Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1991 by The American Society for Biochemistry and Molecular Biolopy, Inc
Vol. 266, No 7, Issue of March 5, pp. 4500-4507,1991 Printed in U. S. A .
Depletion of Cellular Calcium Accelerates Protein Degradation in the Endoplasmic Reticulum*
(Received for publication, October 22, 1990)
Thomas Wileman$$, Lawrence P. Kane$, Gerald R. Carsonll, and Cox TerhorstS From the $Laboratory of Molecular Immunology, Dana Farber Cancer Institute, Boston Massachusetts 02115 and the VProtein Expression Group, T-cell Sciences, Cambridge Massachusetts 02139-4135
In this study the effects of A23187 and thapsigargin on the degradation of T-cell antigen receptor-/3 (TCR- B) and CD3-6 in the endoplasmic reticulum have been studied. Preliminary experiments showed that these drugs had different effects on the secretory pathway. Depletion of cellular calcium pools by incubation of cells with A23187 in calcium-free medium blocked transport between the endoplasmic reticulum and the Golgi apparatus whereas thapsigargin caused a modest increase in transport. When added to cells transfected with TCR-/3 or CD3-6 the drugs caused an immediate stimulation of proteolysis of presynthesized protein and at maximum effective concentrations caused a 3- fold increase in the rate of degradation. They did not affect the lag period of 1 h which precedes degradation of newly synthesized proteins. Chelation of cytosolic calcium also accelerated degradation, suggesting that depletion of calcium from the endoplasmic reticulum was the main stimulus of proteolysis and that increased degradation was not caused by a transient increase in cytosolic calcium levels. The selectivity of degradation in the endoplasmic reticulum was maintained. A23187 had no effect on the stability of CD3-7 nor co-trans- fected 6 - B dimers. Calcium depletion increased the overall rate of degradation in the endoplasmic reticu- lum and increased the rate of proteolysis of an “anchor minus” @ chain. The results suggested that proteolysis within the endoplasmic reticulum may be regulated by the high concentrations of Ca2+ which are stored in the organelle. Ca2+ may be required for protein folding. Calcium depletion may have caused the /3 and 6 chains to adopt a conformation that was more susceptible to proteolysis. Alternatively, calcium depletion may have disrupted the lumenal content of the endoplasmic retic- ulum and increased the access of proteases to potential substrates.
Newly synthesized proteins are retained in the endoplasmic reticulum until they fold correctly (Copeland et al., 1986, 1988; reviewed by Rose and Doms, 1988; Hurtley and Helen- ius, 1989). The endoplasmic reticulum contains enzymes that catalyze protein folding and subunit assembly (reviewed by Rothman, 1989) yet some proteins, particularly mutant pro-
* This work was supported in part by a grant from the Council for Tobacco Research and National Institutes of Health Grant A1-15066. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
5 Recipient of Basil O’Connor Starter Scholar Award 5-760 from the March of Dimes and a Fellow of the King Trust of the Medical Foundation. To whom correspondence should be sent: Rm. 309 JFB, Dana Farber Cancer Institute, 44 Binney St., Boston MA 02115.
teins (Gething et al., 1986) or components of multimeric receptors that are made in excess, fail to attain the confor- mation required for their transport to the Golgi apparatus. Such proteins are retained in the endoplasmic reticulum, and in some cases they are degraded rapidly’ (Lippincott-Schwartz et al., 1988; Amara et al., 1989; Rotundo et al., 1989; Stoller and Shields, 1989; Sato et al., 1990; Sitia et al., 1990). These processes of retention, sorting for secretion, and degradation may be interrelated functions. The first two serve to organize the assembly and transport of proteins from the endoplasmic reticulum, and proteolysis removes proteins that would oth- erwise accumulate in the organelle.
The T-cell antigen receptor (TCR)’ has provided a good system for the study of assembly and degradation in the endoplasmic reticulum. The receptor is composed of six trans- membrane proteins; the TCR-a and -p chains recognize an- tigens presented to the T-cell whereas the associated CD3-y, -6, and -t subunits and the [ chain homodimer are essential for the transport of the receptor to the cell surface. Single subunits and partial complexes are retained in the endo- plasmic reticulum where they show different susceptibilities to proteolysis. The TCR-a and $3 chains and the CD3-6 subunit are degraded, whereas the CD3-y and -6 subunits and the [ homodimer are long lived (Chen et al., 1988; Bonifacino et al., 1989; Wileman et al., 1990a). Interestingly, the assembly of subunits within the endoplasmic reticulum alters their intracellular survival, and associations with the stable CD3- y or -t subunits protect labile chains from proteolysis (Boni- facino et al., 1989; Wileman et al., 1990a). Although the precise site of degradation of newly synthesized T-cell receptor chains is unknown, it seems likely that it takes place in the early compartments of the secretory pathway. The N-linked sugars of degraded proteins are not processed by enzymes of the medial or cis Golgi, and proteolysis is unaffected by agents that block transport from the endoplasmic reticulum (Lippin- cott-Schwartz et al., 1988; Bonifacino et al., 1989; Wileman et al., 1990b).
This present study was stimulated by recent reports dem- onstrating that early events in the secretory pathway can be disrupted by drugs that deplete cells of calcium ions (Lodish and Kong, 1990; Booth and Koch, 1989) or by mutations that affect intracellular calcium homeostasis (Rudolph et al., 1989). Each of these reports has implicated the endoplasmic reticu- lum, an intracellular store of calcium, as the main site of
Le, A., Graham, K. S., and Sifers, R. N., (1990) J. Biol. Chem.
‘The abbreviations used are: TCR, T-cell receptor for antigen; MAPT-AM, 1,2-bis-5-methylaminophenoxylethane- N,N’-tetraace- toxymethyl acetate; ELISA, enzyme-linked immunosorbent assay; SDS, sodium dodecyl sulfate; Endo-H, endoglycosidase-H; IL2, inter- leukin-2; CHO, Chinese hamster ovary; mAb, monoclonal antibody; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid.
265, 14001-14007.
4500
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Effect of Calcium on ER Degradation 4501
action. Calcium ionophores slow the movement of proteins from the endoplasmic reticulum to the Golgi apparatus (Lod- ish and Kong, 1990) whereas prolonged depletion of cellular calcium disrupts the sorting of the resident lumenal proteins of the endoplasmic reticulum and in some cells induces their secretion (Booth and Koch, 1989). Such results are consistent with the observation that yeasts carrying a mutant calcium ATPase (PMR1-) secrete proteins that are retained in the endoplasmic reticulum of wild-type strains (Rudolph e t al., 1989).
These observations have shown that the lumenal calcium ions of the endoplasmic reticulum play an important role in regulating the early functions of the secretory pathway. It is possible that calcium ions are also used to regulate degrada- tion in endoplasmic reticulum. In this study cells transfected with the labile TCR-/3 chain have been used to test this possibility. Intracellular calcium pools were depleted from cells using the calcium ionophore A23187 in calcium-free medium, and the calcium store of the endoplasmic reticulum was drained by incubating cells with thapsigargin, a specific inhibitor of the Ca2+ ATPase of the endoplasmic reticulum (Thastrup et al., 1990). Both procedures stimulated degrada- tion in the endoplasmic reticulum. It is possible that these results reflect a physiological role for lumenal Ca2+ in the regulation of proteolysis in the organelle.
EXPERIMENTAL PROCEDURES
Materials-A23187 and MAPT-AM (1,2-bis-5-methylaminophen- oxylethane- N,N"tetraacetoxymethyl acetate) were purchased from Calbiochem. Thapsigargin was purchased from LC Services Corpo- ration (Woburn, MA). Brefeldin A was purchased from Epicenter Technologies (Madison WI).
Cells and Transfections-The CHO cell line K1 (ATCC CCl 61) was obtained from ATCC (Rockville, MD). The cells were maintained in 5% CO, at 37" C in a base medium of RPMI supplemented with glutamine (2 mM). The African green monkey kidney cell line COS- 1 was a generous gift of Brian Seed (Masschusettes General Hospital, Boston, MA) and was cultured at 37" C in Dulbecco's modified Eagle's medium supplemented with 2 mM glutamine and buffered using 10% CO,. All media contained 8% heat-inactivated fetal calf serum. Tissue culture media were purchased from GIBCO or Sigma. The transfec- tion of cells with the components of the T-cell antigen receptor (TCR-8, CD3-6 and -y and p-e dimers) has been reported previously (Wileman et al., 1990a, 199Ob). Mouse L-cells expressing the full- length 55-kDa chain of the IL2 receptor were a generous gift from Dr. Warner Greene (Duke University, Durham, NC). The construc- tion of "anchor minus" mutants of the IL2 receptor and the TCR-p chain and their transfection into CHO cells have been reported previously (Wileman et al., 1990~).
Antibodies-W76 and IdentiTy &F1@ were provided by T-cell Sciences (Cambridge, MA) and are specific for TCR-8. S P 19 and SP 64 precipitate CD3-6, and S P 34 precipitates CD3-e (Pessano et al., 1985). 7G7 is specific for the human IL2 receptor (Rubin et al., 1985). For use in ELISAs the antibodies were affinity purified from ascites using protein G (SP 19 and S P 64) or protein A (W76 and &F1) Sepharose (Pharmacia LKB Biotechnology Inc.). P-F1 and SP 64 were coupled to periodate-oxidized sugars of horseradish peroxidase.
Metabolic Labeling, Immunoprecipitation, and Electrophresk- Transfected cells were labeled metabolically using Tran3'S-label (ICN Radiochemicals, Cleveland OH). Cells (90% confluent, 10-cm dish) were preincubated for 1 h at 37 "C in methionine and cysteine-free medium, washed, and then pulse labeled for 30 min at 37 "C in the same medium containing 0.2 mCi/ml 35S. Cells were chased with complete medium for the indicated time periods. Labeled cells were lysed at 4" C in 1% Nonidet P-40 in immunoprecipitation buffer containing peptide protease inhibitors. Lysates were clarified by centrifugation, precleared, and immunoprecipitated as described pre- viously (Alarcon et al., 1988). Immunoprecipitates were washed ex- tensively in immunoprecipitation buffer and solubilized in reducing sample buffer in preparation for SDS-polyacrylamide gel electropho- resis. Polyacrylamide gel electrophoresis was performed using 12.5% slab gels (Laemmli, 1970). Gels were fixed and incubated with 1 M salicylic acid prior to autoradiography. To ensure that excess antibody
had been used for the pulse-chase immunoprecipitation analysis, lysates were reprecipitated with the relevant antibodies. In each case less than 10% of the original protein remained in the lysate.
Endoglycosidase Treatment-Endoglycosidase H was purchased from Genzyme (Boston, MA), and digestions were run overnight at 37" C. Immunoprecipitates were first suspended in 10 pl of 1% SDS and denatured by heating to 100" C for 3 min and then diluted to 50 pl with 50 mM phosphate buffer, pH 6.0, containing 50 milliunits of enzyme.
Laser Scanning Densitometry-Quantitation of protein bands was performed by scanning autoradiographs with a laser densitometer (LKB Ultroscan XL). A Gaussian integration method was used to estimate the intensity of bands (Gelscan XL software package, Phar- macia). The actin band was used to normalize data between different lanes and different experiments.
ELISA for TCR-6, CD3-6, and the IL2 Receptor-The precipitating antibodies (W76 for TCR-0 and SP 64 for CD3-6) were applied to Immulon 96-well plates (Dynatec Laboratories, Inc., Chantilly, VA) as 2 pg/ml solution in phosphate-buffered saline. Plates were washed with phosphate-buffered saline and blocked for 2 h with phosphate- buffered saline containing 0.05% Tween 20 and 10% bovine serum albumin. Antigen samples, diluted to 200 pl with blocking buffer, were applied to drained wells and incubated for 2 h at 37" C. Bound antigen was visualized in washed wells using horseradish peroxidase- coupled second antibodies (P-Fl for p and SP 19 for 6) diluted in phosphate-buffered saline containing 30% fetal calf serum, 6% phos- phate-buffered saline, and 0.75% Nonidet P-40. Bound peroxidase was determined colorimetrically by the addition of o-phenylenedi- amine and H,O,. IL2 receptor epitopes were analyzed using the Cell- free@ IL2 receptor assay kit from T-cell Sciences.
Cycloheximide Chase ELISA-The details of this assay have been described previously (Wileman et al., 1990b). Briefly, cells were fed for 1 h with fresh growth medium and then preincubated at 4' C for 30 min with 20 pg/ml cycloheximide in RPMI. To start the chase, cells were warmed to 37" C in RPMI containing 20 pg/ml cyclohexi- mide. At suitable time points the medium was removed, and mono- layers were lysed in 300 p1 of immunoprecipitation buffer. Lysates were left on ice for 1 h, insoluble material was pelleted by centrifu- gation, and supernatants (100-pl aliquots) were assayed for antigen using the relevant ELISA.
RESULTS
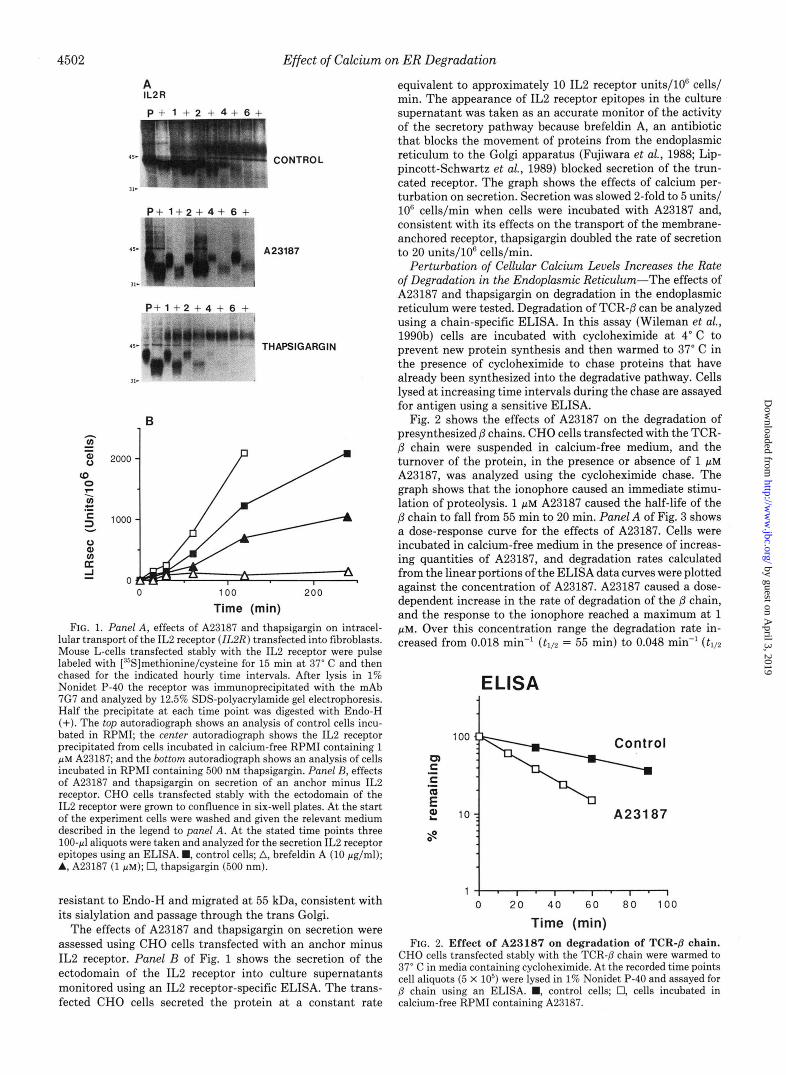
A23187 and Thapsigargin Disrupt the Secretory Pathway- In the first part of this study we have used fibroblasts trans- fected with the IL2 receptor to characterize the actions of A23187 and thapsigargin on the endoplasmic reticulum. The transfected IL2 receptor is a convenient marker for the secre- tory pathway and is transported efficiently from the endo- plasmic reticulum through the Golgi apparatus to the surface of fibroblasts. When the membrane anchor is removed from the protein the ectodomain of the receptor is secreted effi- ciently by cells (Treiger et al., 1986). Panel A of Fig. 1 shows a pulse-chase immunoprecipitation analysis of cells that have been transfected with a full-length IL2 receptor. Under con- trol conditions (top panel), approximately half of the pulse- labeled receptor became resistant to Endo-H after 2 h of chase. During this period the electrophoretic mobility of the protein slowed because of the addition of sialic acid residues to 0-linked oligosaccharides. These post-translational modi- fications indicated that the receptor had been transported through the Golgi apparatus. The center autoradiograph shows the experiment repeated for cells incubated with A23187 in calcium-free medium. The pulse-labeled receptor remained sensitive to Endo-H throughout the chase, suggest- ing that A23187 effectively blocked the transport of the recep- tor to the medial Golgi. The lower panel shows the effects of thapsigargin. In contrast to the action of A23187, it appeared that thapsigargin caused a modest increase in the rate of transport of the receptor to the medial Golgi. This was most evident at 4 h. In control cells, Endo-H-resistant material of 40 kDa was still apparent, but in cells incubated with thap- sigargin all the receptor precipitated at the 4-h time point was
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from
4502 Effect of Calcium on ER Degradation
A IL2R
~ + 1 + 2 + 4 + 6 +
P + 1 + 2 + 4 + 6 +
45- A23187
B 1
THAPSIGARGIN
(D 0 ~
7 . L u)
C .- 3 0 Q) u) a
2000
1000
0 0 100 200
Time (min) FIG. 1. Panel A , effects of A23187 and thapsigargin on intracel-
lular transport of the IL2 receptor (ZL2R) transfected into fibroblasts. Mouse L-cells transfected stably with the IL2 receptor were pulse labeled with [35S]methionine/cysteine for 15 min at 37" C and then chased for the indicated hourly time intervals. After lysis in 1% Nonidet P-40 the receptor was immunoprecipitated with the mAb 7G7 and analyzed by 12.5% SDS-polyacrylamide gel electrophoresis. Half the precipitate at each time point was digested with Endo-H (+). The top autoradiograph shows an analysis of control cells incu- bated in RPMI; the center autoradiograph shows the IL2 receptor precipitated from cells incubated in calcium-free RPMI containing 1 p~ A23187; and the bottom autoradiograph shows an analysis of cells incubated in RPMI containing 500 nM thapsigargin. Panel B, effects of A23187 and thapsigargin on secretion of an anchor minus IL2 receptor. CHO cells transfected stably with the ectodomain of the IL2 receptor were grown to confluence in six-well plates. A t the start of the experiment cells were washed and given the relevant medium described in the legend to panel A. At the stated time points three 100-pl aliquots were taken and analyzed for the secretion IL2 receptor epitopes using an ELISA. M, control cells; A, brefeldin A (10 pglml); A, A23187 (1 pM); 0, thapsigargin (500 nm).
resistant to Endo-H and migrated at 55 kDa, consistent with its sialylation and passage through the trans Golgi.
The effects of A23187 and thapsigargin on secretion were assessed using CHO cells transfected with an anchor minus IL2 receptor. Panel B of Fig. 1 shows the secretion of the ectodomain of the IL2 receptor into culture supernatants monitored using an IL2 receptor-specific ELISA. The trans- fected CHO cells secreted the protein at a constant rate
equivalent to approximately 10 IL2 receptor units/W cells/ min. The appearance of IL2 receptor epitopes in the culture supernatant was taken as an accurate monitor of the activity of the secretory pathway because brefeldin A, an antibiotic that blocks the movement of proteins from the endoplasmic reticulum to the Golgi apparatus (Fujiwara et al., 1988; Lip- pincott-Schwartz et al., 1989) blocked secretion of the trun- cated receptor. The graph shows the effects of calcium per- turbation on secretion. Secretion was slowed 2-fold to 5 units/ lo6 cells/min when cells were incubated with A23187 and, consistent with its effects on the transport of the membrane- anchored receptor, thapsigargin doubled the rate of secretion to 20 units/106 cells/min.
Perturbation of Cellular Calcium Levels Increases the Rate of Degradation in the Endoplasmic Reticulum-The effects of A23187 and thapsigargin on degradation in the endoplasmic reticulum were tested. Degradation of TCR-@ can be analyzed using a chain-specific ELISA. In this assay (Wileman et al., 1990b) cells are incubated with cycloheximide at 4" C to prevent new protein synthesis and then warmed to 37" C in the presence of cycloheximide to chase proteins that have already been synthesized into the degradative pathway. Cells lysed at increasing time intervals during the chase are assayed for antigen using a sensitive ELISA.
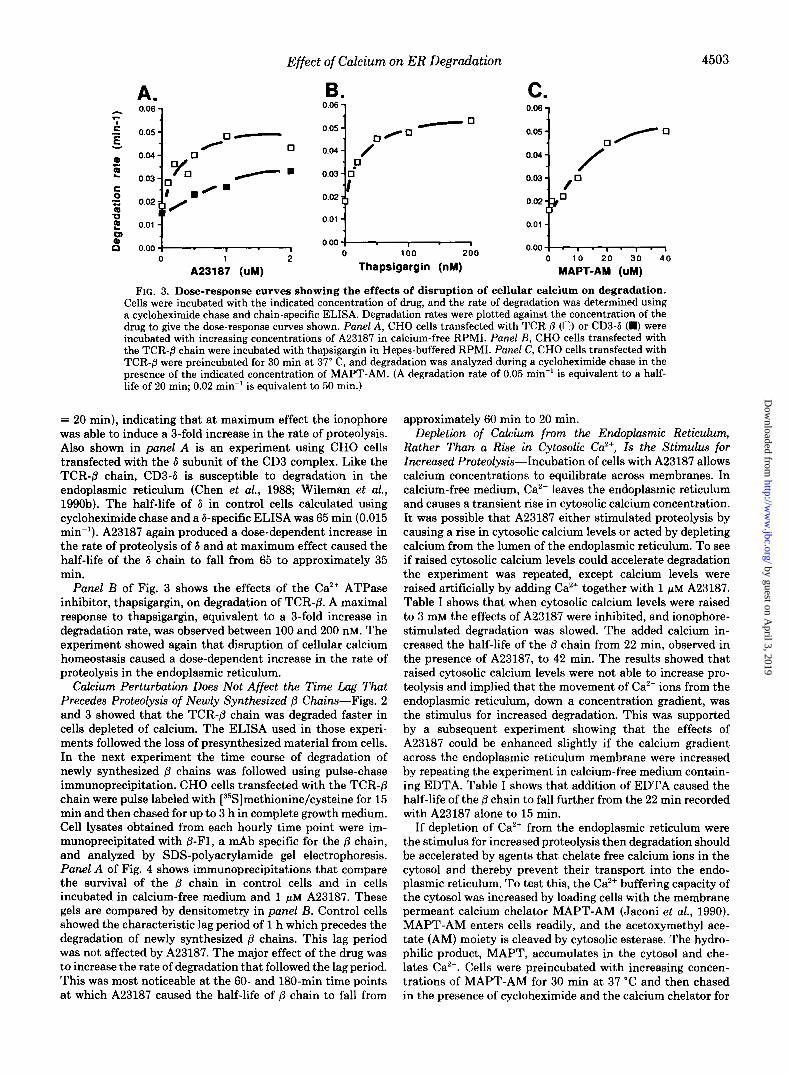
Fig. 2 shows the effects of A23187 on the degradation of presynthesized @ chains. CHO cells transfected with the TCR- p chain were suspended in calcium-free medium, and the turnover of the protein, in the presence or absence of 1 pM A23187, was analyzed using the cycloheximide chase. The graph shows that the ionophore caused an immediate stimu- lation of proteolysis. 1 p~ A23187 caused the half-life of the @ chain to fall from 55 min to 20 min. Panel A of Fig. 3 shows a dose-response curve for the effects of A23187. Cells were incubated in calcium-free medium in the presence of increas- ing quantities of A23187, and degradation rates calculated from the linear portions of the ELISA data curves were plotted against the concentration of A23187. A23187 caused a dose- dependent increase in the rate of degradation of the @ chain, and the response to the ionophore reached a maximum at 1 pM. Over this concentration range the degradation rate in- creased from 0.018 min" (tIl2 = 55 min) to 0.048 min" (t1/2
ELSA 1 1
100 Control
1 1
1 1 0 20 40 60 8 0 100
Time (min) FIG. 2. Effect of A23187 on degradation of TCR-/3 chain.
CHO cells transfected stably with the TCR-P chain were warmed to 37" C in media containing cycloheximide. A t the recorded time points cell aliquots (5 X IO5) were lysed in 1% Nonidet P-40 and assayed for @ chain using an ELISA. M, control cells; 0, cells incubated in calcium-free RPMI containing A23187.
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Effect of Calcium on ER Degradation 4503
A. 0.06 7
0.00 O.O1 1 0 1 2
A23187 (uM)
B. 0.06 1
I:::{ . , , 0.00
0 100 200
Thapsigargin (nM)
C. 0.06 1
0.02 $p
0.00 O.O1 1 0 10 20 30 4 0
MAPT-AM (uU)
FIG. 3. Dose-response curves showing the effects of disruption of cellular calcium on degradation. Cells were incubated with the indicated concentration of drug, and the rate of degradation was determined using a cycloheximide chase and chain-specific ELISA. Degradation rates were plotted against the concentration of the drug to give the dose-response curves shown. Panel A , CHO cells transfected with TCR 0 (0) or CD3-6 (m) were incubated with increasing concentrations of A23187 in calcium-free RPMI. Panel B, CHO cells transfected with the TCR-fi chain were incubated with thapsigargin in Hepes-buffered RPMI. Panel C, CHO cells transfected with TCR-0 were preincubated for 30 min at 37" C, and degradation was analyzed during a cycloheximide chase in the uresence of the indicated concentration of MAPT-AM. (A degradation rate of 0.05 min" is equivalent to a half- life of 20 min; 0.02 min" is equivalent to 50 rnin.)
= 20 min), indicating that at maximum effect the ionophore was able to induce a %fold increase in the rate of proteolysis. Also shown in panel A is an experiment using CHO cells transfected with the 6 subunit of the CD3 complex. Like the TCR-8 chain, CD3-6 is susceptible to degradation in the endoplasmic reticulum (Chen et al., 1988; Wileman et al., 1990b). The half-life of 6 in control cells calculated using cycloheximide chase and a 6-specific ELISA was 65 min (0.015 min"). A23187 again produced a dose-dependent increase in the rate of proteolysis of 6 and at maximum effect caused the half-life of the 6 chain to fall from 65 to approximately 35 min.
Panel B of Fig. 3 shows the effects of the Ca2+ ATPase inhibitor, thapsigargin, on degradation of TCR-p. A maximal response to thapsigargin, equivalent to a %fold increase in degradation rate, was observed between 100 and 200 nM. The experiment showed again that disruption of cellular calcium homeostasis caused a dose-dependent increase in the rate of proteolysis in the endoplasmic reticulum.
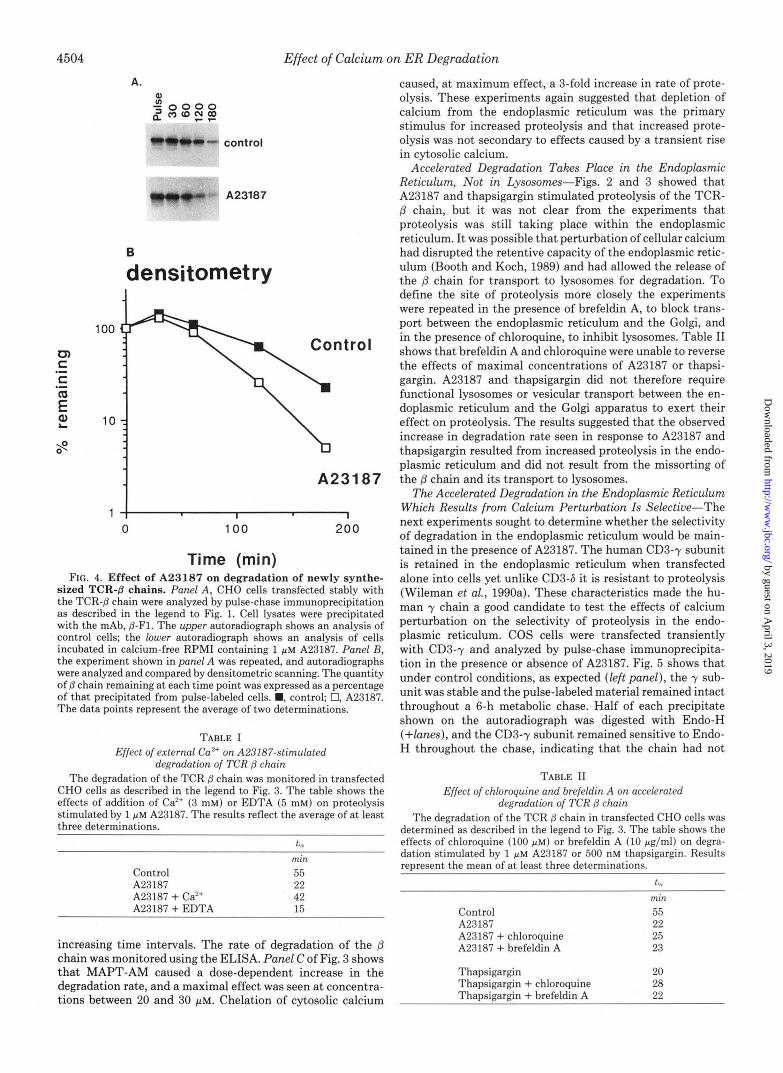
Calcium Perturbation Does Not Affect the Time Lag That Precedes Proteolysis of Newly Synthesized /3 Chains-Figs. 2 and 3 showed that the TCR-8 chain was degraded faster in cells depleted of calcium. The ELISA used in those experi- ments followed the loss of presynthesized material from cells. In the next experiment the time course of degradation of newly synthesized 8 chains was followed using pulse-chase immunoprecipitation. CHO cells transfected with the TCR-p chain were pulse labeled with [35S]methionine/cysteine for 15 min and then chased for up to 3 h in complete growth medium. Cell lysates obtained from each hourly time point were im- munoprecipitated with P-Fl, a mAb specific for the p chain, and analyzed by SDS-polyacrylamide gel electrophoresis. Panel A of Fig. 4 shows immunoprecipitations that compare the survival of the (3 chain in control cells and in cells incubated in calcium-free medium and 1 ~ L M A23187. These gels are compared by densitometry in panel B. Control cells showed the characteristic lag period of 1 h which precedes the degradation of newly synthesized (3 chains. This lag period was not affected by A23187. The major effect of the drug was to increase the rate of degradation that followed the lag period. This was most noticeable at the 60- and 180-min time points at which A23187 caused the half-life of p chain to fall from
approximately 60 min to 20 min. Depletion of Calcium from the Endoplasmic Reticulum,
Rather Than a Rise in Cytosolic Ca", Is the Stimulus for Increased Proteolysis-Incubation of cells with A23187 allows calcium concentrations to equilibrate across membranes. In calcium-free medium, Ca2+ leaves the endoplasmic reticulum and causes a transient rise in cytosolic calcium concentration. It was possible that A23187 either stimulated proteolysis by causing a rise in cytosolic calcium levels or acted by depleting calcium from the lumen of the endoplasmic reticulum. To see if raised cytosolic calcium levels could accelerate degradation the experiment was repeated, except calcium levels were raised artificially by adding Ca2+ together with 1 PM A23187. Table I shows that when cytosolic calcium levels were raised to 3 mM the effects of A23187 were inhibited, and ionophore- stimulated degradation was slowed. The added calcium in- creased the half-life of the p chain from 22 min, observed in the presence of A23187, to 42 min. The results showed that raised cytosolic calcium levels were not able to increase pro- teolysis and implied that the movement of Ca2+ ions from the endoplasmic reticulum, down a concentration gradient, was the stimulus for increased degradation. This was supported by a subsequent experiment showing that the effects of A23187 could be enhanced slightly if the calcium gradient across the endoplasmic reticulum membrane were increased by repeating the experiment in calcium-free medium contain- ing EDTA. Table I shows that addition of EDTA caused the half-life of the p chain to fall further from the 22 min recorded with A23187 alone to 15 min.
If depletion of Ca2+ from the endoplasmic reticulum were the stimulus for increased proteolysis then degradation should be accelerated by agents that chelate free calcium ions in the cytosol and thereby prevent their transport into the endo- plasmic reticulum. To test this, the Ca2+ buffering capacity of the cytosol was increased by loading cells with the membrane permeant calcium chelator MAPT-AM (Jaconi et al., 1990). MAPT-AM enters cells readily, and the acetoxymethyl ace- tate (AM) moiety is cleaved by cytosolic esterase. The hydro- philic product, MAPT, accumulates in the cytosol and che- lates Ca2+. Cells were preincubated with increasing concen- trations of MAPT-AM for 30 min at 37 "C and then chased in the presence of cycloheximide and the calcium chelator for
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

4506 Effect of Calcium on ER Degradation
of the two proteins was unchanged. Under each condition tested the removal of the membrane anchor from the P chain still afforded an approximate 5-fold resistance to proteolysis.
DISCUSSION
The first experiments showed that the movement of pro- teins from the endoplasmic reticulum into the secretory path- way was disrupted when cells were depleted of calcium. A23187 blocked the movement of the transfected IL2 receptor from the endoplasmic reticulum to the medial Golgi and slowed the secretion of the anchor minus IL2 receptor. These results agreed with the report of Lodish and Kong (1990) showing that A23187 slowed the secretion of proteins from HepG2 cells. Surprisingly, the Ca2+ ATPase inhibitor, thap- sigargin, increased the rate of transport of the IL2 receptor to the trans Golgi and doubled the rate of secretion of its ectodomain from cells. The cellular basis for the enhancement of secretion by thapsigargin is not known, but these observa- tions are consistent with the phenotype of yeasts carrying a mutant Ca2+ ATPase (PMR1-). In these cells the control of transport between the endoplasmic reticulum and the Golgi apparatus is disrupted, and they secrete heterologous fusion proteins that are retained in the endoplasmic reticulum of wild-type strains (Rudolph et al., 1989).
In spite of their different effects on secretion, A23187 and thapsigargin both stimulated degradation in the endoplasmic reticulum. Their predominant effect was to increase the rate of proteolysis. The lag period of 1 h, which is observed prior to degradation of proteins in the endoplasmic reticulum, was not affected. The cycloheximide chase assay, which makes use of an ELISA to follow the loss of presynthesized material, showed that the effects of thapsigargin and A23187 were immediate and dose dependent (Figs. 2 and 3). A23187 and thapsigargin both cause a transient rise in cytosolic calcium levels (Thastrup et al., 1990). Increased cytosolic calcium levels were not the stimulus for accelerated proteolysis. First, the actions of A23187 were reversed rather than enhanced when the ionophore was added to cells in the presence of 3 mM Ca2+. Second, reducing the levels of free Ca2+ in the cytosol by incubating cells with the membrane-permeant cal- cium chelator MAPT-AM caused a dose-dependent increase, not decrease, in the rate of degradation in the endoplasmic reticulum. Both results pointed to depletion of calcium from the endoplasmic retculum as being the stimulus for increased proteolysis.
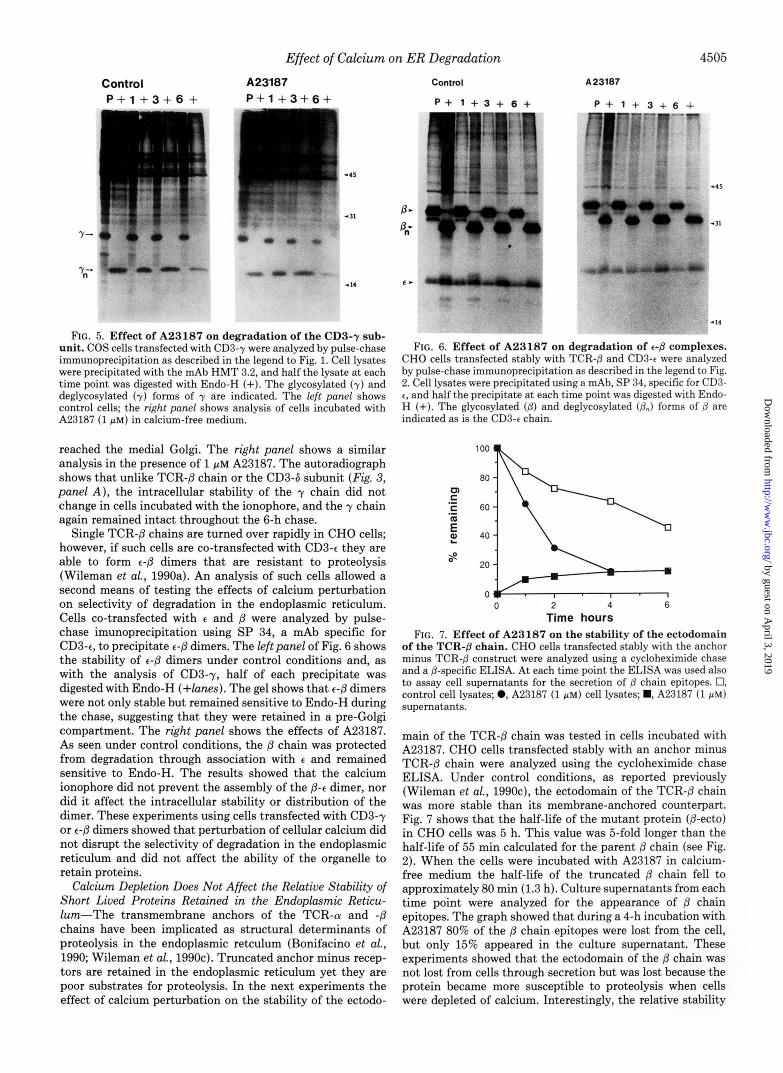
Degradation in the endoplasmic reticulum is selective (Bon- ifacino et d., 1989; Shia and Lodish, 1989; Wileman et d., 1990a), and results from three experiments suggested that this selectivity was maintained in cells incubated with A23187. First, the stimulation of proteolysis observed for the @ chain could be repeated for a second labile chain, CD3-6 (Fig. 3, panel A ) . Second, the ionophore had no effect on the stability or intracellular distribution of the CD3-y subunit, a protein that is resistant to degradation (Wileman et al., 1990a) yet is 70% homologous to CD3-6 (Krissansen et al., 1986). Third, A23187 did not disrupt the protective functions of subunit assembly. Dimers and higher order complexes of T- cell receptors that contain the stable CD3-t or -y subunits are long lived (Bonifacino et al., 1989; Wileman et al., 1990a). Concentrations of A23187 which caused a 3-fold increase in the rate of proteolysis of free /3 chains (Fig. 3, panel A ) had no effect on the assembly or stability of t-P dimers (Fig. 6). Taken together, the results suggested that depletion of cal- cium from the endoplasmic reticulum had a selective stimu- latory effect on proteolysis and did not precipitate indiscrim- inate degradation within the organelle.
One determinant of proteolysis within the endoplasmic reticulum resides in the membrane anchors of the TCR-(U and P-chains (Bonifacino et al., 1990; Wileman et al., 1990~). When these membrane anchors are removed the proteins are still retained in the endoplasmic reticulum, but they become more resistant to degradation. Interestingly, A23187 was able to accelerate the proteolysis of the anchor minus @ chain (Fig. 7) yet it had no effect the stability of the CD3-t or -y subunits (Figs. 5 and 6). These results suggested that the cellular basis for the resistance of anchor minus /3 chain to proteolysis in the endoplasmic reticulum may be different from that of the t and y chains. The most simple explanation for the results would be that t and y chains are not recognized by the proteases. The anchor minus @ chain, on the other hand, is recognized by proteases but is a poor substrate for proteolysis. The protein is, nonetheless degraded faster when the overall rate of proteolysis in the endoplasmic reticulum is increased by calcium depletion.
Prolonged exposure of cells to A23187 or thapsigargin for 4-8 h causes the endoplasmic reticulum to fragment and become dispersed throughout the cytoplasm (Booth and Koch, 1989). During this period resident proteins of the endoplasmic reticulum can be secreted. It seems unlikely from the results reported in this study that fragmentation of the endoplasmic reticulum is the cause of increased proteolysis. The stimula- tion of proteolysis seen in response to calcium depletion was immediate and occurred much sooner than the morphological changes that take place in cells after many hours of exposure to calcium ionophores. The structure of the endoplasmic reticulum did not change during the relatively brief time course (20-60 min) of the experiments on degradation. The reticular structure of the organelle, as judged by the distri- bution of sequestered DiOCs (Lee and Chen, 1988), was un- changed 30 min after the addition of A23187 or thapsigargin (data not shown). The capacity of the endoplasmic reticulum to retain proteins was likewise unaffected during the time course of these experiments. CD3-y and P-t dimers remained sensitive to Endo-H throughout a metabolic chase in the presence of 1 PM A23187 (Figs. 5 and 6).
Although it has not been demonstrated directly, Ca2+ ions may aid the folding of newly synthesized proteins (Lodish and Kong, 1990). This seems a likely site for the action of A23187 and thapsigargin on degradation. It is possible that depletion of Ca2+ disrupts protein folding in the endoplasmic reticulum, and this could cause the P and 6 chains to become trapped in a conformation that is more sensitive to proteolysis. It is also possible that calcium depletion disrupts the structure of the lumen of the endoplasmic reticulum. Recent reports have suggested that the interior of the endoplasmic reticulum con- tains a protein matrix that is cross-linked by Ca2+ (Koch, 1987; Booth and Koch, 1989; Sambrook, 1990). The high concentration of protein in the matrix (100 mg/ml) may restrict the movement of proteins within the organelle. If the matrix were to regulate the interaction between substrates and resident proteases then dissolution of the matrix in response to calcium depletion could have again increased the rate of degradation. There is some evidence that partially folded or incompletely assembled proteins can be retained in the endoplasmic reticulum through associations with the lu- menal content of the organelle. Best characterized is the retention of unassembled immunoglobulin heavy chains through associations with immunoglobulin-binding protein. These associations are transient and serve to catalyze the folding of the heavy chain and its subsequent assembly with light chains (Haas and Wabl, 1983; Bole et aL, 1986; Hender- shot, 1990). In an analogous situation it is possible that single
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Effect of Calcium on ER Degradation 4507
/? or 6 chains are retained and stabilized by the lumenal content of the organelle. This protection may be lost after calcium depletion, and B and 6 are degraded faster. At present the precise role played by immunoglobulin-binding protein in the retention and degradation of T-cell receptor chains has not been resolved. Immunoprecipitations of the different com- ponents of the receptor (Alarcon et al., 1988; Chen et al., 1988) and other proteins trapped in the endoplasmic reticulum (Lodish and Kong, 1990) have yet to reveal strong associations with immunoglobulin-binding protein. These results do not, however, preclude the transient stabilization of T-cell recep- tor proteins by immunoglobulin-binding protein or other lu- menal proteins in uiuo.
In summary, this report has shown that degradation within the endoplasmic reticulum can be accelerated by drugs that disrupt intracellular calcium stores. The experiments have suggested that depletion of calcium from the endoplasmic reticulum, rather than a rise in cytosolic calcium levels, is the stimulus for increased proteolysis. It remains to be determined if the results reported here reflect a physiological role for calcium in the regulation of proteolysis in the endoplasmic reticulum. Calcium is released from the orgenelle by inositol 1,4,5-trisphosphate when cells are activated, and our results raise the possibility that proteolysis in the endoplasmic retic- ulum can be stimulated in activated T-cells. Cytosolic calcium levels oscillate in resting cells because of local, but transient, actions of inositol 1,4,5-trisphosphate on the endoplasmic reticulum (reviewed by Berridge, 1990). Local fluxes of cal- cium may be able to generate regional increases in proteolysis in the endoplasmic reticulum. Finally, the control of Ca" gradients within pre-Golgi compartments could conceivably be used to sort proteins (see Sambrook, 1990; Kelly, 1990) in much the same way that proton gradients are used within the endocytic pathway. If vesicles leaving the endoplasmic retic- ulum had to be drained of Ca2+ before they could deliver their content to a compartment containing proteases, then this would offer another site of action for A23187 or thapsigargin.
REFERENCES Alarcon, B., Berkhout, B., Breitmeyer, J., and Terhorst, C. (1988) J.
Amara, J . F., Lederkremer, G., and Lodish, H. (1989) J. Cell Biol.
Berridge, M. (1990) J. Biol. Chem. 265,9583-9586 Bole, D. G., Hendershot, L. M., and Kearney, J. F. (1986) J. Cell Biol.
Bonifacino, J. S., Suzuki, C. K., Lippincott-Schwartz, J., Weissman,
Bonifacino, J. S., Suzuki, C. K., and Klausner, R. D. (1990) Science
Biol. Chem. 263,2953-2961
109,3315-3324
102.1558-1566
A. M., and Klausner, R. D. (1989) J. Cell Biol. 109 , 73-83
247 , 79-82
Booth, C., and Koch, G. L. E. (1989) Cell 59,729-737 Chen, C., Bonifacino, J. S., Yuan, L. C., and Klausner, R. D. (1988)
Copeland, C. S., Doms, R. W., Bolazau, E. M., Webster, R. G., and
Copeland, C. S., Zimmer, K.-P., Wagner, K. R., Healy, G. A., Mell-
Fujiwara, T., Oda, K., Yokota, S., Takatsuki, A., and Ikehara, Y.
Gething, M.-J., McCammon, K., and Sambrook, J. (1986) Cell 46 ,
Hass I., and Wabl, M. (1983) Nature 306 , 387-389 Hendershot, L. M. (1990) J. Cell Bwl. 1 1 1,829-837 Hurtley, M., and Helenius, A. (1989) Annu. Rev. Cell Bwl. 5 , 277-
Jaconi, M. E. E., Lew, D. P., Carpentier, J.-L., Magnusson, K. E.,
Kelly, R. B. (1990) Nature 345,480-481 Koch, G. L. E. (1987) J. Cell Sci. 8 7 , 491-492 Krissansen, G. W., Owen, M. J., Verbi, W., and Crumpton, M. J.
Laemmli, U. K. (1970) Nature 227 , 680-685 Lee, C., and Chen, L. B. (1988) Cell 5 4 , 37-46 Lippincott-Schwartz, J., Bonifacino, J. S., Yuan, L. C., and Klausner,
Lippincott-Schwartz, J., Yuan, L. C., Bonifacino, J. S., and Klausner,
Lodish, H. F., and Kong, N. (1990) J. Biol. Chem. 265, 10893-10899 Pessano S., Oettegen, H. C., Bhan, A. K., and Terhorst, C. (1985)
Rose, J. K., and Doms, R. W. (1988) Annu. Rev. Cell Biol. 4,257-288 Rothman, J. E. (1989) Cell 59,591-601 Rotundo, R. L., Thomas, K., Porter-Jordan, K., Bensen, R. J. J.,
Fernandez-Valle, C., and Fine, R. E. (1989) J. Bid. Chern. 264,
Rubin, L. A., Kurman, C. C., Biddison, W. E., Goldman, N. D., and Nelson, D. L. (1985) Hybridoma 4,91-102
Rudolph, H. K., Antebi, A., Fink, G. R., Buckley, C. M., Dorman, T . E., LeVitre, J., Davidow, L. S., Mao, J., and Moir, D. T. (1989) Cell
J. Cell Biol. 107 , 2149-2161
Helenius, A. (1986) J. Cell Biol. 103, 1179-1191
man, I., and Helenius, A. (1988) Cell 5 3 , 197-209
(1988) J. Biol. Chem. 2 6 3 , 18545-18552
939-950
307
and Stendhal, 0. (1990) J. Cell Biol. 110,1555-1564
(1986) EMBO J. 5,1799-1808
R. D. (1988) Cell 54,209-220
R. D. (1989) Cell 56,801-813
EMBO J. 4,337-344
3146-3152
58,133-145 Sambrook, J. F. (1990) Cell 6 1 , 197-199 Sato, R., Imanaka, T., Takatsuki, A., and Takano, T. (1990) J. Bid.
Shia, M., and Lodish, H. (1989) Proc. Natl. Acud. Sci. U. S. A. 8 4 ,
Sitia, R., Neuberger, M., Alberini, C., Bet, P., Fra, A., Valetti, C.,
Stoller, T. J., and Shields, D. (1989) J. Cell Biol. 108, 1647-1655 Thastrup, O., Cullen, P. J., Drobak, B. K., Hanley, M. R., and
Dawson, A. (1990) Proc. Natl. Acud Sci. U. S. A. 8 7 , 2466-2470 Treiger, B. T., Leonard, W. J., Svetlik, P., Rubin, L. A., Nelson, D.
L., and Greene, W. G. (1986) J. Immunol. 136,4099-4105 Wileman, T., Carson, G. R., Concino, M., Ahmed, A., and Terhorst,
C. (1990a) J. Cell Biol. 110,973-986 Wileman, T., Pettey, C, and Terhorst, C. (1990b) Int. Immunol. 2,
743-754 Wileman, T., Carson, G. R., Shih, F. F., Concino, M. F., and Terhorst,
C. (1990~) Cell Regul. 1,907-919
Chem. 265,11880-11884
1158-1162
Williams, G., and Milstein, C. (1990) Cell 60, 781-790
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

T Wileman, L P Kane, G R Carson and C Terhorstreticulum.
Depletion of cellular calcium accelerates protein degradation in the endoplasmic
1991, 266:4500-4507.J. Biol. Chem.
http://www.jbc.org/content/266/7/4500Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/266/7/4500.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from





















![Endoplasmic reticulum[1]](https://img.pdfslide.us/doc/110x75/58ed5fc71a28aba1678b4611/endoplasmic-reticulum1.jpg)







