571u 1m diameter single-net; 333u 0.7 diameter BONGO; 333u .7 diameter, respectively). In the North Pacific gyre, hyperiid amphipods have been observed to remain primarily within the upper 100m (Schulenberger 1978). In the Oyashio region, 99% of the population was collected above 300m (Yamada et al. 2004). Chaetognaths can also occur in abundance throughout the water column in the study area. They are important predators of larval fish and copepods, among other plankters, and in coastal BC they can be indicative of the excursion of the typically “offshore” assemblage into fjord inlets (see Mackas & Galbraith 2002). These predatory zooplankters are long (2-120mm, Bone et al. 2001) and slender. 333 micron-mesh nets seem adequate for quantitatively sampling all chaetognath life stages (Terazaki & Miller 1986; Johnson & Terazaki 2003). Oikopleurid larvaceans have been observed in high densities at depth in British Columbia fjords (see Trevarrow et al. 2005). The author has seen salps, as well as ctenophores and cnidarians, in abundance at the surface near the mouths of inlets in the study area. Although these gelatinous plankters are not consumed by the target cetaceans of our study, they are a primary prey of herring. Furthermore, their presence suggests shifts in community dynamics, and could be suggestive of a shifting dominance in phytoplankton size class (Andersen 1998). However, because they break up in towed nets, gelatinous plankters are notoriously difficult to sample quantitatively using our approach. At the most, their presence can be noted and identification should be possible to the class-level. Selecting Target Groups What remains now is a close look at each of our target species to determine the minimum size of relevant life stages; this will constrain maximum appropriate mesh size. While length data was readily available for most target species, this author was unable to find reported data on the widths of copepod life stages. Target copepods:
Neocalanus plumchrus & flemingeri Prosome lengths of these species’ copepodites range from 650µ (C1) to 4.9 mm (C5)(Tsuda et al. 1999). Miller et al.’s (1991) 20-year study in the Gulf of Alaska corroborates these data: mean prosome lengths for these species varied between 3.0 and 3.5mm for N. flemingeri adults, and 3.55 and 3.9mm for N. plumchrus C5s.
Neocalanus cristatus Tsuda et al. (2001) reported prosome lengths for C5 N. cristatus between 6.39 and 7.58mm; for C6, between 5.6 and 6.1mm.
Metridia lucens Metridia is a genus of large-bodied copepod, similar in size to Calanus and Neocalanus. Osgood & Frost (1994) observed that nauplii of M. lucens were missed by 216µ mesh but most of their C1s and above are retained.

20
Metridia lucens is a dominant copepod that may be present at any life stage during the summer. In Dabob Bay, southern British Columbia, Metridia lucens does not appear to enter a diapause state (Osgood & Frost 1994). The fall and winter population was chiefly composed of adult females, which remained at depth and were reproductively immature. Nauplii were found on all sampling dates, meaning reproduction never ceased and M. lucens phenology is more continuous than in other major calanoids in the region (Osgood & Frost 1994). Therefore, accounting for the size range of all life stages for this species would be nice, at least to know what will be missed.
Calanus marshallae Osgood & Frost (1994) states that 216µ mesh cannot be quantitative for nauplii of Calanus marshallae and pacificus. Peterson’s (1979) dissertation on C. marshallae relied on 120µ and 240µ mesh nets. Based on his data collection in the laboratory and in the field, he concluded that this species’ first naupliar stage is ~220µ. The third naupliar stage was the first to be longer than 333µ. The first copepodite stage is ~1mm. C5 is 2.5mm, and C6 can be as little as 3.0mm.
Life Stages Because copepod life stages vary in size, it is difficult to sample all stages in the water column, from egg to nauplius to copepodite, quantitavely. Late copepodites of calanoid copepods can be millimeters in length, but their eggs and nauplii are much smaller. Osgood & Frost (1994) observed that 216µ nets missed calanoid nauplii in the North Pacific’s subpolar gyre. To capture copepodites of Calanus, Harris et al. (2000) recommend a mesh size of no more than 124µ. For copepodites of Pseudocalanus, a smaller genus that is also common to BC waters, they recommend 61µ mesh. Some studies (e.g. Coyle & Pincuk 2005, Peterson 1979, Miller & Clemons 1988) deploy two nets of differing mesh size and diameter in order to capture all life stages present; but the primary objectives of these studies pertain to geographic, vertical, and seasonal patterns. Tsuda et al. (2001) used mesh sizes varying from 333µ to 1mm, the largest found from our literature review in studies of N. plumchrus and flemingeri; while they could not quantify naupliar stages, they maintained that “all later copepodite stages were retained”.
Given that our case study is focused on trophic interactions, that the scale of the study limits manpower and time for sampling, and that there are target species other than copepods that must be considered, we will have to accept that only the most relevant life stages of copepods – copepodites (specifically C4, 5, & 6) and, in some cases, the final naupliar stages, will be sampled.
Fortunately, other studies suggest that the timing of our summer sampling plan is ideal. By late May, when sampling would begin, only copepodites 4 and 5 of the dominant Neocalanus plumchrus are expected to be in the study area (Mackas et al. 2007). The biomass maximum for pre-dormant C5’s of this species can be expected to occur somewhere between late April and late May (Mackas et al. 2007), meaning there may not much need to cater equipment design to earlier, smaller life stages.
Target euphausiids:
The following sizes were reported by Brinton et al. (2000), who provide life tables for each species.
Euphausia pacifica Egg capsules can be as small as 360µ. Life stages become progressively large. The final furcicula stage (7) can be as small as 5.3 mm. Adults can be as small as 11 mm.
Thysanoessa spinifera Eggs are as small as 380µ. Adults are as small as 18mm.
Thysanoessa inspinata Eggs and larvae are undescribed. Adults are as small as 12mm.
Study Area_______________________________________________________________________________

21
The study area (Map 1) in northern British Columbia is a complex of fjords that constitutes a broad intracoastal archipelago of steep-walled islands surrounded by deep, protected channels riddled with sills. Broadly, the northwest coast has been an active area of study regarding the influence of oceanographic processes on zooplankton community dynamics (Mackas & Coyle 2005), but no zooplankton survey has yet been conducted in this remote sector of the British Columbian coast. Its waters are productive (“green”). Strong tides and terrigenous sources of freshwater and nutrients result in impressive currents and onshore-offshore gradients in salinity and temperature. Like most fjords, the chemistry of the water column is probably highly structured. Resources_______________________________________________________________________________ Resources for this study are limited. It is a small operation, crewed by three researchers aboard a 37 ft. cutter rig sailboat. Field conditions range from comfortable to near-freezing downpours and thick fog. Thanks to the area’s protected waterways, swell is only of concern at the outermost sampling stations. Tows must be retrieved manually, meaning that we are limited to single-net tows. The vessel has a davit equipped with block and tackle that provides some mechanical advantage, but time and energy are considerably limited. Study Design_____________________________________________________________________________ To design for our goals within our constraints, we will plan for daytime vertical tows with nets that sample as they fall (“plummet nets”, after Heron 1982 and Daly & Macauley 1988). These tows will complement the prey maps generated using a scientific echosounder recording along standard transect lines (Map 2). Infrequent night-time vertical tows will be attempted if possible, but the risks associated with nighttime intracoastal navigation will limit their number. Our sampling of euphausiids, which may avoid less at night, may be compromised as a result, but options here are few. Although not presented here, a design centered around oblique tows was also considered; documentation of the decision to choose vertical sampling is available upon request.
Step 2: Maximum Mesh Size
As there is much to be said for consistency in methods across studies, and much to be learned from peer-reviewed methods of published articles, we scrutinized the methodologies of the zooplankton literature relevant to our case study region and target taxa (Table 2). The following conclusions were drawn: The limiting target taxon will be the copepods. All life stages of euphausiids, amphipods, and chaetognaths can be sampled with 333µ mesh or even larger. This mesh size seems a standard among regional zooplankton studies. Some copepod studies have also used 333µ mesh (Mackas & Anderson 1986, Miller & Clemons 1988, Miller et al. 1991, Tsuda et al. 1999, Tsuda et al. 2001). However, these studies focused only on the late copepodite stages of target species, with the exception of Miller & Clemons (1988) who employed another net with smaller mesh size (70µ) to quantify egg and naupliar stage dynamics. If we wished to collect mid- to late-naupliar stages of our target copepods, if only for descriptive results, nothing larger than a 220µ mesh should be used -- and the smaller the better (after Mackas 1992, Mackas & Galbraith 2002, Peterson 1979, Trevorrow et al. 2005). But, given all the other dimensions of this study that will require attention and the large amount of samples we will accumulate over the season, our focus must be limited to copepodite stages. Our maximum mesh size is therefore 333µ. According to Figure 2, the porosity of 333µ mesh is 46% from both suppliers.
Step 3: Diameter Concerns of avoidance by euphausiids (our primary targets) must govern our diameter decisions. Avoidance is of much less concern in copepods and amphipods. Most of the literature (Table 2) involving euphausiids used net sizes between .6m and 1m in diameter (Fiedler et al. (1998)’s 2.94m net is an exception, but their field work

22
was done from a large oceanographic vessel). The WP-2 small mesozooplankton net is .57m in diameter (Tranter & Fraser 1968), which may be too avoidable for euphausiids. Harris et al. (2000) recommended a diameter of 0.70m in temperate coastal zones (with a 200µ mesh). Given our focus on euphausiids, anything less than .70m would not do. However, above 1m weight, deck space, and drag become prohibitive. A large mouth would also constrain other parameter options in the effort to maximize OAR. The best range for this case study is therefore a diameter of .70m to 1m.
Step 4: Tow Duration Given that tows will contribute most to our objective of sampling diversity, that safety concerns will confine us to daytime sampling, that zooplankton distribution is structured more vertically than horizontally, that many vertically migrating taxa may be at depth during the hours we would be sampling, that time and crew resources are considerably limited, and that other study objectives require attention while the vessel is underway, vertical tows seem most appropriate for our study. In the end, our tow duration will be determined by desired tow distance. The length of towing line is limited by deck space and the fact that it must be retrieved manually. Seafloor depth is not a limiting factor, as the centers of fjord channels in our study area are 600m deep or more. A target sampling distance of 250m would be comparable to published methods, and should sufficiently sample deeper daytime scattering layers of dominant taxa. Therefore, vertical tows will go to a maximum of 250m where possible, which, with a diameter range of .70m to 1.0 m and a 91.4% initial sampling efficiency, would yield volumes between 75 and 200 m3 (Fig. 6a). These values are low compared to oblique tow volumes from CalCOFI and McGowan & Fraundorf (1966), among others, but they meet or exceed published most vertical tow volumes (Table 2).
Step 5: Minimum Open-Area Ratio
Minimum standards for our IFE dictate that MOAR be 3:1 (Tranter & Smith 1968). If we opt for 333µ mesh, we should boost that minimum to 5:1. Our study is in green waters. According to Figure 7, our desired vertical tow duration of 250m requires an OAR of 5.3:1. This will do if we opt for 333µ mesh. If we had used 150-230µ mesh to catch naupliar copepods, our minimum OAR will be set not by tow duration, but by mesh size itself, at 9:1 (Tranter & Smith 1968 and Ohman 2013).
Step 6: Solving for Net Length With known MOAR, porosity and diameter options, we can now constrain the final variable, net length. We are working with 3 diameter options, 0.70m, 0.85m, and 1.0m. Using Figure 8, we are presented with three potential design configurations:
1. 333µ (46% porosity), 0.70m, 5.3:1, 2.80m length. 2. 333µ (46% porosity), 0.85m, 5.3:1, 3.5m. 3. 333µ (46% porosity), 1.0m, 5.3:1, 4.1m.
Decision Given the small size of our research platform and the fact that crew will be retrieving the net manually, option 1, with a length of 2.8 meters, seems the best choice for us. We are now qualified to take the next steps in designing and obtaining our vertical net. Because ours is a plummet net design, whose cylindrical collar will be cinched with a choke line, the collar must be non-porous and cannot contribute to the 2.8m of length needed to achieve an OAR of 5.3:1. The collar will have to be the length of the mouth’s diameter, giving an overall net length of 3.5m (Fig.9, 10).

23
Literature Cited Andersen, V. 1998. Salp and pyrosomid blooms and their importance in biogeochemical cycles, p. 125-137. In Q.
Bone [ed.], The biology of pelagic tunicates. Oxford University Press, Oxford. Bernardi, Riccardo de. 1984. Methods for the estimation of zooplankton abundance. In ____. Pp. 59-86. Bollens, S.M., B.W. Frost, B.W., and T.S. Lin. 1992. Recruitment, growth, and diel vertical migration of
Euphausia pacifica in a temperate fjord. Marine Biology 114: 219–228. Bone, Q., H. Kapp, and A.C. Pierrot-Bults. [ed.] 1991. The biology of chaetognaths. Oxford University Press,
Oxford. p. 1-173. Boyde, S.H., P.H. Weibe, and J.L. Cox. 1978. Limits of Nematoscelis megalops in the Northwest Atlantic in
relation to Gulf Stream cold core rings. II. Physiological and biochemical effects of expatriation. Journal of Marine Research 36:143-159.
Brinton, E. 1962. Variable factors affecting the apparent range and estimated concentration of euphausiids in the North Pacific. Pacific Sci., 16:374-408.
Brinton, E. 1967. Vertical migration and avoidance capability of euphausiids in the California Current. Limnology & Oceanography 12:451-483.
Brinton, E., M.D. Ohman, A.W. Townsend, M.D. Knight, and A.L. Bridgeman. 2000. Euphausiids of the World Ocean. World Diversity Database CD-ROM Series. Windows Version 1.0. ETI Expert Center for Taxonomic Identification: Mauritskade, Netherlands.
Calkins, D.G. 1986. Marine Mammals. In Hood, D.W., and S.T. Zimmerman, eds. 1986. The Gulf of Alaska: physical environment and biological resources. NOAA Alaska Office OCS Study MMS 86-0095.
Clutter, R.I., and M. Anraku. 1968. Avoidance of samplers. In Zooplankton Sampling, p. 57-76. UNESCO Moograph of Oceanographic Metholodies 2.
Cooney, R.T., 1986. Zooplankton. In: Hood, D.W., Zimmerman, S.T. (Eds.). The Gulf of Alaska, Physical Environment and Biological Resource. MMS/NOAA, Alaska Office, Anchorage, OCS Study MMS 86-0095, pp. 285–303.
Coyle, K.O., and A.I. Pinchuk. 2003. Annual cycle of zooplankton abundance, biomass and production on the northern Gulf of Alaska shelf, October 1997 through October 2000. Fisheries Oceanography 12, 327–338.
Coyle, K.O., A.I. Pinchuk. 2005. Seasonal cross-shelf distribution of major zooplankton taxa on the northern Gulf of Alaska shelf relative to water mass properties, species depth preferences and vertical migration behavior. Deep-Sea Resarch II 32:217-245.
Craig, P.C. 1987a. Forage fishes in the shallow waters of the north Aleutian shelf. In: Forage fishes of the southeastern Bering Sea: Proceedings of a conference; [dates of meeting unknown]; [location unknown]. OCS Study MMS 87-0017. [Place of publication unknown]: [publisher unknown]: 49-54.
Currie, R.I. 1963. The Incean Ocean Standard Net. Deep Sea Research 10(1-2), p.27-32. Daly, K. L. and Macaulay, M. C. 1988. Abundance and distribution of krill in the ice edge zone of the Weddell
Sea, austral spring 1983. Deep-Sea Research 35:21-41. Dolphin, W.F. 1987. Prey densities of humpback whales, Megaptera novaengliae. Experientia 43:468-471. Falkenhaug, T., K.S. Tande, T. Semenova. 1997. Diel, seasonal, and ontogenetic variations in the vertical
distributions of four marine copepods. Marine Ecology Progress Series 149:105-119. Fiedler, P.C., S.B. Reilly, R.P. Hewitt, D. Demer, V.A. Philbrick, S. Smith, W. Armstrong, D.A. Croll, B.R. Tershy,
B.R. Mate. 1998. Blue whale habitat and prey in the California Channel Islands. Deep-Sea research II 45:1781-1801.
Field, L.J. 1988. Pacific sand lance, Ammodytes hexapterus, with notes on related Ammodytes species. In: Wilimovsky, N.J.; Incze, L.S.; Westrheim, S.J., eds. Species synopses, life histories of selected fish and shellfish of the northeast Pacific and Bering Sea. [Place of publication unknown]: [publisher unknown]: 15-33. Available from: Washington Sea Grant Program, 3716 Brooklyn Ave. NE, Seattle, WA 98105.
Fisheries and Oceans Canada. 2010. Recovery Strategy for the North Pacific Humpback Whale (Megaptera novaeangliae) in Canada [DRAFT]. Species at Risk Act Recovery Strategy Series. Fisheries and Oceans Canada, Ottawa. x + 51 pp. Fleminger, A., and R.I. Clutter. 1965. Avoidance of towed nets by zooplankton. Limnology & Oceanography 10:96-104.
Fleminger, A., R.I. Clutter. 1965. Avoidance of towed nets by zooplankton. Limnology and Oceanography 10: 96-104.
Flinn, Rowenna, A.W. Trites, E.J. Gregr. 2002. Diets of fin, sei, and sperm whales in British Columbia: an analysis of commercial whaling records, 1963-1967. Marine Mammal Science 18(3):663-679.

24
Ford, J.K.B., A.L. Rambeau, R.M. Abernethy, M.D. Boodaards, L.M. Nichol, and L.D. Spaven. 2009. An assessment of the potential for recovery of humpback whales off the Pacific Coast of Canada. Canadian Science Advisory Secretariat Research Document 2009/015.
Ford, JKB, G.M. Ellis, L.G. Barrett-Lennard, A.B. Morton, R.S. Palm, K.C. Balcomb III. 1998. Dietary specialization in two sympatric populations of killer whales (Orcinus orca) in coastal British Columbia and adjacent waters. Canadian Journal of Zoology, 76(8): 1456-1471.
Foy, R.J., B.L. Norcross. 1998. Spatial and temporal differences in the diet of juvenile Pacific herring (Clupea pallasi) in Prince William Sound, Alaska. In Juvenile Herring Growth and Habitats: Exxon Valdez Oil Spill Restoration Project Annual Report.
Hargreaves PM, Herring PJ (1992) The response of decapod and mysid crustaceans to artificially lighted trawls. J Mar Biol Assoc. UK 72:621–631
Harris, R.P., P.H. Wiebe, J. Lenz, H.R. Skjoldal, M. Huntley, eds. 2000. ICES Zooplankton Methodology Manual. Academic Press: London.
Harrison, P., J. Fulton, F. Taylor, and T. Parsons. 1983. Review of the biological oceanography of the Strait of Georgia: Pelagic environment. Canadian Journal of Fisheries and Aquatic Science 40: 1064-1094.
Healey, M. C. 1980 The ecology of juvenile salmon in Georgia Strait, British Columbia. In Salmonid Ecosystems of the North Pacific (McNeil, W. J. & Hinsworth, D. C., eds). Oregon State University Press, Corvallis. pp. 203-229.
Heron, A.C. 1982. A vertical free fall plankton net with no mouth obstructions. Limnology and Oceanography 27(3):380-383.
Isaacs, J.D. 1965. Larval sardine and anchovy interrelationships. California Cooperative Oceanic Fisheries Invesitgations Report 10: 102-140.
Johnson, T.B. and M. Terazaki. 2003. Species composition and depth distribution of chaetognaths in a Kuroshio warm-core ring and Oyashio water. Journal of Plankton Research 25(10): 1279-1289.
Kawamura, A. 1980. A review of food of balaenopterid whales. Scientific Report of the Whales Research Institute 32:155-198.
Kawamura, A. 1982. Food habits and prey distributions of three rorqual species in the North Pacific Ocean. Scientific Report of the Whales Research Institute 3$:59-91.
Krieger, K.J., and B.L.Wing. 1985. Location and assessment of humpback whale forage in southeastern Alaska with sonar echo sounding. Unpublished report, Auke Bay Laboratory, NOAA. National Marine Fisheries Service, Auke Bay, AK. 66pp.
Laval, P.H. 1974. Un modele mathematique de l’evitement d’un filet a plankton, son application pratique, et sa verification indirecte en recourant au parasitisme de l’amphipode hyperide Vibilia armata Bovallius. Journal of Experimental Marine Biology and Ecology 14:57-87.
Lorz, H.V., & W.G. Pearcy. 1975. Distribution of hyperiid amphipods off the Oregon coast. J. Fisheries Research Board of Canada 32: 1442-1447.
Mackas, D.L. & E.P. Anderson. 1986. Small-scale zooplankton community variability in a northern British Columbian fjord system. Estuarine, Coastal, and Shelf Science 22:115-142.
Mackas, D.L. and K.O. Coyle. 2005. Shelf-offshore exchange processes, and their effects on mesozooplankton biomass and community composition patterns in the northeast Pacific. Deep Sea Research Part II: Topical Studies in Oceanography 52(5-6): 707-725.
Mackas, D.L., 1992. Seasonal cycle of zooplankton off southwestern British Columbia. Canadian Journal of Fisheries and Aquatic Science 49: 903–921.
Mackas, D.L., and A. Tsuda. 1999. Mesozooplankton in the eastern and western subarctic Pacific: Community structure, seasonal life histories, and interannual variability. Progress in Oceanography 43:335-363.
Mackas, D.L., R. Kieser, M. Saunders, D.R. Yelland, R.M. Brown, and D.F. Moore. 1997. Aggregation of euphausiids and Pacific hake (Merluccius productus) along the outer continental shelf off Vancouver Island. Can. J. Fish. Aquat. Sci. 54: 2080–2096.
Mackas, D.L., R.E. Thomson, and M. Galbraith. 2000. Changes in the zooplankton community of the British Columbia continental margin, 1985-1999, and their covariation with oceanographic conditions. Canadian Journal of Fisheries Aquatic Sciences 58:685-702.
Mackas, David L., & M.D. Galbraith. 2002. Zooplankton Distribution and Dynamics in a North Pacific Eddy of Coastal Origin: I. Transport and Loss of Continental Margin Species. Journal of Oceanography 58:725-738.
Mackas, D.L., S. Batten, M. Trudel. 2007. Effects on zooplankton of a warmer ocean: recent evidence from the northeast Pacific. Progress in Oceanography 75: 223-252.

25
McFarlane, G.A., MacDougall, L., Schweigert, J., Hrabok, C. 2005. Distribution and ecology of Pacific sardines (Sardinops sagax) off British Columbia, CA. CalCOFI Report, Vol. 46.
McGowan, J.A., and V.J. Fraundorf. 1966. The relationship between size of net used and estimates of zooplankton diversity. Limnology and Oceanography 11:456-469.
Miller, C.B., B.W. Frost, H.P. Bathelder, M.J. Clemons, R.E. Conway. 1984. Life histories of large, grazing copepods in a subarctic ocean gyre: Neocalanus plumchrus, Neocalanus cristatus, and Eucalanus bungii in the Northeast Pacific. Progress in Oceangraphy 13: 201-243.
Miller, C.B., M.J. Clemons. 1988. Revised life history analysis for large grazing copepods in the subarctic Pacific ocean. Progress in Oceanography 20: 293-313.
Miller, C.B., J. Fulton, B.W. Frost. 1991. Size variation of Neocalanus plumchrus and Neocalanus flemingeri in a 20-yr sample series from the Gulf of Alaska.
Nemoto, T. 1957. Foods of baleen whales in the northern Pacific. Scientific report of the Whales Research Institute 12:33-89.
Nemoto, T. 1959. Food of baleen whales with reference to whale movements. Scientific Report of the Whales Research Institute 14:149-290.
Nemoto,T. and T. Kasuya. 1965. Foods of baleen whales in the Gulf of Alaska of the North Pacific. Scientific Report of the Whales Research Institute 19:45-51.
O’Connell, M.; Fives, J.M. 1995. The biology of the lesser sand-eel Ammodytes tobianus l. In the Galway Bay area. Biology and Environment. 95b(2): 87-98.
Ohman, M.D., P.E. Smith. 1995. A comparison of zooplankton sampling methods in the Cal-COFI time series. CalCOFI Report Vol. 36.
Ohman, M. 2013. Zooplankton. Crustacean field collection and preservation techniques. DeGrave, Sammy, and Joel Martin, eds. Oxford University Press.
Osgood, K.E., and B.W. Frost. 1994. Comparative life histories of three species of planktonic calanoid copepods in Dabob Bay, Washington. Marine Biology 118: 627-636.
Pankhurst, R.C., D.W. Holder. 1952. Wind tunnel technique. London, Pitman. Pearcy, W.G. 1983. Quantitative assessment of the vertical distribution of micronektonic fishes with
opening/closing midwater trawls. Biological Oceanography 2: 289-310. Peterson, W.T. 1979. Life history and ecology of Calanus marshallae Forst in the Oregon Upwelling Zone.
Dissertation at Oregon State University. Prakash, A. 1962. Seasonal changes in feeding of coho and spring (Chinook) salmon in southern British
Columbian waters. J. Fish. Res. Board. Can. 19:851-866. Pritchard, A.L., and A.L. Tester. 1944. Food of spring and coho salmon in British Columbia. Bull. Fish. Res.
Board Can. 65:23p. Robards, M.D., M.F. Wilson, R.H. Armstrong, J.F. Piatt. 1999. Sand Lance: a review of biology and predator
relations and annotaded bibliography. Res. Pap. PNW-RP-521. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 327 p.
Rogers, D.E.; Rabin, D.J.; Rogers, B.J. [and others]. 1979. Seasonal composition and food web relationships of marine organisms in the nearshore zone of Kodiak Island including icthyoplankton, meroplankton (shellfish), zooplankton, and fish. [Seattle, WA]: University of Washington, Fisheries Research Institute, final report FRI-UW-7925: 291 p.
Schulenberger, E. 1978. Vertical distributions, diurnal migrations, and sampling problems of hyperiid amphipods in the North Pacific central gyre. Deep-Sea Research 25: 605-623.
Scott, J.S. 1973. Food and inferred feeding behavior of northern sand lance (Ammodytes dubius). Journal of theFisheries Research Board of Canada. 30: 451-454.
Shaw, W., and Robinson, C.L.K. 1998. Night versus day abundance estimates of zooplankton at two coastal stations in British Columbia, Canada. Mar. Ecol. Prog. Ser. 175: 143–153.
Smith, P.E., R.I. Clutter. 1965. Hydrodynamics of flow and collection in plankton nets. Ocean science and ocean engineering 1: p.515 (abstract).
Smith, P.E., R.C. Counts, and R.I. Cutter. 1968. Changes in filtering efficiency of plankton nets due to clogging under tow. J. Cons. int. Explor. Mer 32(2):232-248.
Swinney GN, Clarke MR, Maddock L (1986) Influence of an electric light on the capture of oceanic deep-sea fish in Biscay. J Mar Biol Assoc UK 66:483–496
Tanasichuk, R. W. 1998. Interannual variations in the population biology and productivity of Thysanoessa spinifera in Barkley Sound, Canada, with special reference to the 1992 and 1993 warm ocean years. Marine Ecology Progress Series 173: 181-195.

26
Terazaki, M., C.B. Miller. 1986. Life history and vertical distribution of pelagic chaetognaths at Ocean Station P in the subarctic Pacific. Deep-Sea Research 33: 323-337.
Tomilin, A.G. 1957. Kitoobraznye [Cetacea]. Vol. 9 of Sveri SSSR I Prilezhaschchikh stran [Mammals of the U.S.S.R. and Adjacent Countries]. Izedatel’stvo Akademi Nauk SSSR, Moscow. 756 pp. (Translated by Israel Program for Scientific Translations, 1967, 717 pp., NTIS No. TT 65-50086.)
Tranter, D.J., A.C. Heron 1967. Experiments on filtration in p[lankton nets. Australian Journal of marine Freshwater Research 18:89-111.
Tranter, D.J., Fraser, J.H., eds. 1968. Zooplankton Sampling. Monographs on Oceanographic Methodology 2. Paris. UNESCO Press.
Tranter, D.J., P.E. Smith. 1968. Filtration Performance. In “Monographs on oceanographic methodology 2: Zooplankton Sampling”, UNESCO, Switzerland.
Trevorrow, Mark V., D.L. Mackas, M.C. Benfield. 2005. Comparison of multifrequency acoustic and in situ measurements of zooplankton abundances in Knight Inlet, British Columbia. Journal of the Acoustical Society of America 117 (6):3574-3588.
Trumble, R.J. 1973. Distribution, relative abundance, and general biology of selected underutilized fishery resources of the eastern north Pacific Ocean. Seattle, WA: University of Washington. 178 p. M.S. thesis.
Tsuda, A., H. Saito, H. Kasai. 1999. Life histories of Neocalanus flemingeri and Neocalanus plumhcrus (Calanoida: Copepoda) in the western subarctic Pacific. Marine Biology 135: 533-544.
Tsuda, A., H. Saito, H. Kasai. 2001. Geographical variation of body size of Neocalanus cristatus, N. plumchrus, and N. flemingeri in the Subarctic Pacific and its marginal seas: implications for the origin of large form of N. flemingeri in the Oyashio area. Progress in Oceanography 57:341-352.
Weibe, P.H., S.H. Boyd, B.M. Davis, and J.L. Cox. 1982. Avoidance of towed nets by the euphausiid Nematoscelis megalops. Fishery Bulletin 80(1).
Wiebe, P.C, and M.H. Benfield. 2003. From the Hensen net toward four-dimensional biological oceanography. Progress in Oceanography 56(1):7-136.
Yamada, Y., T. Ikeda, A. Tsuda. 2004. Comparative life history study on sympatric hyperiid amphipods (Themisto pacifica and T. japonica) in the Oyashio region western North Pacific. Marine Biology 145: 515-527.
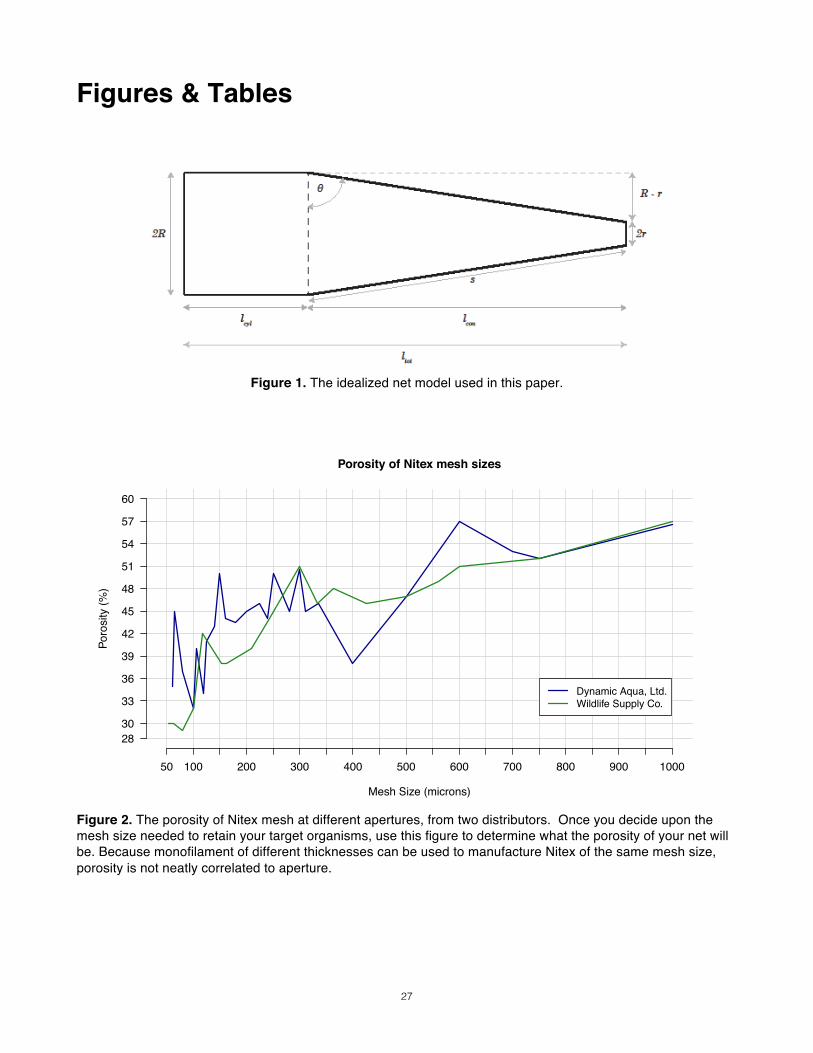
27
Figures & Tables
Figure 1. The idealized net model used in this paper.
Figure 2. The porosity of Nitex mesh at different apertures, from two distributors. Once you decide upon the mesh size needed to retain your target organisms, use this figure to determine what the porosity of your net will be. Because monofilament of different thicknesses can be used to manufacture Nitex of the same mesh size, porosity is not neatly correlated to aperture.
50 100 200 300 400 500 600 700 800 900 1000
2830
33
36
39
42
45
48
51
54
57
60
Mesh Size (microns)
Poro
sity
(%)
Porosity of Nitex mesh sizes
Dynamic Aqua, Ltd.Wildlife Supply Co.
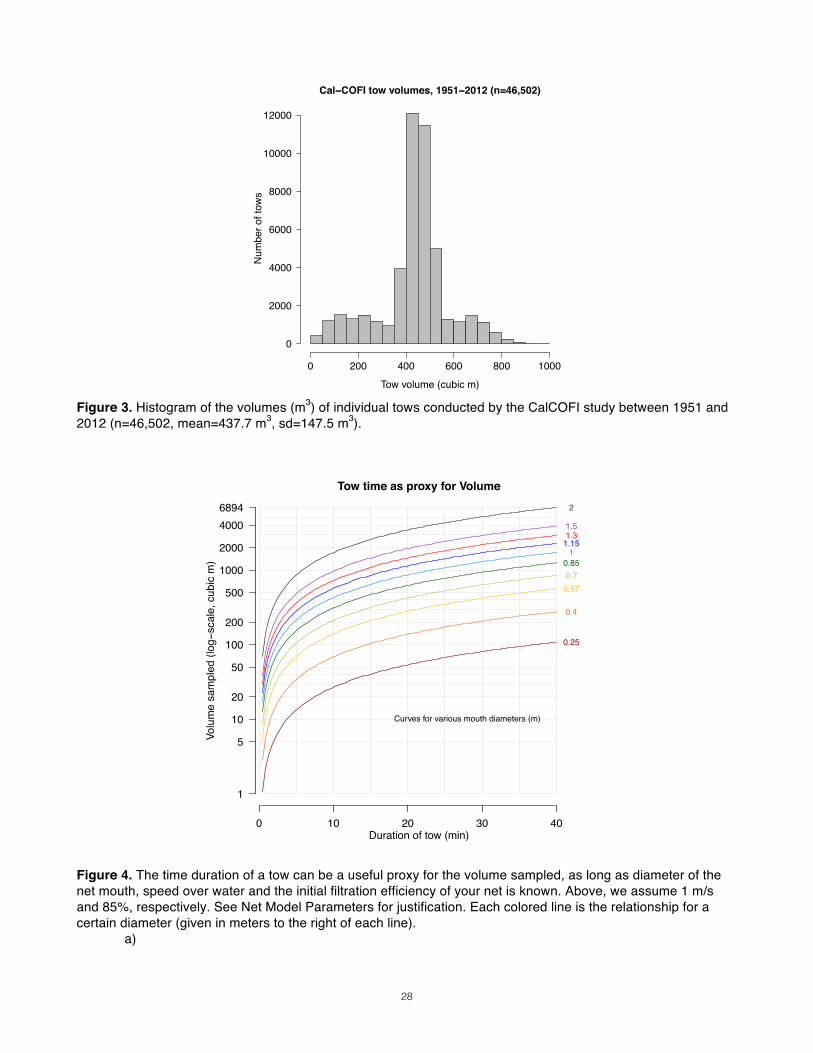
28
Figure 3. Histogram of the volumes (m3) of individual tows conducted by the CalCOFI study between 1951 and 2012 (n=46,502, mean=437.7 m3, sd=147.5 m3).
Figure 4. The time duration of a tow can be a useful proxy for the volume sampled, as long as diameter of the net mouth, speed over water and the initial filtration efficiency of your net is known. Above, we assume 1 m/s and 85%, respectively. See Net Model Parameters for justification. Each colored line is the relationship for a certain diameter (given in meters to the right of each line). a)
0 200 400 600 800 1000
0
2000
4000
6000
8000
10000
12000
Cal−COFI tow volumes, 1951−2012 (n=46,502)
Num
ber o
f tow
s
Tow volume (cubic m)
0.25
0.4
0.570.7
0.851
1.151.31.5
2
Curves for various mouth diameters (m)
0 10 20 30 40
1
5
10
20
50
100
200
500
1000
2000
40006894
Tow time as proxy for Volume
Duration of tow (min)
Volu
me
sam
pled
(log−s
cale
, cub
ic m
)
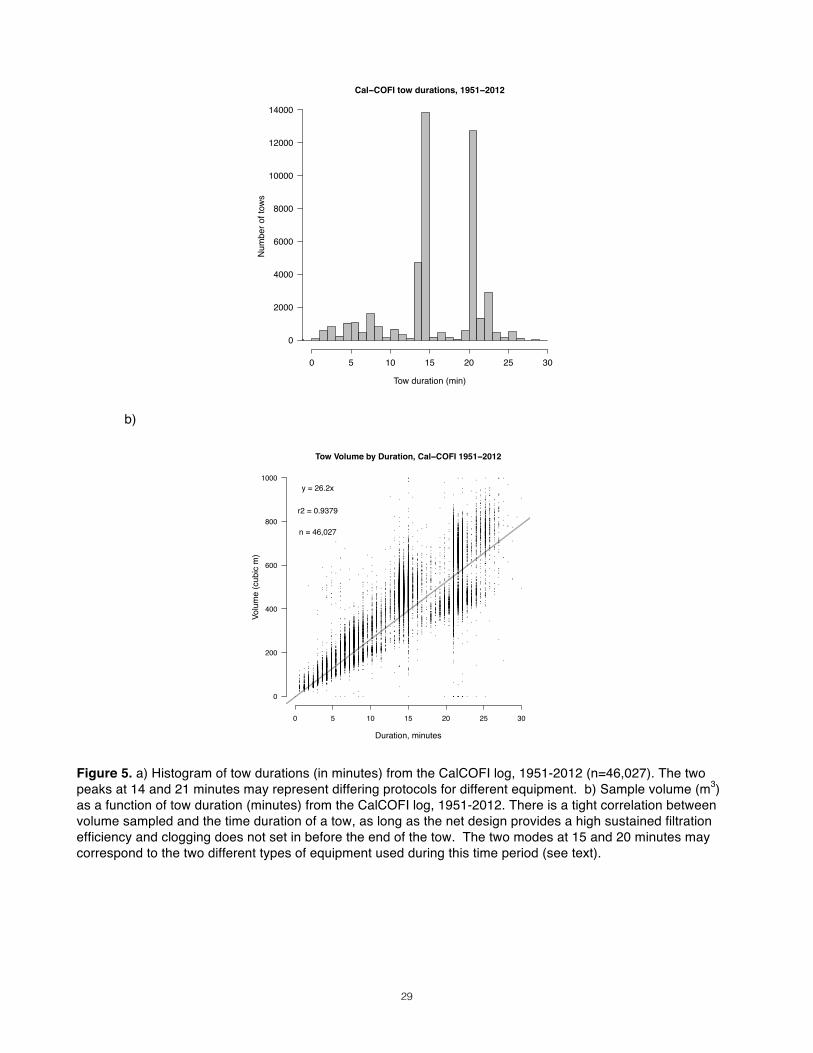
29
b)
Figure 5. a) Histogram of tow durations (in minutes) from the CalCOFI log, 1951-2012 (n=46,027). The two peaks at 14 and 21 minutes may represent differing protocols for different equipment. b) Sample volume (m3) as a function of tow duration (minutes) from the CalCOFI log, 1951-2012. There is a tight correlation between volume sampled and the time duration of a tow, as long as the net design provides a high sustained filtration efficiency and clogging does not set in before the end of the tow. The two modes at 15 and 20 minutes may correspond to the two different types of equipment used during this time period (see text).
0 5 10 15 20 25 30
0
2000
4000
6000
8000
10000
12000
14000
Cal−COFI tow durations, 1951−2012
Num
ber o
f tow
s
Tow duration (min)
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●
● ●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
● ●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●●
●
●
●
●
●●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●● ●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●●
●
●
●
●
●
●
●
● ●
●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●●●●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●● ●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
● ●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●● ●●
●
●
●
●●
●
●●
●
● ●
●●
●
●
●
●
●
●
●
●● ●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●
● ●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●● ●
●
●
●●
●
● ●
●
●
●
●
●
● ●
●
●
●
●●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●●●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●●
●●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
● ●
●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●●●
●●● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●● ●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
● ●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●●●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●
●
●
●
●●
●
●
●
●●
●
● ●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●● ●
●●
●
●
●
●
●●
●
●●
●●
●●
●
●
● ●
●●
●
●
●●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●●
●
●
●●
●●
●
●●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●●
●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●●
●
●
●●
●●●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●● ●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
● ●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
● ●●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
● ●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●● ●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●
●
●
●
●
●
●●
● ●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
● ●
●
●
●
●● ●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●● ●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
● ●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●●●
●
●
●
● ●
●
●
●
●
●
●
●● ●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
● ●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
● ●
●
●●●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
● ● ●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●● ●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●● ●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●●
●●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●●
●●●●
●●
●●
●●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●●
●
●
●
●●
●●
●●●●
●●
●●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●
●●
●●
●
●
●●
●
●●
●●
●
●
●●
●●
●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●
●●
●
●
●●
●●●
●
●
●
●●
●●●●
●●
●
●
●●
●●
●●
●
●●
●●
●●
●●●●
●●●●
●●
●●
●●
●●
●●
●
●●
●
●
●
● ●
●
●●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●●
●
●
●●●●
●
●
●●
●●
●●
●
●
●
●
●
●●
●●
●●
●
●
●
●●
●●
●
●
●
●●
●
●
●●
●●
●●
●●
●●
●
●●●●
●●
●
●●●
●
●
●●
●●
●●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●●●
●
●●
●●
●●
●●
●●
●●
●●
●
●●
●●
●●
●
●
●●
●●
●●
●●
●●●●
●
●●●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●●●●●
●
●
●
●
●
●●
●
●
●●
●
●●
●●
●●
●●
●●
●
●●
●●
●●
●●
●●●●●●
●
●●●●●
●
●●
●
●
●
●●
●● ●●●●
●●
●
●
●●
●●
●●
●●
●●
●
●●
●●
●●
●●
●
●
●●
●
●● ●●
●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●●●●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●●
●●
●●●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●
●●
●●
●●●●
●●
●●
●●●●●●
●●
●●
●●●●
●●
●●
●●
●●
●●●●●●
●●●●●●●●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●●●
●
●
●
●●●
●
●
●
●
●
●●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●●
●●
●●●●
●●
●●
●
●
●
●●
●
●●
●
●
●●●
●
●
●
●
●
●●
●
●●
●
●
●●
●●
●●
●
●
●●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●●
●●
●●
●● ●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●●
●●●
●
●●●●●●
●
●●
●●
●
●
●●
●●
●●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●●●
●●
●
●●
●●
●
●
●
●
●●
●
●
●●
●
●
●●●●
●
●●
●
●●●
●●
●
●●
●
●
●●●●●●
●
●●
●●
●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●●
●
●
●
●
●●
●●●●
●
●
●
●●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●●
●●
●●
●
●●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●●●●●●
●●
●●
●
●
●
●●
●●●
●
●
●
●●
●
●
●●
●●
●●
●
●●●●
●●
●
●
●
●●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●●●
●
●
●●●
●
●●●
●
●●
●
●
●●
●●
●●
●●
●
●●
●
●●
●
●
●●
●
●
●
●●●
●
●
●
●
●●●
●
●●
●
●●
●●
●●
●
●
●
●●●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●●●●●●●●
●●
●●
●●
●●
●●●●
●●●●●●
●●
●●
●●
●
●
●●
●
●
●
●
●●
●●
●●●●
●●
●●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●●●●
●●●●
●
●●
●
●●●●●●●●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●●●●
●
●
●●
●●●●
●●
●
●
●●
●
●●●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●●●
●●●
●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●●●
●●●●●●
●●
●●
●
●
●●
●
●
●●●●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●●●●
●
●
●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●●●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●●
●●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●
●
●●
●
●●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●●●
●●
●●
●●
●●●
●
●●
●●
●● ●●
●
●
●●
●
●
●
●
●●●●●
●
●●
●●●●
●●
●●
●●
●●
●●
●
●
●●
●●●●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●●
●
●
●●
●●
●●
●
●
●●●●
●●●
●
●
●●
●●●
●●
●●
●●
●●
●●
●●●●
●
●●
●
●●
●●
●●
●●
●●
●●
●
●
●●●●
●●●
●
●
●
●●
●●
●●
●●
●●
●● ●●
●
●
●
●
●●
●●
●●●●
●●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●
●
●●
●●
●●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●
●●
●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●●●
●●
●●
●
●
●●
●●
●●
●
●
●
●
●●
●●
●
●●●●●
●●
●●
●●
●●●●●●●●
●●●●●●
●●●●●●●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●●
●
●
●
●●
●
●●
●●
●●●●
●●
●●
●●
●
●
●●
●●
●
●
●●
●
●
●●
●●
●
●
●●
●●
●
●
●●
●
●●●
●●
●
●
●●●
●
●●
●
●
●●●●●●●●
●
●
●
●
●
●
●●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●●
●
●
●●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●●●
●●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●● ●●
●●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●
●
●●
●●
●●●
●●●●●
●●
●●
●●
●●●●
●●
●
●
●
●
●●
●●
●●
●●
●
●●
●
●
●
●●●●
●●
●●
●●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●
●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●● ●●
●●●●
●
●●●
●●
●●
●●
●●
●
●
●●●●●●●●
●●
●●
●●
●●
●
●
●●●●
●
●
●●
●●
●●
●
●●●
●●
●●
●
●
●●
●
●
●●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●●
●
●
●●
●●
●
●●●
●
●
●●●●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●●
●●
●●
●●
●●
●
●
●●●●
●●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●●
●●●
●
●●
●
●
●
● ●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●● ●●
●●
●●
●●●●
●●
●●
●
●
●●
●●
●●
●●●●
●
●
●●
●
●
●●
●●
●●
●
●
●●
●
●
●●
●●
●●
●●
●●
●●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●●
●●
●●●●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●●
●●
●
●
●
●
●
●●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●●
●
●●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●●●●●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●●●●●
●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●●
●●●
●
●
●
●
●
●
●
●●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●●
●●
●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●●
●●
●●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●●
●
●●
●
●
●
●
●●
●● ●
●
●●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●
●
●●●● ●
●
●●●
●
●●
●
●
●
●●
●
●
● ●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●●●●●
●
●
●●
●●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●●● ●●
●
●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●●
●●
●●●
●
●●●●
●● ●
●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●
●
●
●●
●●●
●●
●
●●
●
●
●
●
●
●
●
●●
●●●
●
●●
●●
●●●●●●
●●
●
●
●●●●
●
●●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●
●●
●●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
● ●
●
●●
●
●
●
●
●●●●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●●●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●●●
●
●●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●●
●●●
●●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●●●●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●● ● ●●●
●
●
● ●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●● ●●●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●●
●
●
●
●
● ●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●●●
●●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●●●
●
●●
●●
●●
●●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●●●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
● ●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●●
●●
●
●
●●●
● ●●
●●
●
●
●
●●
●
●
●●●
●
●●●●
●●●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●●
●
●●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●●●
●
●
●
●
●
●
●●●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●●●
●●
●●
●●
●●●
●
●
●
●●
●
●
●●
●
●●●●●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●● ●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●
●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●
●●●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●
●●●
●●
●
●
●●●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●●●
●●●●
●● ●●
●●●●
●●
●●
●
●
●
●
●●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●
●
●●●
●
●
●
●
●●
●
●●
●●
●●●●
●●●●
●●●●
●●
●●
●●
●●
●●
●●
●
●●●
●
●
●
●
●
●●●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●●●●
●●
●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●●●
●
●
●
●●
●
●●
●●
●
●●●
●●
●●●●●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●●●●
●●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●
●●●●
●●
●●
●●
●
●●
●
●●
●
●
●● ●
●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●●●●
●●
●●
●●
●●
●●●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●
●
●
●
●●
●●
●
●
●
●
●●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●●●
●●
●●
●
●
●●
●●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●
●●
●●
●
●
●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●
●●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●●●
●●
●
●
●●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
● ●
●
●
●
●
●
●
●●
●
●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●
●
●
●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●●●
●●
●●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●●●
●
●
●
●●
●
●
●
●●
●●●●
●●●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●
●●
●●
●
●
●●
●
●
●
●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●●
●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●●
●
●
●●●
●
●
●
●
●
●
●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●●
●
●
●●
●●
●●
●
●●
●●
●●●●●●●●
●●
●
●●●
●
●
●
●
●
●
●
●
●●
●●●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●
●●●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●
●●●●●
●
●●
●●
●
●●
●
●●
●
●
●
●●
●
●●
●●●●
●●
●●
●●
●●
●
●
●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●●●
●●
●●●●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●●
●●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●●●●●
●●●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●●
●
●
●
●
●●
●
●●●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●●
●
●
●
●
●●
●
●
●●●●
●●●●
●●
●●
●●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●●●●
●●
●●
●●
●●●●
●
●
●●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●
●●
●●
●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
● ●
●
●
●
●●
●
●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●●
●
●
●●
●●
●●
●● ●●●●
●●
●●●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●●
●●
●●
●●
●●
●●●●●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●
●
●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●●●
●●
●●●●
●●
●●●●
●
●
●
●
●●
●●
●●
●●
●●
●●●●
●●●
●●
●
●
●
●●
●●
●●
●●
●
●
●●
●●
●
●
●●●●●●
●●
●
●●●
●●●●
●●
●
●
●
●●
●●
●●
●●
●
●
●●
●●●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●
●●●●
●
●●
●●
●●
●●
●●
●● ●●
●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●
●●
●
●
●●●
●●
●●●●
●●
●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
● ●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●
●●
●●●●
●
●
●
●
●
●
●●
●●
●●●
●●
●●
●●
●●●●●●
●●
●●●●●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●
●
●
●●
●●●
●●
●●
●●
●●
●●
●●●●
●
●●
●●●●
●●●●
●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●●
●●
●●
●●
●●
●●●●
●●
●●●
●
●●
●●
●●
●●
●●
●●●●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●●
●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●
●●
●●
●
●●●
●
●
●
●
●●●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●●
●●●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●
● ●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●●●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●●●
●
●
●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●
●
●●●●
●●
●●
●●
●●
●●
●●●●
●●●
●
●●
●●
●●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●●●●
●●●●●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●●
●
●
●
●●
●
●
●●
●●
●●
●
●●●
●●
●●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●● ●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●
●
●●
●
●
●●●
●
●
●
●●
●
●
●
●
●●
●●●
●
●
●
●●
●
●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●●●●
●
●
●●
●●
●●
●●
●●●●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●●
●
●
●
●●
●
●●●
●●
●
●
●●
●
●
●
●●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●●●●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●●
●
●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●● ●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●●
●●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●●
●
●
●
● ●●
●●
●
●
●●
●
●
●●
●
●
●
●
●
● ●
●
●
●
●
●●
●
●●
●●
●●
●●
●
●
●●
●●●●●●
●●
●
●●
●
●
●
●●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●
●
●
●
● ●
●
●
●
●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●
● ●
●
●●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●●●●●
●●●●
●●
●●
●
●●●
●●
●●
●●
●●
●●●●●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●●●●●●
●●●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●
●●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
● ●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●●●
●●
●●
●●
●●
●●●●
●●
●●
●●●●
●
●
●
●
●● ●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●●
●●
●●
●●
●●
●●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●●●
●
●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●
●●
●●
●●
●
●
●●
●●
●
●
●
●
●●
●●
●
●
●●
●
●
●●
●●●●
●●
●●
●●
●●●●●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●●●●
●
●●●
●●●●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●
●●●●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●●●
●
●
●
●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●
●
●
●
●●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●●
●
●
●
●
●●●
●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●●●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●●●
●
●●
●●
●●
●●●●
●●●●
●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●
●
● ●
●
●
●
●●
●●●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●●
●●
●●●●●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●●●
●●
●●
●●
●●
●●
●●●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●
●●
●●
●●
●●
●●
●●
●● ●●
●●
●●
●●
●
●
●●
●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●
●
●
●
●
●●
●●
●●
●●
●●●●
●●
●●
●
●
●
●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●●●●●●●
●●
●●
●●
●●
●●
●●
●●●●
●●
●●
●●
●
●●●
●
y = 26.2x
r2 = 0.9379
n = 46,027
0 5 10 15 20 25 30
0
200
400
600
800
1000
Tow Volume by Duration, Cal−COFI 1951−2012
Duration, minutes
Volu
me
(cub
ic m
)
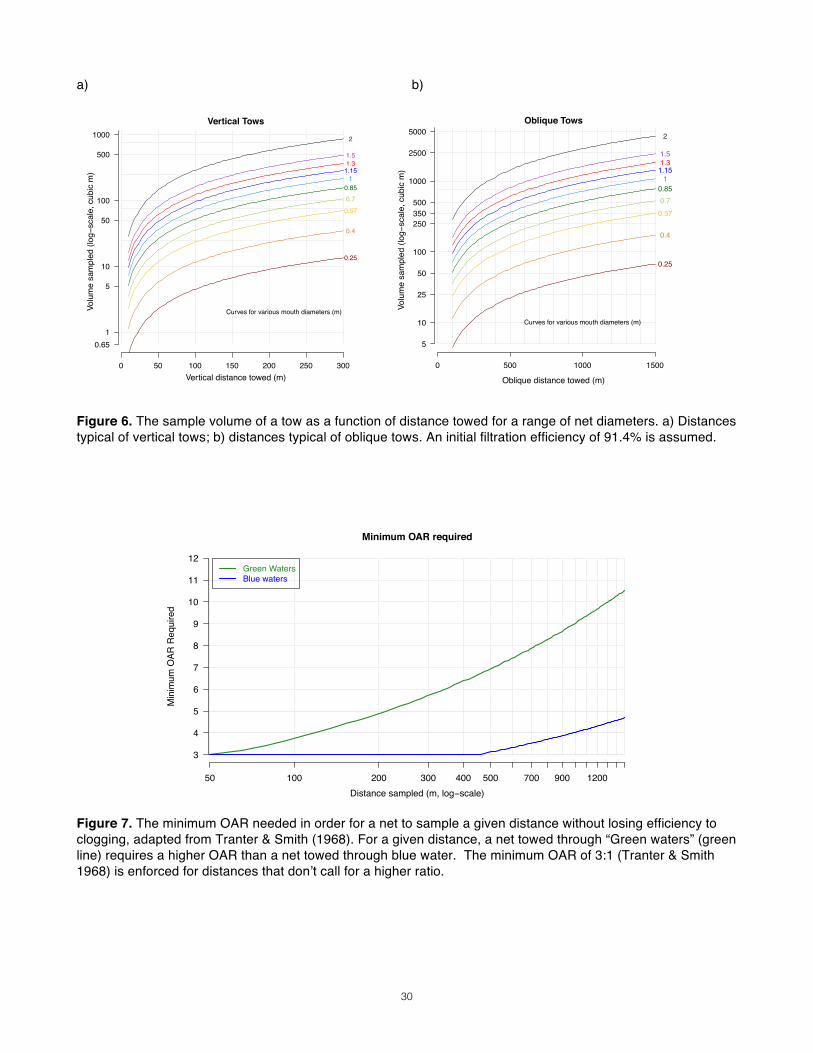
30
a) b)
Figure 6. The sample volume of a tow as a function of distance towed for a range of net diameters. a) Distances typical of vertical tows; b) distances typical of oblique tows. An initial filtration efficiency of 91.4% is assumed.
Figure 7. The minimum OAR needed in order for a net to sample a given distance without losing efficiency to clogging, adapted from Tranter & Smith (1968). For a given distance, a net towed through “Green waters” (green line) requires a higher OAR than a net towed through blue water. The minimum OAR of 3:1 (Tranter & Smith 1968) is enforced for distances that don’t call for a higher ratio.
0.25
0.4
0.570.7
0.851
1.151.31.5
2
Curves for various mouth diameters (m)
0 50 100 150 200 250 300
0.651
5
10
50
100
500
1000Vertical Tows
Vertical distance towed (m)
Volu
me
sam
pled
(log−s
cale
, cub
ic m
)
0.25
0.4
0.570.7
0.851
1.151.31.5
2
Curves for various mouth diameters (m)
0 500 1000 1500
5
10
25
50
100
250350500
1000
2500
5000Oblique Tows
Oblique distance towed (m)
Volu
me
sam
pled
(log−s
cale
, cub
ic m
)
50 100 200 300 400 500 700 900 1200
3
4
5
6
7
8
9
10
11
12
Minimum OAR required
Distance sampled (m, log−scale)
Min
imum
OAR
Req
uire
d
Green WatersBlue waters
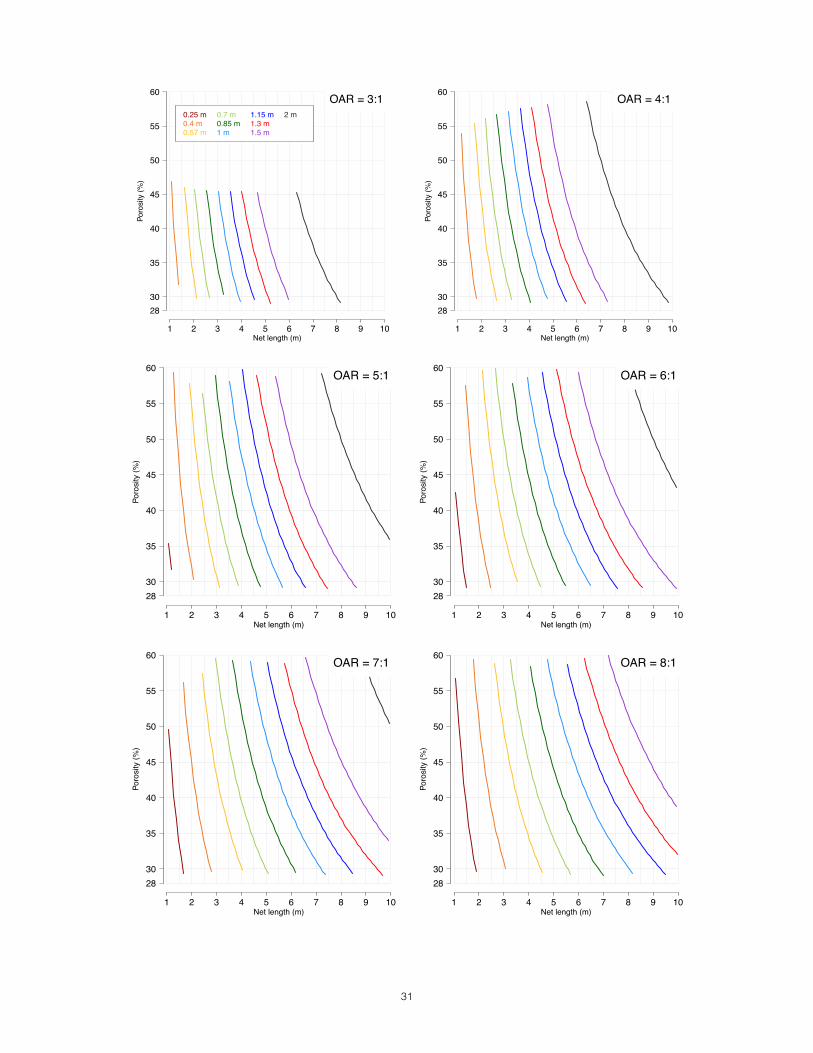
31
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 3:1
Net length (m)
Poro
sity
(%)
0.25 m0.4 m0.57 m
0.7 m0.85 m1 m
1.15 m1.3 m1.5 m
2 m
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 4:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 5:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 6:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 7:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 8:1
Net length (m)
Poro
sity
(%)
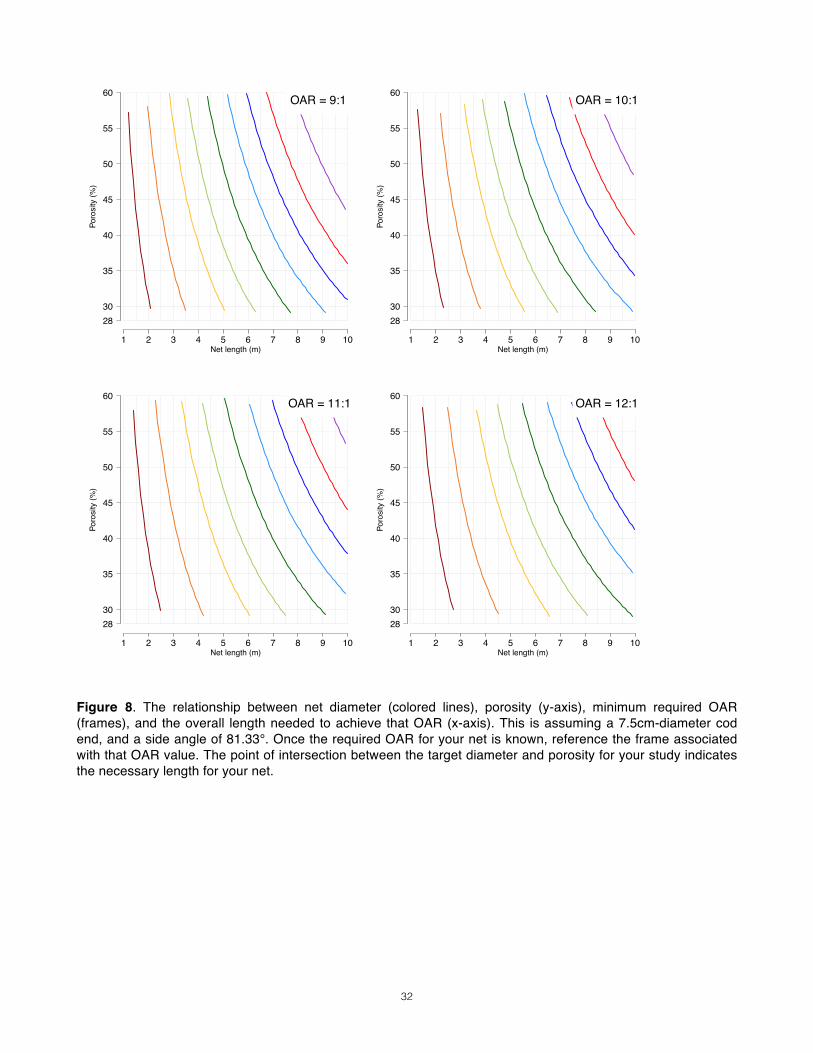
32
Figure 8. The relationship between net diameter (colored lines), porosity (y-axis), minimum required OAR (frames), and the overall length needed to achieve that OAR (x-axis). This is assuming a 7.5cm-diameter cod end, and a side angle of 81.33°. Once the required OAR for your net is known, reference the frame associated with that OAR value. The point of intersection between the target diameter and porosity for your study indicates the necessary length for your net.
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 9:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 10:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 11:1
Net length (m)
Poro
sity
(%)
1 2 3 4 5 6 7 8 9 10
2830
35
40
45
50
55
60 OAR = 12:1
Net length (m)
Poro
sity
(%)

33
Figure 9. The “Heron-Bangarang” sampler in various stages of storage and deployment. This is the final net design for the case study, adapted from Heron (1982). 0.7m-diameter, solid ¾” 316 stainless steel ring with welded eye bolts;; Black Dacron cylindrical collar (.7m length) with chokeline led through caribeners from fore to aft borders of the collar; 2.7m conical section with 333 micron mesh; 7.5cm cod-end.
Figure 10. Realization of the “Heron-Bangarang” design in Fig. 9. Author for scale.
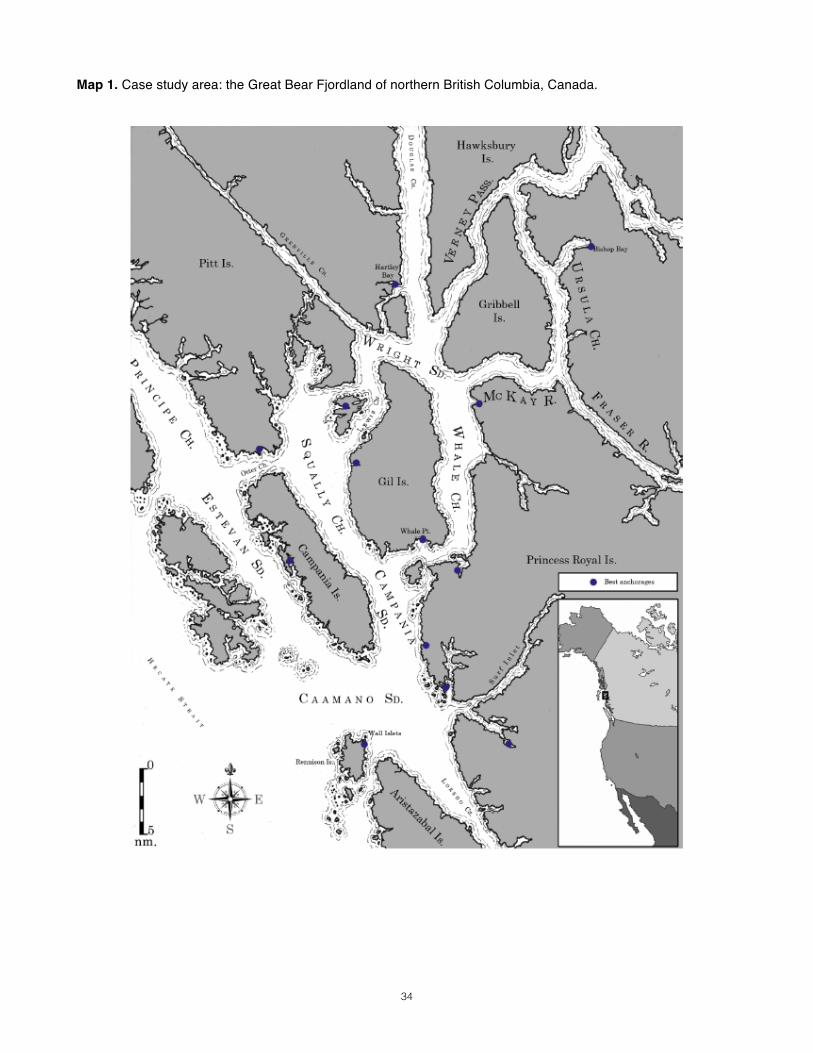
34
Map 1. Case study area: the Great Bear Fjordland of northern British Columbia, Canada.
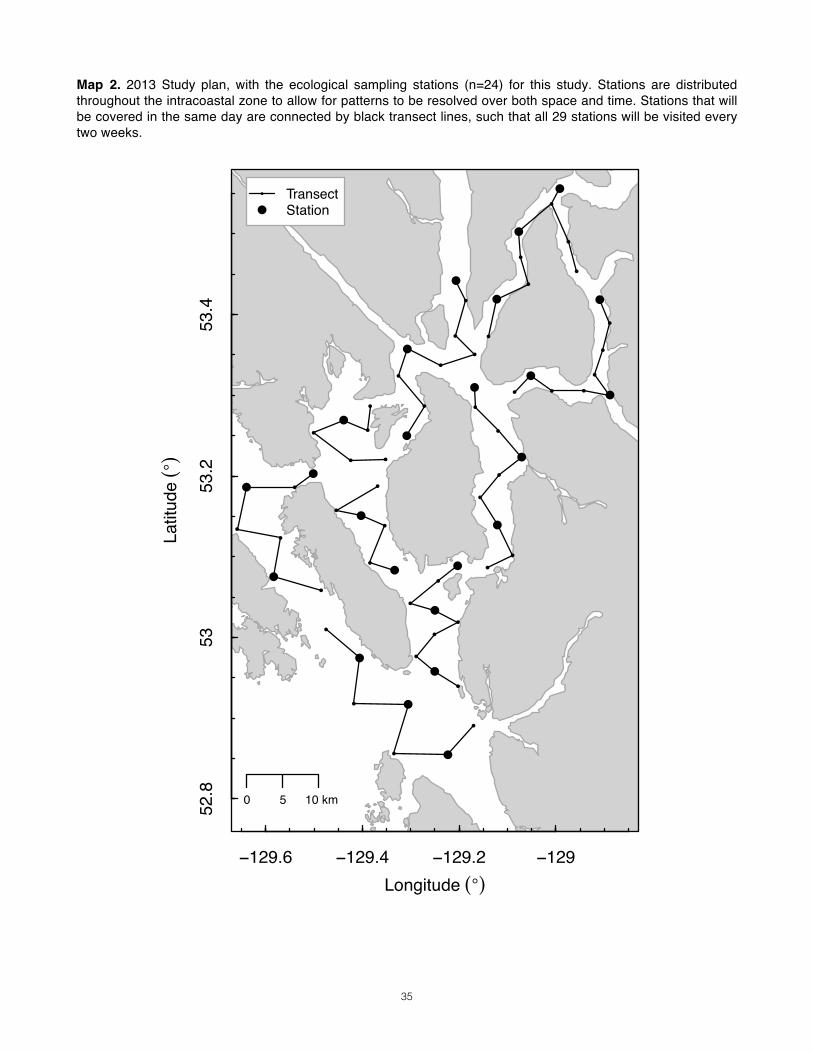
35
Map 2. 2013 Study plan, with the ecological sampling stations (n=24) for this study. Stations are distributed throughout the intracoastal zone to allow for patterns to be resolved over both space and time. Stations that will be covered in the same day are connected by black transect lines, such that all 29 stations will be visited every two weeks.
−129.6 −129.4 −129.2 −129
52.8
5353
.253
.4
Longitude (°)
Latit
ude
(°)
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
●
● ●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
TransectStation
0 5 10 km
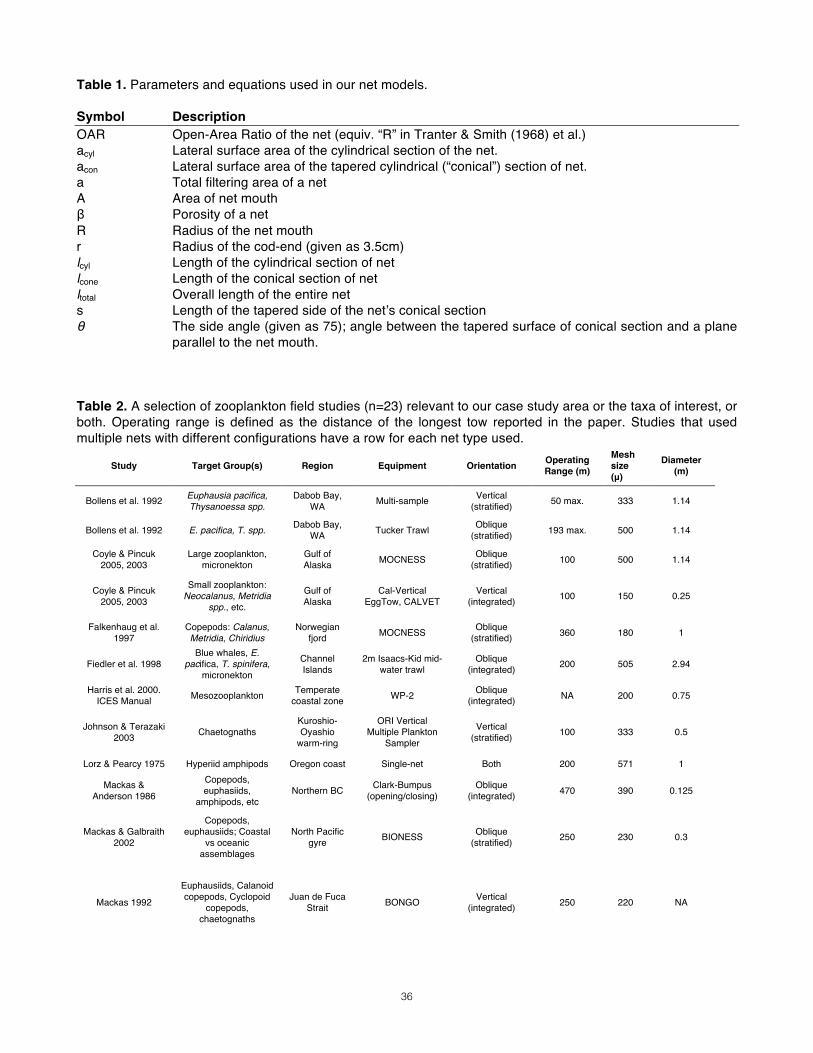
36
Table 1. Parameters and equations used in our net models. Symbol Description OAR Open-Area Ratio of the net (equiv. “R” in Tranter & Smith (1968) et al.) acyl Lateral surface area of the cylindrical section of the net. acon Lateral surface area of the tapered cylindrical (“conical”) section of net. a Total filtering area of a net A Area of net mouth β Porosity of a net R Radius of the net mouth r Radius of the cod-end (given as 3.5cm) lcyl Length of the cylindrical section of net lcone Length of the conical section of net ltotal Overall length of the entire net s Length of the tapered side of the net’s conical section θ The side angle (given as 75); angle between the tapered surface of conical section and a plane
parallel to the net mouth. Table 2. A selection of zooplankton field studies (n=23) relevant to our case study area or the taxa of interest, or both. Operating range is defined as the distance of the longest tow reported in the paper. Studies that used multiple nets with different configurations have a row for each net type used.
Study Target Group(s) Region Equipment Orientation Operating Range (m)
Mesh size (μ)
Diameter (m)
Bollens et al. 1992 Euphausia pacifica, Thysanoessa spp.
Dabob Bay, WA Multi-sample Vertical
(stratified) 50 max. 333 1.14
Bollens et al. 1992 E. pacifica, T. spp. Dabob Bay, WA Tucker Trawl Oblique
(stratified) 193 max. 500 1.14
Coyle & Pincuk 2005, 2003
Large zooplankton, micronekton
Gulf of Alaska MOCNESS Oblique
(stratified) 100 500 1.14
Coyle & Pincuk 2005, 2003
Small zooplankton: Neocalanus, Metridia
spp., etc.
Gulf of Alaska
Cal-Vertical EggTow, CALVET
Vertical (integrated) 100 150 0.25
Falkenhaug et al. 1997
Copepods: Calanus, Metridia, Chiridius
Norwegian fjord MOCNESS Oblique
(stratified) 360 180 1
Fiedler et al. 1998 Blue whales, E.
pacifica, T. spinifera, micronekton
Channel Islands
2m Isaacs-Kid mid-water trawl
Oblique (integrated) 200 505 2.94
Harris et al. 2000. ICES Manual Mesozooplankton Temperate
coastal zone WP-2 Oblique (integrated) NA 200 0.75
Johnson & Terazaki 2003 Chaetognaths
Kuroshio-Oyashio
warm-ring
ORI Vertical Multiple Plankton
Sampler
Vertical (stratified) 100 333 0.5
Lorz & Pearcy 1975 Hyperiid amphipods Oregon coast Single-net Both 200 571 1
Mackas & Anderson 1986
Copepods, euphasiids,
amphipods, etc Northern BC Clark-Bumpus
(opening/closing) Oblique
(integrated) 470 390 0.125
Mackas & Galbraith 2002
Copepods, euphausiids; Coastal
vs oceanic assemblages
North Pacific gyre BIONESS Oblique
(stratified) 250 230 0.3
Mackas 1992
Euphausiids, Calanoid copepods, Cyclopoid
copepods, chaetognaths
Juan de Fuca Strait BONGO Vertical
(integrated) 250 220 NA
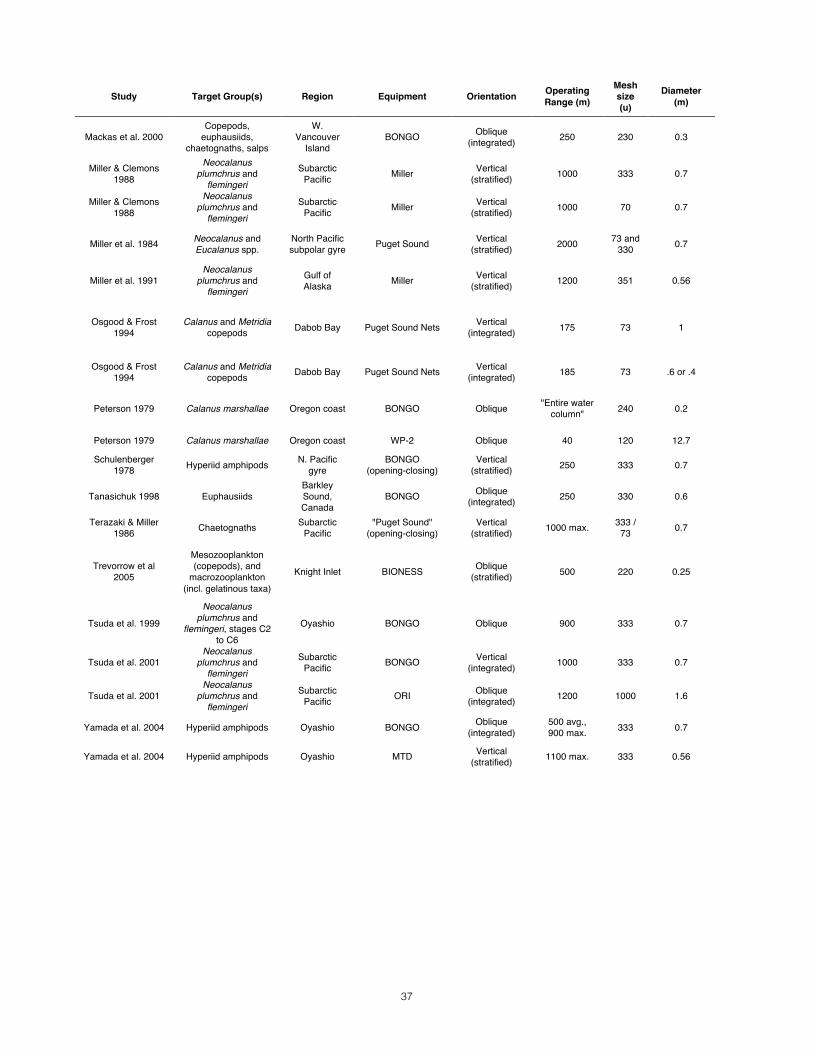
37
Study Target Group(s) Region Equipment Orientation Operating Range (m)
Mesh size (u)
Diameter (m)
Mackas et al. 2000 Copepods,
euphausiids, chaetognaths, salps
W. Vancouver
Island BONGO Oblique
(integrated) 250 230 0.3
Miller & Clemons 1988
Neocalanus plumchrus and
flemingeri
Subarctic Pacific Miller Vertical
(stratified) 1000 333 0.7
Miller & Clemons 1988
Neocalanus plumchrus and
flemingeri
Subarctic Pacific Miller Vertical
(stratified) 1000 70 0.7
Miller et al. 1984 Neocalanus and Eucalanus spp.
North Pacific subpolar gyre Puget Sound Vertical
(stratified) 2000 73 and 330 0.7
Miller et al. 1991 Neocalanus
plumchrus and flemingeri
Gulf of Alaska Miller Vertical
(stratified) 1200 351 0.56
Osgood & Frost 1994
Calanus and Metridia copepods Dabob Bay Puget Sound Nets Vertical
(integrated) 175 73 1
Osgood & Frost 1994
Calanus and Metridia copepods Dabob Bay Puget Sound Nets Vertical
(integrated) 185 73 .6 or .4
Peterson 1979 Calanus marshallae Oregon coast BONGO Oblique "Entire water column" 240 0.2
Peterson 1979 Calanus marshallae Oregon coast WP-2 Oblique 40 120 12.7
Schulenberger 1978 Hyperiid amphipods N. Pacific
gyre BONGO
(opening-closing) Vertical
(stratified) 250 333 0.7
Tanasichuk 1998 Euphausiids Barkley Sound, Canada
BONGO Oblique (integrated) 250 330 0.6
Terazaki & Miller 1986 Chaetognaths Subarctic
Pacific "Puget Sound"
(opening-closing) Vertical
(stratified) 1000 max. 333 / 73 0.7
Trevorrow et al 2005
Mesozooplankton (copepods), and
macrozooplankton (incl. gelatinous taxa)
Knight Inlet BIONESS Oblique (stratified) 500 220 0.25
Tsuda et al. 1999
Neocalanus plumchrus and
flemingeri, stages C2 to C6
Oyashio BONGO Oblique 900 333 0.7
Tsuda et al. 2001 Neocalanus
plumchrus and flemingeri
Subarctic Pacific BONGO Vertical
(integrated) 1000 333 0.7
Tsuda et al. 2001 Neocalanus
plumchrus and flemingeri
Subarctic Pacific ORI Oblique
(integrated) 1200 1000 1.6
Yamada et al. 2004 Hyperiid amphipods Oyashio BONGO Oblique (integrated)
500 avg., 900 max. 333 0.7
Yamada et al. 2004 Hyperiid amphipods Oyashio MTD Vertical (stratified) 1100 max. 333 0.56





























