Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Mar. 1985, p. 774-779 Vol. 47, No. 30019-9567/85/030774-06$02.00/0Copyright © 1985, American Society for Microbiology
Effects of Iron and Desferrioxamine on Infections with Yersiniaenterocolitica
ROY M. ROBINS-BROWNE* AND J. KAYA PRPIC
Department of Microbiology, University of Melbourne, Parkville, Victoria 3052, Australia
Received 20 November 1984/Accepted 6 December 1984
The effects of iron-dextran and the iron chelator desferrioxamine B mesylate (Desferal) on the course andoutcome of experimental yersiniosis were investigated. Yersinia enterocolitica strains representing the threeleading serogroups pathogenic for humans, 03, 08 and 09, were studied. In mice, iron-dextran reduced themedian lethal dose of intraperitoneally administered Y. enterocolitica 03 and 09 ca. 10-fold, whereas besferalreduced this value more than 100,000-fold. Experiments in which Y. enterocolitica was given orally to mice andintraconjunctivally to guinea pigs confirmed that Desferal markedly increased the susceptibility of animals toyersiniosis. Although serogroup 08 yersiniae were inherently more virulent for laboratory animals, they wereless affected by Desferal than were 03 or 09 strains. In vitro experiments indicated that Desferal promotedgrowth of Y. enterocolitica under iron-limiting conditions and suggested that the enhanced vitulence of 08yersiniae may be due to their comparatively low requirement for iron. The adverse effect of Desferal on thecourse of experimental infection with Y. enterocolitica may partly explain the heightened susceptibility ofiron-overloaded patients to systemic yersiniosis.
Iron is an essential growth factor for nearly all bacteria (8,21, 33). The concentration of iron required is generally ca.0.4 to 4 ,uM, an amount readily provided by many environ-ments, including most bacteriological culture media (33). Inmammalian tissues, however, almost all iron is complexedwith other molecules or is tightly bound to carrier proteins,notably transferrin, in serum, and lactoferrin, in mucoussecretions and the granules of polymorphonuclear leuko-cytes (5, 8, 33).To obtain iron under conditions of iron starvation, nmany
bacteria release low-molecular-weight, high-affinity iron che-lators, known collectively as siderophores (21). These ligandsbind and solubilize ferric iron before re-entering the bacterialcell via specific membrane receptors. Siderophores occur intwo broad chemical classes, hydroxanfates and catechols, atleast one of which has been found in almost every aerobic orfacultatively anaerobic bacterial species examined (21).
Desferrioxamine B is a trihydroxamate siderophore ob-tained from Streptomyces pilosus. It is a licensed therapeuticsubstance for use in patients with iron overload resultingfrom acute poisoning or repeated administration in the formof blood transfusions (10, 20).
Because siderophores permit bacterial multiplication inanimals, they may be regarded as virulence factors (30).There are pathogenic bacteria, however, in which sidero-phores have not been detected. One of these is Yersiniaenterocolitica, an important cause of enteritis and mesen-teric adenitis (2, 22). Systemic involvement is unusual withY. enterocolitica, and most patients with this complicationhave an underlying disease, commonly cirrhosis or ironoverload (3, 6, 25, 27, 29). The close relationship betweensystemic yersiniosis and hemosiderosis is believed to reflectthe difficulty Y. enterocolitica has in obtaining iron fromtissues. However, in view of the fact that many patients withiron overload receive desferrioxamine, Melby et al. (19)have suggested that this substance, a proven growth factorfor Y. enterocolitica in vitro (4, 22), may contribute to thedevelopment of systemic yersiniosis. To investigate this
* Corresponding author.
possibility, we examined the influence of iron and desferri-oxamine B, alone and in combination, on the course andoutcome of experimental infections with Y. enterocolitica inmice and guinea pigs. Our findings indicate that desferri-oxamine B markedly increases the susceptibility of animalsto yersiniosis.
(This work was presented in part at the 24th InterscienceConference on Antimicrobial Agents and Chemotherapy, 8to 10 October 1984, Washington, D.C.)
MATERIALS AND METHODS
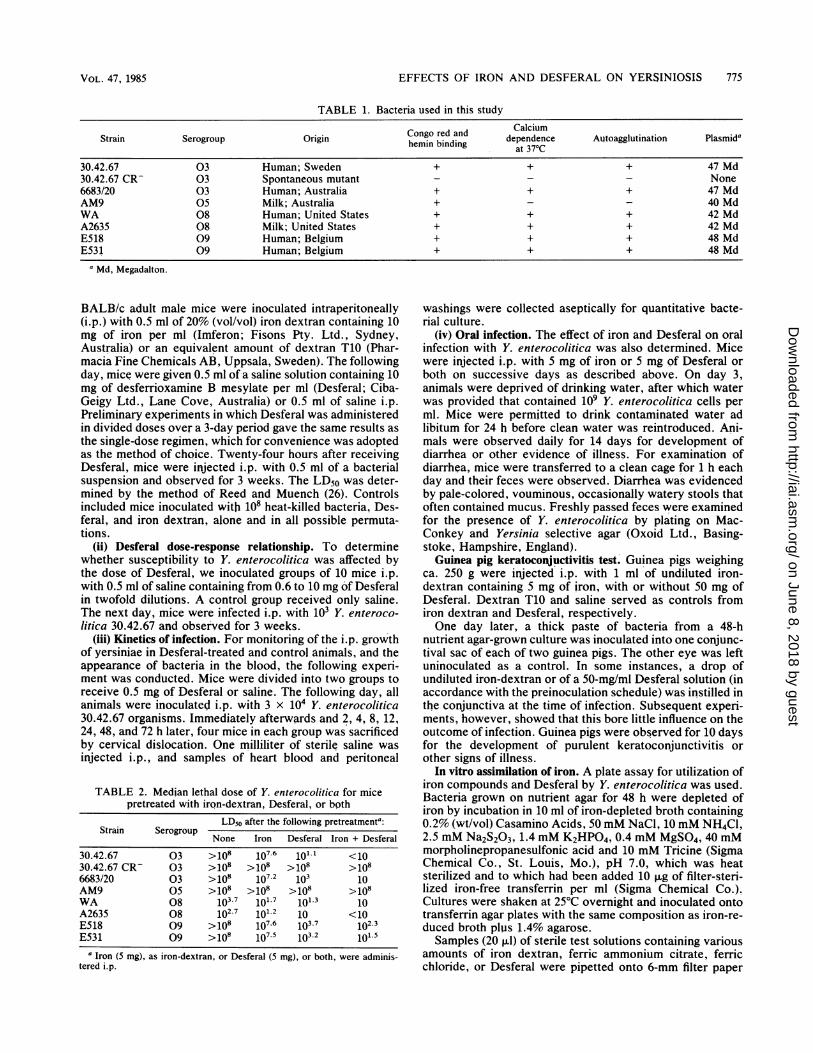
Bacteria. Eight Y. enterocolitica strains were examined.Six comprised two representatives of each of the majorpathogenic serogroups: 03, 08, and 09 (2). One strain,30.42.67 CR-, was a spontaneously occurring, plasmidless,avirulent derivative of a clinical isolate, and the eighth was aserogroup 05 strain obtained from milk (12). Each strain wasexamined for the following virulence-associated attributes asdescribed previously (24): Congo red and hemin binding,calcium dependence at 37°C, autoagglutination, and plasmidcarriage. The characteristics of the bacteria are summarizedin Table 1.
Before animal pathogenicity studies, bacteria were culti-vated at 25°C for 48 h on Congo red acid morpholinepropane-sulfonic acid pigmentation (CRAMP) agar containing 50 ,ugof Congo red per ml (24). Ten to 12 pigmented colonies (or,in the case of Y. enterocolitica 30.42.67 CR-, nonpigmentedcolonies) were selected and plated on nutrient agar. Plateswere incubated at 25°C for 48 h, after which bacteria wereharvested in sterile physiological saline, washed, and ad-justed by comparison with an opacity standard to ca. 8.4 x
108 CFU/ml. This suspension was used to prepare decimaldilutions of bacteria starting from 2 x 108 CFU/ml. Platingon CRAMP and magnesium oxalate agar (24) confirmed thatthe great majority (>95%) of bacteria in each inoculum boreplasmids.Mouse virulence determinations. (i) Estimation of median
lethal dose (LD5s). The influence of iron and desferrioxamineB alone and in combination on the virulence of Y. enteroco-litica for mice was determined as follows. Groups of five
774
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
EFFECTS OF IRON AND DESFERAL ON YERSINIOSIS 775
TABLE 1. Bacteria used in this studyCalcium
Strain Serogroup Origin hemin binding dependence Autoagglutination Plasmidaheminbinding at 370C
30.42.67 03 Human; Sweden + + + 47 Md30.42.67 CR- 03 Spontaneous mutant - - - None6683/20 03 Human; Australia + + + 47 MdAM9 05 Milk; Australia + - - 40 MdWA 08 Human; United States + + + 42 MdA2635 08 Milk; United States + + + 42 MdE518 09 Human; Belgium + + + 48 MdE531 09 Human; Belgium + + + 48 Md
a Md, Megadalton.
BALB/c adult male mice were inoculated intraperitoneally(i.p.) with 0.5 ml of 20% (vol/vol) iron dextran containing 10mg of iron per ml (Imferon; Fisons Pty. Ltd., Sydney,Australia) or an equivalent amount of dextran T10 (Phar-macia Fine Chemicals AB, Uppsala, Sweden). The followingday, mice were given 0.5 ml of a saline solution containing 10mg of desferrioxamine B mesylate per ml (Desferal; Ciba-Geigy Ltd., Lane Cove, Australia) or 0.5 ml of saline i.p.Preliminary experiments in which Desferal was administeredin divided doses over a 3-day period gave the same results asthe single-dose regimen, which for convenience was adoptedas the method of choice. Twenty-four hours after receivingDesferal, mice were injected i.p. with 0.5 ml of a bacterialsuspension and observed for 3 weeks. The LD50 was deter-mined by the method of Reed and Muench (26). Controlsincluded mice inoculated with 108 heat-killed bacteria, Des-feral, and iron dextran, alone and in all possible permuta-tions.
(ii) Desferal dose-response relationship. To determinewhether susceptibility to Y. enterocolitica was affected bythe dose of Desferal, we inoculated groups of 10 mice i.p.with 0.5 ml of saline containing from 0.6 to 10 mg of Desferalin twofold dilutions. A control group received only saline.The next day, mice were infected i.p. with 103 Y. enteroco-litica 30.42.67 and observed for 3 weeks.
(iii) Kinetics of infection. For monitoring of the i.p. growthof yersiniae in Desferal-treated and control animals, and theappearance of bacteria in the blood, the following experi-ment was conducted. Mice were divided into two groups toreceive 0.5 mg of Desferal or saline. The following day, allanimals were inoculated i.p. with 3 x 104 Y. enterocolitica30.42.67 organisms. Immediately afterwards and 2, 4, 8, 12,24, 48, and 72 h later, four mice in each group was sacrificedby cervical dislocation. One milliliter of sterile saline wasinjected i.p., and samples of heart blood and peritoneal
TABLE 2. Median lethal dose of Y. enterocolitica for micepretreated with iron-dextran, Desferal, or both
LD50 after the following pretreatment":None Iron Desferal Iron + Desferal
30.42.67 03 >108 107.6 101.1 <1030.42.67 CR- 03 >108 >108 >108 >1086683/20 03 >108 107.2 1o3 10AM9 05 > 108 > 108 > 108 >108WA 08 103.17ol.7 lol.3 10A2635 08 102.7 1012 10 <10E518 09 >108 107.6 103-7 1o2.3E531 09 >108 505110302.5
a Iron (5 mg), as iron-dextran, or Desferal (5 mg), or both, were adminis-tered i.p.
washings were collected aseptically for quantitative bacte-rial culture.
(iv) Oral infection. The effect of iron and Desferal on oralinfection with Y. enterocolitica was also determined. Micewere injected i.p. with 5 mg of iron or 5 mg of Desferal orboth on successive days as described above. On day 3,animals were deprived of drinking water, after which waterwas provided that contained 101 Y. enterocolitica cells perml. Mice were permitted to drink contaminated water adlibitum for 24 h before clean water was reintroduced. Ani-mals were observed daily for 14 days for development ofdiarrhea or other evidence of illness. For examination ofdiarrhea, mice were transferred to a clean cage for 1 h eachday and their feces were observed. Diarrhea was evidencedby pale-colored, vouminous, occasionally watery stools thatoften contained mucus. Freshly passed feces were examinedfor the presence of Y. enterocolitica by plating on Mac-Conkey and Yersinia selective agar (Oxoid Ltd., Basing-stoke, Hampshire, England).Guinea pig keratoconjuctivitis test. Guinea pigs weighing
ca. 250 g were injected i.p. with 1 ml of undiluted iron-dextran containing 5 mg of iron, with or without 50 mg ofDesferal. Dextran T10 and saline served as controls fromiron dextran and Desferal, respectively.One day later, a thick paste of bacteria from a 48-h
nutrient agar-grown culture was inoculated into one conjunc-tival sac of each of two guinea pigs. The other eye was leftuninoculated as a control. In some instances, a drop ofundiluted iron-dextran or of a 50-mg/ml Desferal solution (inaccordance with the preinoculation schedule) was instilled inthe conjunctiva at the time of infection. Subsequent experi-ments, however, showed that this bore little influence on theoutcome of infection. Guinea pigs were observed for 10 daysfor the development of purulent keratoconjunctivitis orother signs of illness.
In vitro assimilation of iron. A plate assay for utilization ofiron compounds and Desferal by Y. enterocolitica was used.Bacteria grown on nutrient agar for 48 h were depleted ofiron by incubation in 10 ml of iron-depleted broth containing0.2% (wt/vol) Casamino Acids, 50 mM NaCl, 10 mM NH4C1,2.5 mM Na2S203, 1.4 mM K2HPO4, 0.4 mM MgSO4, 40 mMmorpholinepropanesulfonic acid and 10 mM Tricine (SigmaChemical Co., St. Louis, Mo.), pH 7.0, which was heatsterilized and to which had been added 10 ,ug of filter-steri-lized iron-free transferrin per ml (Sigma Chemical Co.).Cultures were shaken at 25°C overnight and inoculated ontotransferrin agar plates with the same composition as iron-re-duced broth plus 1.4% agarose.Samples (20 ,ul) of sterile test solutions containing various
amounts of iron dextran, ferric ammonium citrate, ferricchloride, or Desferal were pipetted onto 6-mm filter paper
VOL. 47, 1985
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
776 ROBINS-BROWNE AND PRPIC
disks (Oxoid Ltd.), which, after drying, were placed on theagar surface. Plates were incubated at 25°C for 72 to 96 h,and the diameter of the zone of exhibition around each diskwas measured. For these experiments, all media and rea-
gents were prepared in sterile plasticware with deionizeddouble-distilled water.
RESULTSMouse virulence studies. The results of the LD50 determi-
nations and the effect of iron dextran and Desferal on thesevalues are summarized in Table 2. The essential findingswere as follows. (i) In contrast to serogroup 08 strains, Y.enterocolitica of serogroups 03 and 09 was avirulent formice not pretreated with iron-dextran or Desferal. (ii) De-pending on the infecting strain, iron-dextran reduced theLD50 ca. 10- or 100-fold. (iii) Desferal reduced the LD50 ofserogroup 03 and 09 strains more than 100,000-fold; itseffect on serogroup 08 bacteria was less pronounced. (iv) Ofall mice infected with Y. enterocolitica, those which re-
ceived Desferal with or without iron-dextran died soonerthan control mice given only iron or dextran. (v) Thevirulence-enhancing effects of Desferal and iron-dextranwere additive insofar as the combination was more effectivethan either agent given alone. (vi) Y. enterocolitica strain30.42.67 CR-, which lacked the virulence-associatedplasmid, was avirulent for mice regardless of the preinfec-tion treatment schedule. The same was true of strain AM9,which despite possessing a 40-megadalton plasmid did notdisplay the virulence characteristics of calcium dependenceand autoagglutination (Table 1).The virulence-enhancing effect of Desferal was dose re-
lated. All mice given 103 bacteria with 2.5, 5, or 10 mg ofDesferal died within 1 week of infection, whereas mice giventhe same bacterial inoculum with 1.25 mg of Desferal or lesssurvived longer than 3 weeks (Fig. 1).The fate of injected bacteria in Desferal-treated and con-
trol mice is shown in Fig. 2. In mice that received Desferal,i.p. numbers of bacteria remained constant for 8 h and thenrose steadily to 2 x 107 per ml at 24 h. Bacteria weredetected in heart blood 12 h after inoculation, and by 24 hthey had reached 2 x 104 per ml. By 48 h, all Desferal-treated animals had died. In control animals, bacterial num-bers held steady for ca. 24 h and then declined; bacteria werenot detected in the blood at any time.
100 -
80 -
0
2 60-
C 40-
a)
20 -
0-
0 0.62 1.25 2.5 5 10
Desferal dose (mg)FIG. 1. Percentage of mice surviving 3 (0), 5 (0), and 7 (A) days
after i.p. inoculation with 103 Y. enterocolitica 30.42.67 cells andvarious doses of Desferal.
8-
7 -
E0C.,o-
0
6 3-0
o 2 -
1 -
OJ0 12 24 48 72
Hours after infection
FIG. 2. Fate of Y. enterocolitica 30.42.67 in mice given Desferal(0, A) or saline (0). Peritoneal contents (0, 0) and blood (A) weresampled at the times shown. Each point is the mean count from fouranimals. All mice that received Desferal died within 48 h. Bacteriawere not detected in the blood of mice that received saline. Thehorizontal line indicates the detection limit of the culture methodused.
The results of experiments in which mice were infectedorally are shown in Table 3. As was the case with i.p.infection, serogroup 03 and 09 strains caused disease onlyin mice pretreated with iron-dextran or Desferal. Two orthree days after ingesting Y. enterocolitica 03 or 09, micegiven iron alone developed diarrhea which cleared sponta-neously within 2 to 4 days. Mice that received Desferal withor without iron-dextran developed diarrhea and died 2 to 3days later. Serogroup 08 strains caused diarrhea and a fatalinfection even when no iron or Desferal was provided,although iron- and Desferal-treated mice did die 1 to 3 daysearlier than control animals. Y. enterocolitica strains 30.42.67CR- and AM9 produced no disease, regardless of thepreinfection treatment schedule. Yersiniae were recoveredfrom the feces of all animals with diarrhea, but only incon-sistently from those with normally formed stools.Guinea pig virulence studies. The results of the guinea pig
keratoconjunctivitis test paralleled those of the mouse viru-
TABLE 3. Outcome of oral Y. enterocolitica infection of micepretreated with iron-dextran, Desferal or botha
Outcomeb of infection after the followingStrain Serogrout, pretreatment:
None Iron Desferal Iron + Desferal
30.42.67 03 d d,D d,D30.42.67 CR- 036683/20 03 d d,D d,DAM9 05 - -WA 08 d,D d,D d,D d,DA2635 08 d,D d,D d,D d,DE518 09 d d,D d,DE531 09 d d,D d,D
a Mice received 5 mg of iron as iron-dextran or 5 mg of Desferal or both onsuccessive days. They were deprived of drinking water for 24 h and then givenwater that contained 109 Y. enterocolitica cells per ml and permitted to drinkad libitum for 24 h befpre clean water was reintroduced.
b _, No effect; d, diarrhea; D, death.
INFECT. IMMUN.
I
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

EFFECTS OF IRON AND DESFERAL ON YERSINIOSIS 777
TABLE 4. Outcome of intraconjunctival inoculation of Y.enterocolitica into guinea pigs pretreated with iron-dextran,
Desferal, or bothOutcomea of inoculation with Y. enteroco-
Strain Serogroup litica after the following pretreatmentbNone Iron Desferal Iron + Desferal
30.42.67 03 C PC,Dc PC,D30.42.67 CR- 03 -6683/20 03 C PC,DC PC,DAM9 05WA 08 PC PC PC,D' PC,DA2635 08 PC PC PC,DC PC,DcE518 09 C PC PCE531 09 - C PC PC
a-, No effect; C, mild conjunctivitis; PC, severe purulent conjunctivitis;D, death.bIron (50 mg) or Desferal (50 mg) or both were administered i.p.c Only one of 2 animals died.
lence studies (Table 4). Y. enterocolitica serogroups 03 and09 were avirulent for animals not given iron-dextran orDesferal. Guinea pigs pretreated with iron developed mildself-limiting conjunctivitis, whereas those given Desferaldeveloped severe purulent conjunctivitis which, in somecases, progressed to panophthalmitis and a fatal systemicinfection. Strains 30.42.67 CR- and AM9 were avirulentunder all test conditions.
In vitro assimilation of iron compounds. None of the Y.enterocolitica strains examined grew on transferrin agarunless iron or Desferal was provided or transferrin wasomitted (Fig. 3). This was in contrast to several strains ofYersinia frederiksenii, Yersinia kristensenii, and Yersiniaintermedia, which produced siderophores and therefore grewluxuriantly on transferrin agar without supplements (manu-script in preparation).The growth response of bacteria to both iron (Fig. 4) and
Desferal (data not shown) was linear with respect to concen-tration. The iron requirements of bacteria in serogroups 03,05, and 09 were similar. Serogroup 08 strains, however,required appreciably less iron for growth than those in theother serogroups (Fig. 4). The requirement for iron and theability of bacteria to utilize iron-bound Desferal as a growth
FIG. 3. Transferrin agar, showing growth of Y. enterocolitica30.42.67 around disks containing iron-dextran (left) and desferri-oxamine B (right).
promoter were not influenced by carriage of the virulence-associated plasmid.
DISCUSSIONIn 1973, Rogers reported that catechol siderophores stim-
ulate bacterial growth in vitro and in vivo and suggested thatthey should be regarded as true virulence factors (30). Sincethen, several researchers have investigated the effect ofcatechols and hydroxamates, including Desferal, on experi-mental infections. Their findings have been conflicting.With some bacteria, notably Neisseria meningitidis (7),
Neisseria gonorrhoeae (9), Listeria monocytogenes (32),and a mutant strain of Salmonella typhimurium defective incatechol synthesis (13), Desferal may afford a degree ofprotection against infection. Recognition of this factprompted Calver et al. to suggest that Desferal may have achemotherapeutic role in severe neisserial disease (7).There is evidence, however, that infection with certain
other bacteria may be aggravated by Desferal. These includewild-type Salmonella typhimurium (13), Klebsiella spp. (17),and now Y. enterocolitica. The disparate effects of Desferalon different infections relate to the ability of the infectingagent to use ferrioxamine, the iron-containing form of Des-feral, as a growth factor. Thus, in infections caused bybacteria lacking both a high-affinity iron uptake system andferrioxamine receptors, e.g., pathogenic neisseriae and L.monocytogenes, Desferal favors the host by binding iron andsequestering it from invading microbes. On the other hand,Desferal will usually stimulate the in vivo multiplication ofbacteria which can utilize ferrioxamine, such as Klebsiellaspp. and Y. enterocolitica.
Y. enterocolitica is predominantly an enteric pathogen (2),and although it lacks a high-affinity iron-chelating system ofits own, it is able to utilize siderophores of other bacteria (4,22). In the intestine, where these compounds are abundant,yersiniae should be able to multiply freely. In tissues,however, a lack of readily assimilable iron would restrict
20 -
- 15-EL-
0
E 10-
N 5-
0 I I I ,0.6 2.5 10 40
Iron-dextran content (,g)160
FIG. 4. Effect of iron-dextran on growth of Y. enterocoliticaserogroups 03, 05, and 09 (0) and serogroup 08 (0) on transferrinagar. Growth was expressed as the diameter of the zone of exhibi-tion around a 6-mm filter paper disk containing various concentra-tions of iron-dextran. Similar results were obtained when ferricammonium citrate, ferric chloride, or Desferal was substituted foriron-dextran, although only 5 ng of Desferal (for serogroup 03, 05,and 09 bacteria) and 0.05 ng of Desferal (for serogroup 08 strains)was sufficient to stimulate growth. Iron and Desferal requirementswere unaffected by plasmid carriage.
VOL. 47, 1985
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

778 ROBINS-BROWNE AND PRPIC
growth. Provision of additional iron or ferrioxamine enablesbacteria to overcome this handicap. Parenthetically, the factthat Y. enterocolitica organisms utilize the siderophores ofother bacteria provides an explanation for our earlier obser-vation of synergisim between Salmonella typhimurium andY. enterocolitica in experimentally infected mice (28).The close correlation between in vitro utilization of Des-
feral and its stimulatory effect on bacterial multiplication invivo indicates that Desferal acts primarily as a growthstimulant and not as an immunosuppressant. Support for thisconclusion is provided by our finding that Desferal had noeffect on the course of infection with Y. enterocoliticastrains lacking virulence-associated plasmids. The effect ofDesferal contrasts with that of iron, which while serving as apromoter of microbial growth may also suppress immunedefenses (14, 16).The finding that serogroup 08 strains were less dependent
on iron for in vitro growth than were their 03 and 09counterparts could account for their innate increased viru-lence for animals and their reduced response to Desferal invivo. The experimental findings agree with clinical experi-ence suggesting that serogroup 08 yersiniae may be morelikely to produce septicemia in previously healthy individu-als (11, 15) than are 03 and 09 strains, which are opportun-ists heavily reliant on the iron status of their host (6, 25, 26,29, 31). Difference in iron requirements or Desferal utiliza-tion, however, cannot explain the lack of pathogenicity ofthe plasmidless serogroup 03 strain, 30.42.67 CR-, or theserogroup 05 strain, AM9, both of which were able to utilizeinorganic iron compounds and Desferal as efficiently as theirvirulent counterparts. Thus, although the virulence-associ-ated plasmid in Y. enterocolitica may encode hemin binding(24) and the synthesis of novel outer membrane proteins(23), there is no evidence that the plasmid governs iron orferrioxamine utilization. Clearly, iron assimilation is onlyone of several factors contributing to the virulence of Y.enterocolitica.
In this study, Desferal was more efficient than iron instimulating growth of Y. enterocolitica in vitro and in vivo.This is probably because Desferal provides Y. enterocoliticawith iron in a soluble, easily assimilable form. The sourcefrom which Desferal obtains iron is not known precisely. Intransferrin agar, it probably competes successfully withtransferrin for the small amount of iron that contaminatesthe constituents of the medium (1). In vivo, it is likely thatDesferal derives iron from sources other than transferrin(18). In iron-overloaded mice, iron-dextran probably sup-plied much of the iron in ferrioxamine.
In summary, this study has shown that iron-dextran andDesferal exert an adverse influence on the course of exper-imental yersiniosis. The pronounced susceptibility of Des-feral-treated mice and guinea pigs to infection with sero-group 03 and 09 Y. enterocolitica provides a convenientmeans for investigating the pathogenicity of these bacteria,which under normal circumstances are avirulent for labora-tory animals. The effect of Desferal probably accounts inpart for the increased susceptibility of iron-overloaded hu-mans to systemic yersiniosis and possibly to other infec-tions. These observations suggest that for patients requiringchelation therapy, iron chelators should be sought thatcannot be used by pathogenic bacteria.
ACKNOWLEDGMENTS
We are indebted to D. Hughes, I. Juhlin, K. Wachsmuth, and G.Wauters for providing the bacteria used in these investigations.
This work was supported by a grant from the Australian NationalHealth and Medical Research Council. J.K.P. is the recipient of anAustralian Public Service Postgraduate Study Award.
LITERATURE CITED1. Aisen, P. 1976. The binding and release of iron by transferrin.
Birth Defects Orig. Artic. Ser. 12:81-95.2. Bottone, E. J. 1977. Yersinia enterocolitica: a panoramic view of
a charismatic microorganism. Crit. Rev. Microbiol. 5:211-241.3. Bouza, A., A. Dominguez, M. Meseguer, L. Buzon, D. Bioxeda,
M. J. Revillo, L. de Rafael, and J. Martinez-Beltran. 1980.Yersinia enterocolitica septicemia. Am. J. Clin. Pathol.74:404-409.
4. Brock, J. H., and J. Ng. 1983. The effect of desferrioxamine onthe growth of Staphylococcus aureus, Yersinia enterocoliticaand Streptococcus faecalis in human serum: uptake of desfer-rioxamine-bound iron. FEMS Microbiol. Lett. 20:439-442.
5. Bullen, J. J. 1981. The significance of iron in infection. Rev.Infect. Dis. 3:1127-1138.
6. Butzler, J. P., M. Alexander, A. Segers, N. Cremer, and D.Blum. 1978. Enteritis, abscess, and speticemia due to Yersiniaenterocolitica in a child with thalassemia. J. Pediatr. 93:619-621.
7. Calver, G. A., C. P. Kenny, and D. J. Kushner. 1979. Inhibitionof the growth of Neisseria meningiditis by reduced ferritin andother iron-binding agents. Infect. Immun. 25:880-890.
8. Finkelstein, R. A., C. V. Sciortino, and M. A. McIntosh. 1983.Role of iron in microbe-host interactions. Rev. Infect. Dis.5:S759-S777.
9. Finkelstein, R. A., and R. J. Yancey. 1981. Effect of sidero-phores on virulence of Neisseria gonorrhoeae. Infect. Immun.32:609-613.
10. Graziano, J. H., and A. Cerami. 1977. Chelation therapy for thetreatment of thalassemia. Semin. Hematol. 14:127-134.
11. Gutman, L. T., E. A. Ottesen, T. J. Quan, P. S. Noce, and S. L.Katz. 1973. An inter-familial outbreak of Yersinia enterocoliticaenteritis. N. Engl. J. Med. 288:1372-1377.
12. Hughes, D. 1979. Isolation of Yersinia enterocolitica from milkand a dairy farm in Australia. J. Appl. Bacteriol. 46:125-130.
13. Jones, R. L., C. M. Peterson, R. W. Grady, T. Kumbaraci, A.Cerami, and J. H. Graziano. 1977. Effects of iron chelators andiron overload and Salmonella infection. Nature (London)267:63-64.
14. Kaplan, S. S., and R. E. Basford. 1979. Exogenous iron andimpairment of intraleukocytic bacterial killing, or the leukocytetells her story, p. 92-95. In D. Schlessinger (ed.), Microbiol-ogy-1979. American Society for Microbiology, Washington,D.C.
15. Keet, E. E. 1974. Yersinia enterocolitica septicemia: source ofinfection and incubation period identified. N.Y. State J. Med.74:2226-2230.
16. Keown, P., and B. Descamps-Latscha. 1983. In vitro suppressionof cell-mediated immunity by ferroproteins and ferric salts. Cell.Immunol. 80:257-266.
17. Khimji, P. L., and A. A. Miles. 1978. Microbial iron-chelatorsand their action on Klebsiella infections in the skin of guineapigs. Br. J. Exp. Pathol. 59:137-147.
18. Lipschitz, D. A., J. Dugard, M. 0. Simon. T. H. Bothwell, andR. W. Charlton. 1971. The site of action of desferrioxamine. Br.J. Haematol. 20:395-404.
19. Melby, K., S. Slordahl, T. J. Guttenberg, and S. A. Nordbo.1982. Septicaemia due to Yersinia enterocolitica after oraloverdoses of iron. Br. Med. J. 285:467-468.
20. Modell, B. 1979. Advances in the use of iron-chelating agents forthe treatment of iron overload. Prog. Hematol. 11:267-312.
21. Neilands, J. B. 1981. Microbial iron compounds. Annu. Rev.Biochem. 50:715-731.
22. Perry, R. D., and R. R. Brubaker. 1979. Accumulation of ironby yersiniae. J. Bacteriol. 137:1290-1298.
23. Portnoy, D. A., H. Wolf-Watz, I. Bolin, A. B. Beeder, and S.Falkow. 1984. Characterization of common virulence plasmidsin Yersinia species and their role in the expression of outermembrane proteins. Infect. Immun. 43:108-114.
24. Prpic, J. K., R. M. Robins-Browne, and R. B. Davey. 1983.
INFECT. IMMUN.
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

EFFECTS OF IRON AND DESFERAL ON YERSINIOSIS 779
Differentiation between virulent and avirulent Yersinia entero-colitica isolates by using Congo red agar. J. Clin. Microbiol.18:486-490; Erratum 19:446.
25. Rabson, A. R., A. F. Hallett, and H. J. Koornhof. 1975.Generalized Yersinia enterocolitica infection. J. Infect. Dis.131:447-451.
26. Reed, L. J., and H. Muench. 1935. A simple method of estimat-ing fifty per cent endpoints. Am. J. Hyg. 27:493-497.
27. Robins-Browne, R. M., A. R. Rabson, and H. J. Koornhof. 1979.Generalized infection with Yersinia enterocolitica and the roleof iron. Contrib. Microbiol. Immunol. 5:277-282.
28. Robins-Browne, R. M., A. R. Rabson, and H. J. Koornhof. 1981.Yersinia enterocolitica in South Africa, p. 193-203. In E. J.Bottone (ed.), Yersinia enterocolitica. CRC Press, Inc., BocaRaton, Fla.
29. Roche, G., B. Leheup, A. Gerard, P. Canton, C. Lion, G.Leichtmann, and J. B. Dureux. 1982. Septicemies a Yersiniaenterocolitica. Revue generale a propos d'un nouvaeu cas chezune jeune femme presentant une thalassemie majeure. Rev.Med. Interne 3:65-74.
30. Rogers, H. J. 1973. Iron-binding catechols and virulence inEscherichia coli. Infect. Immun. 7:445-456.
31. Smith, R. E., A. M. Carey, J. M. Damare, F. M. Hetrick, R. W.Johnston, and W. H. Lee. 1981. Evaluation of iron dextran andmucin for enhancement of the virulence of Yersinia enterocoli-tica serotype 0:3 in mice. Infect. Immun. 34:550-560.
32. Sword, C. P. 1966. Mechanisms of pathogenesis in Listeriamonocytogenes infection. J. Bacteriol. 92:536-542.
33. Weinberg, E. D. 1978. Iron and infection. Microbiol. Rev.42:45-66.
VOL. 47, 1985
on June 8, 2018 by guesthttp://iai.asm
.org/D
ownloaded from


![Vol. in U.S.A. Effect ofIron and Salt Prodigiosin Synthesis · culture on TS slants (1.0% ion agar no. 2 [Colab], 3.0% Trypticase soy broth [BBL]) or on slants of Brain Heart Infusion](https://img.pdfslide.us/doc/110x75/5f6410a6530e2f494935985b/vol-in-usa-effect-ofiron-and-salt-prodigiosin-synthesis-culture-on-ts-slants.jpg)


![T2 mapping and T2* imaging in heart failure...and intravenous desferrioxamine chelation therapy [19]. In cases of severe cardiac iron loading, or when LV function is impaired, a combined](https://img.pdfslide.us/doc/110x75/5f0eeb4a7e708231d441958c/t2-mapping-and-t2-imaging-in-heart-failure-and-intravenous-desferrioxamine.jpg)






![CY-'Proc. Nat. Acad. Sci. USA Vol. 70, No.8, pp. 2429-2433, August 1973 Synthetic Analogs ofthe Active Sites ofIron-Sulfur Proteins. Structure and Properties ofBis[o-xylyldithiolato-MA2-sulfidoferrate(III)],](https://img.pdfslide.us/doc/110x75/5e9dfa63d66a0c175d562bb1/cy-proc-nat-acad-sci-usa-vol-70-no8-pp-2429-2433-august-1973-synthetic.jpg)




![Porphyria Cutanea Tarda (PCT) - Austin Publishing Group · include removal of iron by phlebotomy or low-dose chloroquine, [10] desferrioxamine infusion, cholestyramine, iron chelators,](https://img.pdfslide.us/doc/110x75/5fc0af1be0758e4a687e514c/porphyria-cutanea-tarda-pct-austin-publishing-group-include-removal-of-iron.jpg)





